
Influence of aquaculture on characteristics of phytoplankton community in Liusha Bay
-
摘要:
于2012年8月至2013年1月在流沙湾海区进行了3个季度的浮游植物调查。共检出浮游植物80种,包括硅藻门34属69种,甲藻门5属9种,蓝藻门2属2种,主要优势种为中肋骨条藻(Skeletonema costatum)、菱形海线藻(Thalassionema nitzschioides)和中心圆筛藻(Coscinodiscus centralis)等。浮游植物的细胞丰度为0.001×104~1.495×104个· L-1),夏季最高冬季最低且内港丰度普遍低于外港。扇贝主养区、网箱鱼类养殖区、珍珠贝养殖区的多样性指数普遍高于对照区(无养殖区),且网箱鱼类养殖区最高。夏、秋两季扇贝主养区丰度相对较高,网箱鱼类养殖区浮游植物细胞丰度则与对照区相近,珍珠贝养殖区丰度夏季较高(1.099×104个· L-1)而秋季较低(0.021×104 个· L-1)。结果表明,近年来流沙湾海区浮游植物种类及丰度锐减;扇贝养殖可提高浮游植物丰度,鱼类网箱养殖则反之;密集的网箱设施阻碍了湾口水流交换,减少了营养物质的运输并限制了内港珍珠贝养殖区浮游植物丰度。
Abstract:We studied the characteristics of phytoplankton community in Liusha Bay for three quarters from August, 2012 to January, 2013. A total of 80 phytoplankton species were recorded, with 69 species of diatom belonging to 34 genera, followed by dinoflagellates represented by 9 taxa (5 genera) and blue-green algaes by 2 taxa (2 genera). The most common dominant species were Skeletonema costatum, Thalassionema nitzschioides and Coscinodiscus centralis. The range of phytoplankton density was 0.001×104~1.495×104 cells · L-1 with maximum density in summer, followed by autumn and winter. In general, the phytoplankton density decreased from outer bay to inner bay. Compared with the control (non-cultured area), the diversity indices in the cultured areas were higher, especially in the fish cultured area. In summer and autumn, phytoplankton density was higher in scallop culture area, while lower in fish cage and non-cultured area. However, phytoplankton density in pearl oyster cultured area was higher in summer (1.099×104 cells ·L-1) and low in autumn (0.021×104 cells · L-1). The results indicate that both phytoplankton taxa and density declined steeply in recent years. The phytoplankton density increased and decreased by scallop culture and fish cage culture, respectively. Dense fish cages blocked the current exchange, diminishing the transportation of nutrients across the bay and the phytoplankton density in pearl oyster cultured area of Liusha Bay.
-
Keywords:
- Liusha Bay /
- phytoplankton /
- scallops /
- fish cage /
- principal components analysis
-
流沙湾(22.36°~20.50°N,109.80°~110.02° E)位于广东省湛江市雷州半岛西南部,为西北向呈葫芦形的半封闭海湾,面积约69 km2,是中国著名的海水珍珠育苗与产珠基地[1]。流沙湾拥有丰富的海藻资源,海草床面积约900 hm2,为整个海湾的氮(N)、磷(P)吸收及溶氧增加起到重要作用,使流沙湾在较长时间内能够维持高密度的珍珠贝养殖[2]。
然而,近年来流沙湾超容量养殖产生的负面效益日渐凸显。依据流沙湾马氏珠母贝(Pinctada martensi)养殖的常规密度(1.05×105只· hm-2),其适养面积为461.83 hm2,但2008年调查显示其吊养面积为1 250 hm2,超过养殖容量的1.7倍[1, 3]。超容量的马氏珠母贝养殖对浮游植物群落形成较大压力,马氏珠母贝养殖成活率、珍珠产量及品质均受到严重影响,从业人员纷纷转向网箱鱼类养殖和墨西哥湾扇贝(Argopecten irradians concentricus)养殖等见效快的养殖领域[4]。到2012年,流沙湾水域马氏珠母贝养殖面积已降至272 hm2,但墨西哥湾扇贝养殖面积却达1 132 hm2;网箱养殖面积仅为67.5 hm2,数量却已达到4×104只以上,密布湾口的航道处。
浮游植物对养殖水域中有机物同化和营养物质的移除起着关键作用,是浮游动物、贝类等生物的重要饵料来源[5-6]。浮游植物的群落组成、丰度变化、空间分布等间接反映水体环境的动态变化,是养殖水体健康程度的指标之一[7]。2009年之前针对流沙湾海域展开的浮游植物调查较为密集[2, 8-9],此后则鲜见相关报道。当前扇贝养殖数量已远超珍珠贝养殖,网箱养殖侵占湾口,在这种新的养殖格局下对流沙湾的鱼贝养殖区展开浮游植物的针对性调查,可为流沙湾扇贝养殖业的可持续发展和南珠产业的振兴提供一定的参考依据。
1. 材料与方法
1.1 采样站位点的设定
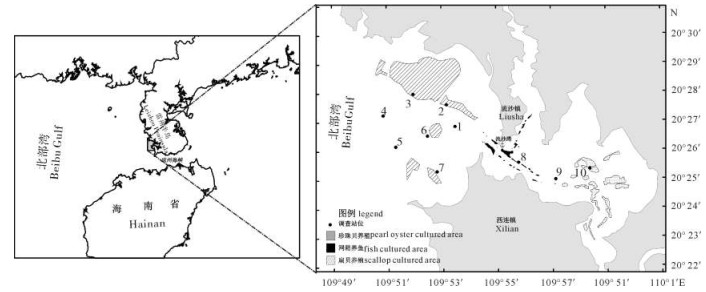
分别于2012年8月(夏季)、11月(秋季)及2013年1月(冬季)对流沙湾内港、外港进行季度性采样调查。采样站位点如图 1所示,数字1~10代表采样站位。
1#~7#站位为外港采样点,包括邻近扇贝(即墨西哥湾扇贝)养殖区(2#、3#、6#和7#),对照区(4#和5#),以及外港最邻近湾口的1#站位,其中3#站位于扇贝主养区。8#~10#站位属于内港采样区,其中8#站位于湾口航道内狭长的网箱(鱼排)养殖区;10#站位于内港深处的珍珠贝(即马氏珠母贝)养殖区。
