
A study on settlement performance of falling-net based on GAM
-
摘要: 为全面了解罩网网具沉降性能,为罩网模型试验的开展及渔具渔法的改进提供参考,该研究于2016年3月在南海中南部进行海上试验,采用多项式回归和广义加性模型 (Generalized Additive Model, GAM) 对罩网沉降性能进行分析研究。结果显示:1) 自由沉降是影响网具总沉降深度的更为关键的因素。2) 90.0~119.9 s是较为适宜的自由沉降时间。3) 自由沉降速度先短时间内增大后逐渐减小,最大值出现在沉降第10 秒;阻尼沉降速度在前50 s相对稳定,之后迅速降低。4) 自由沉降时间、风速、风流向夹角对自由沉降深度影响显著;自由沉降时间、风速、渔船漂流速度、风流向夹角对阻尼沉降深度影响显著。研究表明:1) 风速、风流向夹角对自由沉降深度与阻尼沉降深度的影响是复杂且非线性的,它们可能通过波浪和改变船、网相对位置影响网具沉降。2) 渔船漂流速度对阻尼沉降深度影响显著,它可能会影响罩网网口的闭合程度与网具绞收。Abstract: In order to comprehensively understand the settlement performance of falling-net, as well as to provide references for the development of falling-net model test and improvement of fishing gear and method, we conducted sea trials in the central-southern South China Sea in March 2016, and studied the settlement performance of falling-net based on polynomial regression and Generalized Additive Model (GAM). The results show that: 1) Free settlement was a more critical factor affecting the total settlement depth. 2) 90.0–119.9 s was an appropriate free settlement time based on the settlement performance of the falling-net. 3) The free settlement speed first increased in a short time and then decreased, the maximum value occurring at 10th s. The damping settlement speed remained relatively stable in the first 50 s with little change, but then decreased rapidly. 4) Free settlement time (Tf), wind speed (Sw), and direction angle between wind and current (
$ \alpha $ ) had significant influences on free settlement depth. Tf, Sw, drift speed of fishing boat (Sb) and$ \alpha $ had significant influences on damping settlement depth. The study suggests that: 1) The influences of Sw and$ \alpha $ on free settlement depth and damping settlement depth are complex and non-linear, and may affect the settlement of falling-net through waves and changing the relative position between fishing boat and falling-net. 2) Sb has a significant influence on damping settlement depth. This may be caused by the fact that Sb affects the degree of closure of net-mouth and hauling of purse line.-
Keywords:
- Falling-net /
- Settlement performance /
- Generalized Additive Model
-
黄鳍金枪鱼 (Thunnus albacares) 隶属于鲭科、金枪鱼属,是一种喜集群的大洋性洄游鱼类,因背鳍、臀鳍及小鳍同为橘黄色,又名黄鳍鲔[1]。广泛分布于太平洋、大西洋、印度洋的热带和亚热带海域[2]。众所周知,金枪鱼具有生长速度快、粗蛋白和不饱和脂肪酸含量高的特点,深受消费者喜爱[3]。金枪鱼的丰度还可以支持珊瑚礁、沿海渔业和海水养殖等产业的可持续开发和利用[4]。其种群是组成远洋生态系统的关键部分,也是世界上最大和最有价值的渔业之一[5]。
对水生动物来说,pH是一个重要的环境因子,其极细微的变化也会产生深远影响[6];从而引起生态系统内一系列的化学变化,影响大多数水产生物的生长、繁殖、代谢与生存[7]。当水体pH下降到一定程度,超过机体的最大调控范围,就会出现细胞内酸中毒和环境高碳酸血症[8]。二氧化碳 (CO2)排放过量引起的海洋酸化改变了海洋环境,扰乱了海洋动物机体的新陈代谢、酸碱平衡和生物体内多种酶的活性[9]。有研究表明,水体酸化会使金头鲷 (Sparus aurata) 的抗氧化能力受到抑制[10];过酸的养殖水体会显著影响脊尾白虾 (Exopalaemon carinicauda) 的非特异性免疫力[11];酸化导致了青鳉 (Oryzias melastigm) 幼鱼脂质代谢紊乱[12];严重的酸化条件与舌齿鲈 (Dicentrarchus labrax) 显著升高的最大代谢率有关[13];海洋酸化会导致金枪鱼的生物量减少[14]。
