
Effect of nylon screen mesh as artificial substrate on variation of bacterial community in shrimp culture water
-
摘要:
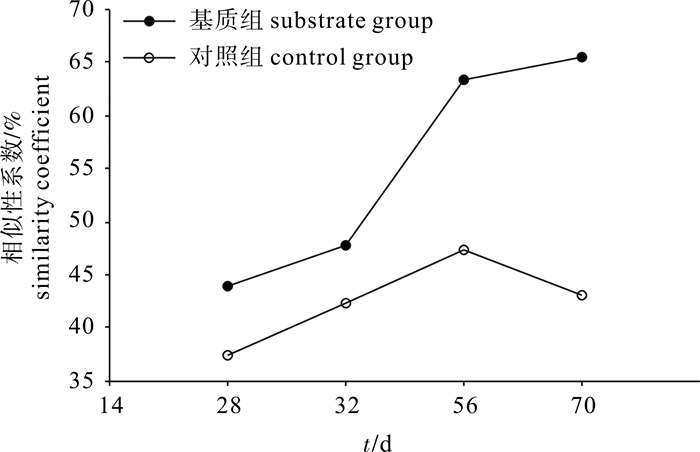
采用基于16S rDNA的PCR-DGGE(变性梯度凝胶电泳)技术,比较分析了添加尼龙筛绢网基质对凡纳滨对虾(Litopenaeus vannamei)精养系统中水体细菌群落结构演替的影响。结果表明:1)基质组水体菌群结构丰富度稍低于对照组;2)基质组水体菌群结构相似性始终高于对照组;随着时间的推移,基质组的相似性系数不断升高,而对照组出现了先升高后下降的波动;3)对照组水体养殖前期优势菌主要为变形菌门和未培养细菌,在养殖后期则以厚壁菌门芽孢杆菌属(Bacillus spp.)细菌为主,基质组水体菌群则在整个养殖周期中均以厚壁菌门芽孢杆菌属细菌和未培养细菌为主。该研究结果可为进一步阐明人工基质养殖系统中微生物的生态调控功能和作用机制提供参考。
Abstract:The bacterial community composition in the substrate-biofilm shrimp culture system was monitored and investigated by pcr-denaturing gradient gel electrophoresis (DGGE). Results show that: 1) The water flora structure of substrate group was less diversified than that of the control. 2) The similarity coefficient of substrate group increased with time, while that of the control increased first then decreased. 3) Uncultured Firmicutes (Bacillus spp.), the dominant bacteria at the beginning of culture in the control, were uncultured Proteobacteria and uncultured bacterium, and the dominant bacteria at the end of culture was uncultured Firmicutes (Bacillus spp.). Uncultured Firmicutes (Bacillus spp.) and uncultured bacterium were the dominant bacteria throughout the culturing period. The results provide references for clarifying the function and mechanism of ecological control of microbes in artificial substrate system.
-
Keywords:
- shrimp /
- artificial substrate /
- bacterial community /
- DGGE
-
南澳岛位于116°53′E~117°19′E,23°11′N~23°32′N,地理位置优越,海洋生物资源丰富。沿岸海域往往是生态系统中的敏感区域,其资源环境受诸多因素的影响。近年来,由于南澳岛近岸资源的不合理开发利用,生态环境已遭受一定程度的破坏,渔业资源明显衰退。
准确高效的渔业资源评估方法是制定科学长效的生态渔业管理机制的先决条件。传统的渔业资源评估主要依托于网捕采样调查,而任何网具对于捕捞对象都具有一定选择性,并且网捕采样通常会对调查水域渔业资源及鱼类栖息环境造成一定的破坏,其应用范围通常受水域生态环境和海底地形等条件的约束[1-2]。近年来,水下记录、摄影等技术成为获取鱼类群落信息,进行鱼类资源生态监管常用的一类方法。由于此类方法易受光照、悬浮颗粒等水域环境条件的限制,导致其评估结果可信度降低,应用范围受限[3-4]。声学评估作为一种后起的生态渔业评估方法正不断推广应用并取得了较好的成效;与传统渔业调查法相比,该方法便捷、高效、采样面积大、对渔业资源和水域生境破坏力度小,更能适应当前生态渔业发展的需求,应用前景十分广阔[5-8]。声学方法自20世纪80年代引入中国,并逐渐成为中国海洋生物资源调查研究的重要手段之一[9-10]。2007—2009年,李娜娜等[11]采用水声学方法对大亚湾杨梅坑人工鱼礁水域资源声量进行了评估;张俊等[12]针对南沙南部陆架海域渔业资源于2013年进行了4个航次的声学调查,分析了鱼类资源量空间分布特征和季节变化趋势;2013年秋季,曾雷等[13]采用声学的方法评估了百色水库渔业资源状况,并分析了鱼类昼夜行为特征。声学评估法与直接捕捞法、水下摄影等方法的有机结合被认为是未来渔业资源评估与生态监测的最佳选择之一[2]。
本研究于2014年秋季和2015年春季,综合采用声学探测和拖网调查对南澳岛部分海域渔业资源进行探查,了解不同季节该海域渔业资源动态、结构组成、空间分布等特征,并分析探讨影响渔业资源评估结果的主要因素,以期为该海域渔业资源的合理开发利用及管理策略的制定提供科学依据。
1. 材料与方法
1.1 调查海域概况与调查设置
研究海域地理空间位置见图1,大致范围为117.021 9°E~117.427 8°E,23.274 3°N~23.635 8°N。累积进行了3个航次的声学调查,调查时间分别为2015年4月25日(春季)、2014年9月10日(秋季)和2014年12月14日(冬季)。由于设备故障,冬季航次声学调查结果不能用作渔业资源评估。本文基于2014年秋季与2015年春季航次声学探测数据对南澳岛海域渔业资源进行评估分析。南澳岛海域秋季声学探测区域水深范围为1.84~38.4 m,平均水深为15.61 m;春季声学走航区域水深范围为2.33~40.2 m,平均水深为24.59 m。