1.2 浮游植物的采集与分析
浮游植物样品的采集与定量参照《海洋调查规范GB 17378.7-2007》[10]。定性样品采用浅水Ⅲ型浮游生物网自海底向海面垂直拖曳,定量样品则用有机玻璃采水器收集表层及底层的水样(各1 L)进行混合,现场经质量分数为5%的甲醛溶液固定后,带回实验室后用浮游生物计数框镜检及计数。
1.3 环境因子的采集与分析
现场测定采用溶氧仪(YSI-556型)实时监测海水中溶解氧(DO)、水温(T)、盐度(S)和pH,采用萨式盘法测定透明度(SD)。
实验室测定营养盐水样经-20 ℃暂存后,迅速带回实验室进行分析。其中亚硝酸盐(NO2-N)采用萘乙二胺分光光度法,氨氮(NH4-N)采用靛酚蓝法,硝酸盐(NO3-N)采用锌(Zn)-镉(Cr)还原法,活性磷酸盐(PO4-P)采用磷钼蓝分光光度法测定,总氮(TN)和总磷(TP)采用过硫酸钾氧化法测定。以上方法均按照《海洋监测规范GB 17378.4-2007》[11]指定方法进行。叶绿素a(Chl-a)及颗粒有机物(POM)的测定方法分别为丙酮萃取法[12]和灼烧称重法[13]。
1.4 多样性分析方法
多样性指数(Shannon and Weaver)[14]
$$ H^{\prime}=-\sum\limits_{i=1}^s P_i \log _2 P_i $$ (1) 均匀度(Pielou)[15]
$$ J=\frac{H^{\prime}}{\log _2 S} $$ (2) 优势度[16]
$$ Y=\left(\frac{n_i}{N}\right) \cdot f_i $$ (3) 式中s为物种数,Pi为第i物种在全部采样中的比例,ni为第i种的个体数,N为所有种的个体总数,S为样品种类总数,fi为第i种在各站点的监测频率。Y≥0.02视为优势种。
1.5 数据处理和分析
采用SPSS 18.0软件进行数据统计与方差分析,以P<0.05作为差异显著性水平。图表采用Origin 8.5绘制,环境因子与浮游植物群落结构多样性的关系的冗余分析(RDA)分析采用CANOCO 4.5软件,相关数据经过log10(x+1)转换。
2. 结果
2.1 浮游植物的种类组成及优势种
流沙湾浮游植物种类组成见表 1。3个航次共鉴定出浮游植物3门41属,共80种。其中硅藻为优势种群(34属,69种),占总数的86.25%;甲藻5属9种;蓝藻2属2种。从季节变化来看,冬季最多(56种),夏、秋两季种类均为45种。统计各季出现频次≥3,且至少在某一个站位的相对丰度>5%的浮游植物物种用CANOCO软件筛选,符合条件的浮游植物共计22种,均属硅藻门,随机编号为Bac01~Bac22(表 1)。
表 1 流沙湾浮游植物种类名录Table 1. Catalogue of phytoplankton species in Liusha Bay种类 species 编码 code 种类 species 编码 code 硅藻门 Bacillariophyceae 短角弯角藻 Eucampia zoodiacus 薄壁几内亚藻 Guinardia flaccida 环纹劳德藻 Lauderia annulata 中华半管藻 Hemiaulus sinensis Bac20 菱形藻 Nitzschia sp. Bac09 半盘藻 Hemidiscus sp. 洛氏菱形藻 N.lorenziana 波罗的海布纹藻 Gyrosigma balticum 柔弱菱形藻 N.debilis 布纹藻 Gyrosigma sp. 新月菱形藻 N.closterium 尖布纹藻 G.acuminatum 长菱形藻 N.longissima 斜布纹藻 G.obliquum 长菱形藻弯端变种 N.longissima var. Bac19 并基角刺藻 Chaetoceros decipiens Bac02 马鞍藻 Campylodiscus sp. 笔尖形根管藻 Rhizosolenia styliformis Bac01 海洋环毛藻 Corethron pelagicum 粗根管藻 R.robusta 尖刺菱形藻 Nitzschia pungens Bac07 刚毛根管藻 R.setigera Bac06 螺旋链鞘藻 Streptotheca thamensis Bac10 斯托根管藻 R.stolterfothii 太阳漂流藻 Planktoniella sol 中肋骨条藻 Skeletonema costatum Bac21 布氏双尾藻 Ditylum brightwellii Bac03 掌状冠盖藻 Stephanopyxis palmeriana 筒柱藻 Cylindrotheca gracilis 奇异棍形藻 Bacillaria paradoxa Bac14 丹麦细柱藻 Leptocylindrus danicus 密联海链藻 Detonula pumila 翼根管藻 Proboscia alata 佛氏海毛藻 Thalassionema frauenfeldii Bac05 小环藻 Cyclotella sp. Bac15 菱形海线藻 T.nitzschioides Bac08 小桩藻 Characium sp. 圆海链藻 Thalassiosira rotula 楔形藻 Licmophora sp. Bac16 钝角盒形藻 Biddulphia obtuse 海洋斜纹藻 Pleurosigma pelagicum 高盒形藻 B.regia 诺马斜纹藻 P.normanii 活动盒形藻 B.mobiliensis 日本星杆藻 Asterionella japonica 颗粒盒形藻 B.granulata 扭鞘藻 Helicotheca tamesis 长耳盒形藻 B.aurita 圆盘藻 Pulvinularia sp. 中华盒形藻 B.sinensis 圆筛藻 Coscinodiscus sp. 茧形藻 Amphiprora sp. 整齐圆筛藻 C.concinnus 紧密角管藻 Cerataulina compacta 中心圆筛藻 C.centralis Bac22 艾氏角毛藻 Chaetoceros eibenii 舟形藻 Navicula sp. 变异辐杆藻 Bacteriastrum varians 甲藻门 Pyrrophyta 短孢角毛藻 Chaetoceros brevis 叉角藻 Ceratium furca 角毛藻 Chaetoceros sp. 大角角藻 C.macroceros 洛氏角毛藻 C.lorenzianus 纺锤梭角藻 C.fusus 冕孢角毛藻 C.subsecundus Bac11 三角角藻 C.tripos 拟旋链角毛藻 C.pseudocurvisetus Bac12 具尾鳍藻 Dinophysis caudata 扭链角毛藻 C.tortissimus Bac13 鳍藻 Dinophysis sp. 无沟角毛藻 C.holsaticus 具刺膝沟藻 Gonyaulax spinifera 细齿角毛藻 C.denticulatus 海洋原多甲藻 Protoperidinium oceanicum 发状角毛藻 C.crinitus Bac04 蓝藻门 Cyanophyta 旋链角毛藻 C.curvisetus Bac17 颤藻 Oscillatoria sp. 圆柱角毛藻 C.teres Bac18 巨大色球藻 Chroococcus gigateus 远距角毛藻 C.distans 调查期间的浮游植物优势种共计14属22种(均为硅藻),且冬季优势种数稍高于夏、秋季。整体而言,各站Y呈现随季节推移逐渐下降,且呈内港普遍高于外港的规律。中肋骨条藻、菱形海线藻、奇异棍形藻和中心圆筛藻在3个季节中均为优势种。其中中肋骨条藻的Y明显高于其他优势种,夏、秋、冬季的平均Y分别为0.460(0.190~0.698)、0.345(0.070~0.684)和0.125(0.012~0.314),夏、冬两季在10个站位出现的频次均超过8。菱形海线藻夏、秋和冬季的平均Y依次为0.050(0.005~0.300)、0.082(0.017~0.230)和0.009(0.014~0.230),仅在外港的个别站位成为优势种。