大气中过多的CO2被海水吸收后形成碳酸,增加了海水酸度,然而少有酸化胁迫对黄鳍金枪鱼幼鱼生理特性影响的报道。随着捕捞量的增加,金枪鱼的资源量急剧下降,无法满足市场需求。本研究通过测定相关酶活性,评估海水酸化胁迫对黄鳍金枪鱼幼鱼抗氧化防御系统和免疫功能的影响,为海洋生态环境和金枪鱼野生种群的保护提供参考。
1. 材料与方法
1.1 实验材料
黄鳍金枪鱼幼鱼平均体长为 (18.21±1.09) cm、平均体质量为 (354.98±149.77) g,由中国水产科学研究院南海水产研究所热带水产研究开发中心提供。实验用水为砂滤后的自然海水,水温 (22.5±1.0) ℃,溶解氧质量浓度 (7.5±0.5) mg·L−1,盐度 (33.00±0.80)‰,氨氮质量浓度 <0.05 mg·L−1,亚硝酸盐质量浓度 <0.05 mg·L−1。
1.2 实验设计
正式实验开始前,先将金枪鱼幼鱼在驯养池中暂养7 d,期间投喂冰鲜杂鱼。随机挑选72尾行为活泼、体表健康的幼鱼,在直径和水深均为1.5 m的圆形钢化桶中进行48 h的酸化胁迫实验,暂养期间与正式实验期间水质保持一致。实验共设置4组,以自然海水pH 8.1为对照组,pH 7.6、7.1、6.6为实验组,每组设3个重复,每个重复6尾鱼。实验期间金枪鱼幼鱼处于停食状态。并通过1.0 mmol·L−1的氢氧化钠 (NaOH) 溶液或1.0 mmol·L−1的盐酸 (HCl) 溶液调节海水pH。使用pH分析仪 (PH848) 测定海水酸度,每2 h矫正1次,使pH变化幅度保持在 ±0.1。实验至第48小时,统计死亡的幼鱼数量 (以鱼体侧躺、沉入水底不再游动为准),计算成活率。
1.3 样品采集与处理
实验结束后每组随机选取6尾黄鳍金枪鱼幼鱼,经丁香酚麻醉后进行解剖,解剖全程在冰盒上进行。分别采集肝、鳃、红肌、皮肤组织于2 mL无菌冻存管中,置于液氮中冷冻,之后保存在−80 ℃冰箱中用于后续免疫和代谢酶的测定。准确称取组织0.1~0.2 g,加入9倍体积的0.9% (w) 盐水,冰水浴条件下机械匀浆后,3 000 r·min−1离心10 min,取上清液测定相关酶活性和总蛋白含量。选取丙二醛 (MDA) 为抗氧化物质的标志物;超氧化物歧化酶 (SOD)、过氧化氢酶 (CAT)、谷胱甘肽过氧化物酶 (GSH-Px) 和过氧化物酶 (POD) 作为抗氧化防御系统的标志物;酸性磷酸酶 (ACP)、碱性磷酸酶 (AKP)、溶菌酶 (LZM) 作为免疫系统的标志物。上述所有检测均使用南京建成生物工程研究所试剂盒,所有指标的测定严格按照说明书进行操作。采用酶标仪 (SYNERGY-HT) 或分光光度计 (UV-1800BPC) 测定各项指标。
1.4 数据处理
采用Excel 2010软件整理数据,以“平均值±标准差 ($\overline { x}\pm s $) ”表示 (n=3)。实验结果通过SPSS 25.0软件进行单因素方差分析 (One-way ANOVA),并用邓肯检验法分析组间差异性,显著性水平α为0.05。使用Origin 2022软件作图。
2. 结果
实验至第48小时,仅pH 6.6处理组的幼鱼出现死亡,成活率为83%。各组之间无显著性差异 (P>0.05,图1)。
![]() 图 1 酸化胁迫对黄鳍金枪鱼幼鱼成活率的影响注:相同字母表示组间无显著性差异 (P>0.05)。Figure 1. Effect of acidification stress on survival rate of juvenile yellowfin tunaNote: Same letters represent no significant differences among groups (P>0.05).
图 1 酸化胁迫对黄鳍金枪鱼幼鱼成活率的影响注:相同字母表示组间无显著性差异 (P>0.05)。Figure 1. Effect of acidification stress on survival rate of juvenile yellowfin tunaNote: Same letters represent no significant differences among groups (P>0.05).2.1 酸化胁迫对黄鳍金枪鱼幼鱼抗氧化能力的影响
与对照组相比,各实验组皮肤的MDA含量均有所下降,红肌和鳃的MDA含量均有所上升。除对照组外,相同pH下肝脏中的MDA含量均显著高于其他组织 (P<0.05),且随着pH的降低呈上升趋势,在pH 6.6时含量最高 (图2)。
![]() 图 2 酸化胁迫对黄鳍金枪鱼幼鱼丙二醛含量的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 2. Effect of acidification stress on MDA of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with same pH (P<0.05).
图 2 酸化胁迫对黄鳍金枪鱼幼鱼丙二醛含量的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 2. Effect of acidification stress on MDA of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with same pH (P<0.05).随着pH的降低,皮肤中的SOD活性呈先下降再上升的趋势,肝中的SOD活性呈下降趋势,二者均在pH 8.1时达到最高。红肌中的SOD活性在pH 7.6时最高。鳃中的SOD活性在pH 7.1时达到最高,且随着pH的降低呈先上升后下降的趋势;除对照组外,相同pH下鳃中的SOD活性均显著高于其他组织 (P<0.05,图3-a)。
![]() 图 3 酸化胁迫对黄鳍金枪鱼幼鱼超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶和过氧化物酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 3. Effects of acidification stress on SOD, CAT, GSH-Px and POD activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).
