
Comparison on resources community of stow-net fishery before and after fishing off season in Huangmaohai Estuary
-
摘要:
根据2015年休渔前后(春季和夏季)利用张网在黄茅海河口获得的渔业资源调查资料,应用数量生物量比较曲线和生物量谱方法分析了其渔业资源群落状况。结果显示,调查共捕获渔获56种,其中鱼类47种,隶属于12目27科38属;甲壳类9种,隶属于2目5科7属。休渔前(春季)渔获种类大于休渔后(夏季),春季优势种为凤鲚(Coilia mystus)、红狼牙

Abundance-biomass comparison curve and biomass spectra methods were applied to analyze the data collected from stow-net fishery before and after fishing off season (spring and summer) in Huangmaohai Estuary so as to evaluate the status of fishery resources community. Fivty-six fishery species were collected during the surveys, including 47 fish species (12 orders, 27 families, 38 genera) and 9 crustacean species (2 orders, 5 families and 7 genera). The catch species in spring were more than that in summer. In spring, the dominant species were Coilia mystus, Odontamblyopus rubicundus and Exopalaemon carinicauda. In summer, the dominant species were C.mystus, Anchoviella commersonii and O.rubicundus. The k-dominance curves of abundance and biomass intersected in spring, and the W-statistic value was -0.015. But the k-dominance curve of abundance was above the biomass curve in summer with the W-statistic value of -0.046. This phenomenon indicates that the present resources community of stow-net fishery was disturbed moderately in spring, but disturbed seriously in summer. Furthermore, the slopes of normalized biomass spectra in spring were higher than -1, and the biomass increased with the increase of the average individual weight, but preferred the opposite in summer. The results indicate that fishing can affect the biomass spectra, and the fishing off season does not play an obviously positive role by the decline of the slope of biomass spectra under overfishing. To maintain the ecosystem health, management and restrictions must be imposed on the fishery.
-
Keywords:
- stow-net /
- fishery resource /
- community structure /
- ABC curves /
- biomass spectra /
- Huangmaohai Estuary
-
近岸水域由于其独特的地理位置及水环境特征,是海洋与陆地相互作用和海洋生物多样性保护研究的重要水域[1]。近年来,由于人类活动和海洋环境变化等,直接导致了近岸水域渔业资源的衰退,优质鱼比例下降,低质鱼成了主要捕捞对象,以及个体减小、性成熟提前等[2]。因此,人类干扰对渔业资源群落的影响越来越受到海洋渔业专家及生态专家的关注。
张网是南海近岸渔业的重要定置渔具,研究张网渔业资源群落有助于制定中国海洋渔具管理决策,特别是对于促进《全国海洋捕捞渔具目录》的实施具有重要意义。目前,已有部分学者对张网的作业概况、网目选择性、渔获组成多样性及渔业资源群落结构等进行了研究[2-9],并探讨了一系列分析群落变化的方法和指标[10]。粒径谱理论就是其中的一种重要方法,是由SHELDON和PARSONS首次提出的[11]。该理论已广泛应用于浮游生物、底栖生物、微型生物的群落特征分析及鱼类的潜在产量估算等方面[12-18]。因此,该研究利用黄茅海河口近岸休渔前后(春季和夏季)的张网调查资料,采用数量生物量比较曲线(abundance-biomass comparison curves,ABC曲线)和生物量谱方法对张网渔业资源群落状况进行了分析,以期有助于南海近岸渔业资源的合理开发和可持续利用。
1. 材料与方法
1.1 数据来源
数据来源于2015年休渔前后(春季和夏季)在珠江口沿岸新会崖门的黄茅海河口开展的张网调查,经纬度为113°06′ E,22°08′ N。休渔前(春季)调查时间为2015年4月16日~27日,有效网次为15网次;休渔后(夏季)航次时间为2015年8月19日~31日,有效网次为12网次。
