
Decadal changes in growth, mortality and maturity parameters of Evynnis cardinalis in Beibu Gulf
-
摘要:
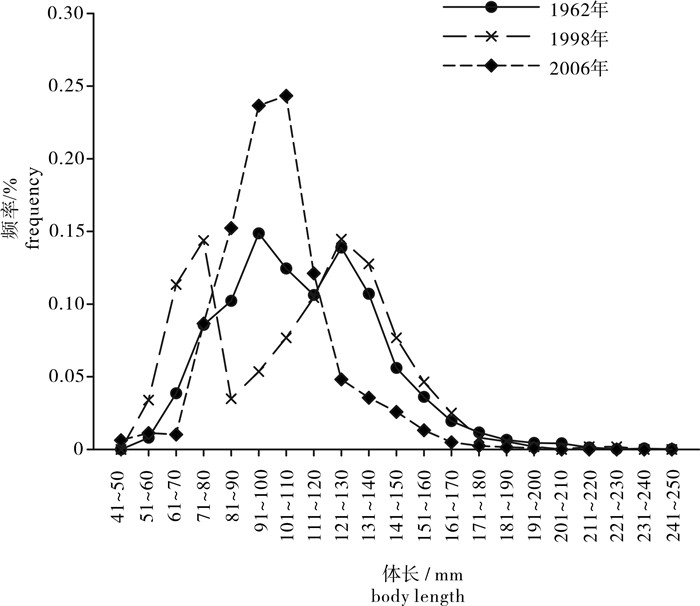
根据1961年~1962年(1962年)、1997年~1999年(1998年)以及2006年北部湾底拖网调查的渔业生物学资料,对3个时期二长棘犁齿鲷(Evynnis cardinalis)的群体结构、初次性成熟体长(L50)以及生长、死亡参数等进行研究。结果表明:1)与1962年和1998年相比,2006年二长棘犁齿鲷的体长频率分布由双峰型转变为单峰型,雌雄比明显增加。2)平均体长由1962年的108.8 mm减小到2006年的102.0 mm;L50由108.9 mm减小到95.9 mm;生长系数由0.35增加到0.50;二长棘犁齿鲷小型化、性成熟提前和生长加快现象明显。3)对比1962年,1998年并未发生平均体长、L50的减小,分析认为这可能是由于这段时期二长棘犁齿鲷并未受到长时间的过度捕捞。4)3个时期的开发率分别为0.49、0.62和0.60,目前二长棘犁齿鲷处于过度开发状态。
Abstract:We evaluated the population structure, first maturity lengths (L50), growth and mortality coefficient of threadfin porgy Evynnis cardinalis in Beibu Gulf using biological data obtained from bottom trawls conducted during 1961~1962(1962), 1997~1999(1998) and 2006. Results show that the bimodal patterns of length frequency distribution in 1962 and 1998 had turned into unimodal type in 2006 and the female-male ratio increased obviously; the mean body length decreased from 108.8 mm in 1962 to 102.0 mm in 2006; L50 decreased from 108.9 mm in 1962 to 95.9 mm in 2006, and growth coefficient (K) increased from 0.35 in 1962 to 0.50 in 2006, which indicates that E.cardinalis tended to be miniaturized with an increase in speed of growth and a decrease in time to mature. Slight increase in mean body length and L50 from 1962 to 1998 indicates longtime overfishing of E.cardinalis did not happened in this period. The exploitation rates (E) in 1962, 1998 and 2006 were 0.49, 0.62 and 0.60, respectively, suggesting that this stock has been over-exploited since the 1990s.
-
Keywords:
- Evynnis cardinalis /
- biological parameters /
- decadal changes /
- Beibu Gulf
-
哈维弧菌(Vibrio harveyi)为革兰氏阴性弧形短杆菌,极生单鞭毛,基因组DNA的G+C含量在46% ~48%之间,菌体大小为0.44~1.38 μm,在3%NaCl的营养琼脂平板上形成的菌落呈圆形灰色,TCBS培养基上为黄色,对O/129敏感[1-2]。哈维弧菌在海洋环境中主要分布于自由水体、浮游动植物体表、海底沉积物中,是一种广泛存在于海洋环境中的条件致病菌,能够感染多种水产经济动物[3-5],给海水养殖业带来巨大的损失。
在养殖中抗菌药物的使用致使大量病原菌产生耐药性,对抗生素表现出强烈抗性,病原菌对抗菌药物的耐药性问题日益显露出来[6-7]。细菌接触抗菌药物可发生变异获得耐药性,其机制主要有4种[8-10]:1)产生抗生素酶,使抗生素失活;2)作用靶位变异,不应答药物;3)外膜通透性改变,阻断药物进入;4)激活外派体系,加速泵出进入体内的药物。研究发现,哈维弧菌含有多种耐药性质粒基因[11],可通过接合的方式传播耐药基因[12-13],同时哈维弧菌具有较强的外排泵作用[14-15],导致多重耐药的发生。