2.2 主成分及冗余分析
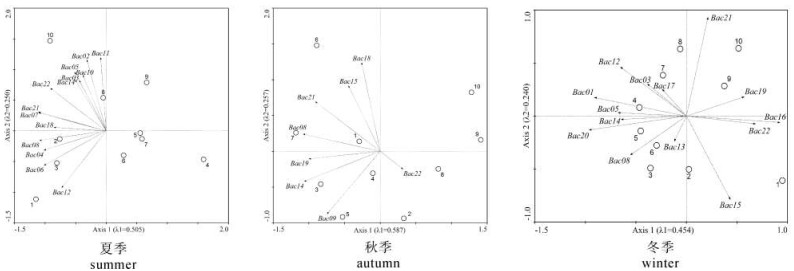
先对物种数据进行去趋势对应分析(detrended correspondence analysis,DCA),结果显示夏、秋和冬季的最长排序轴的轴长分别为2.153(第一排序轴)、1.442(第二排序轴)和1.539(第一排序轴),轴长均<3,适合线性模型。在DCA分析的基础上,应用主成分分析(principal components analysis,PCA)分析浮游植物分布特征(图 2)。排序结果显示,整体而言,内港的8#~10#站位在浮游植物物种分布上特征相似,且与外港区分明显(秋季最为显著)。具体表现为夏季浮游植物物种全部分布于第二、第三象限,以10#站位居多,1#站位次之,而在外港的对照区(4#和5#站位)及非扇贝主养区(6#和7#站位)分布较少;秋季浮游植物物种也主要分布于第二、三象限,且6#站位分布最多,内港的8#~10#站点则分布较少;与夏季和秋季相比,冬季的浮游植物物种在各站点的分布更为均匀,在各个象限内均有分布,但在9#和10#两站所在的第一象限内分布较少。
![]() 图 2 流沙湾浮游植物样方的主成分分析排序图Figure 2. PCA ordination of phytoplankton community composition and samples in Liusha Bay
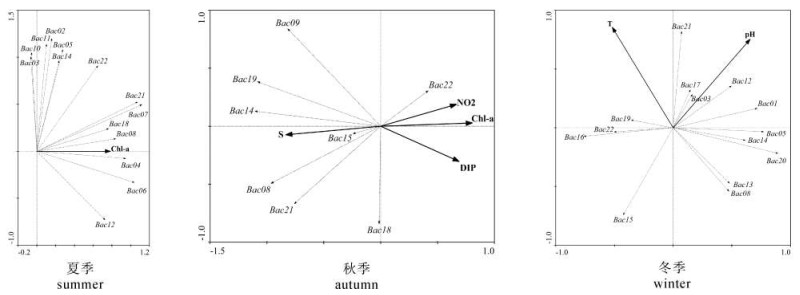
图 2 流沙湾浮游植物样方的主成分分析排序图Figure 2. PCA ordination of phytoplankton community composition and samples in Liusha Bay调查期间流沙湾的环境因子均值见表 2。在冗余分析(redundancy analysis,RDA)(图 3)中,利用向前引入法(forward selection)对所有站位的各环境因子进行逐步筛选,Monte Carlo置换检验结果显示,夏季叶绿素a(F=4.834,P=0.008),秋季盐度(F=7.977,P=0.002)、叶绿素a(F=6.977,P=0.002)、溶解性PO4-P(F=4.637,P=0.01)和NO2-N(F=4.027,P=0.012),冬季pH(F=2.960,P=0.014)和水温(F=2.657,P=0.03),对流沙湾浮游植物分布的影响达到显著水平,而其他环境因子的影响达不到显著水平(P>0.05)。以上数据表明叶绿素a为夏季影响流沙湾浮游植物分布的关键因子,对浮游植物物种变化的解释程度高达37.7%;夏季盐度和叶绿素a对浮游植物物种变化的解释程度分别为49.9%和46.6%,除此二者外,NO2-N和溶解性PO4-P同为夏季的关键环境因子;冬季则以物理指标pH和水温为主要影响因子,但对物种变化的解释程度较低(依次为27.0%和24.9%)。
表 2 夏季至冬季流沙湾主要水质参数Table 2. Main parameters for water quality in Liusha Bay from summer to winter季节
season溶解氧/mg·L-1
DO透明度/m
SD可溶性无机磷/mg·L-1
DIP氨氮/mg·L-1
NH4亚硝酸盐/mg·L-1
NO2硝酸盐/mg·L-1
NO3总磷/mg·L-1
TP总氮/mg·L-1
TN叶绿素a/μg·L-1
Chl-a颗粒有机物/mg·L-1
POM盐度
S温度/℃
T酸碱度
pH夏季 summer 6.650 1.550 0.013 0.025 0.012 0.656 0.039 2.255 7.325 2.640 30.764 30.73 8.38 秋季 autumn 6.813 1.180 0.014 0.098 0.002 0.220 0.085 2.721 3.960 6.180 30.900 26.26 8.50 冬季 winter 7.230 1.410 0.015 0.027 0.009 0.424 0.100 2.061 0.775 4.480 32.131 19.84 8.44 ![]() 图 3 流沙湾浮游植物与环境因子冗余分析排序图Figure 3. RDA ordination of phytoplankton community composition and environmental factors in Liusha Bay
图 3 流沙湾浮游植物与环境因子冗余分析排序图Figure 3. RDA ordination of phytoplankton community composition and environmental factors in Liusha Bay2.3 典型养殖区浮游植物丰度比较
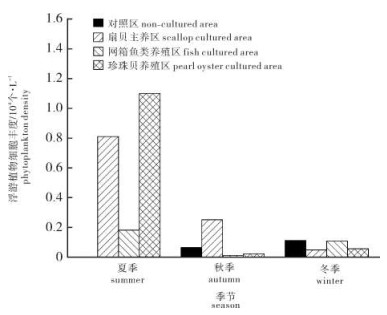
浮游植物丰度随季节推移而降低,10个采样站位的浮游植物丰度范围分别为夏季0.001×104~1.495×104个· L-1(均值0.419×104个· L-1),秋季0.010×104~0.576×104个· L-1(均值0.143×104个· L-1),冬季0.012×104~0.111×104个· L-1(均值0.065×104个· L-1)。图 4比较了对照区(4#)、扇贝主养区(3#)、网箱鱼类养殖区(8#)和珍珠贝养殖区(10#)的浮游植物丰度。结果显示,夏季所有养殖区的浮游植物丰度均高于对照区,扇贝主养区、珍珠贝养殖区高于网箱鱼类养殖区;秋、冬两季浮游植物丰度整体较低,但秋季扇贝主养区浮游植物丰度明显高于其他典型区域。
![]() 图 4 流沙湾典型养殖区浮游植物丰度Figure 4. Phytoplankton density in typical culture areas in Liusha Bay
图 4 流沙湾典型养殖区浮游植物丰度Figure 4. Phytoplankton density in typical culture areas in Liusha Bay2.4 典型养殖区H′与J
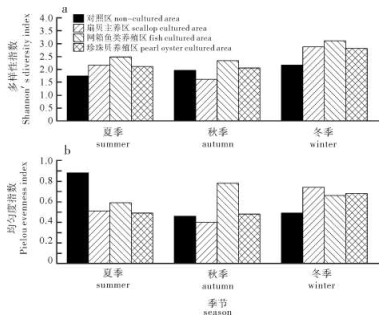
10个采样站位的浮游植物H′分别为夏季1.704~3.740,秋季0.364~2.691,冬季2.167~3.309,而J分别为夏季0.447~0.876,秋季0.091~0.784,冬季0.486~0.827。H′平均值冬季(2.734)>夏季(2.407)>秋季(2.022),J平均值冬季(0.631)>秋季(0.623)>夏季(0.508)。
图 5显示,各典型养殖区的浮游植物H′高于对照区,且网箱鱼类养殖区的指数值最高。J的组间变化规律与H′变化规律相近。夏季对照区的J高于典型养殖区,秋冬两季则反之。
