
Community characteristics of net phytoplankton in water masses in Nansha area during spring of 2013
-
摘要:
根据2013年春季调查资料研究了南沙海域不同水团中网采浮游植物的群落特征。共鉴定网采浮游植物4门62属222种,其中硅藻门和甲藻门种类数分别占67.1%和31.1%。浮游植物平均丰度为(16 655±36 570) 个· m-3,硅藻门平均丰度占93.6%。浮游植物多样性指数(H′)和均匀度指数(J′)平均分别为4.43和0.85。巴拉巴克海峡西侧混合水受苏禄海表层入侵水及冷涡影响,跃层浅,营养盐浓度高,浮游植物种类数和丰度均最高。巽他陆架北缘沿岸混合水受局部上升流的影响,浮游植物丰度次之。中南半岛沿岸混合水中无机氮(DIN)浓度最低,浮游植物种类数和丰度均低,而固氮蓝藻的丰度比例却最高。南沙中央表层水受陆源物质影响小,跃层深且薄,75 m以浅水层硅酸盐浓度最低,浮游植物丰度较低,但甲藻门丰度比例最高,J′最高,浮游植物丰度最低值出现在暖涡区域。
Abstract:Based on the samples collected in Nansha area in spring, 2013, we investigated the net phytoplankton community in order to determine the effects of water masses on their distribution. A total of 222 species, belonging to 62 genera of 4 phyla, were identified. Bacillariophyta and Pyrrophyta accounted for 67.1% and 31.1% in species number, respectively. The average abundance of net phytoplankton was (16 655±36 570) cell · m-3. Bacillariophyta accounted for 93.6% in abundance. The average diversity index and evenness index of net phytoplankton were 4.43 and 0.85. The species number and abundance of net phytoplankton were the highest in the mixed waters in the west of Balabac Strait, characterized by shallow thermocline and high nutrient concentrations resulting from the intrusion of Sulu Sea waters and the appearance of a clod eddy. In the mixed waters in the north edge of the Sunda Shelf, the abundance of net phytoplankton was sub-high due to the influence of upwelling. In the mixed waters from coast of the Indo-China Peninsula with low inorganic nitrogen, the species number and abundance of net phytoplankton were low, but the percentage of Cyanophyta increased to the maximum. In the Nansha central surface waters with less terrigenous matter, deep and thin thermocline, the lowest silicate concentration occurred at 0~75 m, the average abundance of net phytoplankton was low, but the percentage of Pyrrophyta increased to the maximum, and the average evenness index of net phytoplankton was the highest, while the lowest abundance of net phytoplankton appeared in the warm eddies.
-
Keywords:
- Nansha area /
- net phytoplankton /
- water masses /
- redundancy analysis
-
南沙海域位于南海南部,在巴拉望岛、加里曼丹岛和中南半岛之间,东部通过巴拉巴克海峡与苏禄海相通,南部通过巽他陆架北缘与爪哇海相通,是连结西太平洋和印度洋的重要海区,地理环境复杂。研究表明南沙海域上层存在4个不同水团,即南沙中央表层水、巴拉巴克海峡西侧混合水、巽他陆架北缘沿岸混合水和中南半岛沿岸混合水[1-3]。南沙海域受热带季风影响,春季(5月下旬之前)该海域处于冬季风消退、夏季风尚未形成的季风转换期,其海洋表层环流仍受冬季风的影响[4-5]。浮游植物是海洋的初级生产者,在海洋生态系统的物质、能量循环中起着重要的作用。南沙海域的浮游植物已有较多的研究,对其群落结构及其时空变化趋势已有初步的了解[6-11]。文章应用数理统计分析方法探讨了季风转换期南沙海域网采浮游植物的群落特征及其不同水团对网采浮游植物群落分布的影响。