依据美国临床和实验室标准协会(clinical and laboratory standards institute)抗生素敏感试验标准,将菌株对抗生素的敏感性以抑菌圈直径为变量,划分为敏感(S)、中介敏感(I)和耐药(R)3个类别,使菌株对同一种药物获得统一和可对比性的耐药性,同一菌株对多种抗生素可形成一种特定的耐药谱型。随着宿主和时间的改变,细菌耐药谱也会发生变异[16]。有研究发现,水产养殖环境中所分离的哈维弧菌对头孢菌素类的头孢噻吩(KF)、头孢氨苄(CEF)、头孢拉定(CEP)和头孢西丁(FOX)以及新型β-内酰胺单酰胺环类的氨曲南(AZT)等多种抗生素耐药[17-18]。而在海产品中分离的哈维弧菌的耐药谱表现为DOX/AMP/PEN/GEN/CIP/NOR/AK、DOX/AMP/PEN/GEN/CIP[19]。此外,研究者对马来西亚的斑节对虾(Penaeus monodon)哈维弧菌的耐药性研究发现,对氯霉素(CHL)、四环素(TET)和呋喃唑酮(FUR)敏感菌株高达96.7%,80%菌株对卡那霉素敏感,56.7%菌株对磺胺甲基异哑唑(SMZ)敏感[20]。不同来源的哈维弧菌的耐药谱研究证实,不同宿主、地点以及时间所分离的菌株耐药谱存在明显差异。
该研究以2012年~2014年南海沿海养殖区域患病鱼体分离得到的90株哈维弧菌为对象,测定其对12种常见抗生素的敏感性,并根据地点、时间以及宿主情况,分析其耐药谱差异。以形成的抑菌圈直径为变量,对各菌株的耐药性情况进行系统聚类,分析耐药谱的聚类关系,为南海地区的哈维弧菌的聚类情况和分析耐药谱的变迁提供依据。
1. 材料与方法
1.1 菌株来源
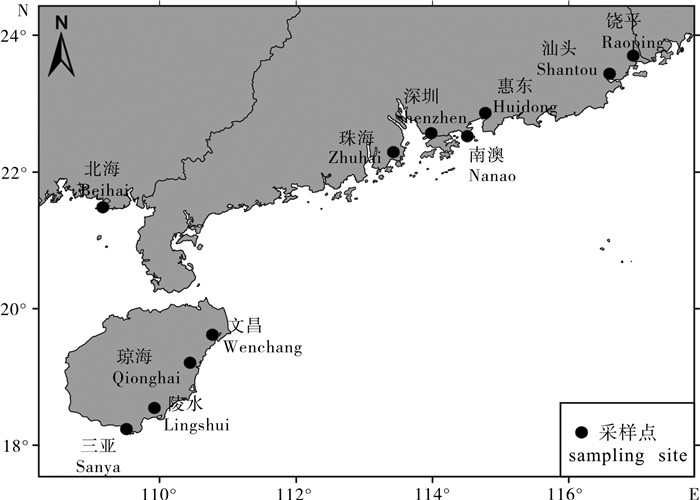
90株哈维弧菌菌株为笔者实验室保存菌株,于2012年7月至2014年5月分离自广东、广西和海南沿岸养殖的患病鱼(图 1),其中海南沿岸(包括海南的东海线陵水新村港、文昌烟堆、文昌铺前、三亚红沙、琼海谭门、黎安)分离株56株,其主要的宿主是乌鲳(Formio niger)、棕点石斑鱼(Epinephelus fuscoguttatus)、珍珠龙趸(鞍带石斑鱼♂×棕点石斑鱼♀)、卵形鲳鲹(Trachinotus ovatus)、点篮子鱼(Siganus guttatus)、布氏鲳鲹(T.blochii)、东星斑(Plectropomus leopardus)、青石斑鱼(E.awoara)、军曹鱼(Rachycentron canadum)和川纹笛鲷(Lutjanus sebae)等;广东沿海地区(包括饶平井州、红山码头、欧边、惠东盐州港、流沙港、深圳南澳、珠海桂山岛等)分离株31株,其宿主主要是浅色黄姑鱼(Nibea coibor)、卵形鲳鲹、点篮子鱼、双棘黄姑鱼(N.diacanthus)、珍珠龙趸、青石斑鱼、斜带髭鲷(Hapalogenys nitens)、芝麻斑(Epinephelus sp.)等;广西北海地区棕点石斑鱼分离到菌株3株。
1.2 药敏试验
采用纸片扩散法(K-B法)进行试验,记录药敏纸片抑菌圈的直径[21]。药敏纸片为12种常见的抗生素(均购于杭州天和微生物试剂有限公司):头孢克肟(CEF,5 μg · 片-1)、庆大霉素(GEN,10 μg · 片-1)、四环素(TET,30 μg · 片-1)、环丙沙星(CIP,5 μg · 片-1)、诺氟沙星(NOR,10 μg ·片-1)、氯霉素(CHL,30 μg · 片-1)、复方新诺明(T/S,23.7/1.25 μg · 片-1)、利福平(RIF,5 μg ·片-1)、呋喃唑酮(FUR,100 μg · 片-1)、阿莫西林(AMO,20 μg · 片-1)、红霉素(ERY,15 μg · 片-1)和恩诺沙星(ENR,10 μg · 片-1)。以大肠杆菌ATCC25922为质控菌,参照美国临床和实验室标准协会[22-24](clinical and laboratory standards institute)抗生素敏感试验标准对耐药谱进行分析。
1.3 耐药谱的分析
统计抗生素所形成的抑菌圈直径大小,将每种抗生素的耐药情况用S、I、R记录,每株菌株对12种抗生素形成唯一的耐药谱。对耐药谱进行统计分类,计算耐药率,并根据地区、时间以及宿主的不同统计分析具有耐3种及以上抗生素的耐药谱型的差异。其中耐药谱统计分类的分析方法如下:
耐药率:某一细菌对抗生素的耐药菌株与检验的总菌株数的比率;
耐药谱型丰富度:菌株具有的耐药谱型数量与菌株数量的比值;
耐药谱型率:部分菌株具有的耐药谱型数量占全部菌株具有的耐药谱型数量的比率。
1.4 哈维弧菌耐药性的聚类分型
采用SPSS 19.0软件对菌株进行聚类分析,以抗生素形成的抑菌圈直径作为变量,采用系统聚类[25]的方法进行聚类分析,以平方Euclidean距离表示菌株间的耐药性同类关系,结果以树状图的形式显示。
2. 结果
2.1 哈维弧菌的耐药谱型
对90株哈维弧菌抗生素的耐药谱型进行分析,得到A~Z共26种耐药谱型,谱型丰富度为28.9%,各谱型对应的耐药谱见表 1,其中A、I和J的耐药谱型均为抗2种抗生素,且3个耐药谱均抗阿莫西林,包含菌株45株,占总菌数的50%,谱型丰富度为11.5%;C型耐药谱仅抗阿莫西林1种抗生素,包含菌株9株,占总菌数10%,谱型丰富度为3.8%;B、D、G、H、V、Y和Z型耐药谱均抗3种抗生素,除Y和Z外,其他耐药谱的均抗阿莫西林,包含菌株共12株,占总菌数的13.3%,谱型丰富度为27%;K型不抗任何抗生素,共1株;其余的14种耐药谱型均为抗3种以上的抗生素,包含菌株共23株,占总菌数的25.6%,谱型丰富度为53.8%。
表 1 哈维弧菌菌株耐药谱分型Table 1. Antibiogram types of V. harveyi strains谱型
type菌株数
number of strain耐药谱
antibiogram谱型
type菌株数