图 3 酸化胁迫对黄鳍金枪鱼幼鱼超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶和过氧化物酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 3. Effects of acidification stress on SOD, CAT, GSH-Px and POD activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).随着pH的降低,皮肤中的CAT活性呈下降趋势,在pH 8.1时活性最高。鳃和红肌中的CAT活性呈先上升再下降的趋势,在pH 7.1时达到最高;除对照组外,相同pH下鳃中的CAT活性均显著高于其他组织 (P<0.05)。肝中的CAT活性变化平稳,在pH 6.6时达到最高。相同pH下,肝和鳃中的CAT活性均显著高于皮肤和红肌 (P<0.05,图3-b)。
随着pH的降低,皮肤、红肌和鳃中的GSH-Px活性变化相似,均呈先上升后下降的趋势,且均在pH 7.1时达到最高,此时皮肤中的GSH-Px活性显著高于其他组织 (P<0.05)。肝中的GSH-Px活性呈上升趋势,在pH 6.6时达到最高 (图3-c)。
随着pH的降低,皮肤中的POD活性呈下降趋势,在pH 8.1时活性最高。红肌中各实验组的POD活性均高于对照组,在pH 6.6时达到最高。鳃中的POD活性呈先上升后下降的趋势,在pH 7.1时达到最高;相同pH下鳃中的POD活性均显著高于其他组织 (P<0.05)。肝中的POD活性呈先下降再上升的趋势,在pH 8.1时活性最高。除对照组外,相同pH下红肌和鳃中的POD活性均显著高于皮肤和肝 (P<0.05,图3-d)。
2.2 酸化胁迫对黄鳍金枪鱼幼鱼免疫酶活性的影响
随着pH的降低,肝和皮肤各实验组的ACP活性与对照组相比均有所下降;相同pH下肝中的ACP活性均显著高于其他组织 (P<0.05)。红肌中的ACP活性呈先上升后下降再上升的趋势,在pH 7.6时达到最高;鳃中的ACP活性呈先上升后下降的趋势,在pH 7.1时达到最高。相同pH下鳃中的ACP活性均显著低于其他组织 (P<0.05,图4-a)。
![]() 图 4 酸化胁迫对黄鳍金枪鱼幼鱼酸性磷酸酶、碱性磷酸酶和溶菌酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 4. Effects of acidification stress on ACP, AKP and LZM activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).
图 4 酸化胁迫对黄鳍金枪鱼幼鱼酸性磷酸酶、碱性磷酸酶和溶菌酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 4. Effects of acidification stress on ACP, AKP and LZM activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).随着pH的降低,鳃、红肌和皮肤中的AKP活性呈先下降后上升的趋势,均在pH 8.1时最高。肝中的AKP活性呈上升趋势,在pH 7.1时达到最高。相同pH下红肌中的AKP活性均显著低于其他组织 (P<0.05,图4-b)。
随着pH的降低,皮肤中的LZM活性呈先上升后下降的趋势,在pH 7.1时达到最高。鳃和红肌中的LZM活性呈上升趋势,均在pH 6.6时达到最高;相同pH下鳃中的LZM活性均显著高于其他组织 (P<0.05)。肝中的LZM活性在pH 7.1时达到最高 (图4-c)。
3. 讨论
3.1 环境胁迫对水产动物不同组织抗氧化能力的影响
MDA是脂质过氧化的重要产物[15],反映机体受氧化损伤的程度[16],其含量随着机体氧化应激水平升高而增加,含量越高表明鱼体所受的压力越大[17]。肝脏有排泄、解毒等多种功能,可清除外来有害物质和机体产生的有害废物[18];且含有大量与免疫相关的细胞,具有重要的免疫调节功能,在机体的免疫应答中发挥重要作用[19]。本研究中肝脏的MDA含量随海水酸度升高呈上升趋势,且累积含量显著高于其他组织。可能是由于金枪鱼处于应激状态,体内产生大量的活性氧自由基,脂质过氧化水平升高,机体出现了氧化损伤,与克氏原螯虾 (Procambarus clarkii)[20]的结果相似。