![]() 图 1 南澳岛海域声学调查区域与拖网站位分布图A3~A14. 秋季拖网位点;S3~S14. 春季拖网位点Figure 1. Survey area of acoustic assessment and distribution of trawl sampling sites in Nan'ao Island areaA3–A14. trawl sampling sites in autumn; S3–S14. trawl sampling sites in spring
图 1 南澳岛海域声学调查区域与拖网站位分布图A3~A14. 秋季拖网位点;S3~S14. 春季拖网位点Figure 1. Survey area of acoustic assessment and distribution of trawl sampling sites in Nan'ao Island areaA3–A14. trawl sampling sites in autumn; S3–S14. trawl sampling sites in spring1.2 声学数据采集
声学数据采集借助挪威Simrad EY60分裂波束科学鱼探仪,工作频率分别为120 kHz (2014年秋季)和70 kHz (2015年春季)。实时经纬度信息通过GPS (Gamin GPSCSx,美国)与声学数据同步采集,各次调查科学鱼探仪的主要技术参数见表1。由于Simrad EY60声学评估硬件系统缺乏长期的稳定性,且换能器性能受不同环境介质的影响[14],故各季度调查前按照国际通用方法对科学鱼探仪系统的收发增益系数进行现场校正[15]。为防震减噪,换能器通过导流罩内固定于船体右舷外侧且不与船体接触,吃水0.8 m,走航速度约5~7 kn。通常声学调查航线依据调查区域的特征设计成平行式或“之”字式,受调查海域自然地理状况及其他人为因素的限制,声学调查航迹并不规则(图1)。参照Aglen[16]覆盖率计算公式
$D{\rm{ = }}L/\sqrt A $ ,式中L为声学航程,A为调查海域面积。2次调查声学评估海域覆盖率D分别为7.34 (2014年秋季)和8.07 (2015年春季)。表 1 EY60科学鱼探仪主要技术参数设定Table 1. Setting of main technical parameters of scientific echosounder EY60技术参数
technical parameter2014年秋季
autumn 20142015年春季
spring 2015换能器频率/kHz transducer frequency 120 70 发射功率/W transmitting power 200 300 脉冲宽度/μs pulse duration 128 512 等效波束角/dB equivalent beam angle –21 –21 换能器增益/dB transducer gain 26.8 25.69 横向波束宽度/(°) athwardship 3 dB beam width 7 6.53 纵向波束宽度/(°) alongship 3 dB beam width 7 6.43 吸收系数/dB·km–1 absorption coefficient 45.3 18.99 声速/m·s–1 sound speed 1 531.82 1 536.75 1.3 生物学数据获取
调查海域渔业资源生物学信息的获取主要通过底拖网进行,用以辅助渔业资源声学回波映像的识别与积分分配。共设置8个站位(图1),底拖网信息见表2。各站位渔获样品均进行现场分类并计数取样,数量少于50尾的物种全部取样;若单一物种渔获数量大于50尾,则根据其大小组成特点按比例随机取样,并测量记录每种渔获的长度(体长/叉长/胴长)和体质量,体长精确至1 cm,体质量精确至1 g。
表 2 主要生物学取样设置Table 2. Main setting of biological sampling主要参数
main parameter2014年秋季
autumn 20142015年春季
spring 2015采样类型 sample type 底拖网 底拖网 发动机功率/kW engine power 280 280 平均拖网速度/km·h–1 average trawling speed 5.56 4.93 平均采样时长/h average trawling time 0.55 0.46 网口宽度/m net width 6 4 网口高度/m net height 2 2 网目/cm net size 3×3 3×3 1.4 数据处理与分析
声学数据的处理主要借助专业分析软件Echoview 6.1 (Myriax),根据回波映像特征,剔除明显的环境噪声数据。利用海底检测功能板块生成探测区域海底轮廓并进行手动编辑。重新设置积分起始水层为1.5 m以屏蔽海表航行噪声干扰,积分终止水层设置为海底之上0.5 m以排除海底回波信号干扰(图2)。参照多种类海洋渔业资源声学评估方法[5,17],基本积分航程单元(elementary distance sampling unit,EDSU,n mile)设置为0.5 n mile,以底拖网采样的渔获物组成信息作为回波积分分配的主要依据。通过阈值响应(threshold response)模块分析Sv阈值的变化对回波面积积分(NASC)的影响,从而大致判断目标群体声学回波的Sv可信阈值区间。目标群体与非目标群体之间Sv并无明显界线,一般为NASC随Sv加速变化的拐点[17]。Sv积分阈值设置为–70 dB,以减小非目标信号对回波积分的影响。同时根据单体回波映像,利用Echoview软件中单体检测和轨迹追踪模块[18],进而获得回波单体大小、频率组成与垂直空间分布特征。
1.5 鱼类资源密度评估方法
1.5.1 回波积分法
1) 在不考虑渔获种类组成及体长结构分布的情况下,根据目标强度现场测定法,各基本积分航程单元内渔业资源数量密度(number density,尾·km–2)计算式为
$$ \quad\quad\quad\quad\quad\quad\quad\overline \rho = \frac{{\rm NASC}}{{4\pi \cdot {\sigma _{bs}} \cdot {{1.852}^2}}} $$ (1) 式中σbs为后向散射截面(backscattering cross-section,m2),它与目标强度(target strength,TS)的关系为
$$ \quad\quad\quad\quad\quad\quad\quad TS = 10\log {\sigma _{bs}} $$ (2) 2) 参照多种类海洋渔业资源声学评估方法,以拖网采样的渔获物信息作为积分值分配的主要依据,分析区域内第i物种的数量密度