1. 材料与方法
1.1 站位设计及采样
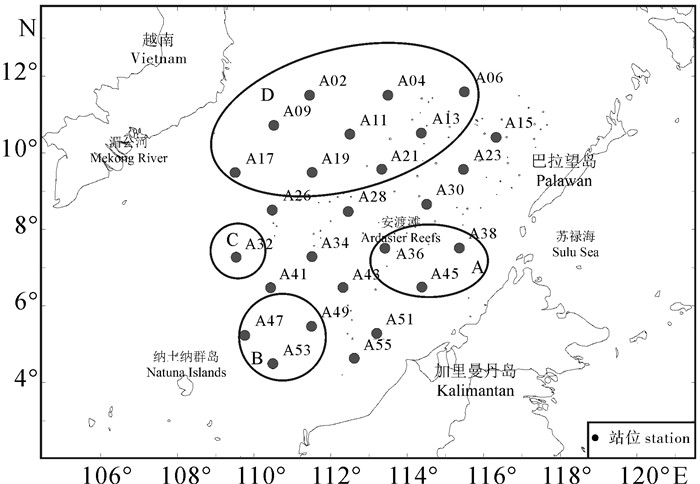
2013年3月~4月“南锋”号调查船在南沙海域(109°30′E~116°30′E,4°30′N~11°30′N)进行了26个站位的生态环境调查(图 1)。调查项目包括温度(T)、盐度(S)、营养盐[硝酸盐(NO3-)、亚硝酸盐(NO2-)、氨氮(NH4+)、磷酸盐(PO43-)和硅酸盐(SiO32-)]、叶绿素a(Chl-a)和网采浮游植物等。
1.2 样品分析鉴定
海水温度、盐度使用CTD(Sea-Bird)测定,营养盐和Chl-a采集5 m、10 m、25 m、50 m、75 m、100 m、150 m和200 m水样[12],使用Lachat QC 8500流动注射比色法测定营养盐,使用Turner Designs 10荧光法测定Chl-a。网采浮游植物用小型浮游生物网采集(网口面积0.1 m2,网目孔径77 μm),水深大于200 m的站位拖网深度为200 m,水深小于200 m的站位拖网深度从底至表,采集到的浮游植物样品用5%的甲醛溶液固定,带回实验室静置浓缩后在Leica DMLB生物显微镜下进行分类鉴定与计数。
1.3 数据分析
网采浮游植物群落结构使用优势度(Y)、Shannon-Wiener多样性指数(H′)和Pielous均匀度指数(J′)[13]。浮游植物与环境因子之间的关系使用Canoco for Windows 4.5软件进行排序分析[11, 14-15]。筛选出现频率≥30%的浮游植物种类数据,并经过log10(X+1)标准化转换,通过降趋对应分析(DCA)得出物种数据特征,最后选用冗余分析(redundancy analysis,RDA)绘制出物种、站位和环境因子的排序图。浮游植物空间分布图使用ArcGIS 10.0软件的地理统计模块绘制。海面高度异常(sea level anomalies,SLA)和地转流数据取自http://www.aviso.oceanobs.com 提供的网格化资料,该资料包括TOPEX/Poseidon,Jason-1 and ERS-1/2,空间分辨率是(1/3)°×(1/3)°。
2. 结果与讨论
2.1 不同水团温盐特征
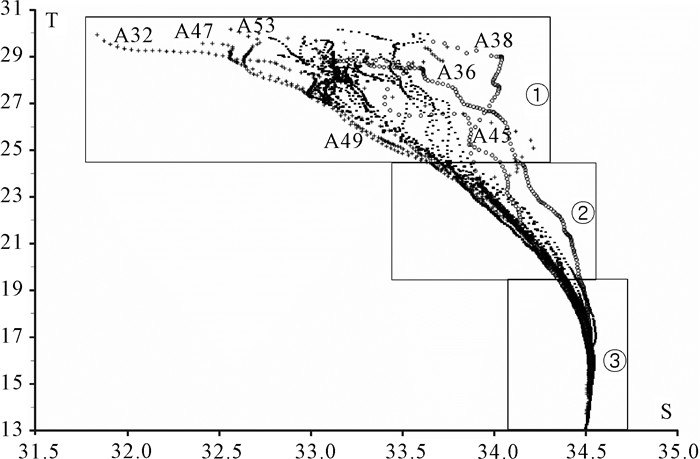
根据不同水层温盐上四分位数及其S-T点聚图变化特征,0~75 m水层75%的数据点在①层。75~120 m水层75%的数据点在②层,120 m以深水层的数据点则基本在③层(图 2)。75 m以浅水层温、盐的变幅最大,75~120 m水层温盐差异开始变小,至120 m以深水层S-T点聚图已基本重合。75 m以浅水层中A36、A38和A45站盐度最高,A32、A47、A49和A53站盐度最低,其他站位盐度介于上述两者之间。基于上述站位间的盐度差异,结合以往水团研究结果[1-3],将调查站位划分A、B、C、D 4个水团(图 1,一些位于水团交汇区的站位未划入4个水团中)。A区是巴拉巴克海峡西侧混合水,为苏禄海与南沙海水混合,盐度最高;B区是巽他陆架北缘沿岸混合水,源于爪哇海,受巽他陆架沿岸冲淡水影响,盐度较低;C区是中南半岛沿岸混合水,受中南半岛沿岸流影响,盐度较低;D区是南沙中央表层水,远离陆地,受沿岸水影响小,盐度介于苏禄海入侵水和沿岸混合水之间。
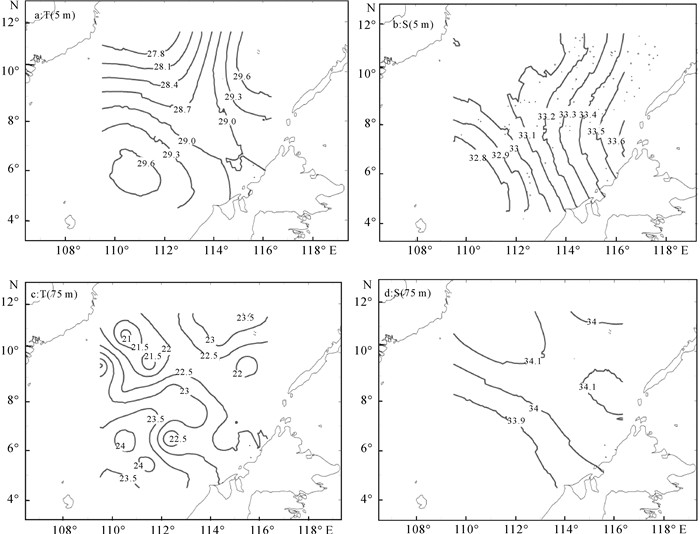
选取温盐空间差异较显著的5 m和75 m水层绘制其平面分布图,结果清晰地显示出表层高温高盐的苏禄海水通过巴拉巴克海峡自东向西锲入南沙海域,高温低盐的巽他陆架北缘沿岸混合水和中南半岛沿岸混合水范围局限在巽他陆架北缘附近,范围最大的南沙中央表层水嵌入于上述3个水团之间(图 3)。75 m水层,除B区和C区2个沿岸混合水影响较大的区域仍保持相对低盐外,其余大部分海域盐度变幅减弱;水温在维持表层平面分布趋势基础上,由于受温跃层的影响局部出现更多的波动,且在B区的A47和A53站发现了一个低温高盐的异常区,水温和盐度与同纬度邻近站位(A49和A55)相差0.77 ℃和0.15以上。
![]() 图 3 南沙海域5 m和75 m的水温和盐度Figure 3. Temperature and salinity in Nansha area at 5 m and 75 m
图 3 南沙海域5 m和75 m的水温和盐度Figure 3. Temperature and salinity in Nansha area at 5 m and 75 m2.2 网采浮游植物群落特征
共鉴定网采浮游植物4门62属222种,其中硅藻门44属149种,甲藻门15属69种,蓝藻门2属3种,金藻门1种。硅藻门和甲藻门分别占总种类数的67.1%和31.1%。
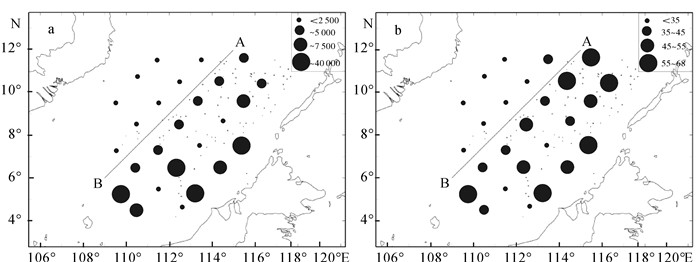
网采浮游植物丰度变化范围为902~85 910个· m-3,平均丰度为(16 655±36 570)个· m-3。硅藻门丰度占优势,占海域浮游植物平均丰度的93.6%。丰度高值区出现在南沙群岛的东部和南部,其丰度均值与西部和北部海域相差将近一个数量级(图 4-a)。不同站位浮游植物种类数为16~68种,以东部、南部和东北部海域种类较多(图 4-b)。H′变化范围为3.44~5.23,平均为4.43。J′变化范围为0.61~0.99,平均为0.85。
