
Community structure of fishery resources from the Nanshawaters of Pearl River Estuary in autumn
-
摘要:
根据2015年11月珠江口南沙海域的渔业资源调查资料,对该海域秋季渔业资源群落组成、优势种和生物多样性等群落结构特征进行了初步研究。结果表明,该海域共出现渔业生物40种,以鱼类和甲壳类为主。其中,底层鱼类21种,中上层鱼类3种,甲壳类16种。相对重要性指数(IRI)可用来度量物种的生态优势度,秋季优势种类有亨氏仿对虾(Parapenaeopsis hungerfordi)和锐齿
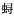 (Charybdis acuta)2种(IRI≥1 000)。该区域内各站位质量渔获率为0.63~11.15 kg·h-1,平均为4.23 kg·h-1;尾数渔获率为84~885尾·h-1,平均为433尾·h-1。Shannon-Wiener多样性指数(H′)为2.07,Margalef丰富度指数(D)为1.64,Peilou均匀度指数(J′)为0.59。分析表明,该海域渔获物以小型经济甲壳动物和经济幼鱼为主,渔获个体偏小,总体上多样性空间差异不大。与该区域2002年同月份的调查数据相比,目前渔获物多样性下降明显,群落结构相对简单。Abstract:
(Charybdis acuta)2种(IRI≥1 000)。该区域内各站位质量渔获率为0.63~11.15 kg·h-1,平均为4.23 kg·h-1;尾数渔获率为84~885尾·h-1,平均为433尾·h-1。Shannon-Wiener多样性指数(H′)为2.07,Margalef丰富度指数(D)为1.64,Peilou均匀度指数(J′)为0.59。分析表明,该海域渔获物以小型经济甲壳动物和经济幼鱼为主,渔获个体偏小,总体上多样性空间差异不大。与该区域2002年同月份的调查数据相比,目前渔获物多样性下降明显,群落结构相对简单。Abstract:According to the fisheries resources data collected from the Nansha waters of Pearl River Estuary in November 2015, we investigated the community structure, dominant species and diversity of fishery resources in this area, and 40 fishery species, mainly fishes and crustaceans, were found, including 21 demersal fishes, 3 pelagic fishes and 16 crustaceans. The index of relative importance (IRI) was used to measure ecological dominance of species to decide the role each species played in the community. The dominant species were Charybdis acuta and Parapenaeopsis hungerfordii in autumn, whose IRIs were higher than 1 000. The range of catch rates in weight was 0.63~11.15 kg·h-1 (average:4.23 kg·h-1). The range of catch rates in individual number was 84~885 ind·h-1 (average:433 ind·h-1). The average Shanon-Wiener diversity index (H′), Pielou evenness index (J′) value and Margalef richness index (D) of fishery resources in autumn were 2.07, 1.64 and 0.59, respectively. The analysis shows that sizes of the catch were generally small, and the catch mainly consisted of small commercial crustaceans and juveniles of commercial fishes. The spatial distribution of diversity of catches had little variation of in all stations. Compared with the fisheries catches in autumn 2002, the species diversity declines and the community structure is relatively simple at present.
-
