
Feeding habits of Symplectoteuthis oualaniensis in the South China Sea
-
摘要:
根据2012年9月~10月南海灯光罩网调查所获得的鸢乌贼(Symplectoteuthis oualaniensis)样本,结合胃含物分析方法和分子检测技术,研究其饵料组成、摄食强度、摄食习性与胴长的关系等。结果表明,鸢乌贼以头足类、鱼类(主要为灯笼鱼科)和甲壳动物为主要摄食对象,同类相食现象明显。头足类和鱼类为鸢乌贼优势饵料生物类群,其中头足类相对重要性指数百分比(%IRI)、个数百分比(N%)、质量百分比(W%)和出现频率(F%)分别为54.38、28.73、58.39和75.60,而鱼类的%IRI、N%、W%和F%分别为41.39、42.54、36.17和63.69。鸢乌贼摄食等级主要以3、4级为主,胃饱满指数平均为3.11%,空胃率平均为2.89%。从各胴长组的N%和F%来看,鸢乌贼偏爱摄食个体相对较大的头足类和鱼类,尤其在高胴长组,头足类和鱼类的优势饵料地位更加明显。随着胴长的增加,鸢乌贼胃含物中小个体饵料类群的N%和F%普遍降低,而大个体饵料类群的N%和F%普遍升高。
Abstract:Based on the samples collected by the light falling-net fishing vessels in the South China Sea during September and October, 2012, we examined the diet composition, feeding intensity and ontogenetic feeding habit shifts of Symplectoteuthis oualaniensis by traditional stomach contents analysis and molecular identification methods. The results show that the diets of S.oualaniensis in the South China Sea were cephalopods, fishes (mainly Myctophidae) and crustaceans, and cannibalism was also detected obviously. Cephalopods and fishes were dominant prey groups; the relative importance index percent (%IRI), individual percentage (N%), quality percentage(W%) and occurrence percentage(F%) of cephalopods were 54.38, 28.73, 58.39 and 75.60, respectively, and those of fishes were 41.39, 42.54, 36.17 and 63.69, respectively. The feeding grades of S.oualaniensis were mainly Grade 3 and Grade 4;the average stomach fullness index was 3.11% and the average percentage of empty stomach was 2.89%. In terms of N% and F% among the mantle length groups, the bigger individuals such as cephalopods and fishes were the main prey groups for S.oualaniensis, especially in the long mantle length groups, the dominant position of cephalopods and fishes was more obvious. With increasing mantle length of S.oualaniensis, N% and F% of the smaller prey groups and of the bigger prey groups in stomach content reduced and increased generally, respectively.
-
黄鳍金枪鱼 (Thunnus albacares) 隶属于鲭科、金枪鱼属,是一种喜集群的大洋性洄游鱼类,因背鳍、臀鳍及小鳍同为橘黄色,又名黄鳍鲔[1]。广泛分布于太平洋、大西洋、印度洋的热带和亚热带海域[2]。众所周知,金枪鱼具有生长速度快、粗蛋白和不饱和脂肪酸含量高的特点,深受消费者喜爱[3]。金枪鱼的丰度还可以支持珊瑚礁、沿海渔业和海水养殖等产业的可持续开发和利用[4]。其种群是组成远洋生态系统的关键部分,也是世界上最大和最有价值的渔业之一[5]。
对水生动物来说,pH是一个重要的环境因子,其极细微的变化也会产生深远影响[6];从而引起生态系统内一系列的化学变化,影响大多数水产生物的生长、繁殖、代谢与生存[7]。当水体pH下降到一定程度,超过机体的最大调控范围,就会出现细胞内酸中毒和环境高碳酸血症[8]。二氧化碳 (CO2)排放过量引起的海洋酸化改变了海洋环境,扰乱了海洋动物机体的新陈代谢、酸碱平衡和生物体内多种酶的活性[9]。有研究表明,水体酸化会使金头鲷 (Sparus aurata) 的抗氧化能力受到抑制[10];过酸的养殖水体会显著影响脊尾白虾 (Exopalaemon carinicauda) 的非特异性免疫力[11];酸化导致了青鳉 (Oryzias melastigm) 幼鱼脂质代谢紊乱[12];严重的酸化条件与舌齿鲈 (Dicentrarchus labrax) 显著升高的最大代谢率有关[13];海洋酸化会导致金枪鱼的生物量减少[14]。