网采浮游植物空间分布特征如下:以图 4中A点(115°E,12°N)、B点(109°E,6°N)连线为界,连线以西浮游植物种类数、丰度和H′偏低,J′较高;而连线以东浮游植物种类数和丰度较高,其中丰度最高的东部和南部海域的H′和J′低于丰度次高的东北部海域。1984年~1988年的调查[6-7]和1993年5月的调查[8]结果同样是南沙群岛东部和南部的浮游植物丰度较高,可见这样的空间分布具有一定的代表性。
![]() 图 4 网采浮游植物丰度(a)和种类数(b)平面分布Figure 4. Abundance (a) and specie number (b) of net phytoplankton in Nansha area
图 4 网采浮游植物丰度(a)和种类数(b)平面分布Figure 4. Abundance (a) and specie number (b) of net phytoplankton in Nansha area2.3 不同水团的网采浮游植物群落特征
4个水团中A区网采浮游植物种类最多,C区种类最少,H′最低,D区J′最高,A区和B区浮游植物丰度明显高于C区和D区,且丰度较高的A区和B区硅藻门丰度比例较高(>94.2%),而丰度较低的C区和D区的甲藻门丰度比例较高(12.1%和18.6%),蓝藻门丰度比例则在C区最高(25.9%),在D区最低(1.3%)(表 1)。
表 1 网采浮游植物生态参数Table 1. Ecological parameters of net phytoplankton in Nansha area水团
water mass种类数
species
number多样性指数(H′)
Shannon-Wiener
diversity index均匀度指数(J′)
Pielous
evenness index丰度 abundance 平均值
average硅藻
Bacillariophyta甲藻
Pyrrophyta蓝藻
Cyanophyta金藻
ChrysophytaA 47 4.43 0.82 34 327 32 545 780 1 002 0 B 42 4.55 0.86 32 797 30 910 978 910 0 C 19 3.50 0.83 2 552 1 584 308 660 0 D 36 4.49 0.90 4 215 3 356 786 55 18 2.4 网采浮游植物和环境的排序分析
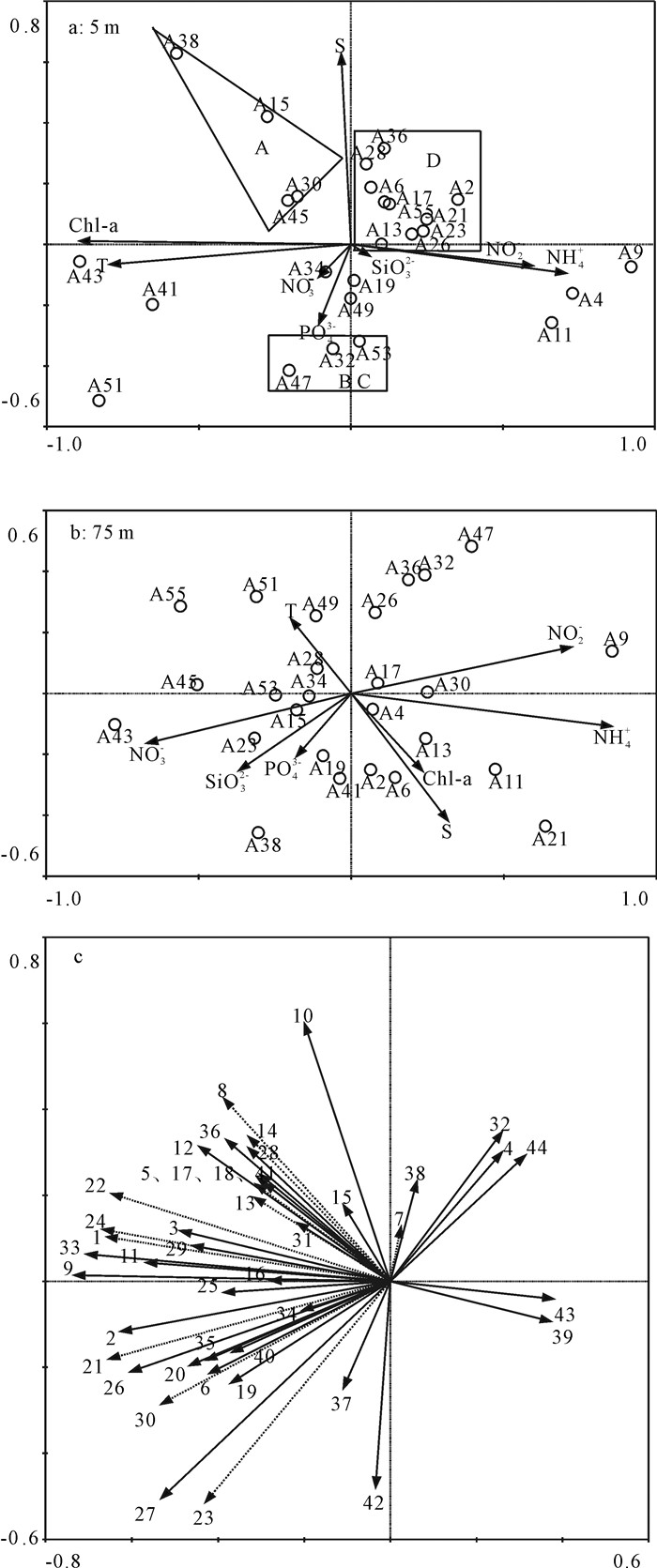
网采浮游植物和环境的排序图显示,表层(5 m)A、D区站位排序接近,B、C区站位排序接近,而A、D区和B、C区之间的距离较远(图 5-a)。这与表 1所述A-B水团和C-D水团组内网采浮游植物群落特征相近,而组间网采浮游植物群落特征相差较远的结果一致。影响表层站位排序的主要因子是水温和盐度。排序图还显示D区氨氮和亚硝酸盐较高,而该区域的网采浮游植物丰度较低,两者不呈正相关关系(图 5-a)。75 m水层站位的排序规律不明显,大致显现A、B区硝酸盐、磷酸盐和硅酸盐偏高趋势,正好与网采浮游植物丰度高值区对应,两者呈正相关关系(图 5-b)。
![]() 图 5 冗余分析排序图a. 5 m水层站位和环境;b. 75 m水层站位和环境;c.种类Figure 5. Redundancy analysis plotsa. stations and environmental factors at 5 m; b. stations and environmental factors at 75 m; c. species
图 5 冗余分析排序图a. 5 m水层站位和环境;b. 75 m水层站位和环境;c.种类Figure 5. Redundancy analysis plotsa. stations and environmental factors at 5 m; b. stations and environmental factors at 75 m; c. species入选排序的主要网采浮游植物种类根据其生态类型分析(表 2),发现近岸种(图中虚线所示)基本位于排序轴原点左侧,说明受沿岸混合水影响的A、B、C区近岸种丰度较高,而受陆源影响最小的D区高盐种丰度较高(图 5-c)。
表 2 入选RDA排序的网采浮游植物种类Table 2. Species of net phytoplankton included for RDA序号
No.种类
species生态习性
ecological habit平均丰度/个·m-3及百分比/%
average abundance
and pencentage出现频率/%
frequency of occurrence1 窄隙角毛藻 Chaetoceros affinis 温带近岸种 1 576/9.5 80.8 2 大西洋角毛藻骨条变种 C.atlanticus var. skeleton 温带外海种 8 80/5.3 50.0 3 奥氏角毛藻 C.aurivillii 暖水外海种 129/0.8 34.6 4 密聚角毛藻 C.coarctatus 暖水外海种 441/2.6 65.4 5 扁面角毛藻 C.compressus 广温近岸种 405/2.4 34.6 6 齿角毛藻 C.denticulatus 暖水外海种 222/1.3 34.6 7 爱氏角毛藻 C.eibenii 暖水近岸种 331/2.0 57.7 8 劳氏角毛藻 C.lorenzianus 暖水近岸种 1147/6.9 73.1 9 短刺角毛藻 C.messanensis 暖水外海种 642/3.9 73.1 10 秘鲁角毛藻 C.peruvianus 广温外海种 74/0.4 34.6 11 嘴状角毛藻 C.rostratus 暖水种 298/1.8 38.5 12 翼根管藻 Rhizosolenia alata 温带外海种 90/0.5 61.5 13 翼根管藻纤细变型 R.alata f. gracillima 广温近岸种 