调查渔船为“粤新会41008”船,木质,船长12.00 m,型宽2.85 m,型深0.83 m,总吨位8.0 GT,净吨位3.0 t,主机功率19.4 kW,2台,每台9.7 kW,携带张网4顶,船上还配备避碰仪、GPS导航仪等设备。
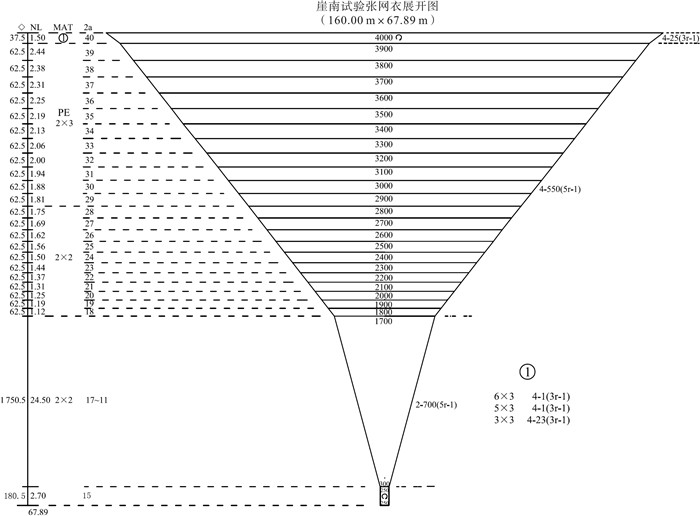
该张网属于双桩竖杆张网,是一种由双桩固定,由竖杆使网口垂直张开的单囊张网,主要张设于沿岸鱼虾类繁育的场所或近海鱼类的洄游通道,是依靠海流将捕捞对象裹挟进入网具达到捕捞目的的被动性网具。其网具主尺度为:40.00 m×67.89 m,即结附网衣的网口纲周长为40.00 m,网衣的纵向拉直总长为67.89 m。网衣展开图如图 1所示。每个网次作业时长约2.5 h。起网后对渔获物进行抽样,取1/8进行种类鉴定、分类计数、称重,并对渔获进行生物学测定,体长、甲宽等以mm为单位,体质量以g为单位。所有样品个体鉴定到种,体质量精确到0.1 g。
1.2 数据处理
1.2.1 物种优势度
采用相对重要性指数(index of relative importance,IRI)来评价渔获种类的优势种,IRI≥1 000为优势种,100≤IRI<1 000为重要种,10≤IRI<100为常见种,IRI<10为少见种[19-20],其计算公式为[19-20]:
$$ \mathrm{IRI}=(N \%+W \%) \times F \% $$ 其中N%表示某种渔获种类的尾数百分比;W%表示生物量百分比;F%表示出现频率百分比。
为进一步分析春夏两季渔获种类组成及每个种类的相对生物量差异,根据平均网次生物量经过平方根转换计算得到Bray-Curtis相似系数矩阵,采用相似性分析(ANOSIM)进行休渔前后(春季和夏季)的渔业资源群落结构差异的显著性检验[21]。
1.2.2 ABC曲线
采用ABC曲线方法来分析渔业资源群落受人类干扰情况,该方法是1986年由WARWICK[22]提出的,是通过在同一坐标系中比较数量优势度曲线和生物量优势度曲线来分析群落在不同干扰状况下的特征[10, 23]。ABC曲线方法反映了r选择和k选择的传统进化的理论背景。在未受干扰(稳定)的状态下,群落主要是以k选择种类(生长慢、性成熟晚的大个体种类)为主,生物量优势度曲线位于数据优势度曲线之上。随着干扰的增加,k选择物种的生物量(或数量)逐渐减少,r选择物种的生物量(或数量)则逐渐增加,当处于中等干扰(或不稳定)的状态时,两条曲线将相交;当群落逐渐变为由r选择的物种(生长快、个体小的种类)为主,此时生物量的优势度曲线在数量优势度曲线之下,则表明群落处于严重干扰的(不稳定)状态[9-10, 23]。
用W值表示ABC曲线中生物量和丰度的相对关系统计量[9-10]:
$$ W=\sum\limits_{i=1}^s \frac{\left(B_i-A_i\right)}{50(S-1)} $$ 式中Bi和Ai为ABC曲线中种类序号对应的生物量和数量的累积百分比,S为出现物种数。
1.2.3 生物量谱
生物量谱是粒径谱理论的变形[14]。随着粒径谱理论的发展,新的粒径谱理论模型相继提出,各种新的粒径谱表示方法也不断革新。P-D模型就是其中的典型模型之一,由PLATTER和DENMAN提出[24],该方法是以对数化生物量为纵坐标,对数化粒径为横坐标所得到的谱线,称为非标准化生物量谱。并提出标准化生物量的概念,某一粒径级的生物量与该粒径级宽度的比值,就称为标准化的生物量。
采用生物量谱方法分析生物量与平均个体质量之间的关系,先计算每个渔获种类的平均体质量(wi),并对其取对数,然后将对数化的平均体质量进行分组,以1 g为单位区间间隔,计算各个区间的总生物量,以对数化的平均个体体质量为x轴,以对数化的各个区间总生物量为y轴,线性回归后可以得到一条直线谱线[12]。谱线的斜率和截距分别表示群落的粒径结构和生物丰度。稳定状态下的生物群落标准化粒径谱是一条斜率为-1的直线,即生物量随着体质量的增加均匀分布;如果斜率大于-1,则生物量随着体质量的增加而增加;如果斜率小于-1,生物量随着体质量的增加而减少[13-14]。
2. 结果
2.1 渔获组成
共捕获渔获种类56种,其中鱼类47种,隶属于1纲12目27科38属;虾类5种,隶属于1纲1目2科3属;虾姑类1种,隶属于1纲1目1科1属;蟹类3种,隶属于1纲1目2科3属。春季捕获渔获种类44种,夏季40种,其中共同种28种。春季张网渔获优势种为凤鲚(Coilia mystus)、红狼牙

渔获种类species 春季spring 夏季summer F/% N/% W/% IRI F/% N/% W/% IRI 斑点鸡笼鲳Drepane punctata 6.67 0.02 0.06 0.51 - - - - 斑 
46.67 0.35 1.67 94.26 75.00 1.09 1.90 224.39 斑头舌鳎Cynoglossus puncticeps 20.00 0.31 0.13 8.80 8.33 0.02 0.02 0.32 赤鼻棱鳀Thrissa kammalensis - - - - 58.33 1.47 2.65 240.84 大甲 
- - - - 8.33 0.01 0.05 0.57 弹涂鱼Periophthalmidae cantonensis - - - - 8.33 0.03 0.04 0.54 刺鲳Psenopsis anomala 6.67 0.02 0.02 0.28 - - - - 聪明关公蟹Dorippe astuta 26.67 0.35 0.10 11.82 - - - - 粗吻海龙Trachyrhamphus serratus 6.67 0.02 0.01 0.16 - - - - 大鳞舌鳎Cynoglossus macrolepidotus 6.67 0.02 0.63 4.31 - - - - 带鱼Trichiurus lepturus 13.33 0.04 0.14 2.31 - - - - 刀额新对虾Metapenaeus ensis 86.67 2.12 1.27 293.83 91.67 1.66 0.78 223.82 杜氏叫姑鱼Johnius dussumieri 66.67 0.53 0.85 91.94 - - - - 杜氏棱鳀Thryssa dussumieri 46.67 0.18 0.24 19.97 - - - - 短吻蛇鳗Ophichthus brevirostris 6.67 0.02 0.01 0.20 - - - - 短吻新银鱼Neosalanx brevirostris 13.33 0.05 0.02 0.97 - - - - 多鳞 