![]() 图 5 流沙湾典型养殖区浮游植物多样性指数(a)和均匀度指数(b)Figure 5. Diversity index (a) and evenness index (b) of phytoplankton in typical culture areas
图 5 流沙湾典型养殖区浮游植物多样性指数(a)和均匀度指数(b)Figure 5. Diversity index (a) and evenness index (b) of phytoplankton in typical culture areas3. 讨论
3.1 浮游植物的群落结构特征
流沙湾浮游植物群落的季节性变化较为明显,细胞丰度夏季较高,秋冬两季较低,其均值从夏季的0.0419×104个· L-1降至冬季的0.065×104个·L-1;冬季的多样性指数和均匀度指数则整体表现为冬季高于夏秋两季。PCA分析显示,内港、外港的浮游植物种类具有明显的空间分布差异,尤其在秋季。近年来流沙湾浮游植物种数呈逐年下降的趋势,2012年~2013年的3次调查检出的浮游植物仅80种,约为2008年前进行的2次调查的一半,其中硅藻在海区浮游植物种类中占绝对优势(表 3)。相比其他全年性的优势种(菱形海线藻、奇异棍形藻、中心圆筛藻),中肋骨条藻显示出更高的出现频次及细胞丰度,此结果与邻近海区的调查结果相一致。2010年夏季的浮游植物调查[17]显示,雷州半岛西南部海域中肋骨条藻的Y高达0.152,丰度达10.78×104个· L-1。与以往调查数据相比,浮游植物丰度(0.001×104~1.495×104个· L-1)降低了1~2个数量级。根据贾晓平等[18]对饵料生物水平等级的划分,流沙湾浮游植物处于低水平(<20×104个· L-1)。
表 3 近年来流沙湾浮游植物群落特征比较Table 3. Comparison of characteristics of phytoplankton community in Liusha Bay in recent years水域
waters采样时间
sampling date硅藻种数/总种数
diatom species/total species丰度/104个·L-1
density多样性指数
H′文献来源
reference流沙湾(外港)Liusha Bay (out bay) 2006~2007 119/159 1.51~90.3 1.11~4.09 程小倪等[8] 流沙湾 Liusha Bay 2008 123/151 0.24~5.72 1.07~3.35 张才学等[2] 流沙湾(内港)Liusha Bay (inner bay) 2008~2009 69/69 3.2~7.0 2.82~2.95 王大鹏等[19] 流沙湾 Liusha Bay 2012~2013 69/80 0.001~1.495 1.62~3.74 该调查 3.2 浮游植物群落结构与环境因子的关系
RDA分析显示叶绿素a为影响夏季流沙湾浮游植物分布的关键因子(正相关),但与秋冬两季的浮游植物丰度的相关性较差。叶绿素a与浮游植物丰度通常有良好的线性关系[20],但这种线性相关特征只有在浮游植物丰度较高时才较为明显,水温、盐度、营养盐及浮游动物摄食等均会影响其关联性,甚至出现负相关趋势[21]。因此,在秋冬两季较低的浮游植物丰度下,二者的相关性远不及夏季明显。
盐度是影响藻类体内渗透压调节能力的重要因素,对盐度调节能力的大小反映出藻类适应的盐度范围并限制了藻类的生长分布[22-23]。RDA分析显示,盐度是影响秋季浮游植物群落的最主要环境因子(正相关)。流沙湾海域平均盐度依次为冬季(32.28)>秋季(31.05)>夏季(30.76)。尽管调查显示的秋季盐度较高,但采样前的第23号强台风“山神”(2012年10月底)登陆造成粤西海域6~8级大风,降雨量超过100 m m,极易造成海水表面的盐度骤降。多项研究表明,中肋骨条藻、菱形海线藻和小环藻等广盐性的浮游植物在低盐环境下有更好的生长表现[24-25]。因此,由于浮游植物对水环境改变的响应的滞后性[26],暴雨引起的盐度骤降对浮游植物的影响可能延后至秋季采样中体现。
与夏秋两季相比,冬季影响浮游植物丰度的环境因子较多,除pH外,叶绿素a、颗粒有机物与浮游植物丰度的相关系数也相对较高。此外,冬季微型浮游动物(<200 μm)对浮游植物生长的控制效应尤为明显:冬季南海北部海域微型浮游动物的摄食压力占浮游植物现存量的42.6%~82.8%[27];北部湾北部海域夏季微型浮游动物对浮游植物现存量的摄食压力达28.1%~66.0%[28]。
营养盐的可获得性对浮游植物丰度产生关键性影响[29-30],尤其是N/P对浮游植物生长有一定的限制作用,N/P>16时浮游植物表现为P限制,<16则表现为N限制[31]。营养加富试验[32]表明,N元素限制的形成是由优势硅藻对N元素的较高需求和N限制的外海水对湾口区产生的显著影响共同决定的。中肋骨条藻的生长主要为氮限制,在N/P>16时的生长速度优于<16[33]。2012~2013年夏季、秋季和冬季的N/P分别为79.9、21.65和32.2,均表现为明显的P限制。因此,N/P的明显升高可能会制约某些种类浮游植物的生长,导致浮游植物种类减少、丰度降低,同时增强中肋骨条藻等硅藻的竞争优势。
3.3 现有养殖格局对浮游植物分布特征的影响
与深圳湾、三亚湾等富营养化海域相比,流沙湾属于典型的贫营养性海湾[2],浮游植物种数和丰度都较低,但多样性指数相对较高(均值2.44),群落结构尚为稳定。与对照区相比,鱼贝养殖区的多样性指数、均匀度指数通常较高,且网箱鱼类养殖区的多样性指数一直为各区最高。
3个季度的调查共检出9种甲藻,夏季(叉角藻、具尾鳍藻、鳍藻等)主要分布在6#站位;秋冬两季(叉角藻、大角角藻、纺锤梭角藻、三角角藻等)则主要分布于对照区(4#和5#)和扇贝非主养区(2#、6#和7#),少量分布于内港(9#和10#)。扇贝主养区和网箱鱼类养殖区甲藻均未检出。张莉红等[34-36]报道,栉孔扇贝(Chlamys farreri)对东海原甲藻的摄食率和摄食选择效率高于中肋骨条藻;扇贝生活过的海水可显著提高中肋骨条藻生长竞争能力,而对东海原甲藻产生一定程度的抑制作用。流沙湾扇贝的养殖周期多为4~6月,夏初投苗秋季收获,扇贝的生理活动可能与栉孔扇贝相似地对浮游植物种类有一定的限制作用,并使骨条藻、圆筛藻、海线藻等常见的小型硅藻优势增强。
养殖设施(网箱、筏架等)对海区水流有明显的阻碍作用[37-38],在一定程度上影响营养盐的分布并对浮游生物分布造成影响。根据2008年的数据模拟结果,流沙湾潮流运动为带有旋转流的往复流运动,落潮流速大于涨潮流速且余速较强;可溶性无机氮(DIN)和可溶性无机磷(DIP)交换率可达97%,海水自净能力很强[39]。然而,2012年进行的水动力建模数据[40]显示,流沙湾港口内、外水体流速差异显著,涨潮时外港水流速度为0.5~1 m · s-1,港口约为0.5 m · s-1,珍珠贝养殖区附近<0.3 m · s-1;退潮时整个内港(包括湾口)的水流速度比涨潮时更低,趋于静止状态。整体而言,外港水流交换相对顺畅,浮游生物可获得的营养物质较多,其丰度普遍高于内港,因此对扇贝养殖区贝类生长的影响也小于对珍珠贝养殖区。夏季珍珠贝养殖区的浮游植物丰度相对较好,但秋季明显低于外港(仅为0.021×104个· L-1)。每年的春季(3月~5月)、秋季(9月~11月)是马氏珠母贝的快速生长期[41],珍珠贝生长所需的饵料生物在秋季得不到快速补充会制约其充分生长。
