
Optimization of high-pressure speece cone selection and accessories development of aquaculture ships
-
摘要:
优质的海水是深远海集约化养殖成功的前提,提升养殖水体的溶解氧对于高密度养殖至关重要。增氧锥是养殖水体最高效的增氧设备之一。针对深远海养殖工船空间紧凑的现状,对增氧锥外形进行优选,并开发相关配件,以达到高效增氧和节约空间的目的。采用流体仿真技术,利用其文丘里结构产生的射流、偏流、回流现象获得增氧效果;通过设计开发微孔曝气射流装置,借助微气泡原理和二次射流效果,进一步提升了增氧锥中氧气溶于水的效率。流体仿真软件结果显示,在其他条件相同的情况下,增氧锥截面角度为28° 时,液速峰值最高,产生的溶解氧效果最佳;有无曝气管射流的对比实验结果显示,加装曝气管的增氧锥增氧速度较快,该配件对于增氧锥的作业有良好的辅助功能,具有较好的应用价值。
Abstract:High-quality seawater is the prerequisite for success of intensive marine aquaculture, and improving dissolved oxygen of aquaculture water is crucial for high-density aquaculture. Speece cone is one of the most efficient oxygenating equipments in aquaculture water. In view of the current situation of compact space of offshore aquaculture ship, we optimized the selection of speece cone, and developed related accessories to achieve the purposes of efficient aeration and space saving. We applied fluid simulation technology to optimize the angle of speece cone, and reasonably used the jet and bias phenomenon caused by venturi structure to increase the dissolved oxygen effect. By the design and development of microporous aeration jet device with help of microbubble principle and secondary jet effect, we improved the efficiency of oxygen dissolved in water in the speece cone. According to the results of fluid simulation software, under the same other conditions, the speece cone section had the highest peak liquid velocity and the best dissolved oxygen at section angle of 28°. According to the comparative experimental results of whether there was an aeration jet device, the speece cone with aeration jet device was faster, and it is concluded that the accessory has good auxiliary function for operation of speece cone and has good value of application.
-
Keywords:
- Aquaculture ships /
- Speece cone /
- Fluid simulation /
- Preference appearance
-
硬骨鱼类的生长主要受生长轴,即生长激素(GH)/胰岛素样生长因子(IGF)的调控,GH为脑垂体分泌的单链肽类激素,通过生长激素受体(GHR)刺激肝脏等组织分泌IGF,进而发生生物学效应。GH居于生长轴的中心位置,在鱼类的个体生长中起关键性作用[1]。研究表明鱼类垂体GH基因的转录和转译水平受性别[2-5]、季节和生理状态[4-7]及外界理化因子[8-10]等因素的影响,且研究结果显示在一定条件下垂体GH基因转录或转译水平高的鱼类其生长速度也快。因此,GH成为研究鱼类生长性能的重要指标。
高密度养殖是提高单位面积产量、实现更大经济效益的有效手段,但不可避免地影响养殖对象的生长速度和健康。而革胡子鲶(Clarias gariepinus)比其他养殖鱼类对高密度养殖的适应性强[11-16]。天津市德仁农业发展有限公司设施化养殖的革胡子鲶,在高密度下仍然能够快速生长(最高产量可达400 kg · m-3水体),推测与其内在的生长调控因子有关。文章通过分析不同养殖密度下革胡子鲶幼鱼的日增质量和垂体GH基因mRNA的表达水平,初步探讨革胡子鲶幼鱼在不同养殖密度下生长差异和生长差异的内在原因,以期为深入探究革胡子鲶的生长调节以及高密度养殖快速生长的机理奠定基础。
1. 材料与方法
1.1 试验材料
试验用鱼为天津市德仁农业发展有限公司从福建漳州购进约1.5 g的革胡子鲶饲养而成,选取大小规格相近的幼鱼为试验对象,平均体质量为10~12 g。
