
Effect of six modifiers on hydrophilicity of biofilm-substrate and membrane-hanging result in prawn pond
-
摘要:
文章旨在研究提高生物膜附着基质亲水性能、缩短挂膜时间的相关改性处理方法,为该项技术应用于水产养殖废水处理工艺提供相关数据参考。研究选择筛绢网(40目)作为生物膜附着基质,选取氢氧化钠(NaOH)、氢氧化钙[Ca(OH)2]、盐酸(HCl)、高锰酸钾(KMnO4)、重铬酸钾(K2Cr2O7)和无水乙醇(C2H5OH)共6种试剂作为试验改性剂。通过基质吸附芽孢杆菌(Bacillus spp.)数量和对虾养殖池内挂膜效果来评价6种改性剂的改性效果。结果表明,10% HCl处理6 h后的基质亲水性更好,虾池挂膜效果最优,为该试验条件下生物膜附着基质亲水性改性的最适改性剂。
Abstract:We discussed the correlate parameters of modifiers enhancing the hydrophilicity of the biofilm-substrate and promoting the mature of the membrane, which may provide references for their application in aquaculture wastewater treatment process. The silk net (aperture 0.425 mm) was chosen as the biofilm-substrate and six modifiers [NaOH, Ca(OH)2, HCl, KMnO4, K2Cr2O7, C2H5OH] were tested as surface treatment reagents of substrate. The amount ofBacillus spp. adhered to substrate and the membrane-hanging effect in aquaculture wastewater provided some key evaluation indice for the six modifiers. The results show that compared with the other treatments, the substrate treated by 10% HCl for 6 h obtained the best hydrophilic ability and better membrane-hanging effect.
-
Keywords:
- biofilm-substrate /
- hydrophilic ability /
- modifier /
- membrane-hanging effect
-
考洲洋位于广东省惠东县,是红海湾向内陆延伸的典型半封闭性浅水海湾,其自然地理条件独特,是粤东地区重要的水产增养殖海域之一[1]。此外,考洲洋还是惠东红树林主要分布区,红树林可为鸟类、昆虫、贝类等提供栖息地,其根系复杂,也是鱼类和甲壳动物等游泳动物重要的庇护和摄食场所[2];红树林与渔业资源的关系也十分紧密,对当地渔业资源产量有着显著的提升作用[3]。当前考洲洋牡蛎养殖过程中建设的大量养殖筏架在一定程度上也能起到人工鱼礁的作用,吸引鱼类聚集[4]。同时,双壳动物强烈的生物沉积作用可为底栖动物提供食物源,从而进一步影响食物网的能量传递[5]。
近年来由于考洲洋海域的人工养殖密度逐渐上升,加之其周边是重要的皮革产区,大量污染物质被排入湾内,导致海水环境质量不断降低,水产品质量安全受到威胁[6-7]的同时,也对考洲洋内物种组成带来了负面影响[1]。此外,由于之前考洲洋内围网和地笼的大量投放,也会严重影响湾内渔业群落的分布和资源更新。针对考洲洋环境和渔业资源现状,自2014年起,有关部门对湾内违规养殖设施以及围网、拦网等网具进行了拆除清理,仅保留刺网作业,并启动红树林生态修复工程,有效地改善了考洲洋的海水环境质量[8]。目前关于考洲洋的研究主要集中在水质条件[9]、沉积物[10]、重金属污染[11]、有机污染物[12]等环境质量监控方面,而对于考洲洋海域的渔业资源状况仍无较为系统的认识。准确了解考洲洋的生态系统渔业资源状况,可为科学评价生态保护和修复工程的实际效果提供客观翔实的基础数据,因此有必要对考洲洋的渔业生物资源群落结构及其时空分布特征进行更全面的调查分析。
由于不同的网具对物种和个体的选择性往往存在一定差异,网具的选择对渔获资源结果也会造成一定影响[13]。不同网具对鱼类的长期选择作用也会影响鱼类的生活史特征,不当的选择范围会对渔业资源造成不可恢复的负面影响[14]。对于类似的封闭型内湾(如广东流沙湾),资源调查常使用定置张网或刺网等方式[15],考虑到考洲洋复杂的底质环境和水深变化,加之考洲洋内大量养殖筏架的存在,为保证调查准确性,本研究采用刺网和地笼2种方式,对当前考洲洋的自然渔业资源状况进行系统调查,分析其资源季节动态及空间分布,为考洲洋渔业资源的科学管理与生物多样性保护提供科学支撑,并为其他地理环境类似的内湾渔业资源调查提供参考。
1. 材料与方法
1.1 采样时间站点布设
考洲洋地处稔平半岛(114°52'E~114°56'E, 22°43'N~22°45'N),总面积44.7 km2,海岸线长65.3 km,通过一条狭长的水道与外界相连,最窄处仅253 m。考洲洋的海底较平坦,平均水深2.2 m,大部分水域水深0.3~1.0 m,受不规则半日潮影响,日平均潮差约为2.31 m。该地平均气温22.1 ℃,年平均雨量 1 944.3 mm,4—10 月为雨季,降雨量可占全年的 82.8%。考洲洋中部区域大部分被牡蛎吊养竹筏覆盖。
本研究于2017年11月和2018年1月、4月、7月,分秋、冬、春、夏四季分别对考洲洋渔业资源状况进行调查。由湾外至湾内,均匀布设5个站点(图1)。
1.2 样品采集与处理
本研究分别采用刺网、地笼方式采集渔获物样品,规格和设置具体为:
刺网每张高1.5 m、长35 m,分为3层,每层网目尺寸分别为42 mm、48 mm和52 mm。刺网放置时间为1 h,每个站点各放置4组重复,每个重复2张刺网。
地笼每个共5节,每节规格10 m,总长50 m。每个地笼网衣分为3层,每层网目均为55目。每个站点放置5个地笼作为重复,放置时间为一个潮水周期 (12 h)。
在每个站点同时使用便携式温度计、盐度计、pH计及溶氧仪对海水理化因子进行测量。采样完成后,对全部渔获物进行种类组成鉴定、分类,计数、称质量,并对渔获物进行生物学测定,包括其体长、全长、体质量、头胸甲长和宽等指标。采用单位捕捞努力量渔获量(catch per unit effort,CPUE)和单位捕捞努力量渔获个体数(number of catches per unit effort,NPUE)作为渔获资源量的指标,计算公式为:
$$ {\rm CPUE} = \frac{C}{{XT}} $$ (1) $$ {\rm NPUE} = \frac{N}{{XT}} $$ (2) 式中C为渔获量,N为捕捞个体数,X为网数,T为放网时间。
水样采集和分析按照《海洋监测规范》(GB/T 17378.4—2007)进行,动物样品处理、保存等按照《海洋调查规范》(GB/T 12763.4—2007)进行,物种所有个体鉴定到种,物种鉴定和生态特征参照《中国海洋鱼类》[16]检索。
1.3 数据处理
1.3.1 优势种
采用Pinkas等[17]提出的相对重要性指数(IRI)来确定群落的优势种。IRI指数计算公式为:
$$ {\rm IRI} = \left( {N + W} \right) \times F $$ (3) 其中N为某物种丰度占总丰度的百分比;W为某一种类的生物量占总生物量的比例;F为该物种出现的频率。
一般而言,IRI大于1 000为优势种;IRI大于100且小于1 000为重要种;IRI大于10且小于100为一般种;IRI小于10为稀有种。
1.3.2 群落生态多样性指数
分别采用Margalef物种丰富度指数(D)、Shannon-Wiener指数(H′)、Pielou均匀度指数(J′)来研究群落的多样性。为
Margalef丰富度[18]:
$$ D = \frac{{S - 1}}{{\ln N}} $$ (4) Shannon-Wiener指数[19]:
$$ H{\rm {'}} = - \sum\limits_{i = 1}^S {{P_i}{\rm ln}{P_i}} $$ (5) Pielou均匀度指数[20]:
$$ J{\rm{{'}}} = \frac{{H'}}{{\ln S}} $$ (6) 其中S为各站点物种种类总数,N为各站点捕捞个体数,Pi为某一种类生物量占总生物量的比例。
1.3.3 群落结构相似度
根据渔获生物量(kg·h–1)数据,经平方根转化后,得到Bray-Curtis相似性系数[21]矩阵,并根据矩阵使用非度量多维标度分析(NMDS)方法分析考洲洋各站点之间的群落结构差异。采用单因子相似性分析(ANOSIM)检验不同站点之间的显著性,并采用stress压力系数来衡量NMDS结果可信度,stress系数小于0.1说明排序较好;stress系数小于0.05说明图形具有很好的代表性[22]。采用Primer 7.0进行上述分析[23]。
1.3.4 粒径谱
水生生物粒径谱(size spectrum)模型是常用于反映水生生态系统生物量分布的模型。本文以经过以2为底的对数转换后的个体体质量数据作为个体尺寸特征,渔获生物量(kg·h–1)按照Sprules和Barth[24]的示例标准化。按对数转换后的个体尺寸分类聚合后,对个体尺寸(x轴)和生物量(y轴)作关系图,得出不同尺寸个体的生物量分布情况。
1.3.5 数据分析
采用双因素方差分析对资源量(CPUE、NPUE)和多样性指数进行显著性检验,自变量为季节、站点。P值显著性水平为0.05。使用LSD法进行事后检验。采用IBM SPSS 25.0软件进行上述分析。
2. 结果
2.1 理化环境因子
调查结果显示,考洲洋海域的理化环境指标季节变化较大(表1)。考洲洋调查水深为0.88~2.45 m,温度14.9~31.5 ℃,盐度8.93~32.0,pH 6.35~8.6。整体上,不同站点间的水深、盐度差异较大,不同季节间的温度、pH差异较大。
表 1 考洲洋四季各站点环境因子Table 1 Hydrological environmental variables in Kaozhouyang Bay during four seasons季节
season站点
station深度
depth温度
temperature盐度
salinitypH 春季
autumnS1 2.00 24.3 28.7 8.1 S2 1.96 24.6 28.5 8.0 S3 1.41 24.5 22.1 8.0 S4 1.60 24.0 19.1 8.2 S5 1.20 23.9 19.9 7.9 冬季
winterS1 0.90 17.1 26.6 6.9 S2 1.35 16.5 24.5 6.8 S3 1.17 15.5 20.6 6.9 S4 1.22 17.0 24.1 6.9 S5 0.76 14.9 18.5 6.9 春季
springS1 1.46 24.6 32.0 6.9 S2 1.92 24.5 31.1 6.7 S3 1.73 24.0 27.8 6.8 S4 2.06 24.7 31.0 6.6 S5 1.63 23.9 25.7 6.4 夏季
summerS1 0.88 31.5 22.5 8.3 S2 2.45 31.5 20.6 8.2 S3 1.41 31.5 13.7 8.2 S4 1.10 31.5 17.4 8.4 S5 1.10 30.5 8.9 8.6 2.2 种类组成及优势种
4个季度调查共记录到渔获物种类2门2纲13目36科93种,其中鱼类11目32科60种,占总物种数的64.52%;甲壳类2目6科33种,占总物种数的35.48% (表2)。其中鲈形目出现种类最多[42种(45.16%)];其次为十足目[29种(31.18%)]、鲱形目[7种(7.52%)]。鱼类物种中,绝大部分(96.7%)为暖水性鱼类。
表 2 考洲洋四季渔获游泳动物物种名录Table 2 List of swimming species caught in Kaozhouyang Bay during four seasons种名 Species 种名 Species 辐鳍鱼纲 Actinopterygii 新月锦鱼 Thalassoma lunare 鳗鲡目 Anguilliformes 弯棘䲗 Callionymus curvicornis 大鳍虫鳗 Muraenichthys macropterus 褐篮子鱼 Siganus fuscescens 海鲢目 Elopiformes 嵴塘鳢 Butis butis 海鲢 Elops saurus 犬牙缰鰕虎鱼 Amoya caninus 鲱形目 Clupeiformes 斑纹舌鰕虎鱼 Glossogobius olivaceus 圆吻海鰶 Nematalosa nasus 拟矛尾鰕虎鱼 Parachaeturichthys polynema 日本海鰶 Nematalosa japonica 孔鰕虎鱼 Trypauchen vagina 花鰶 Clupanodon thrissa 裸项纹缟鰕虎鱼 Tridentiger nudicervicus 黑尾小沙丁 Sardinella melanura 双斑舌鰕虎鱼 Psammogobius biocellatus 