111/0.7 65.4 14 伯氏根管藻 R.bergonii 暖水外海种 58/0.3 42.3 15 距端根管藻 R.calcaravis 暖水广盐种 82/0.5 50.0 16 圆柱根管藻 R.cylindrus 暖水外海种 31/0.2 42.3 17 粗根管藻 R.robusta 暖水外海种 27/0.2 34.6 18 笔尖形根管藻 R.styliformis 暖水外海种 218/1.3 84.6 19 笔尖形根管藻粗径变种 R.styliformis var. latissima 暖水外海种 349/2.1 61.5 20 丛毛辐杆藻 Bacteriastrum comosum 暖水外海种 213/1.3 50.0 21 优美辐杆藻 B.delicatulum 广温近岸种 543/3.3 53.8 22 透明辐杆藻 B.hyalinum 广温近岸种 700/4.2 50.0 23 中华盒形藻 Biddulphia sinensis 暖水近岸种 125/0.7 30.8 24 佛朗梯形藻 Climacodium frauenfeldianum 暖水外海种 802/4.8 53.8 25 海洋棘冠藻 Corethron pelagicum 暖水外海种 122/0.7 65.4 26 地中海指管藻 Dacthliosolen mediterraneus 暖水广盐种 155/0.9 38.5 27 太阳双尾藻 Ditylum sol 暖水外海种 165/1.0 34.6 28 萎软几内亚藻 Guinardia flaccida 暖水近岸种 88/0.5 34.6 29 霍氏半管藻 Hemiaulus hauckii 暖水种 306/1.8 65.4 30 尖刺菱形藻 Nitzschia pungens 广温近岸种 500/3.0 57.7 31 美丽漂流藻 Planktoniella formosa 暖水外海种 104/0.6 69.2 32 太阳漂流藻 P.sol 暖水外海种 27/0.2 34.6 33 菱形海线藻 Thalassionema nitzschioides 广温广盐种 913/5.5 76.9 34 伏氏海毛藻 Thalassiothrix frauenfeldii 广温广盐种 98/0.6 57.7 36 长海毛藻 T.longissima 暖水外海种 91/0.5 69.2 37 细弱海链藻 Thalassiosira subtilis 暖水外海种 184/1.1 53.8 35 铁氏束毛藻 Trichodesmium thiebautii 暖水外海种 142/0.9 57.7 38 二齿双管藻 Amphisolenia bidentata 暖水广盐种 29/0.2 34.6 39 歧分角藻舞姿变型 Ceratium carriense f.volans 暖水外海种 19/0.1 30.8 40 偏转角藻 C.deflexum 暖水外海种 34/0.2 53.8 41 美丽角藻 C.pulchellum 暖水外海种 26/0.2 30.8 42 四叶鸟尾藻 Ornithocercus steinii 暖水外海种 42/0.2 57.7 43 纺锤梨甲藻 Pyrocystis fusiformis 暖水外海种 36/0.2 38.5 44 拟夜光梨甲藻 P.pseudonoctiluca 暖水外海种 133/0.8 73.1 2.5 不同水团对网采浮游植物群落的影响
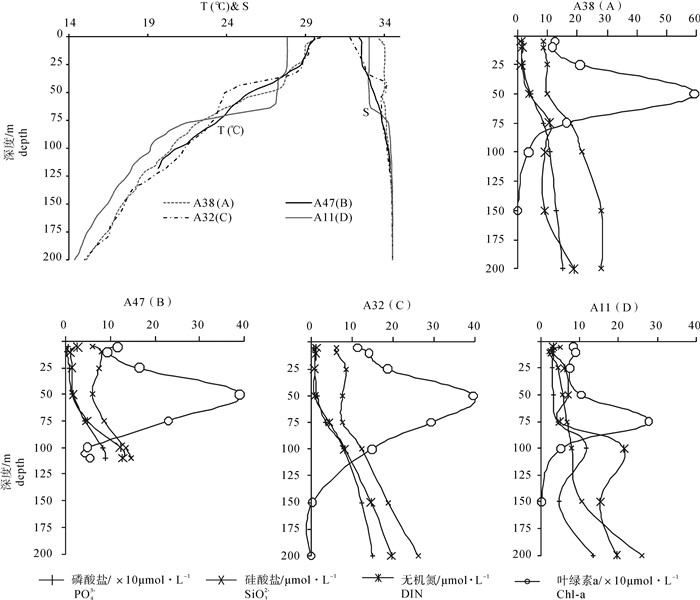
各水团选取1个代表性站位绘制其200 m以浅的环境因子剖面图(图 6),结果显示:A区的A38站盐跃层强度最小,营养盐跃层上界最浅,至50 m水层其无机氮和磷酸盐浓度已明显增高,且营养盐跃层强度最大,这一强大的营养盐跃层对该站网采浮游植物丰度高值区的形成有重要贡献。B区的A47站和C区的A32站温/盐跃层上界较浅,其营养盐跃层深度比A38站稍深,至75 m水层其营养盐有明显的增高。而D区的A11站的温/盐跃层上界最深,营养盐跃层亦较深,在75 m水层其营养盐仍未见明显的增高,而且其Chl-a最大值层也随之变深。与其他3个代表站位50 m层Chl-a大于其他采样水层不同,A11站75 m水层Chl-a大于其他采样水层。经统计D区共有88.9%的站位75 m Chl-a更大,而A、B、C 3个区域71.4%的站位50 m Chl-a更大。可见Chl-a最大值层深度与营养盐跃层深度相对应,反映出跃层屏障作用导致Chl-a在靠近跃层下界上方积累的现象。营养盐跃层深度与网采浮游植物丰度负相关,营养盐跃层强度与网采浮游植物丰度正相关。
![]() 图 6 4个代表性站位环境参数剖面图Figure 6. Profiles of environmental parameters of four representative stations
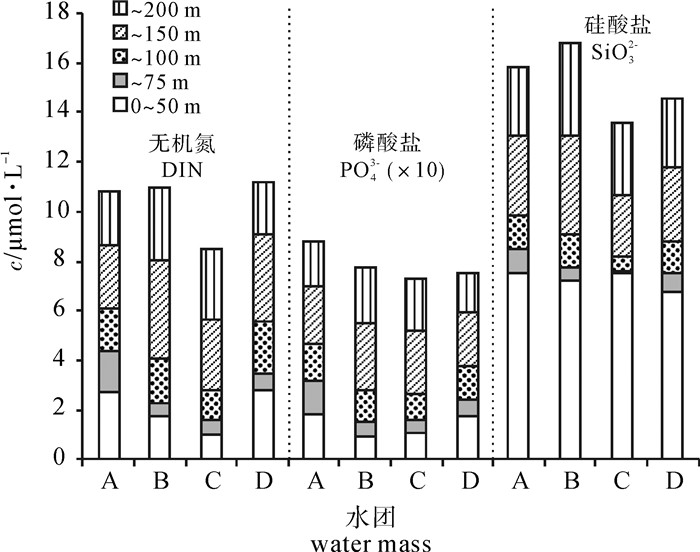
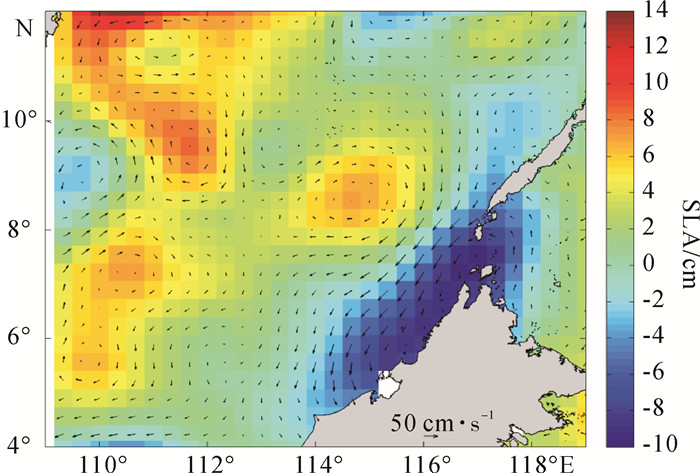
图 6 4个代表性站位环境参数剖面图Figure 6. Profiles of environmental parameters of four representative stations为进一步说明营养盐与网采浮游植物之间的关系,各水团的水柱平均营养盐浓度见图 7。A区巴拉巴克海峡西侧混合水中,200 m以浅的水柱平均营养盐浓度较高,且50~75 m跃层内营养盐增量最高。研究表明该区域受到苏禄海表层入侵水影响[3, 16-17],在春季该表层地转流在6°N~8°N间呈现出西向流的特征[18]。在水温、盐度的平面分布图中亦可见其向西扩展的趋势(图 3)。结合同一时期的海表面高度异常(SLA)分布,发现苏禄海表层入侵水舌西扩范围内及其西南部海域存在一个冷涡(图 8)。这一冷涡附近站位的网采浮游植物群落种类数和丰度均最高。陈楚群等[19]与朱根海等[20]的研究也表明该区域是高Chl-a、高浮游植物丰度所在。