1.2 养殖试验
试验分组及饲养管理同DAI等[17]。即试验用革胡子鲶幼鱼取回后在天津农学院天津市水产生态及养殖重点实验室于水族箱(70 cm×50 cm×50 cm)中暂养2周后,选取发育良好、体质健康的个体[平均体质量为(30.71±0.89)g]用于该试验。试验鱼随机分为35 kg · m-3、65 kg · m-3、95 kg ·m-3和125 kg · m-3 4个密度组,每组3个重复,饲养于水族箱(70 cm×50 cm×50 cm)中,实际水体积为120 L。模拟天津市德仁农业发展有限公司水泥池饲养革胡子鲶幼鱼的实际养殖条件,每日10: 00和18: 00各换水一次,每次换水量为水体积的3/4;养殖水温保持在(27±1)℃。按体质量的2%每天8: 00和16: 00各投喂一次,所用饲料为天津市天祥水产有限公司生产的鲶鱼膨化配合饲料(粗蛋白35%、粗脂肪5%、粗纤维10%、灰分10%)。养殖试验为期60 d,试验期间水体积保持不变,每30 d取样一次。每天观察并记录鱼的摄食及死亡情况。
1.3 相关指标分析方法
1.3.1 革胡子鲶生长的测定
分别于第30和第60天各分析一次。分析时停饲24 h,并以200 mg · L-1的MS-222麻醉,以水族箱为单位称量鱼体的总质量,同时计数鱼体数量,根据下列公式计算鱼的均质量和日增质量:
鱼的均质量=试验结束时每箱鱼总质量(g)/鱼的数量(尾)
日增质量=[试验结束时鱼的均质量(g · 尾-1)-试验开始时鱼的均质量(g · 尾-1)]/养殖天数
1.3.2 GH基因mRNA表达的半定量分析
养殖第30和第60天时,每个养殖密度各取经MS-222麻醉的鱼18尾(每个水族箱取鱼6尾),解剖取出垂体后立即放入液氮中,之后于-80 ℃保存备用。以β-肌动蛋白基因作为内参基因分析垂体GH基因mRNA的相对表达量。试验中以个体为单位,根据王晓梅等[18]报道的方法进行垂体总RNA的提取、cDNA第一条链的合成、GH基因ORF区段和β-肌动蛋白基因片段的扩增以及GH基因mRNA相对表达量分析。
1.3.3 数据统计分析
试验数据以“平均值±标准差”(X+SD)表示,数据采用SPSS 11.5统计软件进行单因素方差分析(One-Way ANOVA),LSD法多重比较各组间数值的差异显著性(P<0.05)。
2. 结果
2.1 生长分析
根据革胡子鲶在不同密度养殖30 d和60 d后体质量(表 1)计算鱼体在0~30 d以及30~60 d 2个养殖期的日增质量。结果表明,试验期间随着养殖密度的增大出现日增质量降低的趋势(图 1),统计分析结果显示,在0~30 d的养殖期间,35 kg ·m-3和65 kg · m-3密度组的日增质量差异不显著(P>0.05),95 kg · m-3和125 kg · m-3密度组的日增质量差异也不显著(P>0.05);但35 kg · m-3和65 kg · m-3密度组的日增质量显著高于95 kg · m-3和125 kg · m-3密度组(P<0.05);养殖30~60 d期间,35 kg · m-3和65 kg · m-3密度组的日增质量显著高于125 kg · m-3密度组(P<0.05),而其余各密度组间的日增质量差异均不显著(P>0.05)。
表 1 养殖密度对革胡子鲶体质量的影响Table 1. Effect of stocking density on body weight of juvenile C.gariepinus指标index 养殖密度/ kg·m-3 stocking density 35 65 95 125 初始体质量/g initial body weight 31.01±0.47 29.77±0.32 30.82±1.17 31.22±0.90 第30天体质量/g body weight on 30th day 64.75±1.21a 60.79±0.43ab 57.40±3.38bc 55.38±2.83c 第60天体质重/g body weight on 60th day 100.94±3.42a 96.64±3.84a 90.70±1.19b 82.87±2.71c 死亡率/% mortality 3.69±0.76a 3.68±1.13a 5.13±0.36b 5.40±0.43b 注:部分数据引自文献[17]。同一行肩标字母相同或无肩标字母者表示差异不显著(P>0.05),肩标字母不同者表示差异显著(P<0.05)
Note:Some data in this table are taken from Reference [17].Values with the same letters or no letter within the same row are not significantly different (P>0.05);whereas those with different letters are significantly different (P<0.05).![]() 图 1 不同养殖密度革胡子鲶日增质量同一线上肩标字母不同者为差异显著(P<0.05),肩标字母相同者为差异不显著(P>0.05)Figure 1. Daily weight gain of C.gariepinus reared at different stocking densitiesValues with different superscripts on the same lines are significantly different (P < 0.05), while those with the same superscripts are not significantly different (P > 0.05).