珠江口南沙海域处于珠江水系的蕉门、虎门、洪奇沥门、横门等水道的入海口,属亚热带海洋季风气候,雨量充沛、气候温和,为众多生物的生长和繁殖提供了良好的环境[1]。由于河口海域内咸淡水交汇,水质肥沃且饵料生物丰富,使得该区域成了多种海洋渔业生物季节性洄游种群和地方性种群重要的产卵场和栖息地,渔业资源丰富,具有重要的渔业经济地位和生态保护价值[2-4]。
渔业资源群落结构及其多样性对维持渔业生态系统的稳定有重要作用[5-7],通过对渔业资源群落结构的研究,可以更好地理解各种群与其所在环境之间的相关性,并掌握近海及河口生态系统对环境变化的响应过程[8-10]。近年来,由于珠江三角洲地区经济的飞速发展[11],珠江口邻近海域的生态环境日渐受到人类活动的影响和破坏,使得该海域的渔业正承受着越来越严重的人口、资源和环境压力,而该海域的鱼类群落及生物组成也随之发生了较为明显的变化[12-14]。因此,有必要了解该海域的鱼类资源结构及生物多样性现状,为珠江口海域的渔业资源保护及可持续性开发利用提供参考,也为进一步开展河口渔业资源对人类胁迫的适应性研究提供基础资料。
1. 材料与方法
1.1 资料来源
数据来自于2015年11月珠江口南沙海域(22°28′N~22°40′N, 113°30′E~113°40′E)底拖网调查资料。该调查结合海湾的地理形状、海湾利用的现状,选取11个拖网作业站位、20个水质调查站位、20个浮游动物调查站位,开展生态环境和渔业资源现状调查(图 1)。
S1、S2、S4、S6、S8、S9、S11、S13、S16、S18、S20站为渔业资源调查站位。调查船功率为280 kW,网具尺寸为30 m×200 m (高×宽)。由于拖网调查站位于航道旁,每站拖曳10~20 min,拖速为2.6~4.3 kn。对拖网所获的渔业生物样品现场进行分类(尽可能至种)及生物学测量,并记录相关信息。
1.2 数据分析
通过Pianka的相对重要性指数(index of relative importance,IRI)来度量珠江口南沙海域的鱼类优势度[15]:
$ {\rm{IRI}} = \left( {N\% + W\% } \right) \times F $
式中N%代表个体数量百分比(每种鱼类个体数量占所捕渔获总数量的比例);W%代表个体质量百分比(每种鱼类质量占所捕渔获总质量的比例);F是出现频率(某一物种出现的站位数占总采样站位数中比例)。定义IRI≥1 000的物种为优势种,100≤IRI<1 000的物种为常见种,优势种和常见种合称为重要鱼种成分[16-17]。
使用丰富度指数(D)[18]、多样性指数(H′)[19]和均匀度指数(J′)[20]来表示珠江口南沙海域的生物多样性,其计算公式分别为:
$ D = {\rm{ }}\frac{{S-1}}{{{\rm{ln}}N}} $
$ H' = - \sum\limits_{i = 1}^S {{P_i}{\rm{lo}}{{\rm{g}}_2}{P_i}} $
$ J' = \frac{{H'}}{{{{\log }_2}S}}$
式中N为渔获物样品总质量,S为渔获物样品种类数量,Pi为第i种渔获物质量在总渔获物质量中所占比例。
使用YSI便携式温盐深仪现场对海水盐度、温度和水深等环境参数进行测定。通过SPSS 17.0软件对鱼类群落结构及多样性参数与环境因子的相关性进行分析。
2. 结果
2.1 种类组成
该次调查共鉴定渔获种类40种,隶属于10目27科36属。鱼类8目18科23属24种,占总渔获种类数的60%,其中以鲈形目居多,有7科12属12种,占鱼类总种类数的50%,占总渔获种类数的30%。甲壳类2目9科13属16种,占总渔获种类数的40%,主要为十足目及口足目,有8科11属14种,占总渔获种类数的35%。
40种渔业生物中,底层鱼类21种,主要有暗纹东方鲀(Takifugu obscurus)、触角沟

从鱼类生长和繁殖的适温性来看,暖水种主要有短吻鲾、卵鳎(Solea ovata Richardson)、孔


2.2 渔获率平面分布
调查区域内各站位渔获率(质量)为0.63~11.15 kg·h-1,高低相差约18倍,平均渔获率(质量)为4.23 kg·h-1,主要为鱼类和甲壳类。S9站渔获率最多(11.15 kg·h-1),其中甲壳类居多,占该站位总渔获率的62.5%。S1、S2、S4、S6、S8及S13站的渔获率均较少,低于其平均水平。S13站渔获量最低(0.63 kg·h-1),其中鱼类居多,占该站位总渔获率的85.3%(图 2)。
调查区域内各站位渔获率为84~885尾·h-1,平均渔获率为433尾·h-1,以甲壳类居多。S6站平均渔获率最大(885尾·h-1),其中甲壳类占85.4%。其次为S8站,渔获率为756尾·h-1,其中甲壳类占87.6%。在11个渔业资源调查站位中,S11和S13站平均尾数密度相对较小,分别为114尾·h-1和84尾·h-1,甲壳类分别占5.7%和71.4%(图 3)。
2.3 主要优势种类
根据IRI对渔获物的优势种类进行分析(表 1)。珠江口南沙海域内的优势种是亨氏仿对虾(Parapenaeopsis hungerfordi)和锐齿

种类
species占总渔获量百分比/%
percentage of biomass占总渔获数量百分比/%
percentage of catch相对重要性指数