number of strain耐药谱
antibiogramA 42 FUR/AMO N 1 T/S/CEF/RIF/TET B 1 FUR/NOR/AMO O 1 FUR/T/S/CEF/RIF/TET C 9 AMO P 1 FUR/T/S/CEF/RIF/TET/ERY/AMO D 1 FUR/CEF/AMO Q 1 FUR/T/S/CEF/RIF/ERY/AMO E 1 FUR/RIF/ERY/AMO R 4 FUR/T/S/CEF/RIF/AMO F 1 FUR/GEN/ERY/AMO S 1 T/S/CEF/RIF/TET/AMO G 5 FUR/RIF/AMO T 4 FUR/T/S/CEF/AMO H 2 FUR/ERY/AMO U 1 FUR/T/S/CEF/ERY/AMO I 1 ERY/AMO V 1 FUR/GEN/AMO J 2 RIF/AMO W 1 FUR/T/S/CEF/TET/ERY/AMO K 1 X 1 FUR/NOR/RIF/CIP L 2 FUR/T/S/CEF/RIF/CHL/TET/ERY/AMO Y 1 FUR/NOR/RIF M 3 FUR/T/S/CEF/RIF/CHL/TET/AMO Z 1 T/S/CEF/RIF 从时间分布上看,2012年、2013年和2014年分别具有10、18和8种耐药谱型,从地区分布上,海南、广东和广西三个地区分别包含17、13和2种耐药谱型。2012年7月~12月分离株22株,具有耐药谱型10种,谱型丰富度38.5%,耐药谱型丰富度45.5%,K型不抗任何抗生素,除Z型一株外其他20株菌株均耐阿莫西林,耐药抗生素种类最多达8种。2013年分离株46株,具有18种谱型,谱型丰富度为69.2%,耐药丰富度为39.1%,耐药广泛,从1种到8种均有,大多包含FUR/T/S/CEF、AMO、FUR/RIF三种之一。2014年1月~5月分离株22株,具有耐药谱型8种,谱型丰富度为30.8%,耐药谱型丰富度为36.4%。宿主间耐药谱差异明显,如珍珠龙趸、棕点石斑鱼和浅色黄姑鱼分离菌株耐药谱分别是17、7和5种,而其他宿主如卵形鲳鲹、点篮子鱼、双棘黄姑鱼、青石斑鱼、军草鱼等耐药谱为1~3种(图 2)。
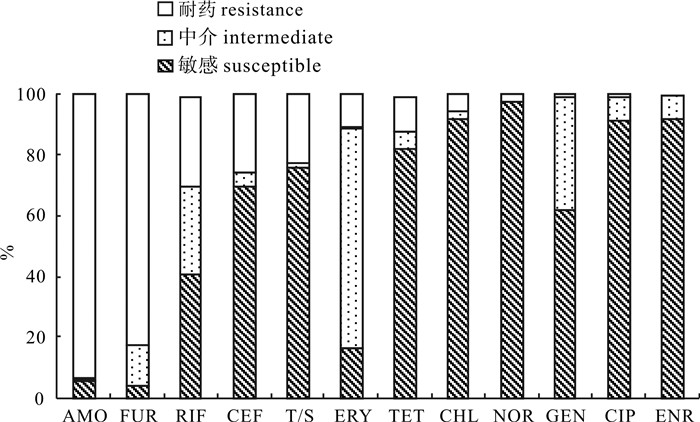
90株哈维弧菌对12种抗生素的耐药情况差异较大(图 3),其中对阿莫西林的耐药率达到93.7%,呋喃唑酮的耐药率也高达82.3%;复方新诺明、头孢克肟和利福平耐药率为22%~30%;庆大霉素、诺氟沙星、氯霉素、四环素、环丙沙星和红霉素的耐药率在10%及以下;而所有菌株均对恩诺沙星不耐药。
![]() 图 3 哈维弧菌菌株对抗生素的耐药性情况比较Figure 3. Comparison of antibiotic resistance of V. harveyi strains
图 3 哈维弧菌菌株对抗生素的耐药性情况比较Figure 3. Comparison of antibiotic resistance of V. harveyi strains2.2 哈维弧菌耐药性的聚类分析
对90株哈维弧菌做样本聚类分析,以12种抗生素的抑菌圈直径具体数值作为变量(图 2),大部分菌株聚类为i~v 5个亚群;当欧式平方达到2时,亚群ⅰ、ⅱ先合并为Group Ⅰ,ⅳ、ⅴ优先合并为一支;当欧式平方达到3时,亚群ⅲ、ⅳ、ⅴ合并为Group Ⅱ。当欧式平方达到5时Group Ⅰ、Group Ⅱ两个组群聚合。另外4株菌株聚类结果欧式距离平方较大,归于其他类。90株哈维弧菌聚类分析得到Group Ⅰ、Group Ⅱ和其他组群,分别含有11、12和4种耐药谱,耐药谱型率分别为16.9%、57.1%和100%。在同一亚群中,包含多种的耐药谱型,亚群间耐药谱型相差大,菌株数量差异也很大。
2.3 哈维弧菌2个耐药组群的比较分析
总体上,组群Group Ⅱ的耐药率要高于Group Ⅰ(表 3)。2个组群在T/S、CEF、RIF、CHL和TET的敏感性存在明显的差异,且Group Ⅰ明显高于Group Ⅱ。2个组群在FUR、NOR、CIP、ENR和AMO耐药性与敏感性均无明显差异。Group Ⅰ与Group Ⅱ2个组群对FUR、TET、CIP和ENR在中介敏感上存在着明显差异。此外,耐药谱型量Group Ⅱ要多于Group Ⅰ,谱型丰富度更高。
表 2 哈维弧菌耐药亚群的耐药谱分布Table 2. Antibiotic resistance patterns of different V. harveyi subgroups组群
group亚群
subgroup菌株数
number of strain谱型数
number of types谱型率/%
type rate谱型丰富度/%
type abundance耐药谱(谱型/菌株数)
antibiogram (type/number of strains)Group Ⅰ ⅰ 58 8 30.7 13.8 C/9,A/40,B/1,D/1,E/1,F/1,G/3,H/2 ⅱ 7 5 19.2 71.4 K/1,A/1,J/2,I/1,G/2 Group Ⅱ ⅲ 6 4 15.4 66.7 L/1,M/3,N/1,O/1, ⅳ 8 5 19.2 62.5 L/1,P/1,Q/1,R/4,S/1 ⅴ 7 4 15.4 57.1 T/4,U/1,V/1,W/1 其他other 4 4 15.4 100 X/1,Y/1,Z/1,A/1 表 3 哈维弧菌耐药菌群对抗生素敏感(S)、中介(I)、耐药(R)情况比较Table 3. Comparison of antibiotic resistance from susceptible, intermediate and resistance of V. harveyi耐药性
