
Acoustic assessment of silver carp and bighead carp in Qinghe Reservoir: comparing echo-counting and echo-integrating methods
-
摘要:
2009年7月下旬使用挪威Simrad公司EY60型(70 kHz)分裂式波束科学鱼探仪对清河水库进行了鲢(Hypophthalmichthys molitrix)和鳙(Aristichthys nobilis)的资源声学调查。使用声学处理软件Echoview 4.0进行声学回波数据的后处理,研究库区内鲢和鳙昼夜分布特性,尝试使用回波计数法与回波积分法进行资源密度的评估,并进行了对比分析,讨论适合于水库鲢和鳙的声学调查方法。结果表明,鲢和鳙分布于水库10 m附近水层,昼间较为集中,单体回波较少,夜间较为分散,可以识别的单体回波较多。深水区域个体较大且密度较低,较浅的区域个体较小且密度较大。声学计数评估更适合夜间进行。根据夜间数据,2种评估方法的结果基本一致,回波积分法获得鱼类密度为0.063 ind · m-3,回波计数法为0.049 ind · m-3,可以确定2种方法均适合鲢和鳙的调查。
Abstract:We conducted an acoustic assessment of silver carp (Hypophthalmichthys molitrix) and bighead carp (Aristichthys nobilis) resources in Qinghe Reservoir in late July of 2009 by using the Simrad EY60 split-beam echosounder. With the software Echoview 4.0, we post-processed the data to analyze the distribution of the species during day and night. Besides, we assessed the resource density by using echo-counting and echo-integrating methods, which were then compared to find out the suitable acoustic investigation method for the two species. The results show that both species gather around 10 m in the reservoir and gather more usually during the day with less single echo but disperse at night with more single echo. Large fish gather in deep water area with relative lower density but small fish in shallow water area with high density. The echo-counting method is more suitable for night assessment, and according to the night data, the estimated density is 0.063 ind · m-3 and 0.049 ind · m-3 obtained by echo-integrating and echo-counting methods, respectively.
-