IRI短吻鲾Leiognathus brevirostris 4.62 7.26 538 红狼牙 1.73 1.53 147 棘头梅童鱼Collichthys lucidus 2.13 5.93 438 孔 5.21 7.77 940 皮氏叫姑鱼Johnius belangerii 9.41 6.03 975 鲈Lateolabrax japonicus 10.68 0.10 97 鲻Mugil cephalus 15.44 2.97 497 亨氏仿对虾Parapenaeopsis hungerfordi 5.02 8.79 1 000 脊尾白虾Palaemon carincauda Holthuis 1.58 8.18 531 巨指长臂虾Palaemon macrodactylus 0.39 5.52 322 锯缘青蟹Scylla serrata 11.03 0.51 311 墨吉对虾Penaeus merguiensis 1.11 3.17 233 锐齿 15.30 13.09 1 795 2.4 多样性特征
通过D、H′和J′来研究珠江口南沙海域渔业资源群落结构的多样性特征(表 2)。计算得出该调查海域内各站位鱼类的平均D是0.96,变化范围是0~1.93。平均H′是1.73,变化范围是0~3.14。平均J′是0.61,变化范围是0~0.88。其中D、H′和J′最高的站位均是S6,最低的站位均是S1。
表 2 珠江口南沙海域各站位丰富度指数(D)、多样性指数(H′)和均匀度指数(J′)Table 2 Diversity indices of fish in the Nansha waters of Pearl River Estuary站位
station丰富度指数(D)
Margalef index多样性指数(H′)
Shannon-Wiener index均匀度指数(J′)
Pielou evenness index鱼类
fish甲壳类
crustacean鱼类
fish甲壳类
crustacean鱼类
fish甲壳类
crustaceanS1 0 1.09 0 1.91 0 0.68 S2 1.17 0.77 2.39 1.72 0.85 0.74 S4 1.06 1.15 1.54 2.16 0.51 0.77 S6 1.93 0.82 3.14 1.53 0.88 0.59 S8 1.29 0.76 2.54 1.52 0.85 0.59 S9 0.66 0.64 1.84 1.08 0.71 0.47 S11 0.55 0 0.55 0 0.24 0 S13 0.44 0.73 0.96 1.30 0.61 0.82 S16 0.91 0.58 1.64 1.54 0.55 0.77 S18 1.52 1.00 2.33 1.69 0.74 0.56 S20 1.01 0.70 2.16 1.40 0.72 0.60 该调查海域内各站位甲壳类的平均D是0.75,变化范围是0~1.15。平均H′是1.44,变化范围是0~2.16。平均J′是0.60,变化范围是0~0.82。其中D和H′最高的站位均是S4,最低的均是S11;J′最高的站位是S13,最低的是S11。
单因子相似性分析(ANOSIM)表明,珠江口南沙海域秋季各站点渔业资源群落结构多样性无显著性差异(P>0.05)。
3. 讨论
3.1 种类组成
海洋鱼类种类组成及其分布主要受温度、盐度等环境因子的影响[21-23]。珠江口附近海区既受沿岸低盐水的影响,亦与南海外海水的消长密切相关,鱼类的种类组成具有自身的特点。调查表明,珠江口南沙海域各站位渔获种类较为相似,主要以鱼类和甲壳类为主,其中底层鱼类较多;从适温性来看,鱼类中以暖水种和暖温种为主;从生态类型上来看,多为沿岸及近海类型鱼类。此外,种类组成具有明显的空间分布特征,靠近内河口种类少而离岸靠海的种类多。离陆地较近的S1、S11和S13等内河口站位的渔获物种类分布均较少,各站位渔获种类仅有5~8种,且主要以甲壳类、鲻和广东鲂等淡水或咸淡水种类为主。而龙穴岛靠外的离岸海域渔获种类相对较多,如站位S4、S6和S16,每个站位渔获种类为12~18种,这可能由于龙穴岛地处珠江口蕉门水道和虎门水道出口交界处,该海域四季内咸淡水时期交替出现,且水深较深,适宜不同生态习性生物的栖息生长和繁殖[24]。因此,这些站点不仅有沿岸种类,如脊尾白虾、红狼牙虎鱼等,同时也出现了皮氏叫姑鱼等海洋性种类(图 4)。
3.2 群落多样性
鱼类的生存环境及其自身的生理活动均会引起鱼类群落结构和多样性发生变化[25-26]。对于珠江口水域的鱼类群落而言,其多样性指数变化范围比较大,且表现出一定的时空差别[2, 27]。而该次调查的珠江口南沙海域各站位鱼类群落多样性总体差异不大,从生物多样性指数来看,H′为0.56~3.05,黄阁镇周边海域除S2站位外,S1、S11和S13站的H′均较低(<1.30),而龙穴岛周边海域站位(S4和S6)的H′相对较高(>1.60),远离陆地海域站位的多样性明显较其他两区偏高,其中S20站的H′达2.49;D′为0.69~2.24,各站点的丰富度差异不大,S1、S11和S13站的丰富度值较小,而S4、S6和S18站的鱼类丰富度较高;J′为0.22~0.85,其最高值和最低值分别出现在S2和S11站位,而各站位的J′分布没有明显特征。