康氏小公鱼 Stolephorus commersonnii 尖鳍寡鳞鰕虎鱼 Oligolepis acutipinnis 汉氏棱鳀 Thryssa hamiltonii 眼瓣沟鰕虎鱼 Oxyurichthys ophthalmonema 鲇形目 Siluriformes 小鳞沟鰕虎鱼 Oxyurichthys microlepis 线纹鳗鲶 Plotosus lineatus 红狼牙鰕虎鱼 Odontamblyopus rubicundus 鲻形目 Mugiliformes 鲽形目 Pleuronectiformes 长鳍骨鲻 Osteomugil cunnesius 卵鳎 Solea ovata 棱鮻 Liza carinata 东方箬鳎 Brachirus orientalis 银汉鱼目 Atheriniformes 鲀形目 Tetraodontiformes 白氏银汉鱼 Hypoatherina valenciennei 纹腹叉鼻鲀 Arothron hispidus 颌针鱼目 Beloniformes 甲壳纲 Crustacea 日本下鱵鱼 Hyporhamphus sajori 十足目 Decapoda 鲉形目 Scorpaeniformes 近缘新对虾 Metapenaeus affinis 鲬 Platycephalus indicus 斑节对虾 Penaeus monodon 粗蜂鲉 Vespicula trachinoides 宽突赤虾 Metapenaeopsis mogiensis 鲈形目 Perciformes 中型新对虾 Metapenaeus intermedius 罗非鱼 Oreochromis sp. 刀额新对虾 Metapenaeus ensis 倒牙魣 Sphyraena putnamae 日本对虾 Marsupenaeus japonicus 眶棘双边鱼 Ambassis gymnocephalus 沙栖新对虾 Metapenaeus joyneri 日本花鲈 Lateolabrax japonicus 短沟对虾 Penaeus scmisulcatus 多鳞 Sillago sihama 墨吉对虾 Penaeus merguiensis 珍鲹 Caranx ignobilis 周氏新对虾 Metapenaeus joyneri 金带细鲹 Selaroides leptolepis 长毛对虾 Penaeus penicillatus 高体若鲹 Caranx equula 秀丽白虾 Palaemon modestus 勒氏枝鳔石首鱼 Dendrophysa russelii 脊尾白虾 Exopalaemon carinicauda 短吻鲾 Leiognathus brevirostris 日本沼虾 Macrobrachium nipponense 颈斑鲾 Nuchequula nuchalis 底栖短桨蟹 Thalamita prymna 短棘鲾 Leiognathus equulus 武士蟳 Charybdis miles 长吻银鲈 Gerres longirostris 日本蟳 Charybdis japonica 长棘银鲈 Gerres filamentosus 近亲蟳 Charybdis affinis 日本十棘银鲈 Gerres japonicus 锐齿蟳 Charybdis acuta 短棘银鲈 Gerres lucidus 拟穴青蟹 Scylla paramamosain 金焰笛鲷 Lutjanus fulviflamma 少刺短桨蟹 Thalamita danae 勒氏笛鲷 Lutjanus russellii 远海梭子蟹 Portunus pelagicus 黄鳍鲷 Acanthopagrus latus 双额短桨蟹 Thalamita sima 灰鳍棘鲷 Acanthopagrus berda 香港蟳 Charybdis hongkongensis 平鲷 Rhabdosargus sarba 变态蟳 Charybdis variegata 黑鲷 Acanthopagrus schlegelii 悦目大眼蟹 Macrophthalmus erato 胡椒鲷 Plectorhinchus pictus 强壮大眼蟹 Macrophthalmus crassipes 细鳞䱨 Terapon jarbua 四齿大额蟹 Metopograpsus quadridentatus 列牙䱨 Pelates quadrilineatus 字纹弓蟹 Varuna litterata 尖突吻䱨 Rhynchopelates oxyrhynchus 口足目 Stomatopoda 黑斑绯鲤 Upeneus tragula 断脊拟虾蛄 Oratosquillina interrupta 金钱鱼 Scatophagus argus 口虾蛄 Squilla orarotia 考洲洋四季优势种组成见表3,其中刺网四季优势种组成基本为鱼类,地笼四季优势种中也包含部分甲壳类。
表 3 考洲洋各季节优势种组成 (IRI > 1 000)Table 3 Dominant species composition in Kaozhouyang Bay in different season网具
gear秋季
autumn冬季
winter春季
spring夏季
summer种类
speciesIRI 种类
speciesIRI 种类
speciesIRI 种类
speciesIRI 地笼 cage 墨吉对虾 6 156.4 短吻鲾 4 706.7 黄鳍鲷 6 779.4 短吻鲾 4 706.7 底栖短桨蟹 2 281.2 长鳍骨鲻 2 648.0 沙栖新对虾 2 083.9 长鳍骨鲻 2 648.0 短吻鲾 1 312.4 黄斑鲾 2 066.1 黄斑鲾 2 066.1 眼瓣沟鰕虎鱼 1 008.3 刺网 gill net 圆吻海鰶 8 254.4 圆吻海鰶 5 680.9 花鰶 3 531.4 花鰶 4 508.4 花鰶 2 816.3 长鳍骨鲻 2 023.5 尖突吻䱨 2 006.4 褐篮子鱼 2 949.6 圆吻海鰶 1 854.7 墨吉对虾 1 620.6 少刺短桨蟹 1 385.7 黄鳍鲷 1 356.4 平鲷 1 191.2 优势种四季出现情况见表4。刺网渔获物中,秋季优势种为圆吻海鰶 (Nematalosa nasus)、花鰶(Clupanodon thrissa);冬季优势种为圆吻海鰶、长鳍骨鲻(Osteomugil cunnesius);春季优势种为花鰶、尖突吻䱨 (Rhynchopelates oxyrhynchus);夏季优势种为花鰶、褐篮子鱼(Siganus fuscescens)、圆吻海鰶、墨吉对虾(Banana prawn)、少刺短桨蟹(Thalamita danae)、黄鳍鲷(Acanthopagrus latus)、平鲷(Rhabdosargus sarba)等7种。