清河水库位于辽宁省铁岭市清河区,是辽宁七大著名水库之一,自东向西略呈扁“V”形,水库最宽处(大坝附近)约1 500 m,最窄处(龟山)约100 m,东西长约15 km,南北宽约2.5 km,最大库容9.71×108 m3,正常养鱼水面3 000 hm2,平均深度11.8 m,最大深度40 m,透明度35.4 cm,淤泥沙底质,是一座以防洪、灌溉为主,结合发电、养鱼等综合利用的大型水利工程。鲢(Hypophthalmichthys molitrix)和鳙(Aristichthys nobilis)为该水库主要增殖品种,比例约占70%~80%。每年10月份向水库投放鱼苗125 t,其中鲢占70%,鳙占30%,投苗规格为50 g · 尾-1,年捕捞量为350~500 t。近年也进行了池沼公鱼(Hypomesus olidus)和银鱼(Hemisalanx prognathus)等品种的增殖放养。因此,确定不同鱼种放养比例获得较高的经济效益,以及进行科学增殖和保持水库渔业可持续发展成为当前研究的重要课题,而准确、高效评估库区内渔业资源现存量是制定增殖计划和科学管理的基础。
传统的大水面淡水渔业资源评估方法主要以网具进行定点站位采样为主,由于网具有鱼种和规格选择性,且采样工作效率较低,直接影响到资源评估的有效性。近年来,新兴的渔业资源声学评估方法在欧、美、日等国家得到了广泛的应用,已成为渔业发达国家海洋渔业资源TAC(总允许捕捞量,total allowable catch)管理的重要参考依据。
中国自1989年起先后对海洋与淡水渔业资源进行了声学调查研究[1-9]。但是,由于不同水域的鱼种及其分布和规格存在差异,针对不同鱼种展开针对性研究不仅是中国渔业水声学发展的必经之路,也是区别于欧美等国家的特点之一。该研究以清河水库为试点,采用水声学测量方法针对北方水库中主要增殖品种鲢和鳙进行资源评估,获得其分布和资源量等信息,并讨论适合的声学调查方法,以期为水库渔业生产和资源管理提供科学依据。
1. 材料与方法
1.1 调查区域及航线
图 1为清河水库的卫星地图,其中P点(42°31'22.72"N,124°17'19.22"E)是水库大坝处的参考点。清河水库龟山(图 1中G点处)以东水深较浅(一般小于5 m),幼鱼分布较多;龟山以西水深相对较深(10~40 m之间,平均12 m左右),根据以往2次调查结果看,此处是鲢和鳙成鱼主要集中分布区。调查范围设定为大坝至龟山深水水域。
![]() 图 1 清河水库声学调查航线,虚线表示昼间航线,实线表示夜间航线Figure 1. Survey routes in Qinghe Reservoir; dotted line indicates day route and solid line indicates night route
图 1 清河水库声学调查航线,虚线表示昼间航线,实线表示夜间航线Figure 1. Survey routes in Qinghe Reservoir; dotted line indicates day route and solid line indicates night route调查分昼间(10:30~11:55)和夜间(18:35~20:20)进行。受左侧大坝到龟山之间夏季所设置的3个三角形定置网具的影响,调查航线采用非固定间距平行航线(图 1)。昼间航线间距约1 km,夜间航线间距最大2 km,最小0.5 km。利用10 m左右的小型平底作业船只进行走航测量,平均航速约4 kn。调查当日天气良好,风浪较小,夜间航行没有光照。航线使用GPS导航仪(GPSmap 60CSx型,Garmin)的定位数据绘制(图 1)。数据通过NMEA格式输出进行保存。调查的水域面积约6.5×106 m2,体积约4.5×107 m3,根据换能器的指向角(7°)、调查线的长度及水深,计算出声学采样体积与调查水域体积之比,白天5.3×10-4,夜间3.9×10 -4。
1.2 调查仪器及数据处理软件
使用分裂式波束科学鱼探仪EY60型(70 kHz)进行声学测量。换能器安放在流线型导流罩内,使用钢管固定悬挂于船侧,吃水1 m。相关参数设置见表 1。使用ER60软件进行原始数据(raw data)格式保存,在后处理过程中,使用Myriax公司的Echoview软件进行数据计算分析。处理方法为:1) 设置表层和底层2条线来屏蔽掉近场回波(表层2 m)和底层附近的不明回波;2) 体积散射强度(volume backscattering strength,SV)和目标强度(target strength,TS)回波图(echogram)的最小阈值设置为-75 dB;在此基础上再屏蔽掉不明的弱散射群;3) 使用TS回波数据进行单体回波检测,确定其TS的分布并进行回波计数,使用SV回波图进行回波积分处理。