群落种类多样性取决于种的数量及其分布的均匀度2个因素,在种类数一定的情况下,各个种群间的数量分布越均匀,其多样性越高[28]。2002年10月调查方法和范围与该次基本一致①,可用来对比分析人类活动对该海域渔业生物的影响(表 3)。与2002年同期渔获物72种相比,该次渔获种类数下降了80%。渔获物种类变化最为明显是头足类。2002年渔获物中头足类有2种,分别为杜氏枪乌贼(Loligo duvaucelii Orbigny)和田乡枪乌贼(L.tagoi Sasaki),而该次调查则没有渔获头足类。鱼类的种类及多样性指数下降明显,鱼类种类数从2002年的61种下降至2015年的54种,多样性指数也从2002年的2.64下降至2015年的1.73。另外,甲壳类种类数虽然明显增加,但多样性指数变化不大。推测当前甲壳类种类增加的主要原因是气候变化和捕捞等人类活动造成鱼类资源衰退后,使得该海域甲壳类被捕食压力释放,以及甲壳类食物可得性增加所导致。总体而言,与2002年相比,当前珠江口南沙海域渔获生物多样性明显下降。这种变化与该区域捕捞、海域污染及工程建设等人类活动对渔业生物产卵场和索饵场的影响密切相关。特别是近10多年来,随着珠江三角洲经济社会的高速发展,生活污水和养殖污染等因素的影响,珠江口海区生态环境受到了严重的污染,赤潮爆发频次明显增加[29]。此外,珠江口作为繁忙的海上交通枢纽,航道疏浚以及高强度的围填海活动[30]等破坏了生物栖息地和产卵场,从而降低了该区域的生物多样性,尤其是底栖鱼类。凤鲚(Clilia mystus)、皮氏叫姑鱼等珠江口海域的底栖鱼类,前者为典型咸淡水洄游种类,均为2002年调查渔获物的优势种之一,而此次调查则没有渔获。这也在一定程度上反映了人类活动对该海域底栖性渔业资源的影响。此外,渔获物规格发生了明显的变化。以棘头梅童鱼为例,它是2个不同年代的常见种,此次调查该种体长为20~77 mm,明显低于2002年22~128 mm。表明该海域渔业资源出现了低龄化和小型化的现象,这与珠江口伶仃洋底层游泳动物变化趋势一致[31]。
表 3 不同年代间珠江口南沙海域物种多样性比较Table 3 Comparison of species diversity indices of fishery resources in the Nansha waters of Pearl River Estuary in different years参数
parameter2002年 2015年 鱼类
fish甲壳类
crustacean头足类
cephalopod鱼类
fish甲壳类
crustacean头足类
cephalopod种类数number of species 61 9 2 24 16 - 多样性指数(H′) Shanncn-Wiener index 2.64 1.42 0.57 1.73 1.44 - ① 中国水产科学研究院南海水产研究所.广州南沙地区附近水域水生生态调查研究报告[R].广州:中国水产科学研究院南海水产研究所,2004:41-61.
3.3 群落特征与环境因子的关系
温度、盐度对海洋鱼类来说是最重要的环境因子[32]。调查区域内各站位水温为24.8~26.6 ℃,平均为25.6 ℃,表层和底层水温温差最大为0.4 ℃,其中距离陆地较远的S8、S9和S20站的水温相对较高;各站位溶解氧的质量浓度为3.40~6.65 mg·L-1,平均为4.55 mg·L-1,溶解氧最高值出现在河口内S13站,最低值出现在离岸S18站,说明在地形的约束和沿岸流的作用下,秋季该海域层水体交换频繁,层化现象不明显,使得溶解氧相对充足。各站位盐度为0.13~22.32,平均为7.94,盐度最高值和最低值分别出现在S20和S13站,盐度分布呈近岸口内站点向口外逐渐增加的态势;各站位pH为7.37~8.10,平均为7.77。从多样性指数与环境因子的相关性分析结果来看(表 4),珠江口南沙海域相同测度指标多样性指数间一般呈显著相关性,而不同测度指标多样性指数和环境因子间的相关性大多不甚显著。其鱼类种类数与表层温度、底层温度、表层盐度和底层盐度均呈显著正相关(P<0.05);D、H′和J′与环境因子不存在显著相关性。说明该海域鱼类群落多样性不存在明显的环境梯度特征,与珠江口其他水域鱼类群落结构和环境因子的关系相比,该海域的群落结构相对简单。
表 4 珠江口南沙海域各站位物种多样性指数与环境因子间的相关性Table 4 Correlation between species diversity indices and environmental factors in the Nansha waters of Pearl River EstuaryF1 F2 F3 F4 F5 F6 F7 F8 F1 1 F2 0.693* 1 F3 0.633* 0.918** 1 F4 0.793* 0.971** 0.965** 1 F5 0.745* 0.939** 0.996** 0.981** 1 F6 0.926** 0.588 0.478 0.666 0.597 1 F7 0.429 0.416 0.262 0.392 0.307 0.624* 1 F8 0.310 0.163 0.460 0.144 0.079 0.271 0.894** 1 注:*. P<0.05;* *. P<0.01。F1.鱼类种类数;F2.表层温度;F3.表层盐度;F4.底层温度;F5.底层盐度;F6.丰富度指数;F7.多样性指数;F8.均匀度指数