6.67 0.02 0.03 0.36 25.00 0.08 0.25 8.31 凤鲚Coilia mystus 100.00 36.51 54.83 9 133.79 100.00 31.70 45.08 7 678.42 弓斑东方鲀Takifugu ocellatus 80.00 4.89 1.94 546.83 41.67 0.07 0.74 33.67 海南 
- - - - 8.33 0.01 0.01 0.15 海鳗Muraenesox cinereus 13.33 0.04 0.62 8.80 - - - - 海鲶Ariussinensis Lacepede 40.00 0.16 0.26 16.96 58.33 0.55 1.13 97.79 褐斑三线舌鳎Cynoglossus trigrammus 80.00 1.48 0.74 177.79 58.33 0.16 0.19 20.63 黑尾吻鳗Rhynchoconger ectenurus 20.00 0.05 0.06 2.37 - - - - 红狼牙虎鱼 100.00 25.44 19.23 4 467.24 91.67 7.09 11.22 1 678.19 花 33.33 0.13 0.56 22.93 - - - - 黄斑鲾Leiognathus bindus 66.67 0.37 0.29 43.78 91.67 5.22 2.78 733.51 黄斑蓝子鱼Siganus oramin 6.67 0.02 0.07 0.57 16.67 0.04 0.08 1.89 黄鳍鲷Sparus latus 6.67 0.02 0.03 0.33 - - - - 棘头梅童鱼Collichthys lucidus 60.00 0.46 2.16 156.80 91.67 2.33 6.09 771.42 脊突猛虾蛄Harpiosquilla raphidea 53.33 0.42 0.54 51.23 8.33 0.01 0.02 0.24 脊尾白虾Exopalaemon carinicauda 100.00 10.15 2.81 1 296.13 91.67 3.70 1.00 431.06 尖吻蛇鳗Ophichthus apicalis 13.33 0.05 0.08 1.76 8.33 0.01 0.01 0.13 近缘新对虾Metapenaeus affinis - - - - 8.33 0.01 0.02 0.25 居氏银鱼Salanx cuvieri 60.00 1.01 0.29 77.82 33.33 0.09 0.06 4.98 锯缘青蟹Scylla serrata - - - - 8.33 0.01 0.28 2.41 康氏小公鱼Anchoviella commersonii 46.67 0.82 0.57 65.15 100.00 32.08 13.75 4 583.75 孔 80.00 3.19 1.28 357.28 41.67 0.25 0.38 25.91 鳓Ilisha elongata 6.67 0.02 0.22 1.61 41.67 0.16 0.48 26.48 丽叶鲹Caranx kalla - - - - 75.00 5.53 5.21 805.43 粒突鳞鲬Onigocia tuberculatus 46.67 0.31 0.10 19.13 50.00 0.14 0.14 14.33 龙头鱼Harpodon nehereus - - - - 41.67 0.29 0.27 23.18 矛尾 86.67 2.40 0.72 270.09 91.67 0.80 1.04 169.13 皮氏叫姑鱼Johnius belengeri - - - - 75.00 0.38 0.48 64.78 乳香鱼Lactarius lactarius - - - - 41.67 0.74 0.23 40.65 少鳞 - - - - 8.33 0.01 0.01 0.16 四指马鲅Eleutheronema tetradactylum - - - - 8.33 0.01 0.26 2.25 纹缟 93.33 0.88 0.46 124.92 25.00 0.04 0.03 1.69 享氏仿对虾Parapenaeopsis hungerfordi 20.00 0.07 0.03 2.06 - - - - 硬头骨鲻Osteomugil strongylocephalus 53.33 0.70 0.61 69.80 50.00 0.21 0.74 47.33 圆颌针鱼Tylosurus melanotus 6.67 0.04 0.08 0.77 8.33 0.06 0.15 1.70 直额鲟Charybdis truncate 86.67 2.99 2.57 481.85 - - - - 中华青鳞鱼Harengula nymphaca 80.00 1.59 3.00 367.34 91.67 0.76 1.14 174.22 中华小公鱼Stolephorus chinensis 6.67 0.15 0.11 1.68 25.00 0.21 0.08 7.16 中线天竺鲷Apogon kallopterus 60.00 1.28 0.38 99.97 83.33 0.72 0.46 98.85 周氏新对虾Metapenaeus joyneri 46.67 0.31 0.19 23.29 75.00 2.25 0.74 224.73 2.2 ABC曲线
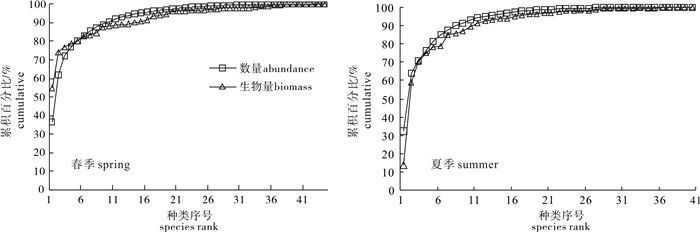
春夏两季渔业资源群落的ABC曲线见图 2。春季渔业资源群落的数量优势度曲线与生物量的优势度曲线相交,处于中等干扰状态,其根据ABC曲线计算的W统计值为-0.015;夏季渔业资源群落的数量优势度曲线在生物量优势度曲线之上,处于严重干扰状态[10],其根据ABC曲线计算的W统计值为-0.046。表明该海域的渔业资源群落中,小型个体或大型鱼类的幼体占绝对优势,且春季该海域的渔业资源群落结构优于夏季。