${\rho _{i,a}}$ (number density,尾·km–2)和资源量密度${\rho _{i,b}}$ (biomass density,t·km–2)分别为$$ \quad\quad\quad\quad\quad\quad\quad {\rho _{i,a}} = {c_i}\frac{{\rm NASC}}{{4\pi \overline \sigma \cdot {{1.852}^2}}} $$ (3) $$ \quad\quad\quad\quad\quad\quad\quad {\rho _{i,b}} = {\rho _{i,a}}\overline {{w_i}} {10^{ - 6}} $$ (4) 式中ci (%)为分析海域内第i物种渔获物数量百分比,
$\overline \sigma $ 为分析海域内所有声学评估种类的平均后向散射截面,$\overline {{w_i}} $ 为第i物种的平均体质量(body mass,g),且有$$ \quad\quad\quad\quad\quad\quad\quad \overline \sigma = \sum\limits_{i = 1}^n {{c_i}{{10}^{\frac{{T{S_i}}}{{10}}}}} $$ (5) $$ \quad\quad\quad\quad\quad\quad\quad T{S_i} = 20\log {L_i} + {b_{20,i}} $$ (6) 式中TSi为第i物种的目标强度,n为参与声学评估的物种数,Li为第i物种平均体长(body length,cm),
${b_{20,i}}$ 为第i物种的参考目标强度。1.5.2 扫海面积法
游泳生物资源量密度B (kg·km–2) 根据扫海面积法估算,公式为
$$ \quad\quad\quad\quad\quad\quad\quad B = Y/A(1-E) $$ (7) 式中Y为平均渔获率 (kg·h–1);B为资源量密度 (kg·km–2);A为每小时扫海面积(km2·h–1);E为逃逸率,取0.5。
2. 结果
2.1 渔获组成特征
2014年秋季,捕获游泳生物和底栖无脊椎动物共73种,其中鱼类40种,头足类5种,虾类6种,蟹类16种,虾蛄类6种,总渔获数量为2 598尾,总渔获量为51.19 kg。2015年春季共捕获游泳生物和底栖无脊椎动物共54种,其中鱼类26种,头足类3种,虾类6种,虾蛄类4种,蟹类15种,总渔获数量为6 251尾,总渔获量为28.33 kg。
为排除海底回波信号干扰,历次调查海底之上0.5 m内均被视为声学探测的盲区,故底栖的鲆鲽类、鰕虎鱼类、蛸类、虾蟹类等非常贴底的生物均不参与声学评估。
根据生物学拖网采样结果,声学评估种类中参与积分分配的种类主要为相对重要性指数(IRI)大于100的常见种和优势种[19]。不同季节,声学评估鱼类组成存在较大差异,2014年秋季参与声学评估的种类主要有39种;2015年春季参与评估的种类主要有18种(表3)。
表 3 声学评估优势种类生物学信息Table 3. Biological information of dominant species in acoustic assessment季节(年份)
season (year)物种
species数量百分比/%
quantity percentage生物量百分比/%
biomass percentage相对重要性指数
IRI体长范围/mm
body length range体质量范围/g
body mass range秋季 (2014)
autumn短尾大眼鲷
Priacanthus macracanthus45.3 34.8 6 678 72~88 8~21 丝背细鳞鲀
Stephanolepis cirrhifer12.0 20.8 2 187 68~107 6~55 丽叶鲹
Caranx kalla9.3 6.5 1 322 60~135 4~85 杜氏枪乌贼
Loligo duvaucelii4.9 6.5 1 134 50~123* 6~58 鹿斑鲾
Secutor ruconius7.4 2.3 647 27~70 2~12 二长棘鲷
Parargyrops edita3.5 9.0 624 95~108 32~56 斑鳍白姑鱼
Argyrosomus pawak6.2 2.2 280 54~105 5.3~30 黄斑篮子鱼
Siganus canaliculatus0.6 0.9 129 74~126 5~74 春季 (2015)
spring鹿斑鲾
Secutor ruconius86.9 68.0 11 613 36~63 1.6~10 二长棘鲷
Parargyrops edita5.9 5.4 845 35~62 1.7~58 杜氏枪乌贼
Loligo duvaucelii1.0 6.9 689 42~182* 4.9~140 竹䇲鱼
Trachurus japonicus3.2 4.1 458 60~90 2.4~11 刺鲳
Psenopsis anomala0.5 2.6 229 65~110 8.5~45 短吻鲾
Leiognathus brevirostris1.1 3.6 179 58~94 4.2~24 皮氏叫姑鱼
Johnius belangerii0.3 2.6 145 92~160 14~75 沙带鱼
Lepturacanthus savala0.5 1.8 114 66~150 3.6~45 注:*. 胴长
Note: *. mantle length参与声学评估种类渔业资源密度扫海面积法计算结果如图3所示:2014年秋季平均数量密度为63 246尾·km–2,生物量密度为1 075.98 kg·km–2;2015年春季平均数量密度为221 984尾·km–2,生物量密度为949.74 kg·km–2。
![]() 图 3 调查海域各拖网位点参与声学评估种类数量与生物量密度Figure 3. Quantity and biomass density of acoustic assessed species in each trawling site of survey area
图 3 调查海域各拖网位点参与声学评估种类数量与生物量密度Figure 3. Quantity and biomass density of acoustic assessed species in each trawling site of survey area2.2 声学评估种类资源密度