Note:F1.number of fish species;F2.sea surface temperature;F3.sea surface salinity;F4.sea bottom temperature;F5. sea bottom salinity;F6.Margalef richness index D;F7.Shanon-Wiener diversity index H′;F8.Pielou evenness index J′ -
![]()
图 3 各站位渔获率(尾数)分布
Figure 3. Catch rate(number of individuals)distribution in each station
表 1 珠江口南沙海域游泳动物的相对重要性指数
Table 1 Indices of relative importance of nekton in the Nansha waters of Pearl River Estuary
种类
species占总渔获量百分比/%
percentage of biomass占总渔获数量百分比/%
percentage of catch相对重要性指数
IRI短吻鲾Leiognathus brevirostris 4.62 7.26 538 红狼牙 1.73 1.53 147 棘头梅童鱼Collichthys lucidus 2.13 5.93 438 孔 5.21 7.77 940 皮氏叫姑鱼Johnius belangerii 9.41 6.03 975 鲈Lateolabrax japonicus 10.68 0.10 97 鲻Mugil cephalus 15.44 2.97 497 亨氏仿对虾Parapenaeopsis hungerfordi 5.02 8.79 1 000 脊尾白虾Palaemon carincauda Holthuis 1.58 8.18 531 巨指长臂虾Palaemon macrodactylus 0.39 5.52 322 锯缘青蟹Scylla serrata 11.03 0.51 311 墨吉对虾Penaeus merguiensis 1.11 3.17 233 锐齿 15.30 13.09 1 795  下载: 导出CSV
下载: 导出CSV
表 2 珠江口南沙海域各站位丰富度指数(D)、多样性指数(H′)和均匀度指数(J′)
Table 2 Diversity indices of fish in the Nansha waters of Pearl River Estuary
站位
station丰富度指数(D)
Margalef index多样性指数(H′)
Shannon-Wiener index均匀度指数(J′)
Pielou evenness index鱼类
fish甲壳类
crustacean鱼类
fish甲壳类
crustacean鱼类
fish甲壳类
crustaceanS1 0 1.09 0 1.91 0 0.68 S2 1.17 0.77 2.39 1.72 0.85 0.74 S4 1.06 1.15 1.54 2.16 0.51 0.77 S6 1.93 0.82 3.14 1.53 0.88 0.59 S8 1.29 0.76 2.54 1.52 0.85 0.59 S9 0.66 0.64 1.84 1.08 0.71 0.47 S11 0.55 0 0.55 0 0.24 0 S13 0.44 0.73 0.96 1.30 0.61 0.82 S16 0.91 0.58 1.64 1.54 0.55 0.77 S18 1.52 1.00 2.33 1.69 0.74 0.56 S20 1.01 0.70 2.16 1.40 0.72 0.60
下载: 导出CSV
表 3 不同年代间珠江口南沙海域物种多样性比较
Table 3 Comparison of species diversity indices of fishery resources in the Nansha waters of Pearl River Estuary in different years
参数
parameter2002年 2015年 鱼类
fish甲壳类
crustacean头足类
cephalopod鱼类
fish甲壳类
crustacean头足类
cephalopod种类数number of species 61 9 2 24 16 - 多样性指数(H′) Shanncn-Wiener index 2.64 1.42 0.57 1.73 1.44 -
下载: 导出CSV
表 4 珠江口南沙海域各站位物种多样性指数与环境因子间的相关性
Table 4 Correlation between species diversity indices and environmental factors in the Nansha waters of Pearl River Estuary
F1 F2 F3 F4 F5 F6 F7 F8 F1 1 F2 0.693* 1 F3 0.633* 0.918** 1 F4 0.793* 0.971** 0.965** 1 F5 0.745* 0.939** 0.996** 0.981** 1 F6 0.926** 0.588 0.478 0.666 0.597 1 F7 0.429 0.416 0.262 0.392 0.307 0.624* 1 F8 0.310 0.163 0.460 0.144 0.079 0.271 0.894** 1 注:*. P<0.05;* *. P<0.01。F1.鱼类种类数;F2.表层温度;F3.表层盐度;F4.底层温度;F5.底层盐度;F6.丰富度指数;F7.多样性指数;F8.均匀度指数