表 1 EY60声学测量的设定参数Table 1. Parameter setting for EY60 acoustic detection参数parameter 设定值setting value 换能器频率frequency 70 kHz 换能器指向角directivity angle 7° 脉冲宽度pulse length 0.256 ms 发射功率transmitting power 500 W 声速sound speed 1 499.26 m·s-1 1.3 科学鱼探仪的校正
调查前选水深30 m左右水域,使用直径38.1 mm的钨钢合金标准球进行系统收发增益和等效波束角的校正。具体步骤为使用普通5号(直径0.37 mm)钓鱼线进行标准球的2点V字形悬挂,放置在换能器下方10 m处,按照EY60型科学鱼探仪的校正程序进行声轴位置的系统收发增益校正,以及将标准球放置在波束角内不同方位进行等效波束角的校正。标准球的TS参考值,根据水温和盐度所计算的声速按FOOTE和MacLENNAN[10]的方法获得。最后将校正文件保存并对科学鱼探仪进行升级操作。
1.4 鲢和鳙资源的声学评估方法
渔业资源声学评估方法一般有回波积分法和回波计数法2种,其中回波积分法主要运用体积混响测量的原理,即对于1个分布远大于波束范围的鱼群,鱼群的回波可以被认为是体积散射。其平均强度值SV正比于鱼群内个体的平均TS和个体的密度。即SV可以被近似为个体的后向散射截面(backscattering cross section,简称σbs)的非相干叠加。假设鱼群内部具有平均后向散射截面<σbs>,平均密度为:
$$ \langle\mathrm{n}\rangle=\langle\mathrm{sv}\rangle /\left\langle\sigma_{\mathrm{bs}}\right\rangle $$ (1) 在广域的实际评估调查中,考虑到不同大小的鱼群以及个体的不同分布,一般在一定的空间范围内对鱼群的SV进行积分,获得平均体积散射强度<sv>。虽然鱼类的后向散射截面受个体形状、体长规格、游泳姿态、内部组成等的影响较为复杂,但是分裂式波束科学鱼探仪可以进行现场测量,给出其平均值的分布。常用的TS是σbs的分贝值[11-12]。该研究根据鱼群回波基本连续的空间分布状况,积分按300 m左右间隔进行,获得不同区域鱼群的SV分布和密度n的分布。另一种回波计数法,是在一定的范围内对声学波束的扫描空间内部进行个体回波计数,根据波束扩展采样角度计算扫描水域的体积,获得该范围内的平均分布密度。波束的扩展角度根据比较不同深度单体回波的长度和船速求得。该研究同样选取300 m左右间隔,使用Echoview分别进行单体回波识别、计数和体积计算,最后获得密度的分布。
2. 结果
2.1 鲢和鳙的垂直和水平分布
使用Echoview处理的坝体附近水域的昼夜间SV回波图中包含表层的近场回波,单体鱼和群体回波以及不明弱混响区(下层面积较大的弱回波区)(图 2)。昼间调查显示了鱼群较为聚集的特征,鲢和鳙集中在8~10 m水层(图 2-a);在4~8 m区间,有少量的单体回波。夜间以个体回波为主,分散于4~10 m水层,集群回波较少(图 2-b),适合单体检测。因此,该研究主要针对夜间的回波图进行回波的积分、单体判别、回波计数以及资源分析评估。图 3显示了夜间各调查航线上每个积分单位(300 m)内的单体鱼个数(图 3-a)和平均SV分布(图 3-b)。在调查区域内鲢和鳙在大坝、龟山和北侧沿岸附近区域个体数量相对较多;龟山附近区域的SV值相对较高。通过对单体回波的TS测量及根据公式(1)可以确定,龟山区域的鲢和鳙个体相对较小且密度较高,中部和坝体附近水域个体较大且密度相对较低。
![]() 图 2 使用Echoview软件表示的清河水库水坝附近昼间(a)、夜间(b)SV回波图Figure 2. Echogram of SV around Qinghe Reservoir dam using Echoview software. day(a), night(b)