results of resistant耐药菌群
group耐药率/% resistance FUR T/S GEN CEF NOR RIF CHL TET CIP ENR ERY AMO S Group Ⅰ 1.54 96.92 58.46 92.31 100 66.15 100 95.38 95.38 96.92 15.38 1.54 Group Ⅱ 9.52 4.76 66.67 0 100 14.29 71.43 38.1 80.95 85.71 14.29 4.76 其他 25 75 100 75 50 0 75 100 75 50 50 75 I Group Ⅰ 18.46 3.08 40 6.15 0 20 0 3.08 4.62 3.08 78.46 0 Group Ⅱ 0 0 33.33 0 0 19.05 4.76 14.29 19.05 14.29 57.14 4.76 其他 0 0 0 0 0 25 25 0 0 50 50 0 R Group Ⅰ 80 0 1.54 1.54 0 13.85 0 1.54 0 0 6.15 96.92 Group Ⅱ 90.48 95.24 0 100 0 66.67 23.81 47.62 0 0 28.57 90.48 其他 75 25 0 25 50 75 0 0 25 0 0 25 2.4 哈维弧菌的多重耐药性
多重耐药谱是菌株抗3类及以上抗生素形成的耐药谱,该研究多重耐药谱21种,包含菌株共35株,占总菌数的38.9%,谱型丰富度为80.8%,包括海南20株、广东12株和广西3株(表 4),分别占该地区菌株总数的21.4%、25.8%和100%。海南地区分离株的多重耐药谱包括FUR/T/S/CEF、以及FUR/GEN/ERY/AMO或FUR/NOR/RIF/CIP两种耐药谱。广东沿海地区的分离株多重耐药谱多数包含T/S/CEF/RIF/TET、RIF/ERY/AMO和RIF/TET/AMO之一。广西沿海地区的分离株耐药谱包括FUR/T/S/CEF/TET/ERY/AMO和FUR/T/S/CEF/AMO两种。
表 4 广东、海南、广西地区抗3种及以上抗生素耐药谱情况Table 4. Three or more than three kinds of antibiotic resistance patterns in Guangdong, Hainan, Guangxi Provinces广东地区Guangdong 海南地区Hainan 广西地区Guangxi 耐药谱型
antibiogram type菌株数
number of strains耐药谱型
antibiogram type菌株数
number of strains耐药谱型
antibiogram type菌株数
number of strainsB 1 D 1 T 2 E 1 F 1 W 1 G 2 G 3 M 2 H 2 N 1 L 2 O 1 M 1 P 1 Q 1 R 1 R 3 S 1 T 2 Y 1 U 1 V 1 X 1 Z 1 3. 讨论
3.1 哈维弧菌的耐药情况
有研究指出,在养殖场中细菌的密度、单位体积内生物量和群落结构等可能会引起细菌耐药性的变化,细菌敏感性试验表明沉积物造就了细菌的高百分比,广谱的耐药性[26-27]。哈维弧菌的耐药谱会随着环境、人为条件的变化不断改变。有学者对水产致病菌耐药性进行了研究,54个受试菌株检出42个耐药株,耐药率高达77.8%[28]。对2012年~2014年采集的90株哈维弧菌分离株,试验中选择的12种抗生素包括国标渔药以及以前可用或禁用的药物,后续的研究工作者可用于借鉴分析养殖中抗生素可能的使用情况、抗生素风险及耐药性污染的评估工作。药物敏感性分析结果显示,菌株的耐药谱型丰富度达28.9%,其中多重耐药谱(耐3类及以上抗生素)21种,包含菌株共35株,占总菌数的38.9%,谱型丰富度为80.8%。耐抗生素种类最高达8种,说明从南海地区养殖鱼分离的哈维弧菌具有复杂多样的耐药谱。
3.2 哈维弧菌耐药性的时空分布
随着时间年份递进,哈维弧菌耐药谱种类也发生着变化更新,耐药谱不断扩大延伸。对2012年、2013年和2014年的耐药谱进行分析(图 2,表 1),3个年份对庆大霉素、诺氟沙星、氯霉素、环丙沙星耐药率低;对利福平的耐药率相近(30%);对呋喃唑酮、阿莫西林的耐药率高(90%以上);对头孢克肟、四环素、复方新诺明的耐药率存在明显的差异,如头孢克肟3年耐药率分别是18.2%、32.6%和13.6%,2013年明显高于其他2年;对红霉素均处于中介敏感,没有明显差异;对恩诺沙星的耐药率为0。2006年研究了香港地区的两个自然保护区分离的所有弧菌对氨苄西林、羧苄青霉素、先锋霉素、克林霉素、硫酸粘菌素、红霉素、梭链孢酸、甲氧西林、呋喃妥英和青霉素的耐药性[29],结果表明哈维弧菌具有广谱的耐药性,与该研究结果相近,说明地区不同,哈维弧菌菌株均具有多重耐药性。2007年~2012年南海地区分离的哈维弧菌对12种常见抗生素的耐药性试验结果显示,共包含26种耐药谱型,多重耐药谱型数量为13种,占总耐药谱型数量的50.0%[30],此试验与2007年~2012年南海地区哈维弧菌多种耐药性比较,哈维弧菌2012年~2014年多种耐药谱数量上升到21种,包含的菌株更多,说明南海地区病原菌的耐药性在增强,抗生素更加广泛,对水产养殖业来说病原菌的危害也更大。