![]() 图 7 4个水团0~200 m水柱平均营养盐浓度Figure 7. Average nutrient concentration at 0~200 m of four water masses
图 7 4个水团0~200 m水柱平均营养盐浓度Figure 7. Average nutrient concentration at 0~200 m of four water masses![]() 图 8 2013年3月26日南沙海域平均海面高度异常和地转流矢量分布图Figure 8. Average sea level anomaly (SLA) with overlaid geostrophic current vectors around Nansha area on 26 March, 2013
图 8 2013年3月26日南沙海域平均海面高度异常和地转流矢量分布图Figure 8. Average sea level anomaly (SLA) with overlaid geostrophic current vectors around Nansha area on 26 March, 2013B区巽他陆架北缘沿岸混合水中,由于中南半岛沿岸流和加里曼丹岛西岸的沿岸流也在这里汇聚[2],水团的交汇致使浮游植物种类较高。另外,沿岸流的离岸作用会引起该区局地底层海水的上升[2]。周文正等[21]研究发现巽他陆架北缘底层冷水受季风推动在春季开始形成,推测是南海次表层水通过巽他陆架纳土纳群岛两侧的深沟爬升到陆架地区的。B区75 m以浅水层的营养盐并不高,而75 m以深其营养盐快速增高,尤其是100 m以深其营养盐增量为4个水团最高。从75 m水层水温、盐度的平面分布中也发现B区存在一个低温高盐异常区,并且其附近站位(A41、A47、A49和A53)的Chl-a平均质量浓度(0.36 μg · L-1)是同纬度邻近站位(A43、A45、A51和A53)Chl-a平均质量浓度(0.14 μg · L-1)的2.6倍。可见底层冷水的涌升为上层海水带来了丰富的营养,并促发形成了网采浮游植物丰度次高值区。柯志新等[22]亦报道过上升流使该地区的生物量较大。
C区中南半岛沿岸混合水中,从海表面高度异常分布发现该区域附近有一暖涡存在(图 8),其200 m以浅水柱无机氮(DIN)、PO43-和SiO32-的平均浓度均为4个水团中最低,尤其是DIN浓度显著低于其他水团,网采浮游植物丰度最低,但C区的固氮蓝藻束毛藻属的丰度比例最高。已有研究表明束毛藻是热带、亚热带大洋寡营养真光层中的主要固氮生物[23],它可将大气中的氮气(N2)转换为浮游植物所需要的化合态氮(NH4+),其固氮量约为80 Mt · a-1[24],是热带、亚热带大洋中新氮的重要来源和新生产力的主要贡献者[25]。
D区南沙中央表层水由于远离陆地,受陆源物质影响小,75 m以浅水层硅酸盐浓度为4个水团最低,网采浮游植物总丰度以及硅藻丰度比例(79.6%)均较低,而甲藻丰度比例则最高(18.6%)。朱根海等[20]的研究也发现该区域浮游植物丰度较低。可能硅酸盐浓度对该海域浮游植物群落组成有一定的影响。D区的海表面高度异常分布图中发现越南沿岸(图 4-A~B连线以西)和8°N~10°N、113°E~115°E之间存在2个暖涡(图 8)。这与程旭华等[26]分析暖涡形成的时间一致。程旭华等认为暖涡在3月开始出现,4月~5月得到完全发展。这2个暖涡区域均为网采浮游植物丰度低值区。另外,D区又深又薄的温/盐跃层会阻碍营养盐垂直向上的对流,即使跃层内营养盐丰富,网采浮游植物丰度高,但由于其跃层薄,对其水柱网采浮游植物平均丰度的提高仍作用较小。邱章等[27]也发现该区域上混合层最厚,虽然D区上混合层的DIN和PO43-浓度在4个水团中属较高水平(图 7),但由于南沙群岛表层是典型的寡营养海区[28],Chl-a最大层都出现在跃层内,因此寡营养水域上混合层偏高的营养盐对水柱网采浮游植物平均丰度的贡献作用较小。
致谢: 对“南锋”号调查船全体工作人员在采样过程中提供的帮助,谨此致谢! -
![]()
图 3 南沙海域5 m和75 m的水温和盐度
Figure 3. Temperature and salinity in Nansha area at 5 m and 75 m
![]()
图 4 网采浮游植物丰度(a)和种类数(b)平面分布
Figure 4. Abundance (a) and specie number (b) of net phytoplankton in Nansha area
![]()
图 5 冗余分析排序图
a. 5 m水层站位和环境;b. 75 m水层站位和环境;c.种类
Figure 5. Redundancy analysis plots
a. stations and environmental factors at 5 m; b. stations and environmental factors at 75 m; c. species
![]()
图 6 4个代表性站位环境参数剖面图
Figure 6. Profiles of environmental parameters of four representative stations
![]()
图 7 4个水团0~200 m水柱平均营养盐浓度
Figure 7. Average nutrient concentration at 0~200 m of four water masses
![]()
图 8 2013年3月26日南沙海域平均海面高度异常和地转流矢量分布图