图 2 使用Echoview软件表示的清河水库水坝附近昼间(a)、夜间(b)SV回波图Figure 2. Echogram of SV around Qinghe Reservoir dam using Echoview software. day(a), night(b)![]() 图 3 夜间各调查航线单体计数值分布(a)及对应的平均SV分布(b)Figure 3. Distributions of individual fish number(a)and corresponding average SV(b)of survey routes at night
图 3 夜间各调查航线单体计数值分布(a)及对应的平均SV分布(b)Figure 3. Distributions of individual fish number(a)and corresponding average SV(b)of survey routes at night2.2 鲢和鳙资源量评估
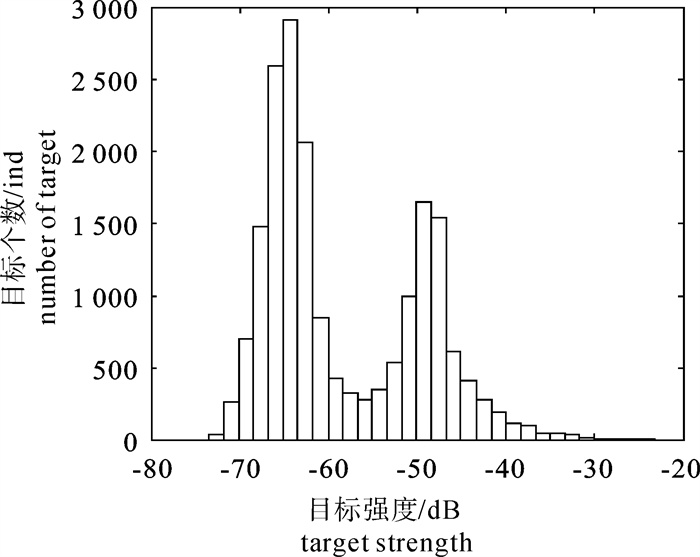
由于回波积分法需要TS值,该研究使用Echoview对各调查航线鲢和鳙分布水层检测到的单体回波进行了TS的现场测量。Echoview可以给出连续单体回波的TS最大值和算术平均值。TS的分布显示为典型的双峰型(图 4)。虽然没有进行网具采样,但是考虑鲢和鳙为有鳔鱼类,可以确定鲢和鳙的TS值集中在-48 dB附近,TS值小于-55 dB为浮游动物,TS值大于-55 dB为鱼类,因此选择大于-55 dB的TS值使用。计算调查水域内平均密度0.063尾· m-3。使用回波计数法计算的平均密度0.049尾· m-3。库区调查范围内的面积约6.5×106 m2,测量的水层深约7 m,计算获得体积约4.5×107 m3。不同方法对应的鲢和鳙分别为2.835×106尾和2.205×106尾。
![]() 图 4 单体回波检测的TS最大值分布图Figure 4. Distribution diagram of maximum TS value of single echo detection
图 4 单体回波检测的TS最大值分布图Figure 4. Distribution diagram of maximum TS value of single echo detection3. 讨论
3.1 调查航线与鱼类分布
最初的设计调查航线采用等间隔平行线,受库区内定置网具设置的影响,只能在走航调查过程中调整,但相对于一般的海洋调查,该研究调查范围很小。航线间隔也远小于一般海洋调查设置的8~10 nmile。从调查结果看,夜间库区内鲢和鳙的单体回波较为分散,区域内的鱼群回波较为连续,基本分布较为分散和连续,调查航线基本满足随机性和相邻航线非相关性的声学采样要求,能够较好地反映鲢和鳙在库区内的水平和垂直分布。对于鲢和鳙的水平分布,深水区域个体较大且密度较低,较浅的区域个体较小且密度较大;对于鲢和鳙的垂直分布,根据传统的捕捞经验,一般认为鲢和鳙夏季主要分布于10 m附近的水层,昼间调查的结果与之相吻合,但是夜间的分布更加分散,个体较大的鲢和鳙具有一定上浮的特点(图 2-b)。
3.2 回波积分法与回波计数法
回波积分法经过近30年的理论研究和技术发展已经成为渔业资源声学评估的主选方法。但是,由于受技术及其他多方面条件的制约,使用该方法进行调查的只能局限于少数研究单位,短期内在国内淡水渔业进行广泛推广困难较大。而回波计数法相对于回波积分法具有原理和技术较简单、使用普通的商用鱼探仪经过简单的改装也可以进行调查和调查成本较低等特点,易于被一般研究者接受。该研究的结果表明,回波积分法和使用Echoview进行的回波计数法的结果基本一致,可以确定2种方法均适合鲢和鳙的调查。该研究也随机抽取10个区域进行了手工计数和软件计数的对比,结果手工计数是软件计数的2倍左右。原因是软件计数根据TS的最小阈值限制了TS较小的单体回波计数,而手工计数时对TS较小的回波判断不准,导致计数结果增大。商用鱼探仪一般没有TS的测量功能,且信号放大倍数较高,所以回波的计数应当进行适当的比对标定,有关此方面的研究还有待于进一步确定。但是,一般商用鱼探仪的波束指向角较科学鱼探仪大,一般为15°左右,后者一般是7°左右,这样可以使单体回波更加易于目测和计数。