大气中过多的CO2被海水吸收后形成碳酸,增加了海水酸度,然而少有酸化胁迫对黄鳍金枪鱼幼鱼生理特性影响的报道。随着捕捞量的增加,金枪鱼的资源量急剧下降,无法满足市场需求。本研究通过测定相关酶活性,评估海水酸化胁迫对黄鳍金枪鱼幼鱼抗氧化防御系统和免疫功能的影响,为海洋生态环境和金枪鱼野生种群的保护提供参考。
1. 材料与方法
1.1 实验材料
黄鳍金枪鱼幼鱼平均体长为 (18.21±1.09) cm、平均体质量为 (354.98±149.77) g,由中国水产科学研究院南海水产研究所热带水产研究开发中心提供。实验用水为砂滤后的自然海水,水温 (22.5±1.0) ℃,溶解氧质量浓度 (7.5±0.5) mg·L−1,盐度 (33.00±0.80)‰,氨氮质量浓度 <0.05 mg·L−1,亚硝酸盐质量浓度 <0.05 mg·L−1。
1.2 实验设计
正式实验开始前,先将金枪鱼幼鱼在驯养池中暂养7 d,期间投喂冰鲜杂鱼。随机挑选72尾行为活泼、体表健康的幼鱼,在直径和水深均为1.5 m的圆形钢化桶中进行48 h的酸化胁迫实验,暂养期间与正式实验期间水质保持一致。实验共设置4组,以自然海水pH 8.1为对照组,pH 7.6、7.1、6.6为实验组,每组设3个重复,每个重复6尾鱼。实验期间金枪鱼幼鱼处于停食状态。并通过1.0 mmol·L−1的氢氧化钠 (NaOH) 溶液或1.0 mmol·L−1的盐酸 (HCl) 溶液调节海水pH。使用pH分析仪 (PH848) 测定海水酸度,每2 h矫正1次,使pH变化幅度保持在 ±0.1。实验至第48小时,统计死亡的幼鱼数量 (以鱼体侧躺、沉入水底不再游动为准),计算成活率。
1.3 样品采集与处理
实验结束后每组随机选取6尾黄鳍金枪鱼幼鱼,经丁香酚麻醉后进行解剖,解剖全程在冰盒上进行。分别采集肝、鳃、红肌、皮肤组织于2 mL无菌冻存管中,置于液氮中冷冻,之后保存在−80 ℃冰箱中用于后续免疫和代谢酶的测定。准确称取组织0.1~0.2 g,加入9倍体积的0.9% (w) 盐水,冰水浴条件下机械匀浆后,3 000 r·min−1离心10 min,取上清液测定相关酶活性和总蛋白含量。选取丙二醛 (MDA) 为抗氧化物质的标志物;超氧化物歧化酶 (SOD)、过氧化氢酶 (CAT)、谷胱甘肽过氧化物酶 (GSH-Px) 和过氧化物酶 (POD) 作为抗氧化防御系统的标志物;酸性磷酸酶 (ACP)、碱性磷酸酶 (AKP)、溶菌酶 (LZM) 作为免疫系统的标志物。上述所有检测均使用南京建成生物工程研究所试剂盒,所有指标的测定严格按照说明书进行操作。采用酶标仪 (SYNERGY-HT) 或分光光度计 (UV-1800BPC) 测定各项指标。
1.4 数据处理
采用Excel 2010软件整理数据,以“平均值±标准差 ($\overline { x}\pm s $) ”表示 (n=3)。实验结果通过SPSS 25.0软件进行单因素方差分析 (One-way ANOVA),并用邓肯检验法分析组间差异性,显著性水平α为0.05。使用Origin 2022软件作图。
2. 结果
实验至第48小时,仅pH 6.6处理组的幼鱼出现死亡,成活率为83%。各组之间无显著性差异 (P>0.05,图1)。
![]() 图 1 酸化胁迫对黄鳍金枪鱼幼鱼成活率的影响注:相同字母表示组间无显著性差异 (P>0.05)。Figure 1. Effect of acidification stress on survival rate of juvenile yellowfin tunaNote: Same letters represent no significant differences among groups (P>0.05).
图 1 酸化胁迫对黄鳍金枪鱼幼鱼成活率的影响注:相同字母表示组间无显著性差异 (P>0.05)。Figure 1. Effect of acidification stress on survival rate of juvenile yellowfin tunaNote: Same letters represent no significant differences among groups (P>0.05).2.1 酸化胁迫对黄鳍金枪鱼幼鱼抗氧化能力的影响
与对照组相比,各实验组皮肤的MDA含量均有所下降,红肌和鳃的MDA含量均有所上升。除对照组外,相同pH下肝脏中的MDA含量均显著高于其他组织 (P<0.05),且随着pH的降低呈上升趋势,在pH 6.6时含量最高 (图2)。
![]() 图 2 酸化胁迫对黄鳍金枪鱼幼鱼丙二醛含量的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 2. Effect of acidification stress on MDA of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with same pH (P<0.05).
图 2 酸化胁迫对黄鳍金枪鱼幼鱼丙二醛含量的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 2. Effect of acidification stress on MDA of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with same pH (P<0.05).随着pH的降低,皮肤中的SOD活性呈先下降再上升的趋势,肝中的SOD活性呈下降趋势,二者均在pH 8.1时达到最高。红肌中的SOD活性在pH 7.6时最高。鳃中的SOD活性在pH 7.1时达到最高,且随着pH的降低呈先上升后下降的趋势;除对照组外,相同pH下鳃中的SOD活性均显著高于其他组织 (P<0.05,图3-a)。
![]() 图 3 酸化胁迫对黄鳍金枪鱼幼鱼超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶和过氧化物酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 3. Effects of acidification stress on SOD, CAT, GSH-Px and POD activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).