1) 假定调查海域渔获种类组成单一,根据目标强度现场测定法,2014年秋季和2015年春季渔业资源数量密度分别为(64 542±132 583)尾·n mile–2和(25 472±45 814)尾·n mile–2;生物量密度分别为(1 053.32±2 163.75) kg·n mile–2和(102.4±184.17) kg·n mile–2。
2) 依据多种类渔业资源积分分配原则,调查海域2014年秋季和2015年春季参与声学积分分配的种类,其平均数量密度分别为(26 806±39 968)尾·n mile–2和(34 416±384 97)尾·n mile–2;平均生物量密度分别为(403.68±601.9) kg·n mile–2和(124.5±139.26) kg·n mile–2。
在假定种类组成单一和混合2种不同条件下,单因素方差分析结果见图4。2015年春季渔业资源数量密度(P=0.113>0.05)与生物量密度(P=0.310>0.05)均未表现出显著性差异。而2014年秋季资源数量密度(P=0.007<0.05)和生物量密度(P=0.004<0.05)在2种不同条件下均表现出显著性差异。
![]() 图 4 种类单一和混合2种不同条件下调查海域渔业资源声学评估误差线. 地理空间差异;**. α=0.05水平下差异显著Figure 4. Acoustic assessment of fishery resources in survey area in single or multi-species assumptionsError bar. geographical spatial difference; **. significant difference at α=0.05 level
图 4 种类单一和混合2种不同条件下调查海域渔业资源声学评估误差线. 地理空间差异;**. α=0.05水平下差异显著Figure 4. Acoustic assessment of fishery resources in survey area in single or multi-species assumptionsError bar. geographical spatial difference; **. significant difference at α=0.05 level根据多种类渔业资源声学评估积分分配原则,调查海域各季度不同种类数量密度与生物量密度组成情况见表4。2014年秋季调查海域优势种类主要为短尾大眼鲷(Priacanthus macracanthus),其数量密度和生物量密度分别为12 142.92 尾·n mile–2和150.22 kg·n mile–2。2015年春季调查海域优势种类主要为鹿斑鲾(Secutor ruconius),其数量密度和生物量密度分别为27 791.47尾·n mile–2和87.42 kg·n mile–2。整体上调查海域渔业资源较为贫乏,且个体偏小,应多予保护。
表 4 调查海域声学评估种类丰度与生物量密度Table 4. Abundance and biomass density of acoustic assessed species in survey area季节(年份)
season (year)种类
speciesb20/dB 丰度密度/尾·n mile–2
abundance density生物量密度/kg·n mile–2
biomass density秋季 (2014)
autumn短尾大眼鲷 Priacanthus macracanthus –68.0 12 142.92 150.22 丝背细鳞鲀 Stephanolepis cirrhifer –76.0 3 216.67 87.92 丽叶鲹 Caranx kalla –72.5 2 492.92 27.80 鹿斑鲾 Secutor ruconius –72.5 1 983.61 9.67 斑鳍白姑鱼 Argyrosomus pawak –68.0 1 661.95 9.46 杜氏枪乌贼 Loligo duvaucelii –80.0 1 313.47 28.32 二长棘鲷 Parargyrops edita –68.0 938.19 39.55 黄斑篮子鱼 Siganus canaliculatus –72.9 160.83 3.50 春季 (2015)
spring鹿斑鲾 Secutor ruconius –72.5 27 791.47 87.42 二长棘鲷 Parargyrops edita –68.0 1 886.88 6.94 竹䇲鱼 Trachurus japonicus –72.5 1 023.39 5.37 杜氏枪乌贼 Loligo duvaucelii –80.0 319.81 9.62 短吻鲾 Leiognathus brevirostris –72.5 351.79 4.58 刺鲳 Psenopsis anomala –80.0 159.90 3.53 沙带鱼 Lepturacanthus savala –66.1 159.90 2.55 皮氏叫姑鱼 Johnius belangerii –68.0 95.94 3.66 注:b20值来自文献记载
Note: The b20 values come from documentary records.调查海域渔业资源数量在地理空间上的分布见图5,其结果表明:1)调查海域渔业资源基本处于较贫乏水平。2)不同季节,声学评估种类存在较大差异,仅鲾类在2个季度调查中均占有较高的比重,枪乌贼类为2个季度常见种。3)不同季节,渔业资源在空间尺度上分布极不均匀,且无明显的空间梯度变化趋势,仅在临近南澳岛东北部(红色标记区域)存在小范围的渔业资源密集区。
2.3 回波单体TS频率组成与垂直分布
据单回波检测结果(图6),以2 dB为分组单元,调查海域2014年秋季回波单体TS变化区间为–64~–33 dB,其中–54 dB以下个体累积数量百分比高达90.37%。此外,根据回波单体在垂直空间上的分布特征,以15 m水深为界可将调查水域大致分为2~15 m (中上层)和15~35 m (中下层)两个水层。中下层水域以小型个体为主,回波单体TS基本在–54 dB以下;中上层个体大小组成相对均匀。