Note:F1.number of fish species;F2.sea surface temperature;F3.sea surface salinity;F4.sea bottom temperature;F5. sea bottom salinity;F6.Margalef richness index D;F7.Shanon-Wiener diversity index H′;F8.Pielou evenness index J′
下载: 导出CSV
-
[1] 李玫, 陈桂珠, 彭友贵.广州南沙湿地生物多样性现状及其保护[J].防护林技科, 2009(3):46-48. http://www.cnki.com.cn/Article/CJFDTOTAL-FHLK200903018.htm [2] 王迪. 珠江口鱼类群落及物种多样性研究[D]. 上海: 上海海洋大学, 2006: 45. http: //www. cnki. com. cn/Article/CJFDTOTAL-NFSC201601015. htm [3] 王迪, 林昭进.珠江口鱼类群落结构的时空变化[J].南方水产, 2006, 2(4):37-45. http://www.cnki.com.cn/Article/CJFDTOTAL-NFSC200604009.htm [4] 晏磊, 谭永光, 杨吝, 等.珠江口水域秋季刺网的渔获组成及多样性分析[J].南方水产科学, 2016, 12(1):111-119. http://www.schinafish.cn/CN/abstract/abstract9289.shtml [5] 朱轶.三峡大坝运行前后西洞庭湖鱼类群落结构特征变化[J].湖泊科学, 2014, 26(6):844-852. doi: 10.18307/2014.0605 [6] 许友伟, 陈作志, 范江涛, 等.南沙西南陆架海域底拖网渔获物组成及生物多样性[J].南方水产科学, 2015, 11(5):76-81. http://www.schinafish.cn/CN/abstract/abstract9388.shtml [7] 韩耀全, 何安尤, 施军, 等.岩滩水域渔业生态环境及鱼类物种多样性现状[J].江苏农业科学, 2015, 43(3):208-212. [8] 杨尧尧, 李忠义, 吴强, 等.莱州湾渔业资源群落结构和多样性的年际变化[J].渔业科学进展, 2016, 37(1):22-29. doi: 10.11758/yykxjz.20150313003 [9] CHU C, MINNS C K, LESTER N P, et al. An updated assessment of human activities, the environment, and freshwater fish biodiversity in Canada[J].Can J Fish Aquat Sci, 2015, 72(14):135-148. doi: 10.1139/cjfas-2013-0609
[10] 蔡文倩, 刘录三, 乔飞, 等.渤海湾大型底栖生物群落结构变化及原因探讨[J].环境科学, 2012, 33(9):3104-3109. http://www.cqvip.com/QK/91181X/201209/43111923.html [11] 皮平凡.构建珠江三角洲会展经济带[J].经济论坛, 2005(6):10-12. http://www.cnki.com.cn/Article/CJFDTOTAL-JJLT200506006.htm [12] 李因强. 珠江口水域鱼类群落结构研究[D]. 湛江: 广东海洋大学, 2008: 50. http: //www. cnki. com. cn/Article/CJFDTOTAL-SEAC803. 012. htm [13] 袁俏君, 苗素英, 李恒翔, 等.珠江口水域夏季小型底栖生物群落结构[J].生态学报, 2012, 32(19):5962-5971. http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_stxb... [14] 林卫强, 李适宇.珠江口水域化学耗氧量, 溶解氧, 无机磷与有机磷的三维水质数学模拟[J].海洋学报, 2003, 25(3):129-137. http://www.cnki.com.cn/Article/CJFDTOTAL-SEAC200303014.htm [15] PINKAS L, OLIPHANT M S, IVERSON I K. Food habits of albacroe, bluefin tuna, and bonito in California waters[J].Cali Dep Fish Game, 1971, 152:1-105. http://www.worldcat.org/title/food-habits-of-albacore-bluefin-tuna-and...