春季数量百分比排在前5位的渔获分别为凤鲚、红狼牙
夏季数量百分比排在前5位的渔获分别为康氏小公鱼、凤鲚、红狼牙
2.3 生物量谱
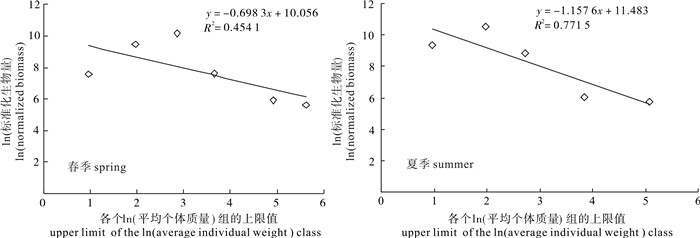
春夏两季标准化生物量谱线性回归分析见图 3,回归方程分别为y=-0.698 3x+10.056(R2= 0.454 1)和y=-1.157 6x+11.483(R2=0.771 5)。春季标准化生物量谱线性回归的斜率大于-1,说明春季张网渔业资源群落的生物量随着体质量的增加不呈均匀分布,而是随着体质量的增加生物量也增加;夏季标准化生物量谱线性回归的结果与春季差异较大,其斜率小于-1,说明夏季张网渔业资源群落的生物量随着体质量的增加而减少,其相关系数R2值也明显高于春季。
![]() 图 3 各季节生物量谱的回归分析Figure 3. Regression analysis for biomass spectra of resources community in spring and summer
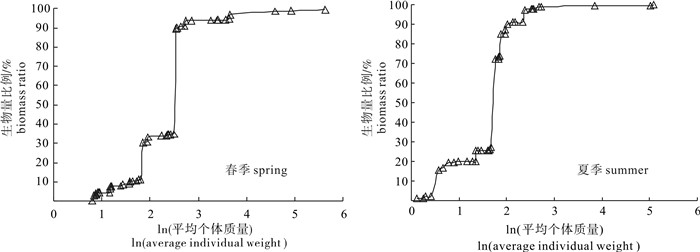
图 3 各季节生物量谱的回归分析Figure 3. Regression analysis for biomass spectra of resources community in spring and summer从生物量随个体平均体质量的变化来看(图 4),春夏两季生物量随着个体平均体质量的增加不呈规律分布,不同个体体质量区间的生物量比例差异较大,小个体鱼种的生物量比例相对较高,其中凤鲚所占生物量百分比跨度最高。春季体质量小于10 g的生物量比例达到33.91%,小于20 g的生物量比例达到93.88%,以个体体质量在10~20 g的生物量比例最高;夏季个体体质量小于10 g的生物量比例达到90.93%,以个体体质量小于10 g的生物量占绝对优势。对比春夏两季渔获的个体体质量,可以看出春季渔获的平均个体体质量明显高于夏季。
![]() 图 4 各季节不同个体体质量区间的生物量比例Figure 4. Biomass spectra of resources community in spring and summer
图 4 各季节不同个体体质量区间的生物量比例Figure 4. Biomass spectra of resources community in spring and summer3. 讨论
张网由于其良好的作业性能和较高的经济效益,目前仍然是中国近海一种重要的作业方式。该研究以张网为研究对象,其原因为:1)张网是在无外界干扰情况下,使渔获物被潮流裹挟入网,渔法上选择性较小,更接近自然分布[25],以该网具研究该海域的渔业资源群落结构更加合理;2)张网目前属于过渡渔具,而专家对其今后划分为准用渔具还是禁用渔具意见不一,该研究提供的基础数据有助于更加准确地把握该种渔具的捕捞现状。
研究显示,ABC曲线特征反映了群落中大型种类和小型种类相对数量的变化以及个体大小组成的变化[23, 26]。珠江口近岸张网渔业资源群落不同季节ABC曲线特征有所差异,春季渔业资源群落的数量优势度曲线与生物量的优势度曲线相交,处于中等干扰状态,而夏季渔业资源群落的数量优势度曲线在生物量优势度曲线之上,处于严重干扰状态。表明该海域的渔业资源群落中,小型个体或大型鱼类的幼体占绝对优势。研究显示休渔对保护经济种类及其幼鱼、幼体的繁衍生长能起到一定的作用[10, 25, 27]。从调查时间来看,春夏两次调查正好是伏季休渔的前后,但是从夏季渔业资源群落的状况来看,休渔在过度捕捞下并没有达到预期效果,主捕对象的比例以及个体体质量反而不如春季。另外该张网作业主要集中于春夏秋3个季节作业,在冬季作业次数少,渔业资源能得到一定程度上的恢复,且该夏季渔获调查并未在休渔结束后立马进行,故存在一定的差异。
生物量谱是生物个体大小的分布模式,是能量在生物群落中的分布[28]。研究显示,捕捞能够影响鱼类粒径谱,捕捞死亡可以导致粒径谱斜率下降,反映了大型鱼类的减少[29-30];反之,鱼类粒径谱也可以反映捕捞的影响,探测该资源群落是否处于过度开发状态[31]。标准化生物量谱线性回归分析结果显示夏季渔业资源群落生物量谱斜率下降,说明该海域张网渔业已处于过度捕捞状态。且从生物量随个体体质量的分布来看,春夏两季种类组成中小个体的比例都非常高,春季以个体体质量在10~20 g的生物量比例最高(59.97%);夏季个体体质量小于10 g的生物量比例达到90.93%。这些小个体的种类包括凤鲚、红狼牙
近岸水域由于其水环境条件的变化,导致群体优势种和一些种类的交替变化[32]。该张网主要在阴历初一和十五前后作业,受月相影响较大。且由于资料的缺失和调查条件的限制,未能从年间变动以及全年序列来分析该海域的渔业资源群落特征,具有一定的局限性。在今后的研究工作中,应加强连续性,并加强对环境因子的影响研究。
-
![]()
图 3 各季节生物量谱的回归分析
Figure 3. Regression analysis for biomass spectra of resources community in spring and summer
![]()
图 4 各季节不同个体体质量区间的生物量比例
Figure 4. Biomass spectra of resources community in spring and summer
表 1 黄茅海河口张网优势种组成
Table 1 Dominant species of stow-net in Huangmaohai Estuary