当海区养殖的滤食性贝类生物量较高,尤其是养殖中后期(8月以后)接近养殖容量时,浮游植物丰度及生物量受到显著的控制作用[42-43]。同时,贝类向水体排泄铵盐等无机营养盐以及粪便、假粪等有机物,沉降后的有机物经矿化作用和再悬浮作用又可重新进入物质循环,为浮游植物生长提供营养[44-45]。据报道,由于养殖筏体对水流的阻碍作用,胶州湾扇贝养殖区的化学耗氧量(COD)、营养盐浓度梯度向区外的延伸不足1 km,对养殖区N贡献率大于60%,对P的贡献率则超过80%[46-47]。对照区、扇贝主养区、网箱鱼类养殖区和珍珠贝养殖区的夏季N/P依次为52、425、56和146,秋季N/P依次为18、97、50和80,冬季N/P依次为73、38、32和45。3个季度各典型区N/P最高区的浮游植物丰度往往最高:夏秋两季扇贝主养区N/P最高,丰度也为各组最高;冬季对照区的N/P较高,丰度也相对较高。扇贝主养区的养殖设施分布最为密集,水体交换能力较差,养殖污染物得不到充分扩散[48],N/P偏高,因此该区中肋骨条藻等硅藻的细胞丰度相应较高。
鱼类网箱养殖增加了水体生态系统中营养物质的输入,对浮游植物的种类组成和丰度变化会产生重要影响[49]。鱼类网箱养殖区N/P略高于对照区,营养盐浓度也与之接近,浮游植物多样性指数较高而丰度却相对较低,可能是由于过于密集的网箱鱼排设施对光照产生较强的遮挡作用,并对浮游植物光合作用形成负面影响。该研究结果与黄长江等[50]的报道相似,柘林湾位于湾口的网箱渔排养殖区叶绿素a浓度普遍低于其他调查站点。
-
![]()
图 2 流沙湾浮游植物样方的主成分分析排序图
Figure 2. PCA ordination of phytoplankton community composition and samples in Liusha Bay
![]()
图 3 流沙湾浮游植物与环境因子冗余分析排序图
Figure 3. RDA ordination of phytoplankton community composition and environmental factors in Liusha Bay
![]()
图 4 流沙湾典型养殖区浮游植物丰度
Figure 4. Phytoplankton density in typical culture areas in Liusha Bay
![]()
图 5 流沙湾典型养殖区浮游植物多样性指数(a)和均匀度指数(b)
Figure 5. Diversity index (a) and evenness index (b) of phytoplankton in typical culture areas
表 1 流沙湾浮游植物种类名录
Table 1 Catalogue of phytoplankton species in Liusha Bay
种类 species 编码 code 种类 species 编码 code 硅藻门 Bacillariophyceae 短角弯角藻 Eucampia zoodiacus 薄壁几内亚藻 Guinardia flaccida 环纹劳德藻 Lauderia annulata 中华半管藻 Hemiaulus sinensis Bac20 菱形藻 Nitzschia sp. Bac09 半盘藻 Hemidiscus sp. 洛氏菱形藻 N.lorenziana 波罗的海布纹藻 Gyrosigma balticum 柔弱菱形藻 N.debilis 布纹藻 Gyrosigma sp. 新月菱形藻 N.closterium 尖布纹藻 G.acuminatum 长菱形藻 N.longissima 斜布纹藻 G.obliquum 长菱形藻弯端变种 N.longissima var. Bac19 并基角刺藻 Chaetoceros decipiens Bac02 马鞍藻 Campylodiscus sp. 笔尖形根管藻 Rhizosolenia styliformis Bac01 海洋环毛藻 Corethron pelagicum 粗根管藻 R.robusta 尖刺菱形藻 Nitzschia pungens Bac07 刚毛根管藻 R.setigera Bac06 螺旋链鞘藻 Streptotheca thamensis Bac10 斯托根管藻 R.stolterfothii 太阳漂流藻 Planktoniella sol 中肋骨条藻 Skeletonema costatum Bac21 布氏双尾藻 Ditylum brightwellii Bac03 掌状冠盖藻 Stephanopyxis palmeriana 筒柱藻 Cylindrotheca gracilis 奇异棍形藻 Bacillaria paradoxa Bac14 丹麦细柱藻 Leptocylindrus danicus 密联海链藻 Detonula pumila 翼根管藻 Proboscia alata 佛氏海毛藻 Thalassionema frauenfeldii Bac05 小环藻 Cyclotella sp. Bac15 菱形海线藻 T.nitzschioides Bac08 小桩藻 Characium sp. 圆海链藻 Thalassiosira rotula 楔形藻 Licmophora sp. Bac16 钝角盒形藻 Biddulphia obtuse 海洋斜纹藻 Pleurosigma pelagicum 高盒形藻 B.regia 诺马斜纹藻 P.normanii 活动盒形藻 B.mobiliensis 日本星杆藻 Asterionella japonica 颗粒盒形藻 B.granulata 扭鞘藻 Helicotheca tamesis 长耳盒形藻 B.aurita 圆盘藻 Pulvinularia sp. 中华盒形藻 B.sinensis 圆筛藻 Coscinodiscus sp. 茧形藻 Amphiprora sp. 整齐圆筛藻 C.concinnus 紧密角管藻 Cerataulina compacta 中心圆筛藻 C.centralis Bac22 艾氏角毛藻 Chaetoceros eibenii 舟形藻 Navicula sp. 变异辐杆藻 Bacteriastrum varians 甲藻门 Pyrrophyta 短孢角毛藻 Chaetoceros brevis 叉角藻 Ceratium furca 角毛藻 Chaetoceros sp. 大角角藻 C.macroceros 洛氏角毛藻 C.lorenzianus 纺锤梭角藻 C.fusus 冕孢角毛藻 C.subsecundus Bac11 三角角藻 C.tripos 拟旋链角毛藻 C.pseudocurvisetus Bac12 具尾鳍藻 Dinophysis caudata 扭链角毛藻 C.tortissimus Bac13 鳍藻 Dinophysis sp. 无沟角毛藻 C.holsaticus 具刺膝沟藻 Gonyaulax spinifera 细齿角毛藻 C.denticulatus 海洋原多甲藻 Protoperidinium oceanicum 发状角毛藻 C.crinitus Bac04 蓝藻门 Cyanophyta 旋链角毛藻 C.curvisetus Bac17 颤藻 Oscillatoria sp. 圆柱角毛藻 C.teres Bac18 巨大色球藻 Chroococcus gigateus 远距角毛藻 C.distans  下载: 导出CSV