表 4 考洲洋四季优势种类出现情况Table 4 Occurrence of seasonal dominant species in Kaozhouyang Bay分层
layer种名
species秋季
autumn冬季
winter春季
spring夏季
summer刺网 gill net 墨吉对虾 √ √ 少刺短桨蟹 √ √ 花鰶 √ √ √ √ 圆吻海鰶 √ √ √ √ 长鳍骨鲻 √ √ √ √ 平鲷 √ 黄鳍鲷 √ √ √ √ 尖突吻䱨 √ √ 褐篮子鱼 √ √ √ √ 地笼 cage 墨吉对虾 √ √ √ √ 沙栖新对虾 √ √ 底栖短桨蟹 √ √ 短吻鲾 √ √ √ √ 黄斑鲾 √ √ √ 长鳍骨鲻 √ √ √ √ 黄鳍鲷 √ √ √ 眼瓣沟鰕虎鱼 √ √ √ √ 眼瓣沟鰕虎鱼 √ √ √ √ 地笼渔获物中,秋季优势种为墨吉对虾、底栖短桨蟹(T. prymna)、短吻鲾(Leiognathus brevirostris)、眼瓣沟鰕虎鱼(Oxyurichthys ophthalmonema)等4种;冬季为短吻鲾、长鳍骨鲻、黄斑鲾(Photopectoralis bindus)等3种;春季优势种为黄鳍鲷、沙栖新对虾(Metapenaeus joyneri);夏季优势种为短吻鲾、长鳍骨鲻、黄斑鲾。
2.3 资源量
资源量调查结果显示,基于刺网与地笼调查的CPUE皆表现出明显的季节性(图2)。夏季[地笼0.014 kg·(h·网)–1,刺网0.128 kg·(h·网)–1]>春季[地笼0.010 kg·(h·网)–1,刺网0.121 kg·(h·网)–1]>秋季[地笼0.007 kg·(h·网)–1,刺网0.091 kg·(h·网)–1]>冬季[地笼0.006 kg·(h·网)–1,刺网0.068 kg·(h·网)–1]。
![]() 图 2 考洲洋各站点间单位捕捞努力量渔获量和单位努力量渔获个体量Fig. 2 Catch per unit effort and number of catches per unit effort at different stations in four seasons
图 2 考洲洋各站点间单位捕捞努力量渔获量和单位努力量渔获个体量Fig. 2 Catch per unit effort and number of catches per unit effort at different stations in four seasons单位时间渔获个体数方面,刺网的季节趋势与渔获率相同,表现出夏季[平均4.5 个·(h·网)–1]>春季[平均2.8 个·(h·网)–1]>秋季[平均2.7 个·(h·网)–1]>冬季秋季[平均1.9 个·(h·网)–1]。地笼同样是夏季最高,平均达到1.96 个·(h·网)–1,其次是冬季[平均1.1 个·(h·网)–1]、春季[平均0.73 个·(h·网)–1]、秋季[平均0.73 个·(h·网)–1]。以季节、站点作自变量,采用双因素方差分析对资源量进行显著性检验。结果显示,季节和站点对CPUE、NPUE影响不显著。
2.4 群落结构相似性
各个季节刺网与地笼群落结构皆不存在站点之间的显著差异 (图3)。除去冬季底层的stress值为0.02外,其余NMDS的stress值皆为0,说明图形具有很好的代表性。因此可以认为,考洲洋各站点之间渔业资源群落结构无显著差异,可以看作一个整体进一步分析多样性变化。
![]() 图 3 考洲洋四季各站点群落结构NMDS分析Fig. 3 NMDS analysis of community structure in Kaozhouyang Bay in four seasons
图 3 考洲洋四季各站点群落结构NMDS分析Fig. 3 NMDS analysis of community structure in Kaozhouyang Bay in four seasons2.5 群落多样性指数
基于刺网的中上层调查中,群落多样性呈现出明显的季节动态。其中D以秋季最高(2.34),其次是冬季(2.20)、春季(1.49)、夏季(0.81);群落H′仍是秋季最高(1.90),其次是冬季(1.18),春季(0.81)与夏季(0.82)接近;J′,秋季、冬季、春季相近,分别为1.69、1.58、1.69,夏季最低(0.79)。
基于地笼的底层调查中,多样性指数变化与中上层略有差异。其中春、秋季的D较高,分别达到3.33、3.63;冬、夏季则分别为2.24、2.93;对于群落H′而言,四季差异较小,但仍以春(2.02)、秋季(2.03)较高,冬季(1.78)和夏季(1.52)较低;J′由秋季至夏季逐渐下降,分别为0.73、0.71、0.68、0.55。
采用双因子方差分析进行显著性检验,结果显示,不同季节、站点之间的D、H′、J′皆无显著性差异(P > 0.05,图4)。
![]() 图 4 基于刺网与地笼调查的多样性指数ns. 四季多样性无显著性差异(P>0.05)Fig. 4 Diversity indices of gill net and cagens. insignificant difference in four seasons (P>0.05)
图 4 基于刺网与地笼调查的多样性指数ns. 四季多样性无显著性差异(P>0.05)Fig. 4 Diversity indices of gill net and cagens. insignificant difference in four seasons (P>0.05)2.6 考洲洋群落粒径谱
考洲洋渔获个体的粒径谱基本呈现双峰的形态,第一峰介于0~8 g,第二峰介于8~64 g (图5)。以第一峰而言,夏季标准化生物量波峰高于其他三季,春、冬季相近,秋季第一峰最弱。各季节波峰个体尺寸基本一致,约为2 g。
以第二峰而言,夏季标准化生物量波峰仍高于其他三季,其次是冬、春、秋季。波峰所处的个体尺寸以冬季最高(32 g),其他三季约为16 g。
3. 讨论
3.1 考洲洋渔业资源种类组成及资源量
考洲洋是粤东地区重要的内洋,为周边惠东地区带来了大规模的水产养殖产业(如牡蛎养殖),也为大量物种提供了关键的生态栖息环境。本次调查的考洲洋内的物种组成以鱼类和甲壳类为主,其中绝大部分鱼类为暖水性鱼类,以鲈形目和鲱形目居多,物种种类与南海其他海域物种较接近[16,25]。考洲洋优势种基本集中在短吻鲾、花鰶、圆吻海鰶、墨吉对虾等物种间,大部分物种都生活在内湾、咸淡水、近海海域内,与考洲洋的海水环境一致。大部分渔获个体质量在100 g以内,鲜有超过500 g的个体。资源量上,考洲洋渔获的平均CPUE不超过0.15 kg·(h·网)–1, 平均个体数不超过5 ind·(h·网)–1,整体渔业资源量较低,仅靠野生渔获难以产生较高的经济价值。但是考洲洋的经济物种数达到96种,物种丰富度最高达到3.63,目前湾内总体的多样性处于较高水平,推测与周边大面积的红树林湿地有一定关系。考洲洋湾内各个站点间的资源量分布均匀,群落结构相似度高,推测原因是考洲洋类似的小型封闭式海湾受潮汐影响较大,因而整体资源分布趋于均匀。