图 1 不同养殖密度革胡子鲶日增质量同一线上肩标字母不同者为差异显著(P<0.05),肩标字母相同者为差异不显著(P>0.05)Figure 1. Daily weight gain of C.gariepinus reared at different stocking densitiesValues with different superscripts on the same lines are significantly different (P < 0.05), while those with the same superscripts are not significantly different (P > 0.05).2.2 GH基因mRNA表达的半定量RT-PCR分析
2.2.1 总RNA分离结果
经1%琼脂糖凝胶电泳检测表明提取的革胡子鲶脑垂体总RNA完整性较好(图 2显示了4个RNA样本的电泳结果)。用核酸蛋白测定仪(德国Eppendorf公司出品,BioPhotometer型)测定出每个RNA样本光密度比(OD260/OD280)为1.80~2.00,表明总RNA纯度较好,RNA样本可以用于后续的试验。
![]() 图 2 革胡子鲶4个个体脑垂体总RNA电泳结果Figure 2. Electrophoretic patterns of total RNA extracts from pituitary of four individuals of C.gariepinus
图 2 革胡子鲶4个个体脑垂体总RNA电泳结果Figure 2. Electrophoretic patterns of total RNA extracts from pituitary of four individuals of C.gariepinus2.2.2 PCR扩增产物电泳检测结果
图 3-a和图 3-b分别为养殖第30和第60天时革胡子鲶部分个体GH和β-actin基因片段扩增后的电泳检测结果。
![]() 图 3 养殖第30天(a)和第60天时(b)革胡子鲶个体GH和β-actin基因片段PCR产物电泳图泳道1~3、4~6、7~8和9~12分别为养殖密度35 kg · m-3、65 kg · m-3、95 kg · m-3和125 kg · m-3的各3个体GH和β-actin基因片段RCR产物电泳图,泳道M为DNA标准分子量(BBI公司)Figure 3. Electrophoretic patterns of PCR products of GH and β-actin gene fragments of individuals on 30th (a) and 60th day nursing(b)Lanes 1~3, 4~6, 7~8 and 9~12 are electrophoretic patterns of PCR products of GH and β-actin gene fragments separately on three individuals at stocking densities of 35 kg · m-3, 65 kg · m-3, 95 kg · m-3 and 125 kg · m-3, respectively; Lane M is DNA Marker (BBI Company)
图 3 养殖第30天(a)和第60天时(b)革胡子鲶个体GH和β-actin基因片段PCR产物电泳图泳道1~3、4~6、7~8和9~12分别为养殖密度35 kg · m-3、65 kg · m-3、95 kg · m-3和125 kg · m-3的各3个体GH和β-actin基因片段RCR产物电泳图,泳道M为DNA标准分子量(BBI公司)Figure 3. Electrophoretic patterns of PCR products of GH and β-actin gene fragments of individuals on 30th (a) and 60th day nursing(b)Lanes 1~3, 4~6, 7~8 and 9~12 are electrophoretic patterns of PCR products of GH and β-actin gene fragments separately on three individuals at stocking densities of 35 kg · m-3, 65 kg · m-3, 95 kg · m-3 and 125 kg · m-3, respectively; Lane M is DNA Marker (BBI Company)2.2.3 GH mRNA表达半定量PCR分析的数据统计