下载: 导出CSV
表 2 夏季至冬季流沙湾主要水质参数
Table 2 Main parameters for water quality in Liusha Bay from summer to winter
季节
season溶解氧/mg·L-1
DO透明度/m
SD可溶性无机磷/mg·L-1
DIP氨氮/mg·L-1
NH4亚硝酸盐/mg·L-1
NO2硝酸盐/mg·L-1
NO3总磷/mg·L-1
TP总氮/mg·L-1
TN叶绿素a/μg·L-1
Chl-a颗粒有机物/mg·L-1
POM盐度
S温度/℃
T酸碱度
pH夏季 summer 6.650 1.550 0.013 0.025 0.012 0.656 0.039 2.255 7.325 2.640 30.764 30.73 8.38 秋季 autumn 6.813 1.180 0.014 0.098 0.002 0.220 0.085 2.721 3.960 6.180 30.900 26.26 8.50 冬季 winter 7.230 1.410 0.015 0.027 0.009 0.424 0.100 2.061 0.775 4.480 32.131 19.84 8.44
下载: 导出CSV
表 3 近年来流沙湾浮游植物群落特征比较
Table 3 Comparison of characteristics of phytoplankton community in Liusha Bay in recent years
水域
waters采样时间
sampling date硅藻种数/总种数
diatom species/total species丰度/104个·L-1
density多样性指数
H′文献来源
reference流沙湾(外港)Liusha Bay (out bay) 2006~2007 119/159 1.51~90.3 1.11~4.09 程小倪等[8] 流沙湾 Liusha Bay 2008 123/151 0.24~5.72 1.07~3.35 张才学等[2] 流沙湾(内港)Liusha Bay (inner bay) 2008~2009 69/69 3.2~7.0 2.82~2.95 王大鹏等[19] 流沙湾 Liusha Bay 2012~2013 69/80 0.001~1.495 1.62~3.74 该调查
下载: 导出CSV
-
[1] 申玉春, 李再亮, 黄石成, 等. 流沙湾海域水产养殖结构与布局调查分析[J]. 中国渔业经济, 2010, 28(1): 105-109. doi: 10.3969/j.issn.1009-590X.2010.01.020 [2] 张才学, 陈慧妍, 孙省利, 等. 流沙湾海草床海域浮游植物的时空分布及其影响因素[J]. 生态学报, 2012, 32(5): 1527-1537. doi: 10.5846/stxb201101110053 [3] 朱春华, 申玉春, 谢恩义, 等. 湛江流沙湾马氏珠母贝的养殖容量[J]. 热带海洋学报, 2011, 30(3): 76-81. doi: 10.3969/j.issn.1009-5470.2011.03.012 [4] 周昌仕, 慕永通. 我国南珠产业的困境及其治理研究[J]. 中国海洋大学学报: 社会科学版, 2012 (4): 49-54. doi: 10.3969/j.issn.1672-335X.2012.04.009 [5] SILVA C, YÁÑEZ E, MARTÍN-DÍAZ M L, et al. Assessing a bioremediation strategy in a shallow coastal system affected by a fish farm culture: application of GIS and shellfish dynamic models in the Rio San Pedro, SW Spain[J]. Mar Pollut Bull, 2012, 64(4): 751-765. doi: 10.1016/j.marpolbul.2012.01.019
[6] ALONGI D M, MCKINNON A D, BRINKMAN R, et al. The fate of organic matter derived from small-scale fish cage aquaculture in coastal waters of Sulawesi and Sumatra, Indonesia[J]. Aquaculture, 2009, 295(1): 60-75. doi: 10.1016/j.aquaculture.2009.06.025
[7] SUN C C, WANG Y S, SUN S, et al. Dynamic analysis of phytoplankton community characteristics in Daya Bay, China[J]. Acta Ecol Sin, 2006, 26(12): 3948-3958. doi: 10.1016/S1872-2032(07)60005-5
[8] 程小倪, 黄良民, 谭烨辉, 等. 流沙湾海域浮游植物群落结构的时空变化[J]. 海洋环境科学, 2011, 30(1): 13-18. doi: 10.3969/j.issn.1007-6336.2011.01.003 [9] 王彦, 申玉春, 叶宁, 等. 流沙湾浮游生物的群落结构与时空分布[J]. 广东海洋大学学报, 2013, 32(6): 66-73. [10] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.7-2007海洋监测规范[S]. 北京: 中国标准出版社, 2008: 3-10. https://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=9C8314F2EADA090A59433F0740B0F438 [11] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.4-2007海洋监测规范[S]. 北京: 中国标准出版社, 2008: 57-121. https://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=9FB14D0EE23D77A96D54A9BDAAF6EA07 [12] 陈宇炜, 高锡云. 浮游植物叶绿素a含量测定方法的比较测定[J]. 湖泊科学, 2000, 12(2): 185-188. doi: 10.3321/j.issn:1003-5427.2000.02.015 [13] 方志山. 综合水产养殖池悬浮颗粒物的动态变化研究[J]. 厦门大学学报: 自然科学版, 2007, 46(1): 131-134. doi: 10.3321/j.issn:0438-0479.2007.01.029 [14] SHANNON C E, WEAVER W. The mathematical theory of communication[M]. London: University of Illionis Press, 1963: 125. doi: 10.1063/1.3067010