海南、广东和广西3个地区分别包含17种、13种和2种耐药谱型,3个地区的耐药谱型差异明显。地区上也存在共性,例如,海南和广东地区分离株对环丙沙星、恩诺沙星、诺氟沙星抗生素敏感性高,对阿莫西林、呋喃唑酮的耐药率高,含有5个相同的耐药谱型A(FUR/AMO)、C(AMO)、R(FUR/T/S/CEF/RIF/AMO)、M(FUR/T/S/CEF/RIF/CHL/TET/AMO)、J(RIF/AMO)。研究发现伊朗波斯湾的海产品分离的哈维弧菌中97.2%的菌株对氨苄青霉素耐药,83.3%菌株对庆大霉素耐药,77.7%对青霉素耐药[19],与该试验得到的耐青霉素的结果一致,但是该试验的菌株对庆大霉素是敏感的,表明地区的差异导致耐药谱的多样性。南海地区养殖鱼类2007年~2012年分离到的哈维弧菌耐药性的研究发现,海南、广东地区的分离株均对沙星类和磺胺类抗生素敏感,对头孢克肟、呋喃唑酮、阿莫西林有较高耐药率[30]。对比2012年以前,同一地区分离株对呋喃唑酮耐药率升高,而对庆大霉素、四环素敏感性有一定升高。在南海地区海水鱼病原菌对头孢克肟、呋喃唑酮、阿莫西林等药物高度耐药,有可能是早期药物的使用导致高耐药菌的产生,高耐药菌常携带大量耐药基因和质粒,会导致耐药性在养殖海域内扩散,危害海域环境微生态安全,对可持续的水产养殖造成威胁。
从宿主看,同一宿主分离到的哈维弧菌耐药谱也具有多样性,且宿主间耐药谱差异明显(图 2)。然而,在不同宿主间也存在着相同的耐药谱,例如棕点石斑鱼、珍珠龙趸和浅色黄姑鱼有A(FUR/AMO)、C(AMO)和G(FUR/RIF/AMO)3种共同的耐药谱。对军曹鱼养殖水体及肠道弧菌耐药性研究发现,氯霉素和庆大霉素对弧菌的抑制效果最佳[31];对海水养殖鱼分离的哈维弧菌耐药性研究发现,青石斑鱼、棕点石斑鱼、卵形鲳鲹和珍珠龙趸耐药谱型分别有6、6、9和10种[31]。综上,从不同海水养殖鱼宿主上分离的哈维弧菌,对抗生素的耐药性有显著差异,呈现多样的耐药谱型。
3.3 哈维弧菌的耐药谱聚类分析
聚类分析是对于静态数据分析的一门技术[32],是把相似的对象通过静态分类的方法分成不同的组别或者更多的子集,让在同一个子集中的成员对象都有相似的一些属性,常见的包括在坐标系中更加短的空间距离等[33]。例如对15株鱼诺卡氏菌聚类分析结果显示,15个菌株在T=23时可分为2个组群,在T=11时2个组群,分别可分为2个和4个亚群[34]。该试验以12种抗生素抑菌圈的直径作为变量,采用SPSS 19.0做耐药性的聚类分析。结果显示将90株哈维弧菌分为3个大群,5个亚群。在欧式距离平方为2时,亚群ⅰ、ⅱ聚类为Group Ⅰ;而达到3时,亚群ⅲ、ⅳ、ⅴ聚类为Group Ⅱ;在距离为5时两个大群合在一起。另外还有4株菌相距远,不能完成聚类,归为其他类。
哈维弧菌耐药谱研究表明,耐药谱聚类简单,在欧式平方为1时分为6个亚群,进而合为2个组群[30]。随着年份的改变,聚类分析更复杂、多样,除了2个主要的组群Group Ⅰ和Group Ⅱ之外,还存在着4株难以聚类的哈维菌株。总体上,组群Group Ⅱ的耐药率要高于Group Ⅰ,而所有菌株对FUR和AMO的抗性均较高。而Group Ⅰ与Group Ⅱ 2个组群对抗生素的敏感性主要在T/S、GEN、CEF、NOR、RIF、CHL、TET、CIP和ENR抗生素存在着明显的差异,且Group Ⅰ明显高于Group Ⅱ。通过对哈维弧菌抗生素敏感性实验研究,利用SPSS软件聚类分析能较好地将耐药性相近的菌株归为一类。
-
![]()
图 1 北部湾二长棘犁齿鲷3个时期体长频率分布的变化
Figure 1. Variation in length frequency of E.cardinalis in Beibu Gulf during three periods
![]()
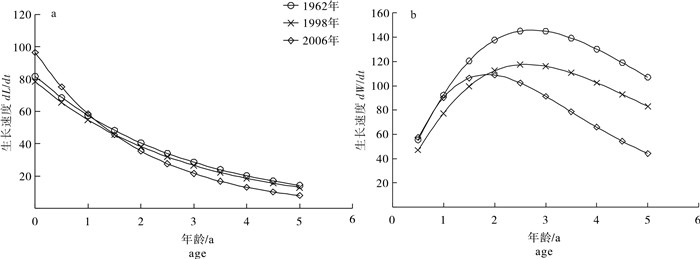
图 2 3个时期北部湾二长棘犁齿鲷体长(a)和体质量(b)生长速度的变化
Figure 2. Variation in growth rates of body length (a) and weight (b) of E.cardinalis in Beibu Gulf during three periods
![]()
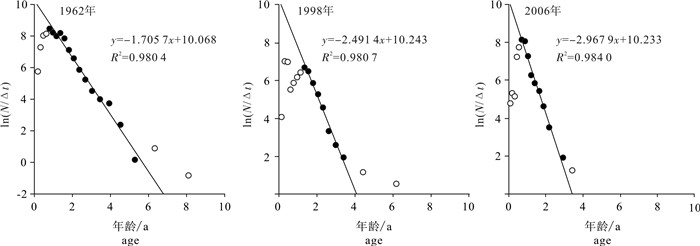
图 3 线性渔获量曲线法估算的北部湾二长棘犁齿鲷3个时期总死亡系数
Figure 3. Estimated total mortality coefficient of E.cardinalis in Beibu Gulf during three periods based on length converted catch curve
![]()
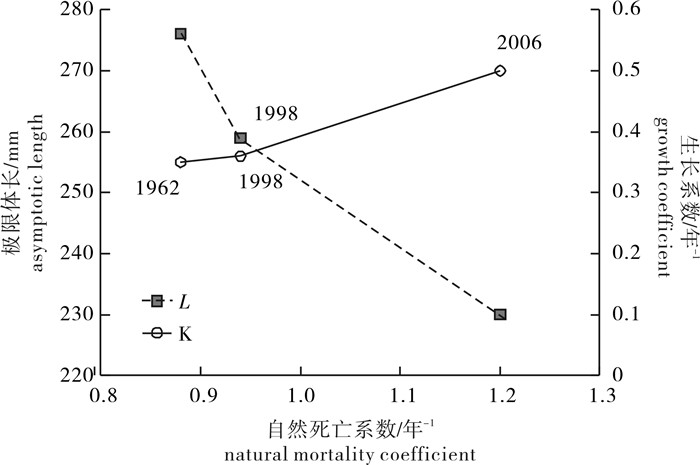
图 4 3个时期北部湾二长棘犁齿鲷极限体长、生长系数与自然死亡系数的关系
Figure 4. Relationship between asymptotic length, growth coefficient and natural mortality coefficient of E.cardinalis in Beibu Gulf during three periods