在使用Echoview时,可以通过限制积分区域的方法屏蔽非研究对象的回波,但是夜间对于浮游动物回波的控制还是较困难,因此,使用夜间数据进行回波积分处理时,会掺杂部分浮游动物的回波,导致评估结果增大。但该研究为了进行2种评估方法的比较而选用了夜间回波。
3.3 鲢和鳙的目标强度与资源量
该研究通过分裂式波束的现场测定给出了库区内鲢和鳙目标强度的比例分布(图 3),该调查选择了主要分布在10 m附近水层的鲢和鳙鱼类进行评估,由于TS的研究工作尚未展开,TS所对应体长的关系式尚不能确定。因鲢和鳙是有管双室鳔鱼类,国际上有关该方面的研究很少,仅HORNE等[13]对以色列某水库进行过一种体长15 cm左右鳊(Mtrogres terraesanctae)的TS研究。因此,有关鲢和鳙鱼类的TS研究有待今后进行细致分析。由于缺少TS的研究,不能分析对应于不同体长鲢和鳙的资源量组成,而是采取了计算平均值的方法进行推算。假定按每尾平均体质量1 kg和2×106尾进行推算,库区可捕鲢和鳙资源量大于2 000 t。库区近年来的年捕捞量为500 t,捕捞规格控制在2~4 kg ·尾-1,因此,可以判断库区鲢和鳙的可持续生产保持较好。
4. 结论
通过研究确定清河水库夏季鲢和鳙的水平分布较为分散,没有大规模鱼群聚集,昼间垂直分布于10 m水层,夜间分布于4~10 m水层。声学资源评估的回波积分法和回波计数法均可以使用,但回波计数法评估鲢和鳙鱼类的调查应选在夜间进行。商用鱼探仪有望使用回波计数法在鲢和鳙的调查中运用。库区的鲢和鳙资源量分布密度在0.05~0.06 ind · m-3。可以确定声学评估法效率较高,即便是增加环境调查的定点采样工作,整个清河库区的调查工作也可在1 d内完成,因此,声学资源评估方法对于水库和湖泊具有较高的推广价值。
致谢: 该研究得到了铁岭市清河水库管理局给予的大力支持和帮助,在此谨向铁岭市银州区李永贵、清河水库管理局薛云刚和李可义致谢。同时感谢日本北海道大学水产学部饭田浩二博士提供使用 Echoview数据处理软件方面的支持。 -
![]()
图 1 清河水库声学调查航线,虚线表示昼间航线,实线表示夜间航线
Figure 1. Survey routes in Qinghe Reservoir; dotted line indicates day route and solid line indicates night route
![]()
图 2 使用Echoview软件表示的清河水库水坝附近昼间(a)、夜间(b)SV回波图
Figure 2. Echogram of SV around Qinghe Reservoir dam using Echoview software. day(a), night(b)
![]()
图 3 夜间各调查航线单体计数值分布(a)及对应的平均SV分布(b)
Figure 3. Distributions of individual fish number(a)and corresponding average SV(b)of survey routes at night
![]()
图 4 单体回波检测的TS最大值分布图
Figure 4. Distribution diagram of maximum TS value of single echo detection
表 1 EY60声学测量的设定参数
Table 1 Parameter setting for EY60 acoustic detection
参数parameter 设定值setting value 换能器频率frequency 70 kHz 换能器指向角directivity angle 7° 脉冲宽度pulse length 0.256 ms 发射功率transmitting power 500 W 声速sound speed 1 499.26 m·s-1  下载: 导出CSV
下载: 导出CSV
-