图 3 酸化胁迫对黄鳍金枪鱼幼鱼超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶和过氧化物酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 3. Effects of acidification stress on SOD, CAT, GSH-Px and POD activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).随着pH的降低,皮肤中的CAT活性呈下降趋势,在pH 8.1时活性最高。鳃和红肌中的CAT活性呈先上升再下降的趋势,在pH 7.1时达到最高;除对照组外,相同pH下鳃中的CAT活性均显著高于其他组织 (P<0.05)。肝中的CAT活性变化平稳,在pH 6.6时达到最高。相同pH下,肝和鳃中的CAT活性均显著高于皮肤和红肌 (P<0.05,图3-b)。
随着pH的降低,皮肤、红肌和鳃中的GSH-Px活性变化相似,均呈先上升后下降的趋势,且均在pH 7.1时达到最高,此时皮肤中的GSH-Px活性显著高于其他组织 (P<0.05)。肝中的GSH-Px活性呈上升趋势,在pH 6.6时达到最高 (图3-c)。
随着pH的降低,皮肤中的POD活性呈下降趋势,在pH 8.1时活性最高。红肌中各实验组的POD活性均高于对照组,在pH 6.6时达到最高。鳃中的POD活性呈先上升后下降的趋势,在pH 7.1时达到最高;相同pH下鳃中的POD活性均显著高于其他组织 (P<0.05)。肝中的POD活性呈先下降再上升的趋势,在pH 8.1时活性最高。除对照组外,相同pH下红肌和鳃中的POD活性均显著高于皮肤和肝 (P<0.05,图3-d)。
2.2 酸化胁迫对黄鳍金枪鱼幼鱼免疫酶活性的影响
随着pH的降低,肝和皮肤各实验组的ACP活性与对照组相比均有所下降;相同pH下肝中的ACP活性均显著高于其他组织 (P<0.05)。红肌中的ACP活性呈先上升后下降再上升的趋势,在pH 7.6时达到最高;鳃中的ACP活性呈先上升后下降的趋势,在pH 7.1时达到最高。相同pH下鳃中的ACP活性均显著低于其他组织 (P<0.05,图4-a)。
![]() 图 4 酸化胁迫对黄鳍金枪鱼幼鱼酸性磷酸酶、碱性磷酸酶和溶菌酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 4. Effects of acidification stress on ACP, AKP and LZM activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).
图 4 酸化胁迫对黄鳍金枪鱼幼鱼酸性磷酸酶、碱性磷酸酶和溶菌酶活性的影响注:相同pH下不同字母表示不同组织间差异显著 (P<0.05)。Figure 4. Effects of acidification stress on ACP, AKP and LZM activities of juvenile yellowfin tunaNote: Different letters represent significant differences among different tissues with the same pH (P<0.05).随着pH的降低,鳃、红肌和皮肤中的AKP活性呈先下降后上升的趋势,均在pH 8.1时最高。肝中的AKP活性呈上升趋势,在pH 7.1时达到最高。相同pH下红肌中的AKP活性均显著低于其他组织 (P<0.05,图4-b)。
随着pH的降低,皮肤中的LZM活性呈先上升后下降的趋势,在pH 7.1时达到最高。鳃和红肌中的LZM活性呈上升趋势,均在pH 6.6时达到最高;相同pH下鳃中的LZM活性均显著高于其他组织 (P<0.05)。肝中的LZM活性在pH 7.1时达到最高 (图4-c)。
3. 讨论
3.1 环境胁迫对水产动物不同组织抗氧化能力的影响
MDA是脂质过氧化的重要产物[15],反映机体受氧化损伤的程度[16],其含量随着机体氧化应激水平升高而增加,含量越高表明鱼体所受的压力越大[17]。肝脏有排泄、解毒等多种功能,可清除外来有害物质和机体产生的有害废物[18];且含有大量与免疫相关的细胞,具有重要的免疫调节功能,在机体的免疫应答中发挥重要作用[19]。本研究中肝脏的MDA含量随海水酸度升高呈上升趋势,且累积含量显著高于其他组织。可能是由于金枪鱼处于应激状态,体内产生大量的活性氧自由基,脂质过氧化水平升高,机体出现了氧化损伤,与克氏原螯虾 (Procambarus clarkii)[20]的结果相似。