![]() 图 6 调查海域回波单体频率组成与垂直分布Figure 6. Frequency and vertical distribution of single echo detection in survey area
图 6 调查海域回波单体频率组成与垂直分布Figure 6. Frequency and vertical distribution of single echo detection in survey area2015年春季调查海域回波单体TS变化范围为–66~–33 dB,各分组单元TS频率分布大致呈逐级下降的变化趋势。在垂直空间上,以15 m水深为界可将调查海域大致分为2~15 m (中上层)和15~35 m (中下层) 2个水层。不同水层,回波单体TS大小组成表现出相似的特征,随回波单体TS的增大其所占数量百分比逐渐下降。
3. 讨论
3.1 渔业资源时空分布特征
各站位拖网渔获分析结果显示,南澳岛海域2014年秋季和2015年春季参与声学评估种类个体均质量分别为16.32 g·尾–1和4.02 g·尾–1。不同季节,渔业资源声学评估呈现出相似的特征,秋季鱼类个体相对较大。此外,南澳岛海域2次调查回波单体TS频率组成亦表现出明显的季节差异。较2014年秋季,2015年春季调查海域回波单体TS变化幅度更大,且小型个体所占比重更高。张俊等[5]基于南海拓林湾渔业资源研究指出,产卵引起的幼鱼增加及休渔结束后对大个体鱼类的大规模捕捞是导致鱼类个体均质量季节差异的主要原因。连玉喜等[20]研究显示,春季是大多数鱼类的繁殖期,通过增殖,水体中仔稚鱼密度大大增加,经过夏季和秋季的生长,鱼类平均个体大小明显大于春季。上述研究结果表明,调查海域鱼类资源的季节动态规律受鱼类繁殖、生长、死亡等因素的共同调节,结果符合生态系统中种群动态理论与能量传递规律[21-24]。
基于扫海面积法,不同站位渔业资源密度在水平空间上存在极大差异,2次调查均有极大异常值出现。结果可能与特定鱼类生理生活习性相关,鹿斑鲾在S14站位及短尾大眼鲷在A3站位的大规模出现,对调查海域不同季节渔业资源水平空间分布差异产生了较大影响。根据回波单体在垂直方向上的大小组成分析可知,以15 m水深为分水岭,秋季中小型个体有明显向中下层水域迁移的趋势。相关研究表明,鱼类在时空尺度上的分布受气候、水文、水质、饵料及人类活动等诸多因素的影响[25-28]。由于缺乏相应的理化环境数据,对于影响调查水域鱼类资源时空变动的主要生物/非生物因子还有待进一步研究。
3.2 声学评估误差分析
声学评估作为海洋渔业资源评估与鱼类行为分析的一种有效方法,其评估结果受诸多因素的影响。传统的声学调查结合调查区域具体的地形地貌特征将调查航线设计为“之”字型或平行断面型2种,由于调查海域地理环境复杂且海上阻隔较多,导致声学调查航迹不规则,增加了声学取样产生随机误差的可能性[11]。受声学近场效应及海底声学探测盲区的影响,调查海域声学积分起始水层设置为海表1.5 m以下至海底0.5 m之上,故临近海表浮游性鱼类和底栖种类不在回波积分范围之内,从而给声学评估结果带来一定的偏差[29-30]。国内外相关研究结果表明,部分鱼类对船舶噪声存在明显的逃避行为,从而使声学评估结果偏小[31-32]。调查海域春季优势群体鹿斑鲾受船舶噪声的影响有明显的跃水现象,可能对声学评估结果带来一定影响。此外,声学回波映像处理结果显示,调查海域各季度均有小型鱼群聚集的现象,2014年秋季在中下层这类集群现象更为显著。据回波积分原理,当研究水域渔业资源存在集群现象时,遮蔽效应往往使资源数量密度评估结果偏低[33]。此外,受调查水域鱼类生理生活习性及理化环境因子的调节,鱼类资源在垂直方向上的组成与分布可能存在一定差异[34-35]。因此,渔业资源底层拖网数据用于整个水域渔业资源回波积分分配是影响渔业资源声学评估结果的另一重要因素。
3.3 不同评估方法的比较
基于回波积分原理分析比较了在鱼种单一和混合2种不同条件下,渔业资源声学评估结果的差异。研究显示,2015年春季渔获种类组成单一和混合2种不同条件下,调查水域渔业资源密度差异不显著;而2014年秋季不同条件下渔业资源声学评估结果存在显著性差异,这可能与调查海域不同季节参与声学评估种类优势集中程度相关。2015年春季调查水域用于声学评估种类优势集中程度极高,鹿斑鲾在渔获数量中所占比重高达86.87%,该时期很可能为鹿斑鲾繁殖盛期。在渔获种类组成单一和混合2种不同的假设前提下,调查海域该季度参与声学评估回波积分分配的种类主要为鹿斑鲾,因而评估结果基本一致。由此可知,在渔获优势集中度较高的特定水域,使用基本积分单元内平均目标强度现场测定法可作为简化渔业资源声学评估方法的一种有效技术手段。而2014年秋季调查水域参与声学评估种类优势集中程度相对较低,第一优势群体所占比重仅45.31%。单因素方差分析结果显示,在2种不同条件下渔业资源声学评估结果存在显著性差异。参照回波积分分配原则,调查海域渔获优势集中程度通过基本声学航程单元内平均声学反射系数(
$\overline \sigma $ )的估算 [公式(5)],对渔业资源声学评估结果产生了影响。此外,本研究分别利用声学评估法和扫海面积法对目标海域不同季节渔业资源密度进行了评估与分析,结果表明,各季度渔业资源声学评估结果明显低于扫海面积法计算结果。扫海面积法作为传统的渔业资源评估方法存在诸多局限。理论上,参照扫海面积法计算公式中,逃逸率E是影响评估结果的重要因子之一。国内外相关研究表明,渔业资源捕捞效率受网具类型、规格、地理形态特征、鱼类行为等众多因素的影响[36-37]。任何调查网具对于渔业资源生物学信息的获取均具有一定的地域局限性和选择性[38-39]。拖网捕捞作为一种主动的渔捞作业方式,其捕捞效率往往高于其他定置渔捞方法。复杂的地理环境往往能为游泳生物提供天然的庇护场所,因此,复杂的水域环境逃逸率往往较高[5,40]。受鱼类昼夜和季节迁移习性的调节,鱼类资源时空分布特征可能存在明显的差异,从而对渔业捕捞效率产生影响。相关研究显示,鱼类在夜间活动较为频繁,且有向表层迁移的现象;而在白天鱼类有向底层迁移的规律,且活动较为稀少[25,37]。此外,鱼类随季节水温的变化有从近岸向深水水域迁移的趋势[34]。因此,针对不同季节,底拖网地理位置的规划设计亦会对捕捞效率造成一定的影响。