[15] PIELOU E C. An introduction to mathematical ecology[M]. New York: Wiley-Interscience, 1966: 1-286. doi: 10.1007/978-3-642-69888-0
[16] 孙军, 刘东艳, 徐俊, 等. 1999年春季渤海中部及其邻近海域的网采浮游植物群落[J]. 生态学报, 2004, 24(9): 2003-2016. doi: 10.3321/j.issn:1000-0933.2004.09.024 [17] 龚玉艳, 张才学, 孙省利, 等. 2010年夏季雷州半岛海岸带浮游植物群落结构特征及其与主要环境因子的关系[J]. 生态学报, 2012, 32(19): 5972-5985. doi: 10.5846/stxb201109011280 [18] 贾晓平, 杜飞雁, 林钦, 等. 海洋渔场生态环境质量状况综合评价方法探讨[J]. 中国水产科学, 2003, 10(2): 160-164. doi: 10.3321/j.issn:1005-8737.2003.02.015 [19] 王大鹏, 曹占旺, 张益峰, 等. 两个珍珠养殖海区浮游植物物种特征及其比较[J]. 广西水产科技, 2010 (2): 16-23. https://xueshu.baidu.com/usercenter/paper/show?paperid=ad16c31fc230f7b56b5576f9c3c5bc96&site=xueshu_se [20] TILZER M M. Secchi disk: chlorophyll relationships in a lake with highly variable phytoplankton biomass[J]. Hydrobiologia, 1988, 162(2): 163-171. doi: 10.1007/BF00014539
[21] 王雨, 林茂, 陈兴群, 等. 海南西部近岸浮游植物的周年变化及主要关联因素[J]. 水生生物学报, 2012, 36(4): 724-734. doi: 10.3724/SP.J.1035.2012.00724 [22] 罗民波, 陆健健, 王云龙, 等. 东海浮游植物数量分布与优势种[J]. 生态学报, 2008, 27(12): 5076-5085. doi: 10.3321/j.issn:1000-0933.2007.12.016 [23] ABID O, SELLAMI-KAMMOUN A, AYADI H, et al. Biochemical adaptation of phytoplankton to salinity and nutrient gradients in a coastal solar saltern, Tunisia[J]. Est Coast Shelf Sci, 2008, 80(3): 391-400. doi: 10.1016/j.ecss.2008.09.007
[24] 王珺, 赖秋明, 姚发壮, 等. 人工培养条件下环境因子对微小小环藻生长的影响[J]. 海洋渔业, 2013, 35(2): 195-201. doi: 10.3969/j.issn.1004-2490.2013.02.011 [25] 霍文毅, 俞志明, 邹景忠, 等. 胶州湾中肋骨条藻赤潮与环境因子的关系[J]. 海洋与湖沼, 2001, 32(3): 311-318. doi: 10.3321/j.issn:0029-814X.2001.03.012 [26] 孟顺龙, 陈家长, 胡庚东, 等. 太湖蠡湖浮游植物群落特征及其对水质的评价[J]. 长江流域资源与环境, 2010, 19 (1): 30-36. https://yangtzebasin.whlib.ac.cn/CN/abstract/abstract9473.shtml [27] 郑丽平, 相卫国, 黄邦钦. 冬季南海北部海域微型浮游动物及其对浮游植物摄食压力研究[J]. 台湾海峡, 2012, 31(1): 72-78. doi: 10.3969/ISSN.1000-8160.2012.01.011 [28] 马璐, 曹文清, 张文静, 等. 北部湾北部海域夏季微型浮游动物对浮游植物的摄食压力[J]. 生态学报, 2014, 34(3): 546-554. https://www.ecologica.cn/html/2014/3/stxb201207201034.htm [29] ERGA S R, SSEBIYONGA N, HAMRE B, et al. Nutrients and phytoplankton biomass distribution and activity at the Barents Sea Polar Front during summer near Hopen and Storbanken[J]. J Mar Syst, 2014, 130: 181-192. doi: 10.1016/j.jmarsys.2012.12.008
[30] ELLWOOD M J, LAW C S, HALL J, et al. Relationships between nutrient stocks and inventories and phytoplankton physiological status along an oligotrophic meridional transect in the Tasman Sea[J]. Deep-Sea Res Pt I, 2013, 72: 102-120. doi: 10.1016/j.dsr.2012.11.001
[31] DEUTSCH C, SARMIENTO J L, SIGMAN D M, et al. Spatial coupling of nitrogen inputs and losses in the ocean[J]. Nature, 2007, 445(7124): 163-167. doi: 10.1038/nature05392
[32] 王晓伟, 李纯厚, 戴明. 大亚湾湾口海域冬季浮游植物生长限制性元素研究[J]. 南方水产, 2007, 3(4): 26-31. doi: 10.3969/j.issn.2095-0780.2007.04.005 [33] 刘东艳, 孙军. 不同氮磷比对中肋骨条藻生长特性的影响[J]. 海洋湖沼通报, 2002 (2): 39-44. doi: 10.3969/j.issn.1003-6482.2002.02.006 [34] 张莉红, 张学雷, 朱明远. 栉孔扇贝对硅藻和甲藻细胞的选择性摄食初探[J]. 海洋科学进展, 2008, 26(3): 372-376. doi: 10.3969/j.issn.1671-6647.2008.03.013 [35] 张莉红, 张学雷, 李瑞香, 等. 桑沟湾扇贝养殖对甲藻数量的影响[J]. 海洋科学进展, 2005, 23(3): 342-346. doi: 10.3969/j.issn.1671-6647.2005.03.013 [36] 张莉红. 扇贝养殖对浮游植物群落影响的初步研究[D]. 青岛: 中国海洋大学, 2005. https://www.doc88.com/p-7748827645555.html [37] ROSLAND R, BACHER C, STRAND Ø, et al. Modelling growth variability in longline mussel farms as a function of stocking density and farm design[J]. J Sea Res, 2011, 66(4): 318-330. doi: 10.1016/j.seares.2011.04.009