![]()
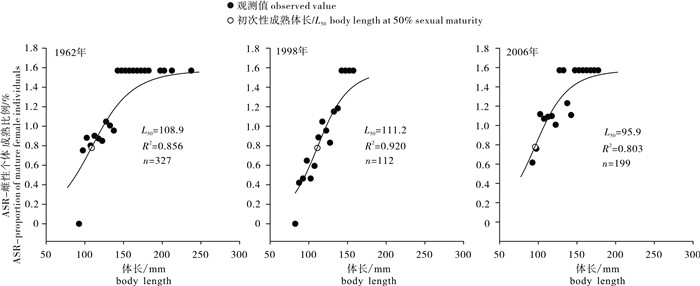
图 5 3个时期北部湾二长棘犁齿鲷雌性个体性成熟比例观测值与ASR模型拟合的逻辑斯蒂曲线
Figure 5. Observed proportion of mature individuals and logistic curve fitted by ASR model of E.cardinalis in Beibu Gulf during three periods
![]()
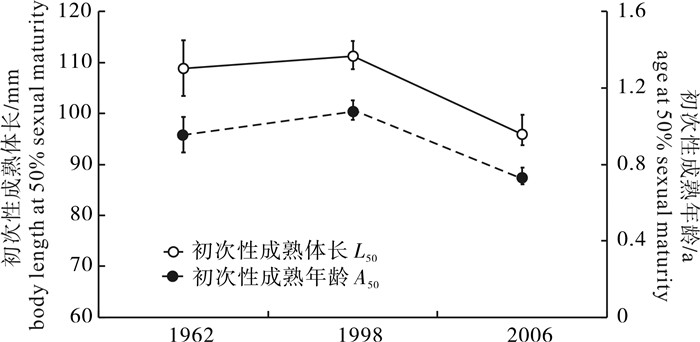
图 6 3个时期北部湾二长棘犁齿鲷雌性个体的L50与A50的变化趋势
误差线为95%置信区间。
Figure 6. Variations in L50 and A50 of female E.cardinalis in Beibu Gulf during three periods
Error bars show the 95% confidence intervals.
表 1 3个时期北部湾二长棘犁齿鲷的群体结构
Table 1 Population structure of E.cardinalis in Beibu Gulf during three periods
采样时间
sampling year尾数
number性比
(♀/♂)
sex ratio体长/mm body length 体质量/g body weight 范围
range均值
mean优势组
dominant范围
range均值
mean优势组
dominant1962年 5 201 1.24 40~240 108.8 91~100;121~130 3.0~589 68.2 31~40;71~90 1998年 1 120 0.82 57~230 110.2 61~80;121~140 3.1~436 70.2 11~20;81~110 2006年 2 055 2.33 41~196 102.0 91~110 2.8~281 48.5 21~60  下载: 导出CSV
下载: 导出CSV
表 2 3个时期北部湾二长棘犁齿鲷死亡参数的变化
Table 2 Variation in mortality parameters of E.cardinalis in Beibu Gulf during three periods
年份year 1962年 1998年 2006年 总死亡系数Z total mortality coefficient 1.71 (1.55~1.87) 2.49 (2.21~2.78) 2.97(2.63~3.31) 自然死亡系数M natural mortality coefficient 0.88 0.94 1.20 捕捞死亡系数F fishing mortality coefficient 0.83 1.55 1.77 开发率E exploitation rate 0.49 0.62 0.60 注:括号内为95%置信区间。
Note:Values in brackets are 95% confidence interval.
下载: 导出CSV
表 3 中国近海4种典型鱼类平均体长、初次性成熟体长和生长系数变化趋势
Table 3 Variations in mean body length, first maturity body length and growth coefficient of four species in coastal waters of China
群体
stock年份
year平均体(肛)长/mm
mean body (anal) length初次性成熟体(肛)长
first maturity body (anal) length生长系数
growth coefficient文献
Reference黄海南部小黄鱼 1960s 245 184 0.26 LI等[9] L.polyactis in the southern Yellow Sea 1980s 144 129 0.40 张国政等[15] 1990s 118 144 0.48 2000s 123 124 0.56 东海带鱼 1960s 239 - 0.27 周永东等[10] T.japonicus in the East China Sea 1970s 227 - 0.31 王垚[26] 1980s 215 - 0.31 1990s 189 - 0.35 北部湾花斑蛇鲻 1960s 186 111 0.26 陈作志等[11] S.undosquamis in the Beibu Gulf 1992~1993 161 105 0.28 1997~1999 133 96 0.33 2000s 131 97 0.39 北部湾二长棘犁齿鲷 1960s 109 109 0.35 此研究 E.cardinalis in the Beibu Gulf 1990s 110 111 0.36 2000s 102 96 0.50 注:小黄鱼平均体长为雌性个体平均体长,若一个年代存在单独两年或以上数据则取平均值。
Note:The mean body length of L.polyactis was from female;the mean value was applied when one decade included separate data of two or more years.