SOD和CAT活性的变化可在一定程度上反映出机体受环境胁迫时免疫机能的变化[21]。GSH-Px在清除过氧化氢 (H2O2)、把脂质过氧化物还原成无毒产物方面起着至关重要的作用[22],其与CAT在清除自由基方面既是互补也是竞争关系[23]。SOD、CAT和GSH-Px协同作用,是抗氧化系统的重要组成部分[24]。POD和生物应激相关,可水解H2O2,对细胞起保护作用[25]。
鳃是鱼类主要的呼吸和免疫器官,表面积大且与水体直接接触,易受有害物质伤害[26];其黏膜中含有丰富的免疫分子,在抵御外界侵袭中发挥重要作用[27]。本研究中,各处理组鳃中的SOD、CAT和POD活性显著高于其他组织,且在pH 7.1时达到最高。说明酸性胁迫下金枪鱼组织中的活性氧自由基含量升高,诱导SOD、CAT和POD的活性升高,鱼体通过激活抗氧化酶系统促进蛋白质合成,改善代谢产物,保护细胞不受损伤,从而使鱼体能够适应低pH环境,达到新的动态平衡。但水体酸度过高时,鱼体会产生过量的氧自由基 (·O2−),机体无法产生足够的SOD和CAT来清除 ·O2−,且动物机体在受到环境胁迫时,SOD和CAT可能会发生抑制的应激变化[28],因此本研究鳃中的CAT活性在pH 6.6时有所回落。与沙塘鳢 (Odontobutis potamophila)[29]、瓦氏黄颡鱼 (Pelteobagrus vachelli)[30]和方斑东风螺 (Babylonia areolata)[31]的研究结果相似。
鱼类皮肤的腺层内含有丰富的腺细胞,分泌的黏液中含有丰富的免疫因子,如溶菌酶、补体类物质等[32],构成了机体的第一道防线[33]。本研究中,皮肤的GSH-Px活性在pH 7.1时达到最高,显著高于其他组织,而CAT活性低于其他组织,可能是由于H2O2没有被CAT及时分解,诱导机体生成更多的GSH-Px来清除多余的H2O2。GSH-Px的活性升高,谷胱甘肽 (GSH) 等抗氧化物质就会消耗过大,可能导致机体受到损伤[34]。本研究中皮肤的GSH-Px活性在pH 7.1时大幅上升,推测此时鱼体无法适应过酸的海水,机体出现了损伤。与尖吻鲈 (Lates calcarifer) 幼鱼[35]和点篮子鱼 (Siganus guttatus) 幼鱼[36]的表现类似。
3.2 环境胁迫对水产动物不同组织免疫酶的影响
磷酸酶又称正磷酸单酯水解酶,可以催化各种含磷化合物的水解,根据他们的特性 (即起催化作用的最适pH),可分为ACP和AKP两类[37],前者主要起催化效果,后者主要起调理和调节的作用[38]。当水体中溶解氧含量低于正常水平时,鱼体发生应激反应,并通过提高AKP和ACP的活性来增强非特异性免疫力[39]。
本研究中,肝脏的ACP活性显著高于其他组织,且随着pH的降低呈下降趋势;但AKP活性呈上升趋势,可能因为二者在肝脏中的作用不同。分析出现上述结果的原因,可能是海水酸度过高,机体为了维持酸碱和离子平衡而采取的一种调节措施,是生物体主动防御的表现。类似的报道还有脊尾白虾[11]和栉孔扇贝 (Chlamys farreri)[21]。鳃中的ACP和AKP变化不一致,可能是此时的环境pH影响了鳃细胞参与免疫的水解酶活性。类似的结果还见于翘嘴鳜 (Siniperca chuatsi) 幼鱼[40]和缢蛏 (Sinonovacula constricta) 成贝[41]。
LZM是衡量鱼类非特异性免疫的指标之一[42],在面对外界环境胁迫时,其活性会升高[43],活性越强,机体的非特异性免疫力就越强[44]。鳃中的LZM活性显著高于其他组织,且随海水酸度的升高呈上升趋势。可能是由于海水酸度的增加使金枪鱼处于应激状态,鱼体通过增加LZM的活性以抵抗外源环境的压力,属于自身的应急保护反应。
4. 结论
本研究通过设置4组酸化梯度 (pH 8.1、7.6、7.1、6.6) 对黄鳍金枪鱼幼鱼进行48 h的环境胁迫,结果显示,酸化胁迫对黄鳍金枪鱼幼鱼的免疫功能和抗氧化防御系统影响显著。抗氧化物质在pH 6.6时达到最高;抗氧化酶的活性多数在pH 7.1时达到最高;非特异性免疫酶变化不一,不同酶的活性因组织不同而呈类似或不同的变化趋势。提示黄鳍金枪鱼幼鱼在pH大于7.1的酸化环境中有一定的调节能力,pH小于7.1时免疫系统发生紊乱。整体而言,肝脏和鳃中的酶活性高于红肌和皮肤,这表明在面对海水酸度升高时,肝脏和鳃中的抗氧化酶和免疫酶能更好地表达,二者抗氧化酶和免疫酶的变化可以代表黄鳍金枪鱼幼鱼机体免疫功能和抗氧化防御系统的变化。
-
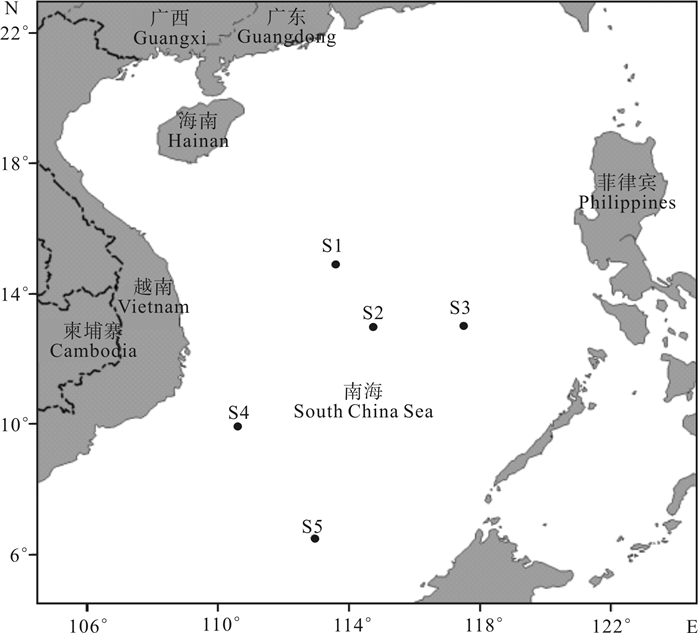
![]()
图 1 南海鸢乌贼采样站位示意图
南海九段线参见彩色宣传页的调查区域图
Figure 1. Sampling sites of S.oualaniensis in the South China Sea
For information about the nine-dash line of the South China Sea, see the survey area map on the colored leaflet.