3.2 季节动态
季节是渔业资源非常重要的影响因素之一。鱼类的季节性洄游会对物种组成和生物量造成直接影响[26];在春、夏产卵季节,一年生的小鱼(如考洲洋的鲾科鱼类)快速发育,对生物量也会有较大改变[27];此外,秋、冬季的藻类和浮游生物有限,对整个生态系统提供的能量输入也较低[28],会导致渔业资源量下降。而且,季节变化也与人类的捕捞强度相关(例如夏季的禁渔期),从而影响考洲洋的渔业资源量。
考洲洋的季节变化体现在资源量、优势种、多样性、个体尺寸分布等4个方面。资源量方面,刺网与地笼的趋势基本一致,夏季CPUE和平均渔获个体数都最高,其次是春季、秋季和冬季,这与长江口海域趋势一致[29]。优势种方面,除夏季刺网外,各季节的优势种基本为2~3种,其中秋季和夏季地笼群落以墨吉对虾等甲壳类为主要优势种,而春、冬季则以短吻鲾、长鳍骨鲻等鱼类为主要优势种;这与单秀娟等[30]的结果一致,黄海海域也以夏、秋季的甲壳类相对资源密度较高。群落多样性方面,春、秋两季的D和H′均高于夏、冬两季。其中冬季的H′和D与环境因子关系较大,冬季水温低,个体基本迁徙或停止产卵,生物量较低[31];而夏季的均匀度最低,因而推测与优势种生物量的爆发有关。生物粒径谱方面,四季的模式较为相似,基本可以按照个体大小分为两大类:1) 以短吻鲾为主的小型个体或幼鱼,集中在5 g以下;2) 以圆吻海鰶、底栖短桨蟹等为主的较大体型的成熟个体,集中在15~60 g。考洲洋夏季的生物量明显较高,因此2个波峰也高于其余三季。这与单秀娟等[32]构建的生物量谱相似。
3.3 网具差异对渔获量的影响
目前,渔业调查方式趋于多元化,除拖网、刺网、地笼、定置张网等传统作业方式外,还有声学探测设备等新兴监测技术,以及针对特定种类的方式(如鼓线用作捕鲨[33])。不同网具对种类和个体大小的选择性不同,调查结果也会存在差异[13],不同的网具和尺寸都会对结果造成一定影响,因此网具的选择对渔业资源调查的准确度十分重要。目前同时使用地笼和刺网进行比较调查的研究较少。徐胜南等[4]利用刺网和地笼分别对人工鱼礁附近的渔获组成、资源量、多样性进行了比较,发现刺网渔获组成主要为鱼类,而地笼的渔获组成则更加均衡。这与本文结果一致,刺网捕获的优势种基本为鱼类(如花鰶、圆吻海鰶),而地笼的优势种则包含数种甲壳类(如墨吉对虾、沙栖新对虾)。除渔获组成外,2种调查方式的资源量也有较大差异,刺网的CPUE和平均渔获个体数都要显著高于地笼,这与徐胜南等[4]调查得到刺网的生物量最高的结果也一致,同时在本次调查中也可能与刺网的调查时间较短有关。群落多样性方面,刺网和地笼的丰富度和多样性指数都没有显著差异,但刺网J′显著高于地笼,这可能因为地笼渔获的总体生物量虽然更高,但是优势种占了大部分,而2种调查方式皆能反映出考洲洋的多样性情况。
相同的网具,不同的尺寸对结果也有影响。国内外多个研究对不同尺寸、结构的刺网作比对,发现网目尺寸影响较大,不同网目的刺网之间的渔获种类、个体数都有差异,但多样性差异较小[34-37]。影响网具对鱼类选择性的因素复杂,除去网目尺寸外,还包括网线缠绕厚度、悬挂率等因素[38]。因此,在开展渔业资源调查以及制定渔获政策时,要充分考虑网具选择及网具结构、尺寸的影响,高选择性的网具可能会降低调查结果的准确度,而过小的网目尺寸则不利于当地渔业的长期发展[33]。
3.4 渔业资源生物量单位及多样性指数比较
在渔业资源调查中,一般以多样性指数作为描述群落结构多样性的参数,而生物量和个体数都可以用作计算多样性指数[39]。陈国宝等[39]的研究显示2种计算方式的结果显著相关,但是部分研究则认为两者无显著相关[36,40]。虽然这2种计算方式的关系很可能与特定的区域环境有关,但各海域间比较多样性高低时,可能因此产生误解。以距考洲洋较近的大亚湾海域为例,孙宝权等[41]计算的H′多样性指数为0.09~2.57,平均1.65;而王雪辉等[25]的结果为2.40~3.82,平均3.15,两文都以2004—2005年的底拖网数据为基础,但结果差异较大,很可能由计算方式不同导致。同理,物种丰富度也会受到影响。以珠江口水域的研究为例,晏磊等[36]的D为3.60~6.73,而袁梦等[42]的D为0~1.93,两者差异较大。虽然结果如上文所讨论,多样性结果和作业方式有一定关系,但是无论是由网具差异还是计算方式导致,结果都不利于海域间的比较;且在以生物量为计算单位时,生物量的标准化也存在不同。谢旭等[43]使用kg·h–1的方式来描述渔获率或生物量大小,有的研究[30]使用kg·m–2或kg·km–2,孙鹏飞等[29]使用CPUE,不利于对不同海域之间的生物量、多样性进行比较。笔者认为,对于刺网、地笼等被动式捕获且不便于计算面积的调查方式,可以使用kg·h–1或CPUE作为资源量单位;对于底拖网、双拖网等主动式的调查方式,则可以使用kg·m–2或kg·km–2作为资源量单位。
4. 结论
基于刺网、地笼2种网具的调查,本文首次对考洲洋进行了四季的渔业资源调查分析。结果显示,考洲洋总体渔业资源量较低,呈现出夏、春高,秋、冬低的趋势,物种丰富度、群落多样性、群落均匀度也存在较大的季节波动,且湾内资源总体空间分布较均匀。本文还通过对比刺网与地笼2种网具的结果,分析了网具选择对渔业资源调查准确度和制定长期渔获管理政策的影响。未来的研究应明确计算方式、统一计量单位,以便于不同海域之间的比较。
-
![]()
图 1 不同化学试剂处理后生物膜附着基质吸附的芽孢杆菌数量
A,B,C,D,E,F和G分别代表 4%氢氧化钠,0.1%氢氧化钙,10%盐酸,5%高锰酸钾,5%重铬酸钾,75% 乙醇和蒸馏水;不同的字母代表不同处理间有显著性差异(P < 0.05),后图同此
Figure 1. Amount of Bacillus spp. attached to substrate with different chemical reagents
A, B, C, D, E, F and G represent 4% sodium hydroxide, 0.1% calcium hydroxide, 10% diluted hydrochloric acid, 5% potassium permanganate, 5% potassium dichromate and 75% ethyl alcohol and distillated water, respectively.Different letters represent significant difference among different treatments (P < 0.05).The same case in the following figures.