扫海面积法采用站点抽样调查的方式,其覆盖率明显低于声学评估方法。渔业资源空间分布存在较大差异时,应尽量增加抽样站位数以削弱随机抽样产生的误差。本研究扫海面积法及声学评估结果均表明,调查海域渔业资源空间分布差异极大。水平方向2014年秋季A3站位短尾大眼鲷资源密度明显高于其他站位,2015年春季S14位点鹿斑鲾资源密度显著高于其他站位。而根据回波单体垂直分布特征,2014年秋季小型个体有向中下层聚集的趋势,故仅基于底拖网渔获数据分析整个水域渔业资源密度可能高估了该水域渔业资源状况。因此,在渔业资源垂直空间分布差异较大时,建议采用分层拖网的取样方法。综上分析可知,渔业资源空间分布特征是影响扫海面积法渔业资源评估结果的另一重要因子。而有关研究结果显示,生物/非生物环境参数是影响渔业资源变动与空间分布的关键要素[26,28,35,41]。因此,在今后的渔业资源研究工作中应同步结合理化环境因子时空梯度变化等方面的研究,以期更加科学地开展渔业资源评估工作。
声学评估法作为一种新兴的生态渔业评估手段,与传统渔业调查法相比,具有科学、便捷、高效且对渔业资源与水域生境的破坏力度小等特点[5-6],因而更能适应现时代渔业调查的需要,其应用前景十分广阔。然而,该方法在种类鉴定、映像识别、噪声消除等方面还有待进一步研究。因此,以声学评估为主体,综合采用传统生物学取样技术是当前渔业资源评估的有效方法,能为渔业资源科学管理及合理开发利用提供重要理论支撑。
-
![]()
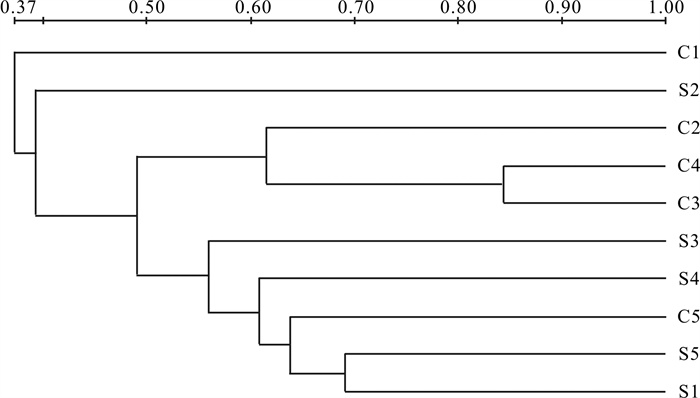
图 2 各处理组水体菌群随时间变化的变性梯度凝胶电泳分析示意图
S1~S5,C1~C5分别代表基质组、对照组不同采样时间样品;带边框为测序条带,编号自上向下依次为S2-1、S4-2、S5-4、C2-7、C5-8、S4-9、C1-10、S2-11、C3-13、C5-15、C2-16、C4-17、C3-18、S3-20、S5-21、C3-22、C5-23、S3-24
Figure 2. Sketch map of DGGE analysis of bacteria in water with time in different groups
S1~S5 represent the experimental groups and C1~C5 represent the control groups at different sampling time. The sequencing bands are framed, and cutting band No. from top to bottom is S2-1, S4-2, S5-4, C2-7, C5-8, S4-9, C1-10, S2-11, C3-13, C5-15, C2-16, C4-17, C3-18, S3-20, S5-21, C3-22, C5-23 and S3-24.
![]()
图 4 水体菌群结构动态变化的移动窗口分析
Figure 4. Variation of microbial communities in water by moving split-window analysis
![]()
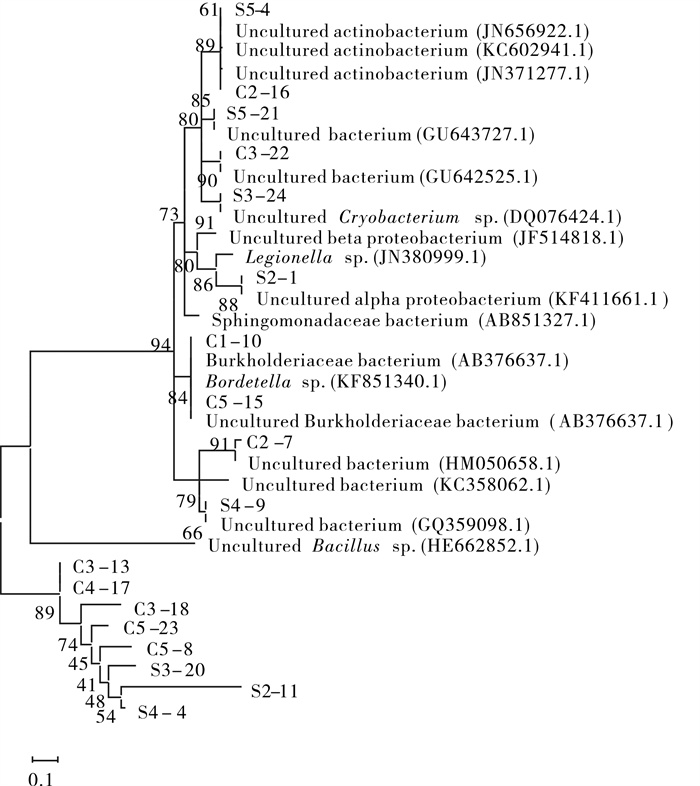
图 5 18条序列与一些相似序列以临接法构建的系统发生树
Figure 5. Phylogeny tree of 18 main sequences and similar ones by neighbor-joining method
表 1 特异性引物序列
Table 1 Specific sequence of primers
目的基因片段
target gene fragment引物名称
primer name引物序列(5′→3′)
primer sequence片段大小/ bp
product size16S rDNA[20] 8f AGAG TTTG ATCC TGGC TAAG 1 500 1492r GGTT ACCT TGTT ACGA CTT 16S rDNA-V3区[21] 341f-GC CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGCCTACGGGAGGCAG CAG 230 341f CCTACGGGAGGCAG CAG 534r ATTACCGCGGCTGCTGG  下载: 导出CSV
下载: 导出CSV
-
[1] 崔和. 我国对虾产销现状与国际对虾市场格局的变化[J]. 中国水产, 2014(8): 28-29. doi: 10.3969/j.issn.1002-6681.2014.08.014 [2] 周远扬, 曹俊明, 万忠, 等. 2013年广东对虾产业发展形势与对策建议[J]. 广东农业科学, 2014(8): 8-11. doi: 10.3969/j.issn.1004-874X.2014.08.003 [3] De SCHRYVER P, CRAB R, DEFOIRDT T, et al. The basics of bio-flocs technology: the added value for aquaculture[J]. Aquaculture, 2008, 277(3/4): 125-137. https://www.semanticscholar.org/paper/The-basics-of-bio-flocs-technology%3A-The-added-value-Schryver-Crab/77fe55196d4452612028f131c87dbe8fafcb174a
[4] 罗亮, 李卓佳, 张家松, 等. 对虾精养池塘碳、氮和异养细菌含量的变化及其相关性研究[J]. 南方水产科学, 2011, 7(5): 24-29. doi: 10.3969/j.issn.2095-0780.2011.05.004 [5] LIU H D, WANG L, LIU M, et al. The intestinal microbial diversity in Chinese shrimp (Fenneropenaeus chinensis) as determined by PCR-DGGE and clone library analyses[J]. Aquaculture, 2011, 317(1/2/3/4): 32-36. https://www.semanticscholar.org/paper/The-intestinal-microbial-diversity-in-Chinese-as-by-Liu-Wang/23054fffb0ae499e01911fe85fc8c0af13d39f65