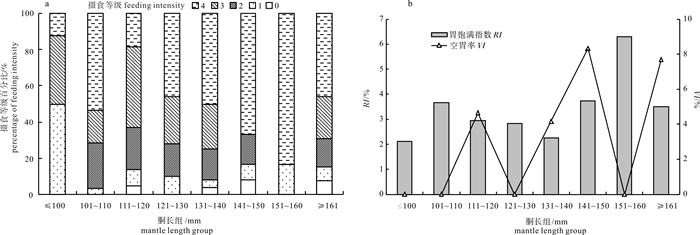
![]()
图 2 不同胴长组鸢乌贼摄食等级百分比(a)、胃饱满指数(RI)和空胃率(VI)(b)
Figure 2. Percentage of feeding intensity (a), stomach fullness index and percentage of empty stomach of S.oualaniensis (b) in different mantle length groups
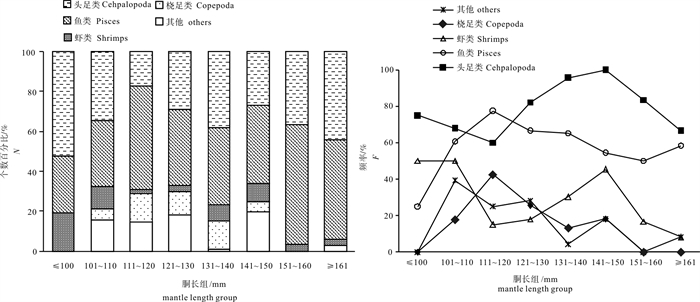
![]()
图 3 不同胴长组鸢乌贼个数百分比(N%)和出现频率(F%)
Figure 3. N% and the F% for major prey groups in diets of S.oualaniensis in different mantle length groups
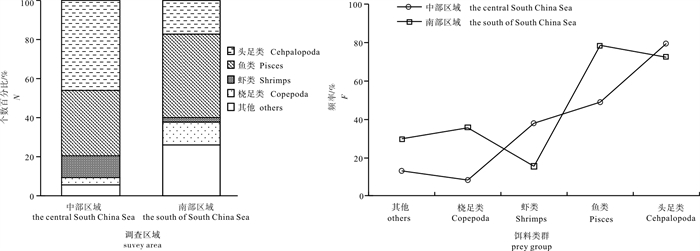
![]()
图 4 不同区域鸢乌贼个数百分比(N%)和出现频率(F%)
Figure 4. N% and the F% for major prey groups in diets of S.oualaniensis in different survey areas
表 1 鸢乌贼的采样数据
Table 1 Sample composition of S.oualaniensis
采样日期
simpling date站位
station经纬度
latitude and longitude胃
stomach数量
number空胃率/%
VI胴长范围/mm
mantle length range平均胴长/mm
average mantle length09-07 S1 113°33.82′E 实胃 34 2.86 92~219 128.8 14°53.45′N 空胃 1 119 119.0 09-17 S2 114°43′E 实胃 7 12.50 133~196 181.0 12°57′N 空胃 1 195 195.0 09-12 S3 117°27.98′E 实胃 43 2.27 103~225 125.1 13°1.73′N 空胃 1 149 149.0 10-03 S4 110°34.5′E 实胃 40 4.76 88~210 126.4 9°55.6′N 空胃 2 114~135 124.5 10-12 S5 112°56.37′E 实胃 44 0 91~154 119.6 6°31.13′N 空胃 0 - - 合计 total 实胃 168 2.89 88~225 127.0 空胃 5 114~195 142.4 总计 173 88~225 127.5  下载: 导出CSV
下载: 导出CSV
表 2 鸢乌贼的饵料组成
Table 2 Diet composition of S.oualaniensis
饵料种类
diet species个数
number个数百分比
number percentage (N%)质量百分比
weight percentage (W%)出现频率
frequency (F%)出现频率百分组成/%
frequency percentage相对重要性指数
index of relative importance IRIIRI比例
index of relative importance percentage (%IRI)甲壳动物 CRUSTACEA 174 21.45 - 51.19 25.37 - - 桡足类 COPEPODA 80 9.86 0.03 22.02 10.91 217.91 1.80 不可辨别桡足类 unidentified copepoda 32 3.95 - 10.12 5.01 - - 伯氏鹰嘴水蚤 Aetideopsis bradyi 1 0.12 - 0.60 0.29 - - 长角深角水蚤 Bathypontia longicornis 1 0.12 - 0.60 0.29 - - 短平头水蚤 Candacia curta 1 0.12 - 0.60 0.29 - - 异尾平头水蚤 C.discaudata 1 0.12 - 0.60 0.29 - - 平头水蚤 Candacia sp. 1 0.12 - 0.60 0.29 - - 奥氏胸刺水蚤 Centropages orsinii 1 0.12 - 0.60 0.29 - - 小基齿哲水蚤 Clausocalanus minor 1 0.12 - 0.60 0.29 - - 长刺大眼水蚤 Corycaeus longistylis 1 0.12 - 0.60 0.29 - - 大眼水蚤 Corycaeus sp. 1 0.12 - 0.60 0.29 - - 真哲水蚤 Eucalanus sp. 1 0.12 - 0.60 0.29 - - 真胖水蚤 Euchirella sp. 2 0.25 - 1.19 0.59 - - 异肢水蚤 Heterorhabdus sp. 3 0.37 - 1.19 0.59 - - 刺额异肢水蚤 H.spinifrons 1 0.12 - 0.60 0.29 - - 瘦新哲水蚤 Neocalanus gracillis 1 0.12 - 