渔获种类species 春季spring 夏季summer F/% N/% W/% IRI F/% N/% W/% IRI 斑点鸡笼鲳Drepane punctata 6.67 0.02 0.06 0.51 - - - - 斑 46.67 0.35 1.67 94.26 75.00 1.09 1.90 224.39 斑头舌鳎Cynoglossus puncticeps 20.00 0.31 0.13 8.80 8.33 0.02 0.02 0.32 赤鼻棱鳀Thrissa kammalensis - - - - 58.33 1.47 2.65 240.84 大甲 - - - - 8.33 0.01 0.05 0.57 弹涂鱼Periophthalmidae cantonensis - - - - 8.33 0.03 0.04 0.54 刺鲳Psenopsis anomala 6.67 0.02 0.02 0.28 - - - - 聪明关公蟹Dorippe astuta 26.67 0.35 0.10 11.82 - - - - 粗吻海龙Trachyrhamphus serratus 6.67 0.02 0.01 0.16 - - - - 大鳞舌鳎Cynoglossus macrolepidotus 6.67 0.02 0.63 4.31 - - - - 带鱼Trichiurus lepturus 13.33 0.04 0.14 2.31 - - - - 刀额新对虾Metapenaeus ensis 86.67 2.12 1.27 293.83 91.67 1.66 0.78 223.82 杜氏叫姑鱼Johnius dussumieri 66.67 0.53 0.85 91.94 - - - - 杜氏棱鳀Thryssa dussumieri 46.67 0.18 0.24 19.97 - - - - 短吻蛇鳗Ophichthus brevirostris 6.67 0.02 0.01 0.20 - - - - 短吻新银鱼Neosalanx brevirostris 13.33 0.05 0.02 0.97 - - - - 多鳞 6.67 0.02 0.03 0.36 25.00 0.08 0.25 8.31 凤鲚Coilia mystus 100.00 36.51 54.83 9 133.79 100.00 31.70 45.08 7 678.42 弓斑东方鲀Takifugu ocellatus 80.00 4.89 1.94 546.83 41.67 0.07 0.74 33.67 海南 - - - - 8.33 0.01 0.01 0.15 海鳗Muraenesox cinereus 13.33 0.04 0.62 8.80 - - - - 海鲶Ariussinensis Lacepede 40.00 0.16 0.26 16.96 58.33 0.55 1.13 97.79 褐斑三线舌鳎Cynoglossus trigrammus 80.00 1.48 0.74 177.79 58.33 0.16 0.19 20.63 黑尾吻鳗Rhynchoconger ectenurus 20.00 0.05 0.06 2.37 - - - - 红狼牙虎鱼 100.00 25.44 19.23 4 467.24 91.67 7.09 11.22 1 678.19 花 33.33 0.13 0.56 22.93 - - - - 黄斑鲾Leiognathus bindus 66.67 0.37 0.29 43.78 91.67 5.22 2.78 733.51 黄斑蓝子鱼Siganus oramin 6.67 0.02 0.07 0.57 16.67 0.04 0.08 1.89 黄鳍鲷Sparus latus 6.67 0.02 0.03 0.33 - - - - 棘头梅童鱼Collichthys lucidus 60.00 0.46 2.16 156.80 91.67 2.33 6.09 771.42 脊突猛虾蛄Harpiosquilla raphidea 53.33 0.42 0.54 51.23 8.33 0.01 0.02 0.24 脊尾白虾Exopalaemon carinicauda 100.00 10.15 2.81 1 296.13 91.67 3.70 1.00 431.06 尖吻蛇鳗Ophichthus apicalis 13.33 0.05 0.08 1.76 8.33 0.01 0.01 0.13 近缘新对虾Metapenaeus affinis - - - - 8.33 0.01 0.02 0.25 居氏银鱼Salanx cuvieri 60.00 1.01 0.29 77.82 33.33 0.09 0.06 4.98 锯缘青蟹Scylla serrata - - - - 8.33 0.01 0.28 2.41 康氏小公鱼Anchoviella commersonii 46.67 0.82 0.57 65.15 100.00 32.08 13.75 4 583.75 孔 80.00 3.19 1.28 357.28 41.67 0.25 0.38 25.91 鳓Ilisha elongata 6.67 0.02 0.22 1.61 41.67 0.16 0.48 26.48 丽叶鲹Caranx kalla - - - - 75.00 5.53 5.21 805.43 粒突鳞鲬Onigocia tuberculatus 46.67 0.31 0.10 19.13 50.00 0.14 0.14 14.33 龙头鱼Harpodon nehereus - - - - 41.67 0.29 0.27 23.18 矛尾 86.67 2.40 0.72 270.09 91.67 0.80 1.04 169.13 皮氏叫姑鱼Johnius belengeri - - - - 75.00 0.38 0.48 64.78 乳香鱼Lactarius lactarius - - - - 41.67 0.74 0.23 40.65 少鳞 - - - - 8.33 0.01 0.01 0.16 四指马鲅Eleutheronema tetradactylum - - - - 8.33 0.01 0.26 2.25 纹缟 93.33 0.88 0.46 124.92 25.00 0.04 0.03 1.69 享氏仿对虾Parapenaeopsis hungerfordi 20.00 0.07 0.03 2.06 - - - - 硬头骨鲻Osteomugil strongylocephalus 53.33 0.70 0.61 69.80 50.00 0.21 0.74 47.33 圆颌针鱼Tylosurus melanotus 6.67 0.04 0.08 0.77 8.33 0.06 0.15 1.70 直额鲟Charybdis truncate 86.67 2.99 2.57 481.85 - - - - 中华青鳞鱼Harengula nymphaca 80.00 1.59 3.00 367.34 91.67 0.76 1.14 174.22 中华小公鱼Stolephorus chinensis 6.67 0.15 0.11 1.68 25.00 0.21 0.08 7.16 中线天竺鲷Apogon kallopterus 60.00 1.28 0.38 99.97 83.33 0.72 0.46 98.85 周氏新对虾Metapenaeus joyneri 46.67 0.31 0.19 23.29 75.00 2.25 0.74 224.73  下载: 导出CSV