SOD和CAT活性的变化可在一定程度上反映出机体受环境胁迫时免疫机能的变化[21]。GSH-Px在清除过氧化氢 (H2O2)、把脂质过氧化物还原成无毒产物方面起着至关重要的作用[22],其与CAT在清除自由基方面既是互补也是竞争关系[23]。SOD、CAT和GSH-Px协同作用,是抗氧化系统的重要组成部分[24]。POD和生物应激相关,可水解H2O2,对细胞起保护作用[25]。
鳃是鱼类主要的呼吸和免疫器官,表面积大且与水体直接接触,易受有害物质伤害[26];其黏膜中含有丰富的免疫分子,在抵御外界侵袭中发挥重要作用[27]。本研究中,各处理组鳃中的SOD、CAT和POD活性显著高于其他组织,且在pH 7.1时达到最高。说明酸性胁迫下金枪鱼组织中的活性氧自由基含量升高,诱导SOD、CAT和POD的活性升高,鱼体通过激活抗氧化酶系统促进蛋白质合成,改善代谢产物,保护细胞不受损伤,从而使鱼体能够适应低pH环境,达到新的动态平衡。但水体酸度过高时,鱼体会产生过量的氧自由基 (·O2−),机体无法产生足够的SOD和CAT来清除 ·O2−,且动物机体在受到环境胁迫时,SOD和CAT可能会发生抑制的应激变化[28],因此本研究鳃中的CAT活性在pH 6.6时有所回落。与沙塘鳢 (Odontobutis potamophila)[29]、瓦氏黄颡鱼 (Pelteobagrus vachelli)[30]和方斑东风螺 (Babylonia areolata)[31]的研究结果相似。
鱼类皮肤的腺层内含有丰富的腺细胞,分泌的黏液中含有丰富的免疫因子,如溶菌酶、补体类物质等[32],构成了机体的第一道防线[33]。本研究中,皮肤的GSH-Px活性在pH 7.1时达到最高,显著高于其他组织,而CAT活性低于其他组织,可能是由于H2O2没有被CAT及时分解,诱导机体生成更多的GSH-Px来清除多余的H2O2。GSH-Px的活性升高,谷胱甘肽 (GSH) 等抗氧化物质就会消耗过大,可能导致机体受到损伤[34]。本研究中皮肤的GSH-Px活性在pH 7.1时大幅上升,推测此时鱼体无法适应过酸的海水,机体出现了损伤。与尖吻鲈 (Lates calcarifer) 幼鱼[35]和点篮子鱼 (Siganus guttatus) 幼鱼[36]的表现类似。
3.2 环境胁迫对水产动物不同组织免疫酶的影响
磷酸酶又称正磷酸单酯水解酶,可以催化各种含磷化合物的水解,根据他们的特性 (即起催化作用的最适pH),可分为ACP和AKP两类[37],前者主要起催化效果,后者主要起调理和调节的作用[38]。当水体中溶解氧含量低于正常水平时,鱼体发生应激反应,并通过提高AKP和ACP的活性来增强非特异性免疫力[39]。
本研究中,肝脏的ACP活性显著高于其他组织,且随着pH的降低呈下降趋势;但AKP活性呈上升趋势,可能因为二者在肝脏中的作用不同。分析出现上述结果的原因,可能是海水酸度过高,机体为了维持酸碱和离子平衡而采取的一种调节措施,是生物体主动防御的表现。类似的报道还有脊尾白虾[11]和栉孔扇贝 (Chlamys farreri)[21]。鳃中的ACP和AKP变化不一致,可能是此时的环境pH影响了鳃细胞参与免疫的水解酶活性。类似的结果还见于翘嘴鳜 (Siniperca chuatsi) 幼鱼[40]和缢蛏 (Sinonovacula constricta) 成贝[41]。
LZM是衡量鱼类非特异性免疫的指标之一[42],在面对外界环境胁迫时,其活性会升高[43],活性越强,机体的非特异性免疫力就越强[44]。鳃中的LZM活性显著高于其他组织,且随海水酸度的升高呈上升趋势。可能是由于海水酸度的增加使金枪鱼处于应激状态,鱼体通过增加LZM的活性以抵抗外源环境的压力,属于自身的应急保护反应。
4. 结论
本研究通过设置4组酸化梯度 (pH 8.1、7.6、7.1、6.6) 对黄鳍金枪鱼幼鱼进行48 h的环境胁迫,结果显示,酸化胁迫对黄鳍金枪鱼幼鱼的免疫功能和抗氧化防御系统影响显著。抗氧化物质在pH 6.6时达到最高;抗氧化酶的活性多数在pH 7.1时达到最高;非特异性免疫酶变化不一,不同酶的活性因组织不同而呈类似或不同的变化趋势。提示黄鳍金枪鱼幼鱼在pH大于7.1的酸化环境中有一定的调节能力,pH小于7.1时免疫系统发生紊乱。整体而言,肝脏和鳃中的酶活性高于红肌和皮肤,这表明在面对海水酸度升高时,肝脏和鳃中的抗氧化酶和免疫酶能更好地表达,二者抗氧化酶和免疫酶的变化可以代表黄鳍金枪鱼幼鱼机体免疫功能和抗氧化防御系统的变化。