利用GeneTools软件分析革胡子鲶个体GH(目标基因)和β-actin(内参基因)基因片段扩增后的电泳图谱,GH与β-actin基因片段扩增产物的信号强度比即为GH基因的相对表达量。
图 4为养殖第30和第60天时不同养殖密度下革胡子鲶GH基因mRNA平均相对表达量的统计分析结果。养殖第30天时,各养殖密度间革胡子鲶GH基因mRNA相对表达量差异均不显著(P>0.05);养殖第60天时,革胡子鲶GH基因mRNA相对表达量在35 kg · m-3和65 kg · m-3密度组间差异不显著(P>0.05),95 kg · m-3和125 kg · m-3密度组间差异也不显著(P>0.05),但35 kg · m-3和65 kg · m-3密度组GH基因mRNA的相对表达量显著高于95 kg · m-3和125 kg · m-3密度组(P<0.05)。
![]() 图 4 不同养殖密度革胡子鲶GH基因mRNA的相对表达量标有不同字母者为差异显著(P<0.05),无字母或字母相同者为差异不显著(P>0.05)Figure 4. Relative expression of GH mRNA in pituitary of C.gariepinus reared at different densitiesDifferent letters above the bar indicate significant difference (P < 0.05);while the same letters or no letter above the bar indicate no significant difference (P > 0.05)
图 4 不同养殖密度革胡子鲶GH基因mRNA的相对表达量标有不同字母者为差异显著(P<0.05),无字母或字母相同者为差异不显著(P>0.05)Figure 4. Relative expression of GH mRNA in pituitary of C.gariepinus reared at different densitiesDifferent letters above the bar indicate significant difference (P < 0.05);while the same letters or no letter above the bar indicate no significant difference (P > 0.05)3. 讨论
鱼体的生长状况是水产养殖中备受关注的重要经济性状,而GH与鱼类的生长密切相关,在鱼类的个体生长及生长调节中起关键性作用。马细兰等[2]指出垂体GH基因在转录水平上反映GH的合成速度,是反映动物生长速度的有效指标,生长快速的鱼其垂体GH mRNA表达量高;并且应用半定量RT-PCR法分析得出尼罗罗非鱼(Oreochromis niloticus)雄鱼垂体GH mRNA表达水平明显高于雌鱼,认为这可能是尼罗罗非鱼雄鱼生长快于雌鱼的主要内在原因之一。性成熟的金钱鱼(Scatophagus argus)雌鱼生长快于雄鱼,其生长差异与垂体GH mRNA的水平也呈显著的正相关[3]。同样,生长快速的雌性黄鲈(Perca flavescens)垂体GH mRNA水平显著高于雄鱼,并且在生长快速的春季显著高于秋季[4]。性成熟的鲤(Cyprinus carpio)垂体GH mRNA表达也呈现出季节性的周期变化,生长较快的季节(水温18~20 ℃)垂体GH mRNA表达比生长相对较慢的季节(水温8~10 ℃)高3倍[6]。鲇形目鳠科的大鳍鳠(Mystus macropterus)脑垂体GH的含量分别在3月和8月出现峰值,与该鱼春季和秋季的快速生长一致[7]。该目鲇科的野生鲇(Silurus asotus)垂体GH的含量在3月和7月各出现一个高峰值,研究者认为这可能与野生鲇春季和产卵后生长加快有关;并且雌性鲇鱼垂体GH的周年含量显著高于雄鱼,与雌鱼生长速度显著快于雄鱼的现象相符合[5]。SHIN等[8]研究发现光的波长可影响观赏性鱼类——克氏双带锯齿盖鱼(Amphiprion clarkii)的生长速度和垂体GH mRNA水平、生长速率与垂体GH mRNA的水平呈正相关。李云等[9]研究显示饲料中添加一定量的半胱胺盐酸盐(1 mg · g-1饲料)可显著提高斜带石斑鱼(Epinephelus coioides)的体质量增长率和垂体GH的转录水平。马细兰等[10]发现促黄体素释放激素类似物(LHRH-A)可显著上调尼罗罗非鱼生长轴相关基因(GH、GHR和IGF)的表达,从而促进鱼类的生长。上述研究说明鱼体的生长速度与垂体GH基因的转录水平呈正相关。