[6] 吴伟, 陈家长, 胡庚东, 等. 利用人工基质构建固定化微生物膜对池塘养殖水体的原位修复[J]. 农业环境科学学报, 2008, 27(4): 1501-1507. doi: 10.3321/j.issn:1672-2043.2008.04.039 [7] ANAND P, KUMAR S, PANIGRAHI A, et al. Effects of C : N ratio and substrate integration on periphyton biomass, microbial dynamics and growth of Penaeus monodon juveniles[J]. Aquac Int, 2013, 21(2): 511-524. doi: 10.1007/s10499-012-9585-6
[8] THOMPSON F L, ABREU P C, WASIELESKY W. Importance of biofilm for water quality and nourishment in intensive shrimp culture[J]. Aquaculture, 2002, 203(3/4): 263-278. https://europepmc.org/article/AGR/IND23334984
[9] 陈亮亮, 董宏标, 张家松, 等. 6种改性剂对生物膜附着基质亲水性及其虾池挂膜效果的影响[J]. 南方水产科学, 2014, 10(2): 42-47. https://xueshu.baidu.com/usercenter/paper/show?paperid=d533da41798b09b38c3af4423ad14266&site=xueshu_se&hitarticle=1 [10] BRATVOLD D, BROWDY C L. Effects of sand sediment and vertical surfaces (AquaMatsTM) on production, water quality, and microbial ecology in an intensive Litopenaeus vannamei culture system[J]. Aquaculture, 2001, 195(1): 81-94. https://www.researchgate.net/publication/223661252_Effects_of_sand_sediment_and_vertical_surfaces_AquaMatsTM_on_production_water_quality_and_microbial_ecology_in_an_intensive_Litopenaeus_vannamei_culture_system
[11] OTOSHI C A, MONTGOMERY A D, MATSUDA E M, et al. Effects of artificial substrate and water source on growth of juvenile Pacific white shrimp, Litopenaeus vannamei[J]. J World Aquac Soc, 2006, 37(2): 210-213. doi: 10.1111/j.1749-7345.2006.00029.x
[12] 宋协法, 马真, 万荣. 纳米生态基在凡纳滨对虾养殖中的应用研究[J]. 中国海洋大学学报(自然科学版), 2010, 40(10): 24-28. https://xueshu.baidu.com/usercenter/paper/show?paperid=06931221c5f7354233d001b4313f5530&site=xueshu_se&hitarticle=1 [13] ZHANG B. Influence of the artificial substrates on the attachment behavior of Litopenaeus vannamei in the intensive culture condition[J]. Int J Anim Vet Adv, 2011, 3(1): 37-43. https://www.semanticscholar.org/paper/Influence-of-the-Artificial-Substrates-on-the-of-in-Zhang/b43d19eb558d482b050e09e33dd1f2e74a7647dd
[14] SCHVEITZER R, ARANTES R, BALOI M F, et al. Use of artificial substrates in the culture of Litopenaeus vannamei (Biofloc System) at different stocking densities: effects on microbial activity, water quality and production rates[J]. Aquac Eng, 2013, 54(3): 93-103. https://www.researchgate.net/publication/256974162_Use_of_artificial_substrates_in_the_culture_of_Litopenaeus_vannamei_Biofloc_System_at_different_stocking_densities_Effects_on_microbial_activity_water_quality_and_production_rates
[15] ZHANG J, CHEN L, DONG H, et al. Artificial substrates in zero-water-exchange culture system regulate the rearing performance of Pacific white shrimp Litopenaeus vannamei (Boone, 1931)under the winter indoor condition[J]. Aquac Res, 2014: 1-10. DOI: 10.1111/are.12473.