-
![]()
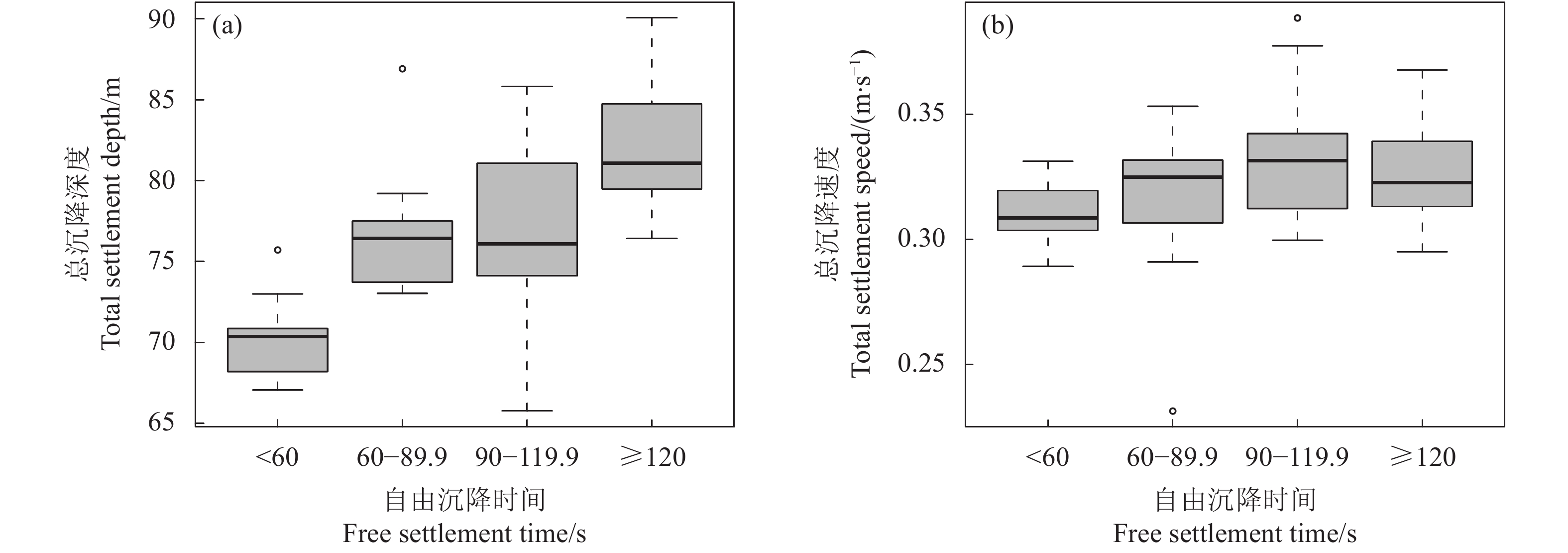
图 3 不同自由沉降时间对应的总沉降深度、速度的箱型图
Figure 3. Box plots of total settlement depth speed at different free settlement time
![]()
图 4 自由沉降深度、自由沉降速度、阻尼沉降深度和阻尼沉降速度随时间变化趋势图
Figure 4. Trend chart of free settlement depth, free settlement speed, damping settlement depth and damping settlement speed along with time
![]()
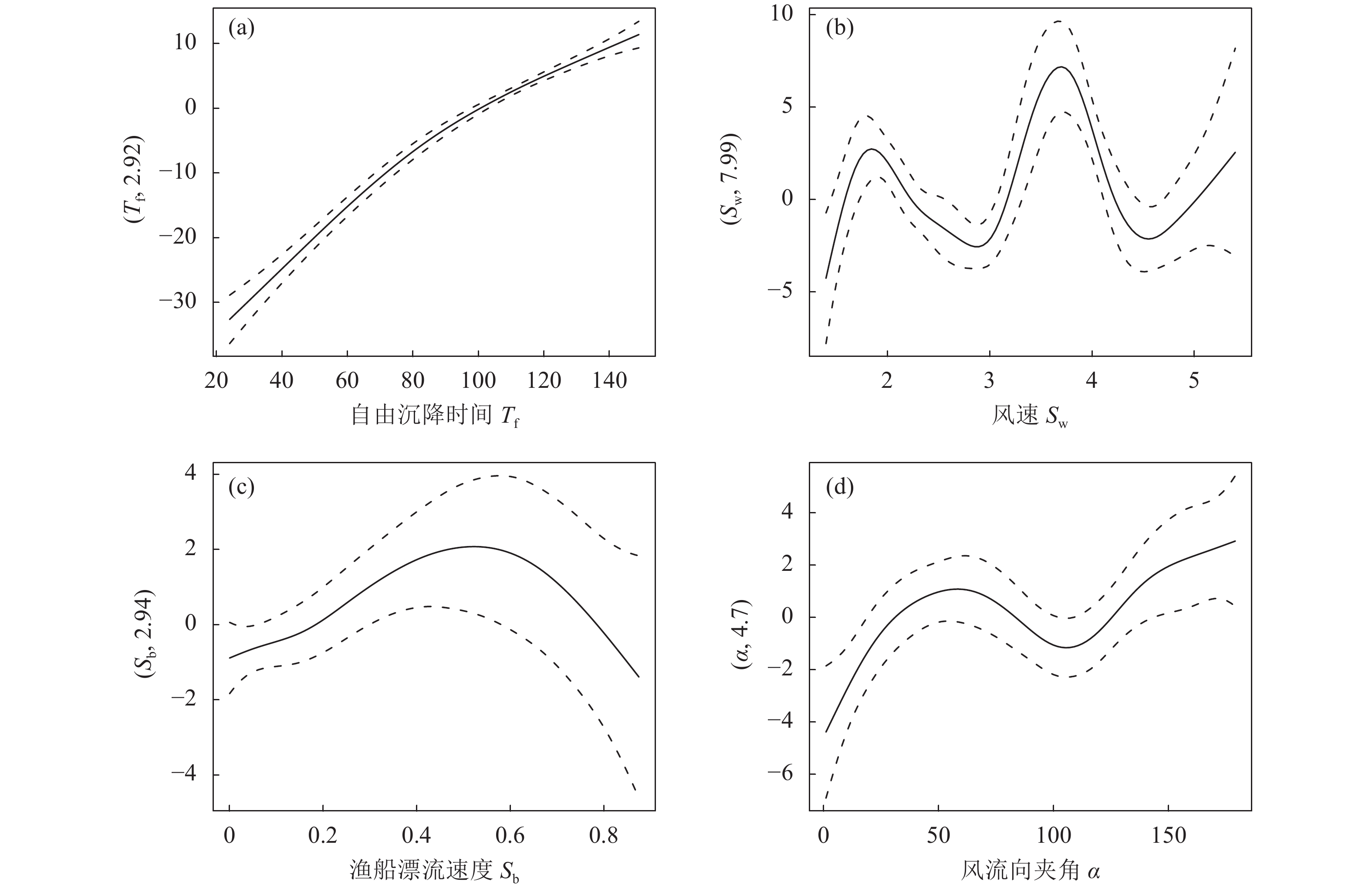
图 6 自由沉降深度与4个因子间的广义加性模型分析图
Figure 6. GAM-derived effects of four predictors on free settlement depth
![]()
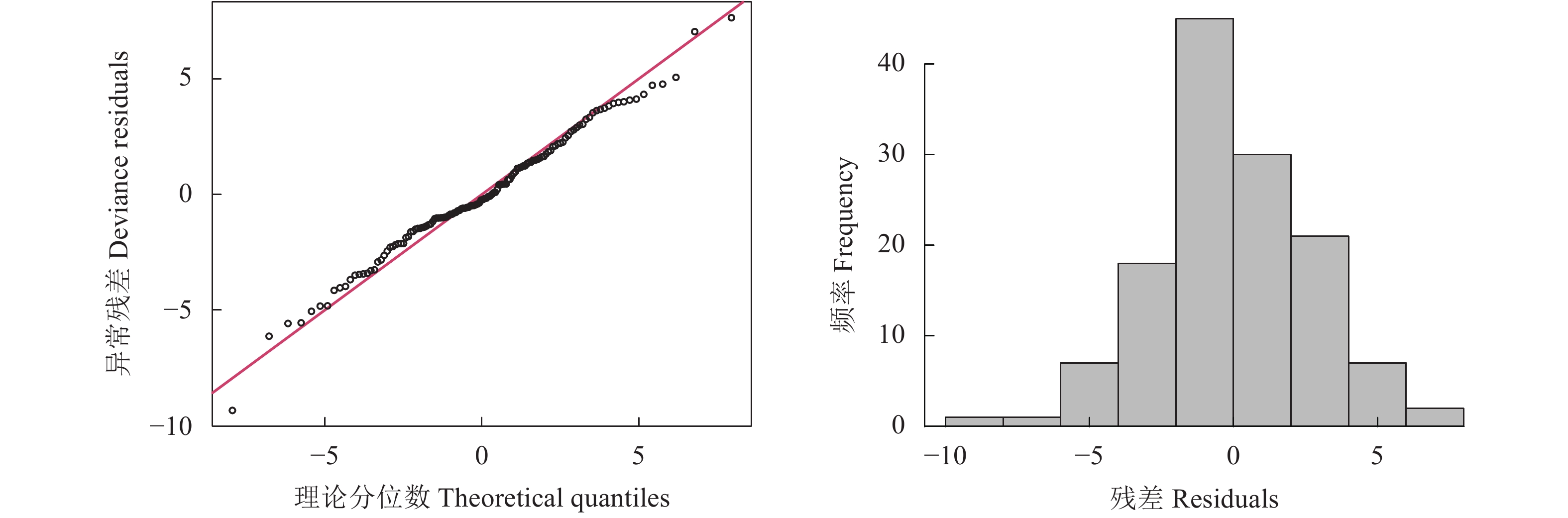
图 7 阻尼沉降深度最优广义加性模型残差图
Figure 7. Residual plot of optimal GAM of damping settlement depth
![]()
图 8 阻尼沉降深度与4个因子间的广义加性模型分析图
Figure 8. GAM-derived effects of four predictors on damping settlement depth
表 1 自由沉降深度广义加性模型统计参数
Table 1 Statistical characteristics of GAM of free settlement depth
公式
Formular决定系数
R2方差解释
Deviance explained/%赤池信息准则
AIC自由沉降深度~自由沉降时间 Df−Tf 0.873 87.600 717.038 自由沉降深度~自由沉降时间+风速 Df−Tf+Sw 0.904 91.300 687.941 自由沉降深度~自由沉降时间+风速+渔船漂流速度 Df−Tf+Sw+Sb 0.911 92.100 680.633 自由沉降深度~自由沉降时间+风速+渔船漂流速度+风流向夹角 Df−Tf+Sw+Sb+$ \alpha $ 0.922 93.300 666.362  下载: 导出CSV
下载: 导出CSV
表 2 自由沉降深度最优广义加性模型检验值
Table 2 F-test for significance of non-parametric effects on optimal GAM of free settlement depth
n=132
下载: 导出CSV
表 3 阻尼沉降深度广义加性模型统计参数
Table 3 Statistical characteristics of GAM of damping settlement depth
公式
Formular决定系数
R2方差解释
Deviance explained/%赤池信息准则
AIC阻尼沉降深度~自由沉降时间 Dd−Tf 0.706 71.700 755.699 阻尼沉降深度~自由沉降时间+风速 Dd−Tf+Sw 0.797 82.300 716.660 阻尼沉降深度~自由沉降时间+风速+渔船漂流速度 Dd−Tf+Sw+Sb 0.840 86.400 688.788 阻尼沉降深度~自由沉降时间+风速+渔船漂流速度+风流向夹角 Dd−Tf+Sw+Sb+$ \alpha $ 0.846 87.000 684.163
下载: 导出CSV
表 4 阻尼沉降深度最优广义加性模型检验值
Table 4 F-test for significance of non-parametric effects on optimal GAM of damping settlement depth