0.60 0.29 - - 粗新哲水蚤 N.robustior 1 0.12 - 0.60 0.29 - - 长腹剑水蚤 Oithona sp. 1 0.12 - 0.60 0.29 - - 丽隆水蚤 Oncae venusta 24 2.96 - 10.71 5.31 - - 裸拟哲水蚤 Paracalanus nudus 1 0.12 - 0.60 0.29 - - 粗拟异肢水蚤 Paraheterorhabdus robustus 1 0.12 - 0.60 0.29 - - 角水蚤 Pontella sp. 1 0.12 - 0.60 0.29 - - 双叶伪胖水蚤 Pseudochirella bilobata 1 0.12 - 0.60 0.29 - - 异尾宽水蚤 Temora discaudata 1 0.12 - 0.60 0.29 - - 不可辨别等足类 unidentified isopoda 4 0.49 - 2.38 1.18 - - 不可辨别端足类 unidentified amphipoda 1 0.12 - 0.60 0.29 - - 不可辨别虾类 unidentified shrimps 45 5.55 5.41 26.79 13.27 293.54 2.42 短尾类大眼幼虫 Megalopa larvae 42 5.18 - 14.88 7.37 - - 长尾类幼体 Macruran larvae 2 0.25 - 1.19 0.59 - - 软体动物 MOLLUSCA 59 7.27 - 11.31 5.60 - - 强卷螺 Agadina stimpsoni 1 0.12 - 0.60 0.29 - - 马蹄螔螺 Limacina trochiformis 40 4.93 - 10.71 5.31 - - 软体动物幼体 Mollusca larvae 18 2.22 - 4.17 2.06 - - 鱼类 PISCES 345 42.54 36.17 63.69 31.56 5 013.08 41.39 不可辨别鱼类 unidentified Pisces 33 4.07 - 19.64 9.73 - - 诺贝珍灯鱼 Lampanyctus nobilis 30 3.70 - 5.95 2.95 - - 七星底灯鱼 Benthosema pterotum 1 0.12 - 0.60 0.29 - - 眶暗虹灯鱼 Bolinichthys pyrsobolus 2 0.25 - 1.19 0.59 - - 尾明角灯鱼 Ceratoscopelus warmingi 3 0.37 - 1.19 0.59 - - 条带眶灯鱼 Diaphus brachycephalus 2 0.25 - 0.60 0.29 - - 天蓝眶灯鱼 D.coeruleus 3 0.37 - 1.19 0.59 - - 巴氏眶灯鱼 D.parri 1 0.12 - 0.60 0.29 - - 华丽眶灯鱼 D.perspicillatus 14 1.73 - 4.17 2.06 - - 瓦氏眶灯鱼 D.watasei 2 0.25 - 1.19 0.59 - - 真巨口鱼 Eustomias sp. 9 1.11 - 4.17 2.06 - - 珍灯鱼 Lampanyctus sp. 20 2.47 - 3.57 1.77 - - 粗鳞灯笼鱼 Myctophum asperum 4 0.49 - 1.19 0.59 - - 金焰灯笼鱼 M.aurolaternatum 1 0.12 - 0.60 0.29 - - 短颌灯笼鱼 M.brachygnathym 2 0.25 - 0.60 0.29 - - 闪光灯笼鱼 M.nitidulum 14 1.73 - 6.55 3.24 - - 钝吻灯笼鱼 M.obtusirostris 4 0.49 - 1.79 0.88 - - 粗短灯笼鱼 M.selenoides 1 0.12 - 0.60 0.29 - - 光彩标灯鱼 Symbolophorus evermanni 190 23.43 - 30.36 15.04 - - 鱼类未定种2 unidentified Pisces 2 1 0.12 - 0.60 0.29 - - 鱼类未定种4 unidentified Pisces 4 1 0.12 - 0.60 0.29 - - 鱼类未定种5 unidentified Pisces 5 3 0.37 - 1.19 0.59 - - 鱼类未定种6 unidentified Pisces 6 1 0.12 - 0.60 0.29 - - 鱼类未定种7 unidentified Pisces 7 3 0.37 - 1.19 0.59 - - 头足类 CEHPALOPODA 233 28.73 58.39 75.60 37.46 6 585.85 54.38 不可辨别头足类 unidentified Cehpalopoda 232 28.61 - 75.00 37.17 - - 安达曼钩腕乌贼 Abralia andamanica 1 0.12 - 0.60 0.29 - - 注:-. 缺少质量的饵料生物无法计算其相关指标
Note:-. The diets lacking of weight can not be calculated their relevant index.