[38] 蒋增杰, 方建光, 门强, 等. 桑沟湾贝类筏式养殖与环境相互作用研究[J]. 南方水产, 2006, 2(1): 23-29. doi: 10.3969/j.issn.2095-0780.2006.01.005 [39] 张静, 刘玉, 马志华, 等. 流沙湾海域潮汐潮流的三维数值模拟和海域环境容量研究[J]. 海洋通报, 2013, 32(1): 34-44. doi: 10.11840/j.issn.1001-6392.2013.01.006 [40] 罗昭林, 朱长波, 郭永坚, 等. 流沙湾表层沉积物中碳、氮、磷的分布特征和污染评价[J]. 南方水产科学, 2014, 10(3): 1-8. doi: 10.3969/j.issn.2095-0780.2014.03.001 [41] 王如才, 张昭萍. 海水贝类养殖学[M]. 青岛: 中国海洋大学出版社, 2008: 276. https://xueshu.baidu.com/usercenter/paper/show?paperid=1c4x0gm0sn6x0x70fy3206u0xg679683&site=xueshu_se [42] MACISAAC H J, SPRULES G, JOHANNSON O E, et al. Filtering impacts of larval and sessile zebra mussels (Dreissena polymorpha) in western Lake Erie[J]. Oecologia, 1992, 92(1): 30-39. doi: 10.1007/BF00317259
[43] 赵宇, 刘佳, 高伟明. 扇贝养殖对海洋浮游植物的影响——以河北昌黎县为例[J]. 安徽农业科学, 2012, 40(10): 5966-5968. doi: 10.3969/j.issn.0517-6611.2012.10.090 [44] 秦培兵, 卢继武. 滤食性贝类对浅海养殖系统中营养盐循环的影响[J]. 海洋科学, 2001, 25(5): 27-29. doi: 10.3969/j.issn.1000-3096.2001.05.010 [45] CATALDO D, O′FARRELL I, PAOLUCCI E, et al. Impact of the invasive golden mussel (Limnoperna fortunei) on phytoplankton and nutrient cycling[J]. Aquat Invasions, 2012, 7(1): 91-100. doi: 10.3391/ai.2012.7.1.010
[46] 杨卫华, 高会旺, 刘红英, 等. 胶州湾扇贝养殖对海域环境影响的初步研究[J]. 海洋湖沼通报, 2007 (2): 86-93. doi: 10.3969/j.issn.1003-6482.2007.02.014 [47] 杨卫华, 高会旺. 胶州湾扇贝养殖对环境影响的数值模型研究[J]. 海洋环境科学, 2010, 29(4): 559-563. doi: 10.3969/j.issn.1007-6336.2010.04.024 [48] 王增焕, 柯常亮, 王许诺, 等. 流沙湾贝类养殖海域环境质量评价[J]. 南方水产科学, 2011, 7(3): 24-30. doi: 10.3969/j.issn.2095-0780.2011.03.005 [49] LAUER P R, FERNANDES M, FAIRWEATHER P G, et al. Benthic fluxes of nitrogen and phosphorus at southern bluefin tuna Thunnus maccoyii sea-cages[J]. Mar Ecol Prog Ser, 2009, 390: 251-263. doi: 10.3354/meps08186
[50] 黄长江, 董巧香, 吴常文, 等. 大规模增养殖区柘林湾叶绿素a的时空分布[J]. 海洋学报, 2005, 27(2): 127-134. doi: 10.3321/j.issn:0253-4193.2005.02.016 -
期刊类型引用(12)
1. 陈君,宋芬,彭祺,文威. 洪湖自然保护区浮游生物空间分布及与水质关系. 水生态学杂志. 2022(01): 63-70 .  百度学术
百度学术
2. 苏家齐,朱长波,李俊伟,李婷,陈素文,颉晓勇,张博. 流沙湾浮游动物群落特征及与鱼贝养殖的关系. 海洋渔业. 2019(03): 278-293 . 百度学术
3. 田洪阵,刘沁萍,Joaquim I.Goes,Helgado Rosario Gomes,杨萌萌. 近20年渤海叶绿素a浓度时空变化. 海洋学报. 2019(08): 131-140 . 百度学术
4. 粟丽,陈作志,黄梓荣,许友伟. 2015年春季南海北部陆架海域网采浮游植物群落结构及其与环境因子关系. 海洋学研究. 2019(03): 86-96 . 百度学术
5. 张映雪,王瑞,屈霄,夏文彤,辛未,郭传波,陈宇顺. 长江中游湖泊不同鱼类养殖方式对浮游植物群落的影响(英文). 水生生物学报. 2018(06): 1135-1143 . 百度学术
6. 苏家齐,朱长波,李俊伟,李婷,陈素文,颉晓勇,张博. 流沙湾浮游植物群落特征季节变化及其与养殖活动的关系. 渔业科学进展. 2018(06): 11-23 . 百度学术
7. 粟丽,黄梓荣,陈作志. 广东沿岸不同海域浮游植物群落结构特征的比较分析. 海洋环境科学. 2017(01): 61-65 . 百度学术
8. 武宇辉,王庆,魏南,刘之威,欧林坚,杨宇峰. 不同鲍养殖模式下浮游植物群落结构与水质特征的比较. 南方水产科学. 2017(06): 73-81 . 本站查看
9. 孙志伟,邱丽华,曹煜成,李卓佳,胡晓娟,苏浩昌,蔡卓平,文国樑. 蓝藻水华对水产养殖业影响的研究进展. 生态科学. 2017(01): 231-235 . 百度学术
10. 韩婷婷,齐占会,戴明,廖秀丽,吴风霞,巩秀玉,付贵权,黄洪辉. 大鹏澳不同区域尿素浓度与浮游植物脲酶活性的研究. 南方水产科学. 2016(02): 7-12 . 本站查看
11. 戴明,巩秀玉,刘华雪,廖秀丽,齐占会,黄洪辉. 2013年春季南沙海域不同水团中网采浮游植物群落特征. 南方水产科学. 2015(05): 38-46 . 本站查看
12. 陈露,李纯厚,魏小岚,刘永,陈作志,戴明,肖雅元,林琳. 南沙海域夏季氮磷添加模拟实验中浮游植物群落的变化. 南方水产科学. 2015(05): 56-66 . 本站查看
其他类型引用(7)
计量
- 文章访问数: 3319
- HTML全文浏览量: 173
- PDF下载量: 1894
- 被引次数: 19
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