[16] PACE N R. A molecular view of microbial diversity and the biosphere[J]. Science, 1997, 276(5313): 734-740. doi: 10.1126/science.276.5313.734
[17] 罗鹏, 胡超群, 张吕平, 等. 凡纳滨对虾海水养殖系统内细菌群落的PCR-DGGE分析[J]. 中国水产科学, 2009, 16(1): 31-38. https://xueshu.baidu.com/usercenter/paper/show?paperid=073cb871c675aff240a054b145c665c6&site=xueshu_se&hitarticle=1 [18] 姚延丹, 李谷, 陶玲, 等. 复合池塘循环水养殖系统微生物群落结构分析[J]. 中国水产科学, 2011, 18(2): 407-415. https://xueshu.baidu.com/usercenter/paper/show?paperid=ffb8870503ceef6c39572b93c79fbdbc&site=xueshu_se&hitarticle=1 [19] 王亭芳, 金风杰, 马士禹, 等. DGGE技术对南美白对虾养殖水体中微生物多样性的研究[J]. 生物技术通报, 2012(10): 131-136. https://xueshu.baidu.com/usercenter/paper/show?paperid=db9b5e97433278bdfeadd56a0639f81f&site=xueshu_se&hitarticle=1 [20] PORTEOUS L A, ARMSTRONG J L. Recovery of bulk DNA from soil by a rapid, small-scale extraction method[J]. Curr Microbiol, 1991, 22(6): 345-348. doi: 10.1007/BF02092152
[21] MUYZER G. DGGE/TGGE a method for identifying genes from natural ecosystems[J]. Curr Opin Microbiol, 1999, 31(2): 7-22. https://www.semanticscholar.org/paper/DGGE-TGGE-a-method-for-identifying-genes-from-Muyzer/970566056daf1a863a5a515d63827a3440b7e374
[22] 张瑜, 韩玲, 谢露蓉. 基于移动窗口分析法的西安市热岛效应研究[J]. 测绘科学, 2012, 37(5): 148-150. [23] 李敬美, 金若菲, 周集体, 等. 活性污泥与生物膜复合工艺处理石化废水的研究[J]. 环境工程学报, 2011, 5(3): 523-527. http://hjhx.rcees.ac.cn/article/id/20110310 [24] FOUAD M, BHARGAVA R. Mathematical model for the bio-film-activated sludge reactor[J]. J Environ Eng, 2005, 131(4): 557-562. doi: 10.1061/(ASCE)0733-9372(2005)131:4(557)
[25] ZOKAEIFAR H, BABAEI N, SAAD C R, et al. Administration of Bacillus subtilis strains in the rearing water enhances the water quality, growth performance, immune response, and resistance against Vibrio harveyi infection in juvenile white shrimp, Litopenaeus vannamei[J]. Fish Shellfish Immunol, 2014, 36(1): 68-74. doi: 10.1016/j.fsi.2013.10.007
[26] DONG H B, SU Y Q, MAO Y, et al. Dietary supplementation with Bacillus can improve the growth and survival of the kuruma shrimp Marsupenaeus japonicus in high-temperature environments[J]. Aquac Int, 2014, 22(2): 607-617. doi: 10.1007/s10499-013-9688-8
[27] 李卓佳, 郭志勋, 冯娟, 等. 应用芽孢杆菌调控虾池微生态的初步研究[J]. 海洋科学, 2006, 30(11): 28-31. doi: 10.3969/j.issn.1000-3096.2006.11.007 [28] 陆家昌, 李活, 黄翔鹄. 枯草芽孢杆菌对水质及凡纳滨对虾幼体免疫指标影响的研究[J]. 南方水产, 2010, 6(1): 19-24. doi: 10.3969/j.issn.1673-2227.2010.01.004 -
期刊类型引用(5)
1. 石娟,刘永,李纯厚,王腾,赵金发,宋晓宇,谢宏宇. 珠江口棘头梅童鱼营养生态位研究. 南方水产科学. 2024(03): 56-65 .  本站查看
本站查看
2. 郑秋实,刘永,李纯厚,吴鹏,肖雅元,林琳,刘玉,邹剑. 季节变化对珠江口棘头梅童鱼肠道和水体中细菌群落结构的影响. 南方水产科学. 2024(04): 77-87 . 本站查看
3. 马菁菁,陈海刚,张喆,田斐,唐振朝,熊倩,张林宝. 2018—2023年珠江口鱼类群落结构变化及其与环境因子的关系. 南方水产科学. 2024(06): 62-73 . 本站查看
4. 熊朋莉,徐姗楠,陈作志,张帅,蒋佩文,范江涛. 珠江河口棘头梅童鱼时空分布以及影响因素的研究进展. 海洋科学. 2022(08): 79-87 . 百度学术
5. 汪润芝,原峰,李崇淑,龙焰,许清燕,奚冰柔,冼嘉俊,叶锦韶. 海上风电场建设与运行对渔业资源群落结构及空间分布的影响研究. 海洋开发与管理. 2022(12): 98-106 . 百度学术
其他类型引用(3)






















计量
- 文章访问数: 3266
- HTML全文浏览量: 144
- PDF下载量: 1459
- 被引次数: 8
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