n=132 因子
Factor估计自由度
edfF P 自由沉降时间 Tf 3.007 57.289 <2.00×10−16* 风速 Sw 7.860 6.220 6.00×10−6* 渔船漂流速度 Sb 8.519 6.623 1.06×10−7* 风流向夹角 $ \alpha $ 1.000 5.395 0.022*
下载: 导出CSV
-
[1] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. [2] 杨吝, 张旭丰, 张鹏, 等. 南海区海洋小型渔具渔法[M]. 广州: 广东科技出版社, 2007: 103-115. [3] 张鹏, 曾晓光, 杨吝, 等. 南海区大型灯光罩网渔场渔期和渔获组成分析[J]. 南方水产科学, 2013, 9(3): 74-79. doi: 10.3969/j.issn.2095-0780.2013.03.012 [4] 冯波, 李忠炉, 侯刚, 等. 南海大眼金枪鱼和黄鳍金枪鱼生物学特性及其分布[J]. 海洋与湖沼, 2014, 45(4): 886-894. doi: 10.11693/hyhz20130600081 [5] 张鹏, 陈森, 李杰, 等. 灯光罩网渔船兼作金枪鱼延绳钓捕捞试验[J]. 南方水产科学, 2016, 12(4): 110-116. doi: 10.3969/j.issn.2095-0780.2016.04.014 [6] 许柳雄, 王敏法, 叶旭昌, 等. 金枪鱼围网沉降特性[J]. 中国水产科学, 2011, 18(5): 1161-119. [7] 晏磊, 张鹏, 杨吝, 等. 南海灯光罩网沉降性能研究[J]. 上海海洋大学学报, 2014, 23(1): 146-153. [8] 唐浩, 许柳雄, 周成, 等. 基于GAM模型研究金枪鱼围网沉降性能影响因子[J]. 水产学报, 2013, 37(6): 944-950. [9] 唐浩, 许柳雄, 王学昉, 等. 两种典型渔法金枪鱼围网网具性能差异[J]. 水产学报, 2015, 39(2): 275-283. [10] WIDAGDO A, LEE C W, LEE J. Calculating and measuring the sinking performance of small-scale purse seine gear in Java, Indonesia, to improve the gear[J]. Fish Aquat Sci, 2015, 18(2): 221-227.
[11] 徐国强, 许柳雄, 周成, 等. 金枪鱼围网下纲提升特性的研究[J]. 南方水产科学, 2015, 11(3): 22-28. doi: 10.3969/j.issn.2095-0780.2015.03.004 [12] TANG H, XU L X, ZHOU C, et al. The effect of environmental variables, gear design and operational parameters on sinking performance of tuna purse seine setting on free-swimming schools[J]. Fish Res, 2017, 196: 151-159. doi: 10.1016/j.fishres.2017.08.006
[13] 许柳雄, 兰光查, 叶旭昌, 等. 下纲重量和放网速度对金枪鱼围网下纲沉降速度的影响[J]. 水产学报, 2011, 35(10): 1563-1571. [14] ZHOU C, XU L X, HU F X, et al. The kinetic deformation of tuna purse seine: a model experiment on different shooting patterns at uniform current[J]. Fish Res, 2015, 169: 18-25. doi: 10.1016/j.fishres.2015.04.008