![]()
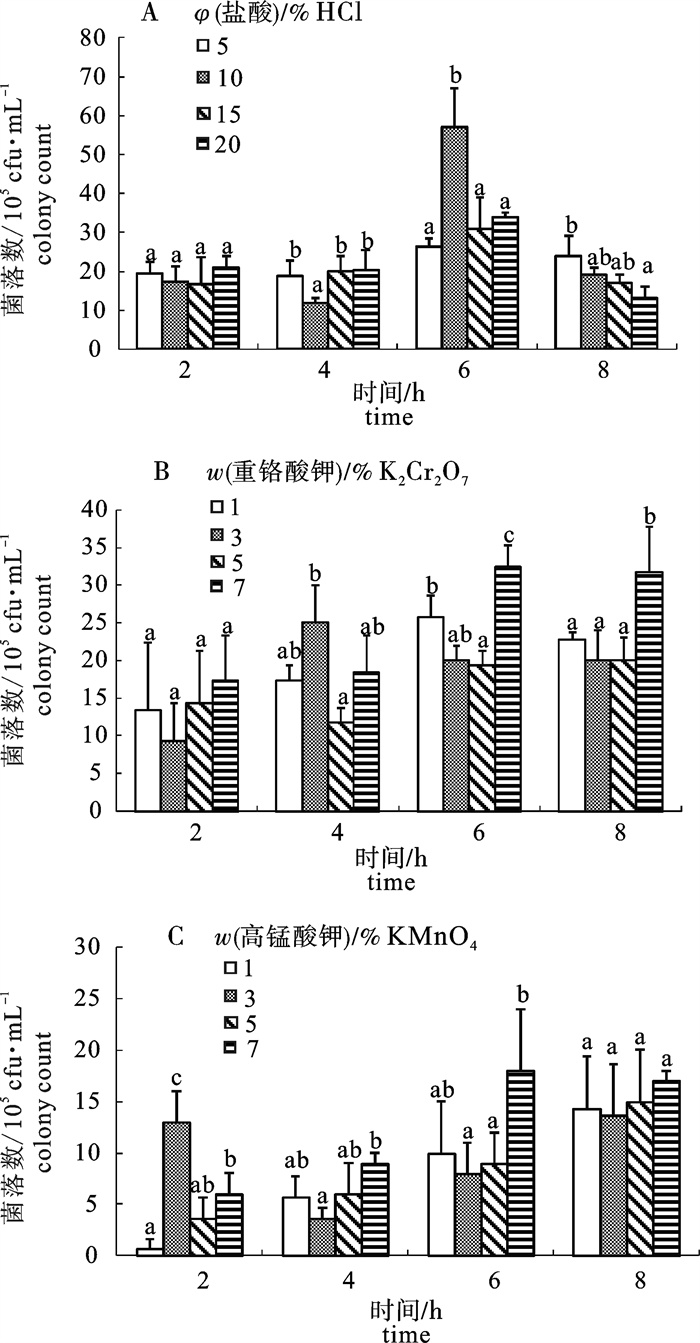
图 2 基质经不同浓度盐酸(A)、重铬酸钾(B)和高锰酸钾(C)处理不同时间后吸附细菌数
Figure 2. Bacterial amount attached to substrate treated with different immerse time and levels of diluted hydrochloric acid(A), potassium dichromate(B) and potassium permanganate (C)
![]()
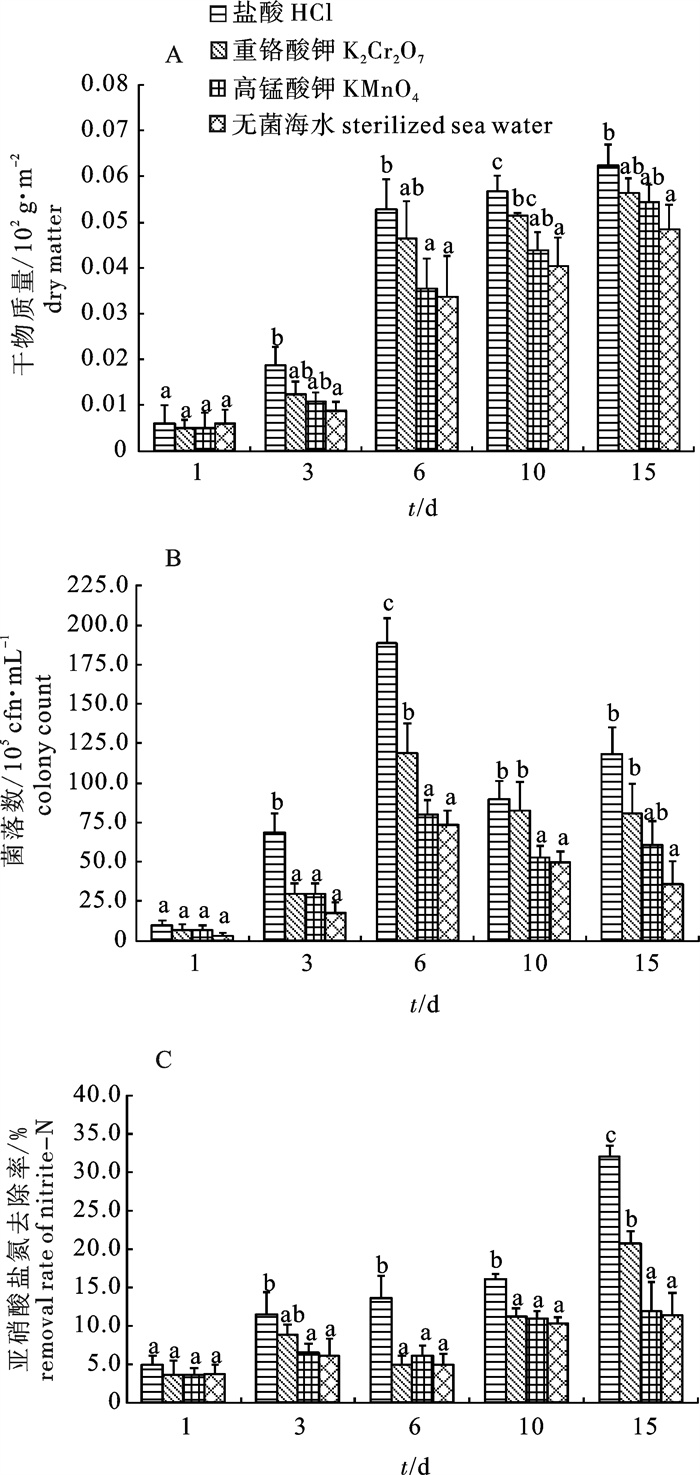
图 3 挂膜阶段各处理组基质表面干物质(A)、吸附异养细菌数(B)和对养殖废水中亚硝酸盐氮去除率(C)的变化
Figure 3. Variation of dry matter (A), total heterotrophic bacteria amount (B) and removal rate of nitrite-N in aquaculture wastewater (C) on substrate in different treatments during membrane formation period
-
[1] 宋志文, 王玮, 赵丙辰, 等. 海水养殖废水的生物处理技术研究进展[J]. 青岛理工大学学报, 2006, 27(1): 13-17. doi: 10.3969/j.issn.1673-4602.2006.01.003 [2] 刘朝阳, 孙晓庆. 生物控制法在水产养殖水质净化中的综合应用[J]. 南方水产, 2007, 3(1): 69-74. doi: 10.3969/j.issn.2095-0780.2007.01.012 [3] 陈金玲, 赖秋明, 苏树叶, 等. 精养虾池主要生态因子变化特点与相关性分析[J]. 南方水产科学, 2012, 8(4): 49-56. doi: 10.3969/j.issn.2095-0780.2012.04.008 [4] 宋玉芝, 秦伯强, 高光. 附着生物对富营养化水体氮磷的去除效果[J]. 长江流域资源与环境, 2009, 18(2): 180-185. doi: 10.3969/j.issn.1004-8227.2009.02.015 [5] STEWART N T, BOARDMAN G D, HELFRICH L A. Treatment of rainbow trout (Oncorhynchus mykiss) raceway effluent using baffled sedimentation and artificial substrates[J]. Aquac Eng, 2006, 35(2): 166-178. doi: 10.1016/j.aquaeng.2006.01.001
[6] ZHANG B. Influence of the artificial substrates on the attachment behavior of Litopenaeus vannamei in the intensive culture condition[J]. Int J Anim Vet Adv, 2011, 3(1): 37-43. https://www.researchgate.net/publication/49615903_Influence_of_the_Artificial_Substrates_on_the_Attachment_Behavior_of_Litopenaeus_vannamei_in_the_Intensive_Culture_Condition
[7] 林武, 陈敏, 罗建中, 等. 生态工程技术治理污染水体的研究进展[J]. 广东化工, 2008, 35(4): 42-46. doi: 10.3969/j.issn.1007-1865.2008.04.015 [8] 司友斌, 彭军. 固定化微生物技术及其在污染土壤修复中的应用[J]. 土壤, 2007, 39(5): 673-676. doi: 10.3321/j.issn:0253-9829.2007.05.001 [9] MA F, GUO J, ZHAO L, et al. Application of bioaugmentation to improve the activated sludge system into the contact oxidation system treating petrochemical wastewater[J]. Bioresour Technol, 2009, 100(2): 597-602. doi: 10.1016/j.biortech.2008.06.066
[10] 马立群. 难粘高分子材料的表面处理技术[J]. 化学与粘合, 1999(1): 23-26. doi: 10.3969/j.issn.1001-0017.1999.01.011 [11] 董高峰. 难粘塑料的表面处理[J]. 有机氟工业, 2005(3): 26-28. doi: 10.3969/j.issn.1001-0017.2002.04.017 [12] 李卓佳, 林亮, 杨莺莺, 等. 芽孢杆菌制剂对虾池环境微生物群落的影响[J]. 农业环境科学学报, 2007, 26(3): 1183-1189. doi: 10.3321/j.issn:1672-2043.2007.03.075 [13] WAHL M. Marine epibiosis. I. Fouling and antifouling: some basic aspects[J]. Mar Ecol Prog Ser, 1989, 58(1): 175- 189. doi: 10.3354/meps058175