[1] 赵宪勇, 陈毓桢. 狭鳕(Theragra chalcogramma Palas) 目标强度的现场测定[J]. 中国水产科学, 1996, 2 (4) : 19-27. https://xueshu.baidu.com/usercenter/paper/show?paperid=1n6s0e30rj330m80aw3t0mg0sr157335&site=xueshu_se&hitarticle=1 [2] 唐启升, 王为祥, 陈毓桢, 等. 北太平洋狭鳕资源声学评估调查研究[J]. 水产学报, 1995, 19 (1) : 8-20. [3] 李永振, 陈国宝, 赵宪勇, 等. 南海北部海域小型非经济鱼类资源声学评估[J]. 中国海洋大学学报, 2005, 35(2): 206-212. doi: 10.3969/j.issn.1672-5174.2005.02.007 [4] 陈国宝, 李永振, 赵宪勇, 等. 南海5类重要经济鱼类资源声学评估[J]. 海洋学报, 2006, 28 (2) : 128-134. doi: 10.3321/j.issn:0253-4193.2006.02.016 [5] 谭细畅, 史建全, 张宏, 等. EY60回声探测仪在青海湖鱼类资源量评估中的应用[J]. 湖泊科学, 2009, 21 (6) : 865-872. [6] 张慧杰, 杨德国, 危起伟, 等. 葛洲坝至古老背江段鱼类的水声学调查[J]. 长江流域资源与环境, 2007, 16 (1) : 86-91. https://xueshu.baidu.com/usercenter/paper/show?paperid=676157be3348d8ba97d5ab60f58b7a7e&site=xueshu_se [7] 陶江平, 陈永柏, 乔晔, 等. 三峡水库成库期间鱼类空间分布的水声学研究[J]. 水生态学杂志, 2008, 1 (1) : 25-33. https://xueshu.baidu.com/usercenter/paper/show?paperid=e4a400bdedd1f8ec9d0332bdab20c9e1&site=xueshu_se [8] 谭细畅, 夏立启, 立川贤一, 等. 东湖放养鱼类时空分布的水声学研究[J]. 水生生物学报, 2002, 26 (6) : 585-590. doi: 10.3321/j.issn:1000-3207.2002.06.002 [9] 谭细畅, 李新辉, 赖子尼, 等. 青皮塘产卵场广东鲂繁殖群体的水声学探测研究[J]. 生态学杂志, 2008, 27 (5) : 785-790. [10] FOOTE K G, MacLENNAN D N. Comparison of copper and tungsten carbide calibration spheres[J]. J Acoust Soc Am, 1984, 75 (2): 612-616. doi: 10.1121/1.390489
[11] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. 中华人民共和国国家标准《海洋调查规范, 第6部分: 海洋生物调查GB/T 12766.6-2007. 》[S]. 北京: 中国标准出版社, 2008: 89-97. https://xueshu.baidu.com/usercenter/paper/show?paperid=106g06q0b11h00g0577w04q0qg095323&site=xueshu_se [12] SIMMONDS J, MacLENNAN D. Fisheries acoustics: theory and practice[M]. 2nd ed. Oxford: Blackwell Science, 2005: 51-53. doi: 10.1002/9780470995303