下载: 导出CSV
-
[1] 任一平, 徐宾铎, 叶振江, 等. 青岛近海春、秋季渔业资源群落结构特征的初步研究[J]. 中国海洋大学学报(自然科学版), 2005, 35(5): 792-798. https://xueshu.baidu.com/usercenter/paper/show?paperid=2c4de7de79c60670ad3766363f14b469&site=xueshu_se [2] 张旭丰, 杨吝, 张鹏, 等. 南海北部张网主要技术参数的变化分析[J]. 南方水产, 2008, 4(6): 108-112. doi: 10.3969/j.issn.2095-0780.2008.06.016 [3] 晏磊, 杨吝, 张旭丰, 等. 南海北部张网作业状况及其管理探讨[J]. 广东农业科学, 2014, 41(6): 140-143. doi: 10.3969/j.issn.1004-874X.2014.06.036 [4] 晏磊, 杨吝, 张旭丰, 等. 南海北部竖杆张网网囊网目选择性研究[J]. 广东农业科学, 2014, 41(8): 171-176. doi: 10.3969/j.issn.1004-874X.2014.08.039 [5] 孙中之, 周军, 王俊, 等. 黄渤海区张网渔业[J]. 渔业科学进展, 2012, 33(3): 94-101. doi: 10.3969/j.issn.1000-7075.2012.03.014 [6] 邹莉, 苗振清, 俞存根, 等. 帆张网渔获物组成及其多样性分析[J]. 南方水产, 2010, 6(6): 46-53. doi: 10.3969/j.issn.1673-2227.2010.06.008 [7] 刘尊雷, 汤建华, 林龙山, 等. 江苏沿岸定置张网主要渔获组成以及对经济鱼类幼体的损害分析[J]. 海洋渔业, 2009, 31(1): 16-26. doi: 10.3969/j.issn.1004-2490.2009.01.003 [8] 张壮丽, 王茵. 闽南海区张网作业渔获物组成分析[J]. 海洋渔业, 2005, 27(2): 129-132. https://d.wanfangdata.com.cn/periodical/ChpNaW5lclBlcmlvZGljYWxDSEkyMDIzMDMyMRINaHl5eTIwMDUwMjAwNxoIcXNzYjQ1NWE%3D [9] 晏磊, 谭永光, 杨吝, 等. 南海珠江口沿岸张网渔业资源群落结构分析[J]. 生物学杂志, 2015, 32(5): 52-57. https://m.fx361.cc/news/2015/0322/20783973.html [10] 李圣法. 以数量生物量比较曲线评价东海鱼类群落的状况[J]. 中国水产科学, 2008, 15(1): 136-144. doi: 10.3321/j.issn:1005-8737.2008.01.018 [11] SHELDON R W, PARSONS T R. A continuous size spectrum for particulate matter in the sea[J]. J Fish Res Bd Can, 1967, 24(5): 909-915. doi: 10.1139/f67-081
[12] MAGNUSSEN E. Demersal fish assemblages of Faroe Bank: species composition, distribution, biomass spectrum and diversity[J]. Mar Ecol Prog Ser, 2002, 238: 211-225. https://www.academia.edu/93244695/Demersal_fish_assemblages_of_Faroe_Bank_species_composition_distribution_biomass_spectrum_and_diversity
[13] MACPHERSON E, GORDOA A, GARCÍA-RUBIES A. Biomass size spectra in littoral fishes in protected and unprotected areas in the NW Mediterranean[J]. Est Coast Shelf Sci, 2002, 55(5): 777-788. https://digital.csic.es/handle/10261/42031
[14] 周林滨, 谭烨辉, 黄良民, 等. 水生生物粒径谱/生物量谱研究进展[J]. 生态学报, 2010, 30(12): 3319-3333. https://www.cnki.com.cn/Article/CJFDTotal-STXB201012030.htm [15] ZHOU M. What determines the slope of a plankton biomass spectrum?[J]. J Plankton Res, 2006, 28(5): 437-448. https://academic.oup.com/plankt/article-abstract/28/5/437/1457472?redirectedFrom=fulltext&login=false