[16] 单秀娟, 金显仕.长江口近海春季鱼类群落结构的多样性研究[J].海洋与湖沼, 2011, 42(1):32-40. doi: 10.11693/hyhz201101005005 [17] 单秀娟, 陈云龙, 戴芳群, 等.黄海中南部不同断面鱼类群落结构及其多样性[J].生态学报, 2014, 34(2):377-389. http://www.ecologica.cn/stxb/ch/html/2014/2/stxb201301110084.htm [18] MARGALEF R.Information theory in ecology[J].Gen Syst, 1958(3):36-71. doi: 10.1890/06-1736.1
[19] SHANNON C E, WEAVER W.The mathematical theory of communication[J].Addison-Wesley Pub, 1981, 97(3):29-125. doi: 10.1063/1.3067010
[20] PIELOU E C.The use of information theory in the study of ecological succession[J].J Theor Biol, 1966(10):370-383. https://www.cabdirect.org/cabdirect/abstract/20103209685
[21] 郑亮. 黄河口海域鱼类群落结构初步研究[D]. 上海: 上海海洋大学, 2014: 44. http: //cdmd. cnki. com. cn/Article/CDMD-10264-1014051578. htm [22] ZHAO S J, LU B Q, LI R W, et al.A preliminary analysis of fishery resource exhaustion in the context of biodiversity decline[J].Sci China Earth Sci, 2016, 59(2):223-235. doi: 10.1007/s11430-015-5193-4
[23] 汪洋.中街山列岛岩礁海域鱼类群落结构和生物多样性研究[J].海洋与湖沼, 2015, 46(4):776-785. http://cdmd.cnki.com.cn/Article/CDMD-10340-1015302110.htm [24] 王建明.龙穴岛渔业生产之探析[J].江西水产科技, 2003(5):36-37. http://www.cqvip.com/QK/82860X/200305/8891998.html [25] 刘修泽, 董婧, 于旭光, 等.辽宁省近岸海域的渔业资源结构[J].海洋渔业, 2014, 36(4):289-299. http://www.cqvip.com/QK/93342X/201404/661790431.html [26] MAO Z G, GU X H, ZENG Q F.The structure of fish community and changes of fishery resources in Lake Hulun[J].J Lake Sci, 2016, 28(2):387-394. doi: 10.18307/2016.0219
[27] 李永振, 陈国宝, 孙典荣.珠江口鱼类组成分析[J].水产学报, 2000, 24(4):312-317. http://www.cnki.com.cn/Article/CJFDTOTAL-SCKX200004003.htm [28] PIELOU E C.The measurement of diversity in different types of biological collections[J].J Theor Biol, 1966, 13:131-144. doi: 10.1016/0022-5193(66)90013-0
[29] 杨渡远.污染给珠江口渔业造成严重损失[J].江西水产科技, 1995(1):1-2. http://www.cqvip.com/QK/82860X/199501/12096066.html [30] 于杰, 陈作志, 徐姗楠.围填海对珠江口南沙湿地资源与生物资源的影响[J].中国水产科学, 2016, 23(3):661-671. http://www.docin.com/p-1754328777.html [31] 李开枝, 尹健强, 黄良民, 等.珠江口伶仃洋海域底层游泳动物的季节变化[J].生态科学, 2012, 31(1):1-7. http://www.cqvip.com/QK/97324X/201201/41092250.html [32] 殷名称.鱼类生态学[M].基隆:水产出版社, 1998:171-187. -
期刊类型引用(26)
1. 初建松,郑卫东,孙利元,彭道民,索安宁,朱玉贵. 基于生物资源的万山海洋牧场生境适宜性评估. 海洋科学. 2024(01): 75-84 .  百度学术
百度学术