[14] AZIM M E, RAHAMAN M M, WAHAB M A, et al. Periphyton-based pond polyculture system: a bioeconomic comparison of on-farm and on-station trials[J]. Aquaculture, 2004, 242(1): 381-396. doi: 10.1016/j.aquaculture.2004.09.008
[15] EBELING J M, TIMMONS M B, BISOGNI J J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems[J]. Aquaculture, 2006, 257(1): 346-358. doi: 10.1016/j.aquaculture.2006.03.019
[16] OTOSHI C A, MONTGOMERY A D, MATSUDA E M, et al. Effects of artificial substrate and water source on growth of juvenile pacific white shrimp, Litopenaeus vannamei[J]. J World Aquac Soc, 2006, 37(2): 210-213. doi: 10.1111/j.1749-7345.2006.00029.x
[17] THOMPSON F L, ABREU P C, WASIELESKY W. Importance of biofilm for water quality and nourishment in intensive shrimp culture[J]. Aquaculture, 2002, 203(3): 263-278. doi: 10.1016/S0044-8486(01)00642-1
[18] VIAU V E, OSTERA J M, TOLIVIA A, et al. Contribution of biofilm to water quality, survival and growth of juveniles of the freshwater crayfish Cherax quadricarinatus(Decapoda, Parastacidae)[J]. Aquaculture, 2012, 324/325: 70-78. doi: 10.1016/J.AQUACULTURE.2011.10.009
[19] 范福洲, 康勇. 水处理用纤维素基载体的降解性能研究[J]. 高分子材料科学与工程, 2006, 22(6): 126-129. doi: 10.3321/j.issn:1000-7555.2006.06.032 [20] 金冬霞, 田刚, 施汉昌. 悬浮填料的选取及其性能试验研究[J]. 环境科学学报, 2002, 22(3): 333-337. doi: 10.3321/j.issn:0253-2468.2002.03.013 [21] 马兆昆, 刘杰. 改性碳纤维及表面含氧官能团对反硝化菌固着化的影响[J]. 功能材料, 2003, 34(5): 592-594. https://xueshu.baidu.com/usercenter/paper/show?paperid=718862f9d7edd03be61f69b2842b1b4e&site=xueshu_se&hitarticle=1 [22] 丰民义, 方涛, 吴娟, 等. 不同载体的藻-菌生物膜应用于水体净化[J]. 水处理技术, 2007, 33(3): 59-61. [23] 薛松松. 纳米生态基在水产养殖中的应用研究[D]. 青岛: 中国海洋大学, 2011. 10.7666/d.y1926840 [24] 刘灿灿, 金吴云, 沈耀良. 上流式曝气生物滤池两种填料启动挂膜的试验研究[J]. 江苏环境科技, 2008, 21(1): 61-64. doi: 10.3969/j.issn.1674-4829.2008.01.018 -
期刊类型引用(34)
1. 李珊,高延奇,赵立宁,王莎,陈善真. 丁香酚和2-苯氧乙醇对不同品系罗非鱼的麻醉效果比较. 水产学杂志. 2025(02): 64-73 .  百度学术
百度学术
2. 宋霖,陈小江,高鹏. 常温下三卡因和丁香酚对条纹小鲃的麻醉效果研究. 水产科学. 2024(01): 144-151 . 百度学术
3. 吴冰,董秀金,徐一力,许建和. 丁香酚慢性麻醉对红鲫血液生理及肝脏hsp70和hsp90mRNA表达量的影响. 河北渔业. 2024(05): 17-21 . 百度学术
4. 董秀金,徐一力,吴冰,许建和. 鱼类麻醉保活运输研究进展. 河北渔业. 2024(06): 41-46 . 百度学术
5. 谢希尧,区又君,徐淑玉,林欣,李加儿,肖裕杰. MS-222和丁香酚胁迫对四指马鲅幼鱼鳃及肝脏组织微观结构的影响. 广东农业科学. 2024(05): 144-154 . 百度学术
6. 黄琼叶,张建明,朱友芳,许秋龙. 丁香酚对细点石斑鱼麻醉效果及肝脏生理生化指标的影响. 河北渔业. 2024(10): 24-29 . 百度学术
7. 周烨,骆小年,段友健,李姣,罗来幸,曾雪,刘佳欣. 2种麻醉剂对马口鱼幼鱼麻醉效果研究. 水产科学. 2024(06): 965-973 . 百度学术
8. 吕曼,董义超,李蒙,于会国,郭建东,石英. MS-222对墨瑞鳕麻醉效果及血清生化指标的影响. 浙江海洋大学学报(自然科学版). 2024(04): 352-358 . 百度学术