[16] BASEDOW S L, TANDE K S, ZHOU M. Biovolume spectrum theories applied: spatial patterns of trophic levels within a mesozooplankton community at the polar front[J]. J Plankton Res, 2010, 32(8): 1105-1119. https://www.peeref.com/zh/works/12460925
[17] SABEEL R A, VANREUSEL A. Potential impact of mangrove clearance on biomass and biomass size spectra of nematode along the Sudanese Red Sea coast[J]. Mar Environ Res, 2015, 103: 46-55. https://www.researchgate.net/publication/267815891_Potential_impact_of_mangrove_clearance_on_biomass_and_biomass_size_spectra_of_nematode_along_the_Sudanese_Red_Sea_coast
[18] HUA E, ZHANG Z, WARWICK R M, et al. Pattern of benthic biomass size spectra from shallow waters in the East China Sea[J]. Mar Biol, 2013, 160(7): 1723-1736. http://www.irgrid.ac.cn/handle/1471x/1154772
[19] PINKAS L, OLIPHANT M S, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. Calif Dept Fish Game: Fish Bull, 1971, 152: 1-105. https://searchworks.stanford.edu/view/8590885
[20] 程济生. 黄海无脊椎动物资源结构及多样性[J]. 中国水产科学, 2005, 12(1): 68-75. doi: 10.3321/j.issn:1005-8737.2005.01.013 [21] 梅春, 徐宾铎, 薛莹, 等. 胶州湾中部海域秋、冬季鱼类群落结构及其多样性研究[J]. 中国水产科学, 2010, 17(1): 110-118. https://d.wanfangdata.com.cn/periodical/ChpNaW5lclBlcmlvZGljYWxDSEkyMDIzMDcyNRIPemdzY2t4MjAxMDAxMDEzGggyMmh0NHBiYQ%3D%3D [22] WARWICK R M. A new method for detecting pollution effects on marine macrobenthic communities[J]. Mar Biol, 1986, 92(4): 557-562. doi: 10.1007/BF00392515
[23] YEMANE D, FIELD J G, LESLIE R W. Exploring the effects of fishing on fish assemblages using abundance biomass comparison (ABC) curves[J]. ICES J Mar Sci, 2005, 62(3): 374-379. http://tvhdh.vnio.org.vn:8080/xmlui/handle/123456789/6016
[24] PLATT T, DENMAN K. Organisation in the pelagic ecosystem[J]. Helgoländer wiss Meeresunters, 1977, 30(1/2/3/4): 575-581. doi: 10.1007/BF02207862
[25] 张庆生, 马永钧. 浙江沿海张网作业区幼带鱼分布、损害及休渔效果的分析[J]. 海洋科学, 1987(6): 43-47. [26] BLANCHARD F, LELOC'H F, HILY C, et al. Fishing effects on diversity, size and community structure of the benthic invertebrate and fish megafauna on the Bay of Biscay coast of France[J]. Mar Ecol Prog Ser, 2004, 280(1): 249-260. doi: 10.3354/meps280249
[27] 张龙, 徐汉祥, 王甲刚, 等. 舟山沿岸定置张网作业休渔前后鱼类组成分析[J]. 浙江海洋学院学报(自然科学版), 2011, 30(1): 1-8. https://wenku.baidu.com/view/6380067ff46527d3240ce073.html?_wkts_=1730270673364 [28] 陈峻峰. 南黄海浮游动物生物量谱及其应用[D]. 上海: 上海海洋大学, 2013: 1-6.https://d.wanfangdata.com.cn/thesis/D372726 [29] SHIN Y J, CURY P. Using an individual-based model of fish assemblages to study the response of size spectra to changes in fishing[J]. Can J Fish Aquat Sci, 2004, 61(3): 414-431. doi: 10.1139/f03-154
[30] BLANCHARD J L, JENNINGS S, LAW R, et al. How does abundance scale with body size in coupled size-structured food webs?[J]. J Anim Ecol, 2009, 78(1): 270-280.