[13] HORNE J K, WALLINE P D, JECH J M. Comparing acoustic model predictions to in suit backscatter measurements of fish with dual-chambered swimbladders[J]. J Fish Biol, 2000, 57(5): 1105-1121. doi: 10.1111/j.1095-8649.2000.tb00474.x
-
期刊类型引用(19)
1. 张丽媛,杨剑虹,熊清海,田敏,王慧,曲品,蒋荣明,寇春妮,武智. 基于水声学的阳宗海鱼类行为特征及其资源评估. 南方水产科学. 2024(01): 110-119 .  本站查看
本站查看
2. 李栋,蔺丹清,王召根,李佩杰,潘杰,叶昆,徐跑,刘凯. 基于水声学频差技术的镇江长江豚类省级自然保护区鱼类资源时空特征. 水生生物学报. 2023(01): 121-132 . 百度学术
3. 汤勇. 中国渔业资源声学评估研究与进展. 大连海洋大学学报. 2023(02): 185-195 . 百度学术
4. 孙扬,汤勇,邢彬彬,李泓泉,毕福洋,马壮. 基于基尔霍夫射线模型法的高白鲑目标强度研究. 大连海洋大学学报. 2021(02): 310-316 . 百度学术
5. 王思凯,李九奇,刘永虎,刘崇焕. 海洋牧场开发的设备需求分析. 安徽农学通报. 2020(09): 92-94 . 百度学术
6. 徐川,胡正春. 核电厂冷源安全渔业资源声学调查. 电力安全技术. 2020(10): 55-59 . 百度学术
7. 孔德平,秦涛,范亦农,陈小勇. 邛海鱼类资源与空间分布的水声学调查. 水生态学杂志. 2019(01): 22-29 . 百度学术
8. 杨洋,朱国平. 海洋渔业资源声学评估技术及在南极磷虾资源声学评估中的应用. 海洋渔业. 2018(03): 368-376 . 百度学术
9. 荆丹翔,韩军,王杰英,王晓丁,徐志伟. 基于成像声呐的鱼类三维空间分布. 水产学报. 2018(06): 996-1005 . 百度学术
10. 武智,李新辉,李捷,陈方灿,朱书礼. 红水河岩滩水库鱼类资源声学评估. 南方水产科学. 2017(03): 20-25 . 本站查看
11. 李斌,汤勇,孙建富,李佩杰,胡亚东,张剑波. 基于声学方法的黄河三门峡水库渔业资源空间分布研究. 大连海洋大学学报. 2016(05): 563-571 . 百度学术
12. 李斌,李佩杰,汤勇,孙建富,胡亚东,张剑波. 黄河小浪底水库主河道水域渔业资源声学评估. 水产学报. 2015(08): 1134-1143 . 百度学术
13. 屈泰春,陈帅,黄洪亮,汤勇,刘健,吴越,李灵智,冯春雷,戴阳. 南极大磷虾声学调查数据噪声处理与资源密度评估. 极地研究. 2014(04): 451-459 . 百度学术
14. 张赞,蔺丹清,汤勇,孙建富,李九奇. 基于声学测量方法的大伙房水库鱼类资源季节变动特征. 南方水产科学. 2014(06): 12-19 . 本站查看
15. 吴斌,方春林,贺刚,傅培峰. FiSATⅡ软件支持下的体长股分析法探讨. 南方水产科学. 2013(04): 94-98 . 本站查看
16. 杨丽丽,何光喜,胡忠军,史文,刘其根,陈来生,王金鹏,杨卫明. 鲢鳙占优势的千岛湖浮游动物群落结构特征及其与环境因子的相关性. 水产学报. 2013(06): 894-903 . 百度学术
17. 屈泰春,黄洪亮,汤勇,陈帅,刘健,李灵智,冯春雷,吴越,徐冰. 渔业声学数据后处理中噪声剔除的研究进展. 渔业信息与战略. 2013(03): 208-213 . 百度学术
18. 牟洪民,姚俊杰,倪朝辉,方贵镇,安苗,马珊. 红枫湖鱼类资源及空间分布的水声学调查研究. 南方水产科学. 2012(04): 62-69 . 本站查看
19. 任玉芹,王珂,段辛斌,阴双雨,李世健,刘绍平,陈大庆. 鳙目标强度和行为特征的水声学研究. 淡水渔业. 2011(02): 3-9 . 百度学术
其他类型引用(17)



计量
- 文章访问数: 4672
- HTML全文浏览量: 162
- PDF下载量: 2718
- 被引次数: 36
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