下载: 导出CSV
-
[1] 陈新军, 刘必林, 王尧耕. 世界头足类[M]. 北京: 海洋出版社, 2009: 312-313. [2] 张鹏, 晏磊, 杨炳忠, 等. 春季南沙海域鸢乌贼种群结构特征的研究[J]. 南方水产科学, 2015, 11(5): 11-19. doi: 10.3969/j.issn.2095-0780.2015.05.002 [3] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. doi: 10.3969/j.issn.1673-2227.2010.01.012 [4] 陈新军, 钱卫国. 印度洋西北部海域鸢乌贼资源密度分布的初步分析[J]. 上海水产大学学报, 2004, 13(3): 218-223. [5] 陈新军, 钱卫国, 田思泉. 阿拉伯海北部公海海域鸢乌贼资源密度及其分布[J]. 海洋科学进展, 2006, 24(3): 360-364. doi: 10.3969/j.issn.1671-6647.2006.03.014 [6] 陈新军, 叶旭昌. 印度洋西北部海域鸢乌贼渔场与海洋环境因子关系的初步分析[J]. 上海水产大学学报, 2005, 14(1): 55-60. [7] 陈新军, 刘金立. 利用形态学方法分析印度洋西北部海域鸢乌贼种群结构[J]. 上海水产大学学报, 2007, 16(2): 174-179. [8] SIRIRAKSOPHON S, SUKRAMONGKOL N, NAKAMURA Y. Exploration of oceanic squid, Sthenoteuthis oualaniensis resources in the South China Sea, Vietnamese waters[C]//Proceedings of the SEAFDEC Seminar on Fishery Resources in the South China Sea, Area Ⅲ: Western Philippines. Bangkok: Southeast Asian Fisheries Development Center, 1999: 181-197.
[9] BASIR S. Biological feature of an oceanic squid, Sthenoteuthis oualaniensis in the South China Sea, Area Ⅲ: Western Philippines [C]//Proceedings of the SEAFDEC Seminar on Fishery Resources in the South China Sea, Area Ⅲ: Western Philippines. Bangkok: Southeast Asian Fisheries Development Center, 2000: 135-147.
[10] PARRY M P. The trophic ecology of two ommastrephid squid species, Ommastrephes bartramii and Sthenoteuthis oualaniensis, in the North Pacific sub-tropical gyre[D]. Manoa: University of Hawaii, 2003: 1-287.
[11] MÉNARD F, POTIER M, ROMANOV E, et al. New information from predator diets on the importance of two Ommastrephidae Sthenoteuthis oualaniensis in the Indian Ocean and Hyaloteuthis pelagica in the Atlantic Ocean[J]. Role Squid Open Ocean Ecosys, 2007, 24: 49-52.
[12] PARRY M. Feeding behavior of two ommastrephid squids Ommastrephes bartramii and Sthenoteuthis oualaniensis off Hawaii[J]. Mar Ecol Prog Ser, 2006, 318: 229-235. doi: 10.3354/meps318229
[13] ZHANG J, CHEN Z Z, GUO B C, et al. Hydroacoustic studies on the commercially important squid Sthenoteuthis oualaniensis in the South China Sea[J]. Fish Res, 2015, 169: 45-51. doi: 10.1016/j.fishres.2015.05.003
[14] FANG Z, XU L L, CHEN X J, et al. Beak growth pattern of purpleback lying squid Sthenoteuthis oualaniensis in the eastern tropical Pacific equatorial waters[J]. Fish Sci, 2015, 81(3): 443-452. doi: 10.1007/s12562-015-0857-8
[15] 颜云榕, 冯波, 卢伙胜, 等. 南沙群岛北部海域鸢乌贼夏季渔业生物学研究[J]. 海洋与湖沼, 2012, 43(6): 1177-1186. [16] 张宇美, 颜云榕, 卢伙胜, 等. 西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究[J]. 广东海洋大学学报, 2013, 33(3): 56-64. [17] 纪东平, 卞晓东, 宋娜, 等. 荣成俚岛大泷六线鱼摄食生态研究[J]. 水产学报, 2014, 38(9): 1399-1409. [18] 纪东平, 卞晓东, 宋娜, 等. 荣成俚岛斑头鱼摄食生态研究[J]. 中国水产科学, 2015, 22(1): 88-98. [19] REED J, TOLLIT D. Microsatellite markers allow species discrimination from faecal and recognizable manatees on the Atlantic coast of Florida[M]//O′SHEA T J, ACKERMAN B B, PERCIVAL H F. Population biology of the florida Manatee. Washington, D.C. : Information and Technology Report 1, 1995: 289.
[20] DEAGLE B E, TOLLIT D J, JARMAN S N, et al. Molecular scatology as a tool to study diet: analysis of prey DNA in scats from captive Steller sea lions[J]. Mol Ecol, 2005, 14(6): 1831-1842.