下载: 导出CSV
-
[1] 伍汉霖, 邵广昭, 赖春福. 拉汉世界鱼类名典[M]. 基隆: 台湾水产出版社, 1999: 376-379. https://www.las.ac.cn/front/book/detail?id=47c09593707d56d90e314e16ffa9fe6a [2] 成泰庆, 郑葆珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987: 334-336. https://mmcs.cast.org.cn/kxjfc/kxjfc/cwx/zylzml1665/art/2023/art_46257a8c3e27463e92e833465620aa4a.html [3] 贾晓平, 李纯厚, 李永振, 等. 南海专属经济区和大陆架渔业生态环境与渔业资源[M]. 北京: 科学出版社, 2004: 535-542. https://www.las.ac.cn/front/book/detail?id=42fe2fc1c537854dac87610b13d7d5a0 [4] 陈作志, 邱永松. 北部湾二长棘鲷生长和死亡参数估计[J]. 水产学报, 2003, 27(3): 251-257. doi: 10.3321/j.issn:1000-0615.2003.03.010 [5] 陈作志, 邱永松. 北部湾二长棘鲷的资源变动[J]. 南方水产, 2005, 1(3): 26-31. doi: 10.3969/j.issn.2095-0780.2005.03.004 [6] 侯刚, 朱立新, 卢伙胜. 北部湾二长棘鲷生长、死亡及其群体组成[J]. 广东海洋大学学报(自然科学版), 2008, 28(3): 50-55. http://gdou.irtree.com/articles/article_detail.aspx?id=ba3cadb5edae4d40bdb3488eaaaf3a5a [7] 冯波, 侯刚, 卢伙胜, 等. 北部湾二长棘鲷资源开发保护策略研究[J]. 农业系统科学与综合研究, 2009, 25(1): 114-119. doi: 10.3969/j.issn.1001-0068.2009.01.024 [8] 詹秉义. 渔业资源评估[M]. 北京: 农业出版社, 1995: 25-120. [9] LI Z L, SHAN X J, JIN X S, et al. Long-term variations in body length and age at maturity of the small yellow croaker (Larimichthys polyactis Bleeker, 1877) in the Bohai Sea and the Yellow Sea, China[J]. Fish Res, 2011, 110(1): 67-74. doi: 10.1016/j.fishres.2011.03.013
[10] 周永东, 徐汉祥, 刘子藩, 等. 东海带鱼群体结构变动的研究[J]. 浙江海洋学院学报(自然科学版), 2002, 21(4): 314-320. https://cstj.cqvip.com/Qikan/Article/Detail?id=7353504&from=Qikan_Article_Detail [11] 陈作志, 邱永松, 徐姗楠, 等. 北部湾花斑蛇鲻生物学特征的演化[J]. 中国水产科学, 2012, 19(2): 321-328. http://www.fishscichina.com/zgsckx/article/abstract/4984?st=alljournals [12] 陈作志, 邱永松, 贾晓平, 等. 捕捞对北部湾海洋生态系统的影响[J]. 应用生态学报, 2008, 19(7): 1604-1610. https://wenku.baidu.com/view/582ea796dd88d0d233d46acd?fr=xueshu_top [13] 邱永松, 曾晓光, 陈涛, 等. 南海渔业资源与渔业管理[M]. 北京: 海洋出版社, 2008: 153-156. [14] CHEN Y, PALOHEIMO J E. Estimating fish length and age at 50% maturity using a logistic type model[J]. Aquat Sci, 1994, 56: 206-219. doi: 10.1007/BF00879965
[15] 张国政, 李显森, 金显仕, 等. 黄海中南部小黄鱼生物学特征的变化[J]. 生态学报, 2010, 30(24): 6854-6861. https://www.ecologica.cn/stxb/article/abstract/stxb200910281414 [16] 陈作志, 孔啸兰, 徐姗楠, 等. 北部湾深水金线鱼种群参数的动态变化[J]. 水产学报, 2012, 36(4): 584-591. http://scxuebao.ijournals.cn/scxuebao/article/pdf/20110807663?st=article_issue [17] 陈大刚. 渔业资源生物学[M]. 北京: 中国农业出版社, 1997: 57-58. https://www.pwsannong.com/booklib/bookIntroduce?SiteID=123&ID=18708832 [18] 殷名称. 鱼类生态学[M]. 北京: 中国农业出版社, 2000: 203-208. https://mooc1.chaoxing.com/mooc-ans/course/207225969.html [19] OLSEN E M, HEINO M, LILLY G R, et al. Maturation trends indicative of rapid evolution preceded the collapse of northern cod[J]. Nature, 2004, 428(6986): 932-935. doi: 10.1038/nature02430
[20] QUINN T J, HODGSON S, FLYNN L, et al. Directional selection by fisheries and the timing of sockeye salmon (Oncorhynchus nerka) migrations[J]. J Appl Ecol, 2007, 17(3): 731-739. https://ui.adsabs.harvard.edu/abs/2007EcoAp..17..731Q