9. 谢希尧,区又君,徐淑玉,林欣,李加儿,肖裕杰. 丁香酚对四指马鲅幼鱼的麻醉效果及抗氧化影响. 中国渔业质量与标准. 2024(04): 1-13 . 百度学术
10. 马凯,朱晓磊,张澜澜,卫明亮,赵成,何宝全,尹家胜,张永泉. 黑龙江茴鱼对几种常用消毒药物和麻醉剂的耐受性. 水产学杂志. 2024(06): 38-44 . 百度学术
11. 唐忠林,张佳佳,周国勤,陈树桥,徐钢春,徐跑,强俊,王佩佩. 丁香酚对“优鲈3号”幼鱼运输水质及其血液、肌肉生理指标的影响. 水产科技情报. 2023(01): 44-52 . 百度学术
12. 韦政坤,董宏标,赵文,陈飞,张传镶,陈健,宫保华,朱长波,张家松. 两种麻醉剂对凡纳滨对虾麻醉效果及组织氧化损伤的影响. 南方水产科学. 2023(01): 136-146 . 本站查看
13. 胡望娇,王翠华,冯广朋,庄平,郑跃平,季强. 丁香酚和MS-222对松江鲈的麻醉效果. 海洋渔业. 2023(02): 181-190 . 百度学术
14. 林晨,吴敏,王凯,曾三妹,吴佳雯,钟莉萍,凌淞江. QuEChERS-超高压液相色谱-串联质谱法同时测定水产品中的6种丁香酚类麻醉剂. 食品科学. 2023(08): 324-329 . 百度学术
15. 章霞,徐志进,李伟业,殷小龙,王易帆,陈爽,马雪彬. MS-222对大黄鱼幼鱼麻醉效果、组织结构及抗氧化酶活性的影响. 大连海洋大学学报. 2023(02): 267-274 . 百度学术
16. 陈军平,武慧慧,沈方方,张佳鑫,于若梦,付永杰,赵道全,谢国强,孙文喜. MS-222与丁香酚对黄尾鲴麻醉效果的比较. 湖北农业科学. 2023(08): 133-139 . 百度学术
17. 陈欣怡,王好学,徐开达,李鹏飞,周永东. 丁香酚对褐菖鲉的麻醉效果. 广东海洋大学学报. 2023(06): 146-152 . 百度学术
18. 李娇妮,廖秀睿,李曦,吕布,杨守国,臧战,VASQUEZ Herbert Ely,王爱民,郑兴,顾志峰. 运输方式对马氏珠母贝(Pinctada fucata)稚贝存活率、生长、消化及抗氧化性能的影响探究. 海洋与湖沼. 2022(02): 414-420 . 百度学术
19. 戴晓娜,陆亦宽,蔡杨杨,孙虹,卢瑛,谢晶. 基于磁性纳米探针的渔用麻醉剂三卡因的快速检测方法. 食品与发酵工业. 2022(08): 245-253 . 百度学术
20. 朱晓玲,张菊,刘杰,韩智,吴婉琴,张莉,王会霞,江丰. 湖北省水产品兽药残留状况分析. 食品安全质量检测学报. 2021(01): 69-77 . 百度学术
21. 谢晶,曹杰. 渔用麻醉剂在鱼类麻醉保活运输中应用的研究进展. 上海海洋大学学报. 2021(01): 189-196 . 百度学术
22. 胡发文,王晓龙,高凤祥,李莉,菅玉霞,王雪,樊英,潘雷,郭文. 温度、盐度和两种麻醉剂对大泷六线鱼幼鱼耗氧率、排氨率的影响. 海洋科学. 2021(01): 54-61 . 百度学术
23. 刘阳,贾贵芳,杨飞,李忠利. 麻醉剂MS-222对运输胁迫黄颡鱼抗氧化系统的影响. 安徽农学通报. 2021(05): 67-69 . 百度学术
24. 陈旭,左涛,周胜杰,杨蕊,于刚,秦传新,马振华. 尖吻鲈幼鱼运输密度和时长对运输水质及其复苏率和抗氧化能力的影响. 南方水产科学. 2021(02): 122-128 . 本站查看
25. 曹杰,王琪,梅俊,谢晶. 有水与无水保活运输对大菱鲆生理应激及鱼肉品质的影响. 水产学报. 2021(07): 1034-1042 . 百度学术
26. 王维鑫,崔爱君,徐永江,李海宁,柳学周,王滨,姜燕,刘新富. MS-222和丁香油对黄条(鱼师)麻醉效果研究. 渔业科学进展. 2021(06): 205-214 . 百度学术
27. 曹杰,谢晶. 间氨基苯甲酸乙酯甲磺酸盐在大菱鲆模拟保活运输中的作用效果. 食品科学. 2021(23): 246-253 . 百度学术
28. 王金林,王万良,张忭忭,谭大明. MS-222和丁香酚对亚东鲑鱼麻醉效果的比较. 甘肃农业大学学报. 2021(06): 26-32+40 . 百度学术
29. 管敏,张德志,饶军,唐大明. 温度对中华鲟(Acipenser sinensis)幼鱼麻醉效果的影响. 生态学杂志. 2020(02): 576-585 . 百度学术
30. 朱之发,边力,刘心田,常青,阮晓红,陈四清,李凤辉,葛建龙,刘长琳. 12种麻醉剂对长蛸的麻醉效果. 中国水产科学. 2020(02): 195-203 . 百度学术
31. 黄小林,杨洁,戴超,李涛,杨育凯,黄忠,虞为,林黑着,孙莘溢,舒琥. 丁香酚对紫红笛鲷幼鱼的麻醉效果. 中国渔业质量与标准. 2020(02): 51-58 . 百度学术
32. 陈小江,熊俐灵,吴建顾,亓鲁,王权. MS-222和丁香酚对四川华吸鳅耗氧率和排氨率的影响. 南方水产科学. 2020(04): 69-74 . 本站查看
33. 王文豪,董宏标,孙彩云,段亚飞,李华,刘青松,张家松,曾祥兵. 石菖蒲挥发油和水溶性氮酮对鱼用麻醉剂的促皮渗透效果研究. 南方水产科学. 2020(04): 62-68 . 本站查看
34. 杨洁,朱晓玲. 丁香酚在水产品中的残留及风险评估研究进展. 食品安全质量检测学报. 2020(18): 6523-6529 . 百度学术
其他类型引用(7)

 下载:
下载:













计量
- 文章访问数: 3265
- HTML全文浏览量: 131
- PDF下载量: 1736
- 被引次数: 41
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