[21] 洪巧巧. 长江口中国花鲈的食性及分子生物学在食性分析上的应用[D]. 上海: 华东理工大学, 2012: 1-65. [22] 陈大纲. 渔业资源生物学[M]. 北京: 中国农业出版社, 1995: 94. [23] 颜云榕, 杨厚超, 卢伙胜, 等. 北部湾宝刀鱼的摄食生态[J]. 生态学报, 2011, 31(3): 654-665. [24] 龚玉艳, 陈作志, 张俊, 等. 南海北部陆坡海域秋季金鼻眶灯鱼的摄食习性[J]. 南方水产科学, 2015, 11(5): 90-99. doi: 10.3969/j.issn.2095-0780.2015.05.011 [25] CORTÉS E J. A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes[J]. Can J Fish Aquat Sci, 1997, 54(3): 726-738.
[26] 叶旭昌, 陈新军. 印度洋西北海域鸢乌贼生物学特性初步研究[J]. 上海水产大学学报, 2004, 13(4): 316-322. [27] 张宇美. 基于碳氮稳定同位素的南海鸢乌贼摄食生态与营养级研究[D]. 湛江: 广东海洋大学, 2014: 1-63. [28] OKUTANI T, TUNG I. Reviews of biology of commercially important squids in Japanese and adjacent waters. I. Symplectoteuthis oualaniensis(Lesson)[J]. Veliger, 1978, 21(1): 87-94.
[29] 钱卫国, 陈新军, 刘必林, 等. 印度洋西北海域秋季鸢乌贼渔场分布与浮游动物的关系[J]. 海洋渔业, 2006, 28(4): 265-271. [30] 薛莹, 金显仕, 张波, 等. 黄海中部小黄鱼摄食习性的体长变化与昼夜变化[J]. 中国水产科学, 2004, 11(5): 420-425. [31] IBÁÑEZ C M, KEYL F. Cannibalism in cephalopods[J]. Rev Fish Biol Fish, 2010, 20(1): 123-136.
[32] SHCHETINNIKOV A. Feeding spectrum of squid Sthenoteuthis oualaniensis(Oegopsida)in the eastern Pacific[J]. J Mar Biol Assoc UK, 1992, 72(4): 849-860.
-
期刊类型引用(11)
1. 肖佳程,赵一方,石耀华,顾志峰. 养殖密度对宝蓝鹦鹉鱼生长、生理与繁殖的影响. 热带生物学报. 2025(02): 304-311 .  百度学术
百度学术
2. 汪海燕,张永明,靳桂双,赵璐. 植物多糖的生物学功能及其在水产养殖中的应用. 饲料研究. 2024(09): 173-177 . 百度学术
3. 高建伟,汪东升,邵锋锋. 加减大柴胡汤对胆总管结石患者湿热证候及肝功能的影响. 中医药临床杂志. 2024(06): 1144-1148 . 百度学术
4. 夏伦斌,马龙龙,乔德亮,何燕飞,蒋平. 三角帆蚌多糖对肉仔鸡生长性能、抗氧化及免疫功能的影响. 浙江农业学报. 2023(03): 547-555 . 百度学术
5. 田瑜,揭育鵾,曾祥兵,乐燕,刘广鑫,程长洪,马红玲,郭志勋. 密度对拟穴青蟹抗氧化、非特异性免疫能力的影响. 南方水产科学. 2023(03): 60-67 . 本站查看
6. 刘清豪,张丹,陈卓,韦婉婷,王城城,金言,修彦凤. HPLC法同时测定炮制前后鸡内金中6种黄酮类成分. 中成药. 2023(08): 2618-2622 . 百度学术
7. 李成辉,董宏标,郑晓婷,桂福坤,曾祥兵,明俊超,陈飞,陈健,张家松. 春砂仁精油对尼罗罗非鱼幼鱼生长、消化、抗氧化能力和血清生化指标的影响. 南方水产科学. 2023(06): 51-59 . 本站查看
8. 黄健彬,迟艳,周传朋,黄小林,黄忠,虞为,荀鹏伟,吴杨,张宇,林黑着. 褐藻寡糖对卵形鲳鲹幼鱼生长性能、抗氧化能力和免疫功能的影响. 南方水产科学. 2022(03): 118-128 . 本站查看
9. 贾慧凝,侍苗苗,卞永乐,侍崇敬,刘恒蔚,宋学宏,秦粉菊. 纳米硒对低氧胁迫下中华绒螯蟹免疫保护和抗氧化能力的影响. 南方水产科学. 2022(06): 100-109 . 本站查看
10. 张慧,董宏标,孙彩云,陈健,黄聪灵,李勇,段亚飞,张家松. 饲料中添加月桂酸单甘酯对尖吻鲈脂质代谢与肝脏功能的影响. 海洋渔业. 2022(06): 736-746 . 百度学术
11. 范思远,张海瑜,周永雨,尹秀雯,刘紫嫣,魏峰. 香菇菌丝体多糖对肉鸡生长性能、免疫功能及抗氧化性能的影响. 饲料研究. 2021(23): 41-44 . 百度学术
其他类型引用(6)




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 17
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