2. 汪慧娟,徐姗楠,张文博,黄洪辉,齐占会,程琪,刘华雪. 基于碳氮稳定同位素的珠江口南沙海域渔业生物群落营养结构研究. 生态科学. 2024(02): 42-50 . 百度学术
3. 王颤. 珠江入海口空间生态性水体变化对鱼类多样性的影响. 湖北农业科学. 2024(04): 127-135 . 百度学术
4. 田思泉,柳晓雪,花传祥,王寅,杜涣洋. 南海渔业资源状况及其管理挑战. 上海海洋大学学报. 2024(03): 786-798 . 百度学术
5. 马菁菁,陈海刚,张喆,田斐,唐振朝,熊倩,张林宝. 2018—2023年珠江口鱼类群落结构变化及其与环境因子的关系. 南方水产科学. 2024(06): 62-73 . 本站查看
6. 谢雨芳,吴鹏,刘永,肖雅元,唐广隆,王腾,林琳,李纯厚. 珠江河口凤鲚的栖息地适宜性评价. 南方水产科学. 2023(01): 22-29 . 本站查看
7. 刘文瑜,黄建强,戴国贤,刘国强,雷学铁,周永东,叶永昌. 珠江口伶仃洋春季鱼类群落结构及其与环境因子的RDA分析. 渔业信息与战略. 2023(01): 32-41 . 百度学术
8. 裴精花,陈清华,范金金,刘伟杰,郭照良,隋昊志. 珠江口海域游泳动物群落结构及多样性特征. 南方农业学报. 2023(12): 3727-3738 . 百度学术
9. 蒋佩文,李敏,张帅,陈作志,徐姗楠. 基于线粒体COI和12S rDNA基因构建珠江河口鱼类DNA宏条形码数据库. 南方水产科学. 2022(03): 13-21 . 本站查看
10. 唐广隆,刘永,吴鹏,孙典荣,肖雅元,王腾,谢雨芳,李纯然,石娟,钟智辉,李纯厚. 珠江口万山群岛海域春季渔业资源群落结构特征及其与环境因子的关系. 中国水产科学. 2022(08): 1198-1209 . 百度学术
11. 李雷,马波,金星,金洪宇,吴松,陈中祥,程磊,王念民,郝其睿. 西藏雅鲁藏布江大峡谷墨脱江段鱼类群落结构及多样性的空间分布特征. 中国水产科学. 2022(09): 1326-1336 . 百度学术
12. 蒋佩文,李敏,张帅,陈作志,徐姗楠. 基于环境DNA宏条码和底拖网的珠江河口鱼类多样性. 水生生物学报. 2022(11): 1701-1711 . 百度学术
13. 周卫国,丁德文,索安宁,何伟宏,田涛. 珠江口海洋牧场渔业资源关键功能群的遴选方法. 水产学报. 2021(03): 433-443 . 百度学术
14. 徐鹏,谢木娇,周卫国,孙英婷,丁德文,索安宁. 近30年珠江口海域游泳动物经济物种群落结构变化特征. 应用海洋学学报. 2021(02): 239-250 . 百度学术
15. 熊朋莉,陈作志,侯刚,张帅,邱永松,范江涛,徐姗楠. 珠江河口棘头梅童鱼生物学特征的年代际变化. 南方水产科学. 2021(06): 31-38 . 本站查看
16. 于南京,俞存根,菅康康,许永久,孙蓓蓓,张佩怡,刘坤,王慧君. 嵊泗列岛邻近海域鱼类种类组成及多样性分析. 渔业研究. 2020(04): 293-301 . 百度学术
17. 李嘉盛,刘复荣,刘明珠,张宏刚,崔金明,蒙海林. 珠江口水产源乳酸菌资源库的构建及优良乳酸菌的高通量筛选. 食品安全质量检测学报. 2020(16): 5627-5634 . 百度学术
18. 林坤,麦广铭,王力飞,王学锋. 2015—2018年珠江口近岸海域鱼类群落结构及其稳定性. 水产学报. 2020(11): 1841-1850 . 百度学术
19. 李娟,朱勤东. 福建省县域渔业地理时空格局演变研究. 测绘与空间地理信息. 2019(03): 47-52+57 . 百度学术
20. 刘岩,吴忠鑫,杨长平,单斌斌,刘胜男,孙典荣. 基于Ecopath模型的珠江口6种增殖放流种类生态容纳量估算. 南方水产科学. 2019(04): 19-28 . 本站查看
21. 应锐,蒋力,尹芳敏,黄建荣,黎祖福,王英永,冯建祥. 基于刺网、地笼的考洲洋游泳动物群落结构和多样性分析. 南方水产科学. 2019(04): 1-10 . 本站查看
22. 郭建忠,陈作志,田永军,张魁,许友伟,徐姗楠,李纯厚. 胶州湾海域鱼类群落种类组成及多样性. 生态学报. 2019(19): 7002-7013 . 百度学术
23. 蔡研聪,徐姗楠,陈作志,许友伟,江艳娥,杨长平. 南海北部近海渔业资源群落结构及其多样性现状. 南方水产科学. 2018(02): 10-18 . 本站查看
24. 曾雷,陈国宝,于杰. 南澳岛海域渔业资源声学评估与空间分布. 南方水产科学. 2018(02): 26-35 . 本站查看
25. 谢志超,孙典荣,刘永,林琳,王腾,肖雅元,李纯厚. 江门海域游泳动物群落组成及其多样性初步分析. 南方水产科学. 2018(05): 21-28 . 本站查看
26. 黄卉,杨丽芝,杨贤庆,李来好,郝淑贤,魏涯,王锦旭. 南海鸢乌贼墨汁多糖分离纯化及组分分析. 食品科学. 2017(24): 118-123 . 百度学术
其他类型引用(11)




计量
- 文章访问数: 3715
- HTML全文浏览量: 1150
- PDF下载量: 815
- 被引次数: 37
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