[21] 陈再超, 刘继兴. 南海经济鱼类[M]. 广州: 广东科技出版社, 1982: 1-266. https://book.sciencereading.cn/shop/book/Booksimple/show.do?id=BBD16415ED1F4B85DE053010B0A0A034A000 [22] 孙典荣, 林昭进. 北部湾主要经济鱼类资源变动分析及保护对策探讨[J]. 热带海洋学报, 2004, 23(2): 62-68. https://d.wanfangdata.com.cn/periodical/ChpNaW5lclBlcmlvZGljYWxDSEkyMDIyMTIwNhINcmRoeTIwMDQwMjAwOBoING4zZDlsc2s%3D [23] 王跃中, 袁蔚文. 南海北部底拖网渔业资源的数量变动[J]. 南方水产, 2008, 4(2): 26-33. https://www.schinafish.cn/article/id/c0e14e66-314f-438e-a9bc-dc850a20add3 [24] QIU Y S, LIN Z J, WANG Y Z. Responses of fish production to fishing and climate variability in the northern South China Sea[J]. Prog Oceanogr, 2010, 85(3): 197-212. http://www.onacademic.com/detail/journal_1000035078018610_06c2.html
[25] 朱晓光, 房元勇, 严力蛟, 等. 高捕捞强度环境下海洋鱼类生态对策的演变[J]. 科技通报, 2009, 25(1): 51-55. https://www.zhangqiaokeyan.com/academic-journal-cn_detail_thesis/0201297985461.html [26] 王垚. 伏季休渔条件下东海带鱼资源评估[D]. 舟山: 浙江海洋学院, 2010: 1-51.https://cdmd.cnki.com.cn/Article/CDMD-10340-2010107863.htm [27] GULLAND J A. Fish stock assessment: a manual of basic methods[M]. New York: Wiley Online Library, 1983: 1-220. doi: 10.1016/0006-3207(91)90020-A
[28] WANG X H, QIU Y S, DU F Y, et al. Population parameters and dynamic pool models of commercial fishes in the Beibu Gulf, northern South China Sea[J]. Chin J Oceanol Limnol, 2012, 30(1): 102-117. doi: 10.1007/s00343-012-1017-y
[29] 邹建伟, 林丕文, 王强哲. 南海北部湾2012年捕捞产量估算[J]. 南方水产科学, 2013, 9(4): 75-81. doi: 10.3969/j.issn.2095-0780.2013.04.013?viewType=citedby-info -
期刊类型引用(19)
1. 姜芳燕,谢浪萍,杨宁,黄海. 金鲳鱼肠炎病的病原菌分离鉴定及其中草药防治. 信阳师范学院学报(自然科学版). 2024(02): 228-233 .  百度学术
百度学术
2. 张婷,王瑞旋,孙敬锋,黄嘉健,林小植,周飞,马细兰. 哈维弧菌对皱纹盘鲍不同组织内细菌群落结构的影响. 热带海洋学报. 2023(01): 56-65 . 百度学术
3. 范嗣刚,孙珍珠,郭靖,吴昊,邓维德,李翔,黎源,陈燕飞,文菁. 日本对虾IKKβ基因克隆及其表达分析. 南方农业学报. 2023(06): 1837-1846 . 百度学术
4. 滕爽爽,张炯明,张利兵,谢尚庶,王瑶华,肖国强,杨千元,方军. 泥蚶致病性哈维氏弧菌的分离与鉴定. 水产科学. 2022(04): 547-555 . 百度学术
5. 陈秀程,王江勇,孙敬锋. 盐度对哈维弧菌耐药性的影响及耐药基因gyrB的表达实验研究. 生态科学. 2022(06): 123-129 . 百度学术
6. 王楠,尹纪元,王英英,李莹莹,吴斯宇,石存斌,李家豪,曹际振,王庆. 草鱼源乳酸菌的分离鉴定及其生物学特性研究. 南方水产科学. 2021(06): 74-84 . 本站查看
7. 何嘉乐,叶仕根,赵小然. 哈维氏弧菌生物膜生成的影响因素研究进展. 水产养殖. 2021(12): 12-17 . 百度学术
8. 魏文娟,赵姝,王元,周俊芳,李新苍,房文红. 副溶血弧菌养殖对虾分离株耐药性及耐药基因分析. 南方水产科学. 2020(01): 9-16 . 本站查看
9. 刘晓靖,王瑞旋,凌慧,姚托,王江勇. 方斑东风螺“急性死亡症”病原及其毒力基因研究. 海洋环境科学. 2019(01): 7-15 . 百度学术
10. 林彦伶,梁枫,胡高垚,韦孟咪,许士兰,龙文芳. 亚抑菌浓度抗生素对海水浴场来源肺炎克雷伯菌的接合及intI突变影响. 中国卫生检验杂志. 2019(04): 389-392+399 . 百度学术
11. 赵小慧,苏洁,樊景凤,马悦欣. 海洋环境中细菌耐药性研究进展. 中国抗生素杂志. 2019(04): 406-412 . 百度学术
12. 陈凯,朱璐丹,谭宏亮,黄东宇,习丙文,潘良坤,谢骏. 2株乳酸菌抑菌作用研究及安全性评价. 南方水产科学. 2019(05): 118-125 . 本站查看
13. 纪佳丽,刘广锋,冯娟,杨文斌,管淑玉. 广藿香对耐药哈维氏弧菌生物膜形成及相关基因的影响研究. 广东药科大学学报. 2018(02): 239-243 . 百度学术
14. 黄艳华,马沙,韩书煜,黄伟德,韦慕兰,蒙兰丽,吕小丽,覃惠明,李德壮,梁静真,黄钧. 2011~2016年广西罗非鱼源无乳链球菌的耐药谱型分析. 南方农业学报. 2018(05): 1023-1031 . 百度学术
15. 贝蕾,苏友禄,赵超,徐力文,刘广锋,王雨,郭志勋,冯娟. 哈维弧菌rbsB基因的克隆与表达. 南方水产科学. 2018(02): 75-82 . 本站查看
16. 李富祥,王鹏飞,闫路路,邱丽华. 花鲈irak4基因cDNA的克隆与表达分析. 南方水产科学. 2018(05): 70-79 . 本站查看
17. 钟志鸿,王菲,陈永贵,邓恒为,王世锋,孙云,陈雪芬,郭伟良,周永灿. 基于响应面法分析乌梅和抗生素对哈维氏弧菌的协同抑制作用. 南方水产科学. 2018(06): 81-88 . 本站查看
18. 刘开放,步君,章翔,谢珍玉. 华南地区海水养殖水域中哈维氏弧菌的抗药性分析. 海南大学学报(自然科学版). 2017(02): 131-139 . 百度学术
19. 蒋魁,徐力文,苏友禄,马红玲,刘广锋,郭志勋,高芳,冯娟. 两株珍珠龙趸病原性哈维弧菌(Vibrio harveyi)的分离与鉴定. 生态科学. 2017(06): 16-24 . 百度学术
其他类型引用(7)

计量
- 文章访问数: 3319
- HTML全文浏览量: 187
- PDF下载量: 974
- 被引次数: 26
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