[15] 李灵智, 黄洪亮, 陈帅, 等. 基于静水池模型试验的金枪鱼围网沉降性能研究[J]. 中国海洋大学学报, 2015, 45(3): 48-53. [16] 唐浩, 许柳雄, 王学昉, 等. 基于网具模型试验的金枪鱼围网性能分析[J]. 中国水产科学, 2015, 22(3): 563-573. [17] TANG H, XU L X, HU F X, et al. Effect of mesh size modifications on the sinking performance, geometry and forces acting on model purse seine nets[J]. Fish Res, 2019, 211: 158-168. doi: 10.1016/j.fishres.2018.11.018
[18] LEE C W, LEE J H, CHA B J, et al. Physical modeling for underwater flexible systems dynamic simulation[J]. Ocean Eng, 2005, 32(3/4): 331-347.
[19] KIM H Y, LEE C W, SHIN J K, et al. Dynamic simulation of the behavior of purse seine gear and sea-trial verification[J]. Fish Res, 2007, 88(1/2/3): 109-119. doi: 10.1016/j.fishres.2007.08.007
[20] HOSSEINI S A. The sinking performance of the tuna purse seine gear with large-meshed panels using numerical method[J]. Fish Sci, 2011, 77(4): 503-520. doi: 10.1007/s12562-011-0371-6
[21] RIZIOTIS V A, KATSAOUNIS G M, PAPADAKIS G, et al. Numerical and experimental analysis of the hydroelas behavior of purse seine nets[J]. Ocean Eng, 2013, 58: 88-105. doi: 10.1016/j.oceaneng.2012.09.022
[22] ZHOU C, XIU L X, ZHANG X F, et al. Application of numerical simulation for analysis of sinking characteristics of purse seine[J]. J Ocean Univ China, 2015, 14(1): 135-142. doi: 10.1007/s11802-015-2384-8
[23] 李杰, 晏磊, 陈森, 等. 灯光罩网网口沉降与闭合性能研究[J]. 南方水产科学, 2015, 11(5): 117-124. doi: 10.3969/j.issn.2095-0780.2015.05.014 [24] 李杰, 晏磊, 陈森, 等. 基于不同配重的罩网沉降性能研究[J]. 南方水产科学, 2016, 12(5): 16-22. doi: 10.3969/j.issn.2095-0780.2016.05.003 [25] 李杰, 晏磊, 杨炳忠, 等. 罩网自由沉降过程的数值模拟研究[J]. 南方水产科学, 2017, 13(4): 105-114. doi: 10.3969/j.issn.2095-0780.2017.04.013 [26] 晏磊, 李杰, 杨炳忠, 等. 光诱罩网沉降性能对鸢乌贼渔获率的影响[J]. 南方水产科学, 2018, 14(4): 10-16. doi: 10.3969/j.issn.2095-0780.2018.04.002 [27] ZAGAGLIA C R, LORENZZETTI J A, STECH J L. Remote sensing data and longline catches of yellowfin tuna (Thunnus albacares) in the equatorial Atlantic[J]. Remote Sens Environ, 2004, 93(1/2): 267-281.