[31] RICE J, GISLASON H. Patterns of change in the size spectra of numbers and diversity of the North Sea fish assemblage, as reflected in surveys and models[J]. ICES J Mar Sci, 1996, 53(6): 1214-1225. https://paperity.org/p/88053184/patterns-of-change-in-the-size-spectra-of-numbers-and-diversity-of-the-north-sea-fish
[32] 李永振, 陈国宝, 孙典荣. 珠江口鱼类组成分析[J]. 水产学报, 2000, 24(4): 312-317. http://scxuebao.ijournals.cn/scxuebao/article/abstract/20000404 -
期刊类型引用(15)
1. 黄央央,杨毕铖,戴桂香. 互花米草入侵对漳江口红树林保护区的影响研究. 海洋开发与管理. 2023(02): 115-124 .  百度学术
百度学术
2. Guangjie FANG,Haolin YU,Xinmeng WANG,Huaxiang SHENG,Yanli TANG,Changdong LIU,Chuanxi CHEN,Zhenlin LIANG. Impact of summer moratorium on set-net fishery in Haizhou Bay, China. Journal of Oceanology and Limnology. 2022(04): 1678-1691 . 必应学术
3. 陈静,黄德练,王雪辉,徐磊,张健,李亚芳,宁加佳,王亮根,刘双双,林昭进,杜飞雁. 基于DNA条形码的江门近岸海域春季鱼卵的种类鉴定及其形态. 南方水产科学. 2022(06): 10-18 . 本站查看
4. 徐姗楠,郭建忠,陈作志,张魁,许友伟,李纯厚. 胶州湾鱼类生物量粒径谱特征. 水产学报. 2020(04): 596-605 . 百度学术
5. 陈梓林,李纯厚,肖雅元,刘永,林琳,王九江,全秋梅. 江门近岸海域大型底栖动物群落结构的分布特征. 南方水产科学. 2020(04): 18-27 . 本站查看
6. 徐姗楠,郭建忠,范江涛,许友伟,粟丽,李纯厚. 大亚湾夏季鱼类生物量粒径谱年际变化特征. 南方水产科学. 2020(04): 28-38 . 本站查看
7. 徐姗楠,郭建忠,陈作志,许友伟,周林滨,柯志新,李纯厚. 大亚湾鱼类生物量粒径谱特征. 中国水产科学. 2019(01): 34-43 . 百度学术
8. 苏莹佳,陈国宝,周艳波,马胜伟,吴洽儿. 2015—2017年南海海域伏季休渔制度实施效果评价. 南方水产科学. 2019(02): 20-28 . 本站查看
9. 韦朋,王雪辉,马胜伟,周艳波,黄应邦,苏莹佳,吴洽儿. 我国南海区海洋捕捞现状分析. 上海海洋大学学报. 2019(06): 976-982 . 百度学术
10. 余景,胡启伟,袁华荣,陈丕茂. 基于遥感数据的大亚湾伏季休渔效果评价. 南方水产科学. 2018(03): 1-9 . 本站查看
11. 晏磊,杨吝,谭永光,杨炳忠,张鹏,李杰. 黄茅海河口张网网囊网目对凤鲚的选择性研究. 上海海洋大学学报. 2018(02): 274-283 . 百度学术
12. 张楠,吴娜,郭华阳,朱克诚,刘永,李纯厚,杨静文,江世贵,张殿昌. 基于DNA条形码技术对江门沿岸海域夏季鱼卵的鉴定. 中国水产科学. 2018(04): 721-727 . 百度学术
13. 谢志超,孙典荣,刘永,林琳,王腾,肖雅元,李纯厚. 江门海域游泳动物群落组成及其多样性初步分析. 南方水产科学. 2018(05): 21-28 . 本站查看
14. Yu Jing,Hu Qiwei,Chen Pimao,Mang Qi. Effect Evaluation of Summer Fishing Moratorium (SFM) in Daya Bay Based on Remote Sensing and Survey Data. Animal Husbandry and Feed Science. 2018(04): 219-225 . 必应学术
15. 郭建忠,陈作志,徐姗楠. 鱼类粒径谱研究进展. 海洋渔业. 2017(05): 582-591 . 百度学术
其他类型引用(7)
计量
- 文章访问数: 3360
- HTML全文浏览量: 219
- PDF下载量: 1000
- 被引次数: 22
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