Figure 8. Average sea level anomaly (SLA) with overlaid geostrophic current vectors around Nansha area on 26 March, 2013
表 1 网采浮游植物生态参数
Table 1 Ecological parameters of net phytoplankton in Nansha area
水团
water mass种类数
species
number多样性指数(H′)
Shannon-Wiener
diversity index均匀度指数(J′)
Pielous
evenness index丰度 abundance 平均值
average硅藻
Bacillariophyta甲藻
Pyrrophyta蓝藻
Cyanophyta金藻
ChrysophytaA 47 4.43 0.82 34 327 32 545 780 1 002 0 B 42 4.55 0.86 32 797 30 910 978 910 0 C 19 3.50 0.83 2 552 1 584 308 660 0 D 36 4.49 0.90 4 215 3 356 786 55 18  下载: 导出CSV
下载: 导出CSV
表 2 入选RDA排序的网采浮游植物种类
Table 2 Species of net phytoplankton included for RDA
序号
No.种类
species生态习性
ecological habit平均丰度/个·m-3及百分比/%
average abundance
and pencentage出现频率/%
frequency of occurrence1 窄隙角毛藻 Chaetoceros affinis 温带近岸种 1 576/9.5 80.8 2 大西洋角毛藻骨条变种 C.atlanticus var. skeleton 温带外海种 8 80/5.3 50.0 3 奥氏角毛藻 C.aurivillii 暖水外海种 129/0.8 34.6 4 密聚角毛藻 C.coarctatus 暖水外海种 441/2.6 65.4 5 扁面角毛藻 C.compressus 广温近岸种 405/2.4 34.6 6 齿角毛藻 C.denticulatus 暖水外海种 222/1.3 34.6 7 爱氏角毛藻 C.eibenii 暖水近岸种 331/2.0 57.7 8 劳氏角毛藻 C.lorenzianus 暖水近岸种 1147/6.9 73.1 9 短刺角毛藻 C.messanensis 暖水外海种 642/3.9 73.1 10 秘鲁角毛藻 C.peruvianus 广温外海种 74/0.4 34.6 11 嘴状角毛藻 C.rostratus 暖水种 298/1.8 38.5 12 翼根管藻 Rhizosolenia alata 温带外海种 90/0.5 61.5 13 翼根管藻纤细变型 R.alata f. gracillima 广温近岸种 111/0.7 65.4 14 伯氏根管藻 R.bergonii 暖水外海种 58/0.3 42.3 15 距端根管藻 R.calcaravis 暖水广盐种 82/0.5 50.0 16 圆柱根管藻 R.cylindrus 暖水外海种 31/0.2 42.3 17 粗根管藻 R.robusta 暖水外海种 27/0.2 34.6 18 笔尖形根管藻 R.styliformis 暖水外海种 218/1.3 84.6 19 笔尖形根管藻粗径变种 R.styliformis var. latissima 暖水外海种 349/2.1 61.5 20 丛毛辐杆藻 Bacteriastrum comosum 暖水外海种 213/1.3 50.0 21 优美辐杆藻 B.delicatulum 广温近岸种 543/3.3 53.8 22 透明辐杆藻 B.hyalinum 广温近岸种 700/4.2 50.0 23 中华盒形藻 Biddulphia sinensis 暖水近岸种 125/0.7 30.8 24 佛朗梯形藻 Climacodium frauenfeldianum 暖水外海种 802/4.8 53.8 25 海洋棘冠藻 Corethron pelagicum 暖水外海种 122/0.7 65.4 26 地中海指管藻 Dacthliosolen mediterraneus 暖水广盐种 155/0.9 38.5 27 太阳双尾藻 Ditylum sol 暖水外海种 165/1.0 34.6 28 萎软几内亚藻 Guinardia flaccida 暖水近岸种 88/0.5 34.6 29 霍氏半管藻 Hemiaulus hauckii 暖水种 306/1.8 65.4 30 尖刺菱形藻 Nitzschia pungens 广温近岸种 500/3.0 57.7 31 美丽漂流藻 Planktoniella formosa 暖水外海种 104/0.6 69.2 32 太阳漂流藻 P.sol 暖水外海种 27/0.2 34.6 33 菱形海线藻 Thalassionema nitzschioides 广温广盐种 913/5.5 76.9 34 伏氏海毛藻 Thalassiothrix frauenfeldii 广温广盐种 98/0.6 57.7 36 长海毛藻 T.longissima 暖水外海种 91/0.5 69.2 37 细弱海链藻 Thalassiosira subtilis 暖水外海种 184/1.1 53.8 35 铁氏束毛藻 Trichodesmium thiebautii 暖水外海种 142/0.9 57.7 38 二齿双管藻 Amphisolenia bidentata 暖水广盐种 29/0.2 34.6 39 歧分角藻舞姿变型 Ceratium carriense f.volans 暖水外海种 19/0.1 30.8 40 偏转角藻 C.deflexum 暖水外海种 34/0.2 53.8 41 美丽角藻 C.pulchellum 暖水外海种 26/0.2 30.8 42 四叶鸟尾藻 Ornithocercus steinii 暖水外海种 42/0.2 57.7 43 纺锤梨甲藻 Pyrocystis fusiformis 暖水外海种 36/0.2 38.5 44 拟夜光梨甲藻 P.pseudonoctiluca 暖水外海种 133/0.8 73.1