[28] CHIU T Y, CHIU T S, CHEN C S. Movement patterns determine the availability of Argentine shortfin squid Illex argentinus to fisheries[J]. Fish Res, 2017, 193: 71-80. doi: 10.1016/j.fishres.2017.03.023
[29] 杨胜龙, 范秀梅, 伍玉梅, 等. 基于GAM模型的阿拉伯海鲐鱼渔场分布与环境关系[J]. 生态学杂志, 2019, 38(8): 2466-2470. [30] 石永闯, 朱清澄, 花传祥, 等. 基于海上实测的秋刀鱼舷提网沉降和提升性能研究[J]. 海洋通报, 2018, 37(4): 459-467. doi: 10.11840/j.issn.1001-6392.2018.04.012 [31] 武胜男, 陈新军, 刘祝楠. 基于GAM的西北太平洋日本鲭资源丰度预测模型建立[J]. 海洋学报, 2019, 41(8): 36-42. [32] 石永闯, 朱清澄, 花传祥, 等. 秋刀鱼舷提网渔具性能模型试验与海上实测结果的比较评估[J]. 海洋学报, 2019, 41(2): 123-133. -
期刊类型引用(11)
1. 肖佳程,赵一方,石耀华,顾志峰. 养殖密度对宝蓝鹦鹉鱼生长、生理与繁殖的影响. 热带生物学报. 2025(02): 304-311 .  百度学术
百度学术
2. 汪海燕,张永明,靳桂双,赵璐. 植物多糖的生物学功能及其在水产养殖中的应用. 饲料研究. 2024(09): 173-177 . 百度学术
3. 高建伟,汪东升,邵锋锋. 加减大柴胡汤对胆总管结石患者湿热证候及肝功能的影响. 中医药临床杂志. 2024(06): 1144-1148 . 百度学术
4. 夏伦斌,马龙龙,乔德亮,何燕飞,蒋平. 三角帆蚌多糖对肉仔鸡生长性能、抗氧化及免疫功能的影响. 浙江农业学报. 2023(03): 547-555 . 百度学术
5. 田瑜,揭育鵾,曾祥兵,乐燕,刘广鑫,程长洪,马红玲,郭志勋. 密度对拟穴青蟹抗氧化、非特异性免疫能力的影响. 南方水产科学. 2023(03): 60-67 . 本站查看
6. 刘清豪,张丹,陈卓,韦婉婷,王城城,金言,修彦凤. HPLC法同时测定炮制前后鸡内金中6种黄酮类成分. 中成药. 2023(08): 2618-2622 . 百度学术
7. 李成辉,董宏标,郑晓婷,桂福坤,曾祥兵,明俊超,陈飞,陈健,张家松. 春砂仁精油对尼罗罗非鱼幼鱼生长、消化、抗氧化能力和血清生化指标的影响. 南方水产科学. 2023(06): 51-59 . 本站查看
8. 黄健彬,迟艳,周传朋,黄小林,黄忠,虞为,荀鹏伟,吴杨,张宇,林黑着. 褐藻寡糖对卵形鲳鲹幼鱼生长性能、抗氧化能力和免疫功能的影响. 南方水产科学. 2022(03): 118-128 . 本站查看
9. 贾慧凝,侍苗苗,卞永乐,侍崇敬,刘恒蔚,宋学宏,秦粉菊. 纳米硒对低氧胁迫下中华绒螯蟹免疫保护和抗氧化能力的影响. 南方水产科学. 2022(06): 100-109 . 本站查看
10. 张慧,董宏标,孙彩云,陈健,黄聪灵,李勇,段亚飞,张家松. 饲料中添加月桂酸单甘酯对尖吻鲈脂质代谢与肝脏功能的影响. 海洋渔业. 2022(06): 736-746 . 百度学术
11. 范思远,张海瑜,周永雨,尹秀雯,刘紫嫣,魏峰. 香菇菌丝体多糖对肉鸡生长性能、免疫功能及抗氧化性能的影响. 饲料研究. 2021(23): 41-44 . 百度学术
其他类型引用(6)









计量
- 文章访问数: 802
- HTML全文浏览量: 275
- PDF下载量: 49
- 被引次数: 17
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