下载: 导出CSV
-
[1] 赵焕庭. 南沙群岛自然地理[M]. 北京: 科学出版社, 1996: 186-187, 191-192. [2] 方文东. 南海南部海洋的环流结构与季节变化[D]. 青岛: 中国科学院海洋研究所, 2000. https://xueshu.baidu.com/usercenter/paper/show?paperid=22a9e80e7a0873bbfff1944681fac2c7&site=xueshu_se&hitarticle=1 [3] 方文东, 黄企洲, 邱章, 等. 春夏季风转换期间南海南部的异常表层水[J]. 热带海洋学报, 2001, 20(1): 78-81. doi: 10.3969/j.issn.1009-5470.2001.01.010 [4] 林锡贵. 影响南沙及其邻近海区的越赤道气流[J], 海洋与湖沼, 1990, 21(24): 328-336. https://ols.iocasjournals.cn/thesisDetails?columnId=36512802&Fpath=home&index=0&lang=zh [5] 林锡贵, 张庆荣. 南沙及其邻近海区的天气气候特征[J]. 热带海洋, 1990, 9(1): 9 -16. [6] 林秋艳, 林永水, 袁文彬. 浮游植物[R]//中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区综合调查研究报告(一)下卷. 北京: 科学出版社, 1989: 652-659. https://xueshu.baidu.com/usercenter/paper/show?paperid=4350d903d67fb203b5672e1115c3a9f6&site=xueshu_se [7] 林永水, 林秋艳. 南沙群岛海区浮游植物的分布特征[C]//中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物研究论文集②. 北京: 海洋出版社, 1991: 66-88. https://www.las.ac.cn/front/book/detail?id=4e3f8a47efab2f02c070a1dffa14e66f [8] 钱树本, 陈国蔚. 南沙群岛海区浮游植物多样性研究[C]//中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物多样性研究Ⅱ. 北京: 海洋出版社, 1996: 11-27. https://www.las.ac.cn/front/book/detail?id=de8872707ae91d5f09a3acbc2a1e7548 [9] 宋星宇, 黄良民, 钱树本, 等. 南沙海区春夏季浮游植物多样性及其分布[J]. 生物多样性, 2002, 10(3): 258-268. doi: 10.3321/j.issn:1005-0094.2002.03.002 [10] 李开枝, 郭玉洁, 尹健强, 等. 南沙群岛海区秋季浮游植物物种多样性及数量变化[J]. 热带海洋学报, 2005, 24(3): 25-30. doi: 10.3969/j.issn.1009-5470.2005.03.004 [11] 戴明, 刘华雪, 廖秀丽, 等. 初秋南沙群岛海域网采浮游植物群落特征及其与环境因子的关系[J]. 应用生态学报, 2013, 24(12): 3553-3560. [12] 国家技术监督局. GB/T12763-2007海洋调查规范[M]. 北京: 中国标准出版社, 2007: 12763.2-2007: 5-8;12763.4-2007: 13-23;12763.6-2007: 11-14, 30-37. http://www.zhongkejc.net [13] 马克平. 生物群落多样性的测度方法[M]//钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994: 141-165. https://xueshu.baidu.com/usercenter/paper/show?paperid=4b889beef7ab1258296336b8fdd58d19&site=xueshu_se [14] 廖秀丽, 陈丕茂, 马胜伟, 等. 大亚湾杨梅坑海域投礁前后浮游植物群落结构及其与环境因子的关系[J]. 南方水产科学, 2013, 9(5): 109-119. doi: 10.3969/j.issn.2095-0780.2013.05.017 [15] 郭永坚, 罗昭林, 朱长波, 等. 水产养殖对流沙湾浮游植物群落特征的影响[J]. 南方水产科学, 2015, 11(2): 57-65. doi: 10.3969/j.issn.2095-0780.2015.02.008 [16] WYRTKI K. Physical oceanography of the Southeast Asian waters[R]. NAGA Report, Vol. 2. Scientific results of marine investigations of the South China Sea and Gulf of Thailand 1959-1961. Scripps Institution of Oceanography. La Jolla, California: The University of Califonia, 1961: 33-47, 165. https://xueshu.baidu.com/usercenter/paper/show?paperid=c0b110cd17abe1297495cc2d61a5b0a2&site=xueshu_se&hitarticle=1
[17] CHU P C, FAN C W, LOZANO C J, et al. An airborne expendable bathythermograph (AXBT) survey of the South China Sea, May 1995[J]. J Geophys Res, 1998, 103 (C10): 21637-21652. doi: 10.1029/98JC02096
[18] 黄企洲. 南沙群岛海区的海流[C]//南沙群岛海区物理海洋研究论文集Ⅰ. 北京: 海洋出版社, 1994: 10-27. https://www.shukui.net/book/1476702.html [19] 陈楚群, 施平, 毛庆文. 南海海域叶绿素a浓度分布特征的卫星遥感分析[J]. 热带海洋学报, 2001, 20(2): 66-70. [20] 朱根海, 宁修仁, 蔡昱明, 等. 南海浮游植物种类组成和丰度分布的研究[J]. 海洋学报, 2003, 25(增刊2): 8-23. https://xueshu.baidu.com/usercenter/paper/show?paperid=fd9b3e4daad5ffc8b28a9ea8c702eabf&site=xueshu_se&hitarticle=1 [21] 周文正, 于非, 南峰, 等. 南海西南部巽他陆架底层冷水及其季节变化分析[J]. 海洋与湖沼, 2015, 46(1): 1-8. doi: 10.11693/hyhz20131200216 [22] 柯志新, 黄良民, 李刚, 等. 春末夏初巽他陆架表层水体的叶绿素粒级结构及其和营养盐的关系[J]. 海洋学报, 2012, 34(3): 190-196. [23] JENKINS W J. Nitrate flux into the euphotic zone near Bermuda[J]. Nature, 1988, 331: 521-523. doi: 10.1038/331521a0
[24] CAPONE D G, ZHER J P, PAERL H W, et al. Trichodesmium, a globally significant marine cyanobacterium[J]. Science, 1997, 276: 1221-1229. doi: 10.1126/science.276.5316.1221
[25] KARL D, LETELIER R, TUPAS L, et al. The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean[J]. Nature, 1997, 388: 533-538. doi: 10.1038/41474
[26] 程旭华, 齐义泉, 王卫强. 南海中尺度涡的季节和年际变化特征分析[J]. 热带海洋学报, 2005, 24(4): 51-59. doi: 10.3969/j.issn.1009-5470.2005.04.008 [27] 邱章, 黄企洲. 南沙群岛海区温跃层时空分布的分析[C]//中国科学院南沙综合科学考察队. 南沙群岛海区物理海洋研究论文集Ⅰ. 北京: 海洋出版社, 1994: 64-80. https://www.las.ac.cn/front/book/detail?id=19bb576144d73c14a9a2046e37a308f4 [28] 蔡平河, 黄奕普, 沈国英, 等. 南沙海域初级生产力与水柱垂直稳定度的关系[J]. 海洋学报, 2000, 22(5): 137-140. doi: 10.3321/j.issn:0253-4193.2000.05.018 -
期刊类型引用(5)
1. 戴明, 刘华雪, 吴风霞, 巩秀玉, 廖秀丽, 黄洪辉. 万山群岛海域网采浮游植物与环境因子的关系. 海洋环境科学. 2019(04): 540-547+554 .  百度学术
百度学术
2. 张静, 严武科, 吕少梁, 曾嘉维, 王学锋. 2015年防城港近岸海域浮游桡足类群落结构的季节变化. 广东海洋大学学报. 2018(06): 18-28 . 百度学术
3. 郭术津, 孙晓霞, 郑珊, 罗璇, 朱明亮, 梁俊华. 热带西太平洋Y3和M2海山区网采浮游植物群落结构特征. 海洋科学. 2018(10): 31-40 . 百度学术
4. 吕少梁, 王学锋, 曾嘉维, 陈海刚, 王力飞, 贾晓平. 防城港海域浮游植物群落结构及其环境适应性. 南方水产科学. 2017(04): 17-25 . 本站查看
5. 徐姗楠, 杨玉滔, 粟丽, 龚玉艳, 陈作志. 珠江口南沙海域浮游植物群落结构特征. 南方水产科学. 2017(04): 26-33 . 本站查看
其他类型引用(1)
计量
- 文章访问数: 3168
- HTML全文浏览量: 197
- PDF下载量: 915
- 被引次数: 6
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