由于脑垂体合成GH后经过血液运送至靶器官,再通过GHR和IGF的介导进而达到促进鱼体生长的作用,因此鱼类的生长速度并不能完全通过GH水平来体现,还与GHR和IGF的水平有关。如银大麻哈鱼(Oncorhynchus kisutch)在禁食3周后,比正常投喂组肝脏GHR mRNA水平低,提示长时间禁食可引起肝脏GHR基因表达的下降[19]。SAERA-VILA等[20]报道金鲷(Sparus aurata)肝脏和脂肪组织GHRⅠ基因表达最丰富,但禁食后可引起GHRⅠmRNA水平的下降。斑点叉尾(Ictalurus punctatus)在投喂外源皮质醇或者禁食4周后,与对照组相比,肝脏GHR mRNA的丰度以及血清中IGF的浓度均显著下降,且肝脏GHR mRNA的丰度和血清中IGF的浓度呈正相关;投喂外源皮质醇后鱼的生长显著下降,禁食组鱼的体质量减少[21]。华益民和林浩然[22]报道鲤饥饿32 d导致生长受阻,肝组织IGF-ⅠmRNA表达水平下降;再投喂后,其生长和肝组织IGF-ⅠmRNA丰度都逐渐恢复,表现出鲤的生长状况与肝组织IGF-ⅠmRNA水平的同步变化。尼罗罗非鱼饥饿28 d内血清GH含量反倒显著升高,而血清IGF含量和肝胰脏IGF mRNA表达丰度显著下降,饥饿后再投喂,第14天时血清GH含量显著升高,继续投喂7 d血清GH含量又表现出显著下降,但仍与饥饿第7天时的GH水平相当,而血清IGF含量和肝胰脏IGF mRNA表达丰度在第21天恢复投喂过程中显著升高,并恢复到饥饿前水平[23]。鲮(Cirrhinus molitorella)在投喂不同蛋白水平的饲料时,在8周的养殖期内其特定生长率与肝组织IGF mRNA的表达量呈极显著的正相关[24]。但强俊等[25]在分析温度和饲料蛋白水平对吉富品系尼罗罗非鱼幼鱼生长的影响时则发现血清GH水平与鱼的特定生长率相关性较低。黄国强等[26]研究也发现褐牙鲆(Paralichthys olivaceus)幼鱼在低温胁迫10 d导致的生长减缓能在恢复至最适生长温度的30 d内获得完全补偿生长,但血浆GH和IGF水平与生长率不存在明显的相关性。
该研究通过分析不同养殖密度下革胡子鲶幼鱼的生长以及垂体GH基因的表达,探讨养殖密度对二者的影响以及二者的相关性。该研究的数据统计分析结果显示,0~30 d养殖期内,35 kg · m-3和65 kg · m-3密度组间的日增质量以及95 kg · m-3和125 kg · m-3密度组间的日增质量差异均不显著,但前2个密度组的日增质量显著高于后2个密度组(图 1)。但养殖第30天时各密度组间GH基因mRNA的相对表达量并未受养殖密度的影响,即各密度组间的GH基因的相对表达量差异不显著(图 4)。30~60 d养殖期内,35 kg · m-3和65 kg · m-3密度组间的日增质量与95 kg · m-3密度组差异不显著,但显著高于125 kg · m-3密度组(图 1)。而养殖第60天时,GH mRNA表达量的分析结果显示35 kg · m-3和65 kg · m-3密度组显著高于95 kg · m-3和125 kg · m-3密度组(图 4)。上述结果表明在该研究中革胡子鲶幼鱼的日增质量与垂体GH基因mRNA相对表达量不存在紧密的相关性。结合前人的研究结果,在该研究中笔者推测,仅就生长轴GH-GHR-IGF而言,革胡子鲶在较短的养殖时间内(30 d),养殖密度增大时,由于空间不足、摄食不均等原因可能影响鱼体生存状态和营养水平,而导致GH下游的生长调控因子,如GHR和IGF的合成与分泌减少,最终使鱼体的日增质量降低。但是随着养殖时间的延长(60 d),养殖密度增大到一定程度时则会影响处于生长轴中心位置的因子——GH mRNA表达的下降,进而GH的合成与分泌减少,这可能成为影响鱼体生长速度的主要内在因素。对于GH-GHR-IGF这一生长轴在革胡子鲶生长中的调节作用以及三者的相互补偿关系仍需进一步深入探究。
-
![]()
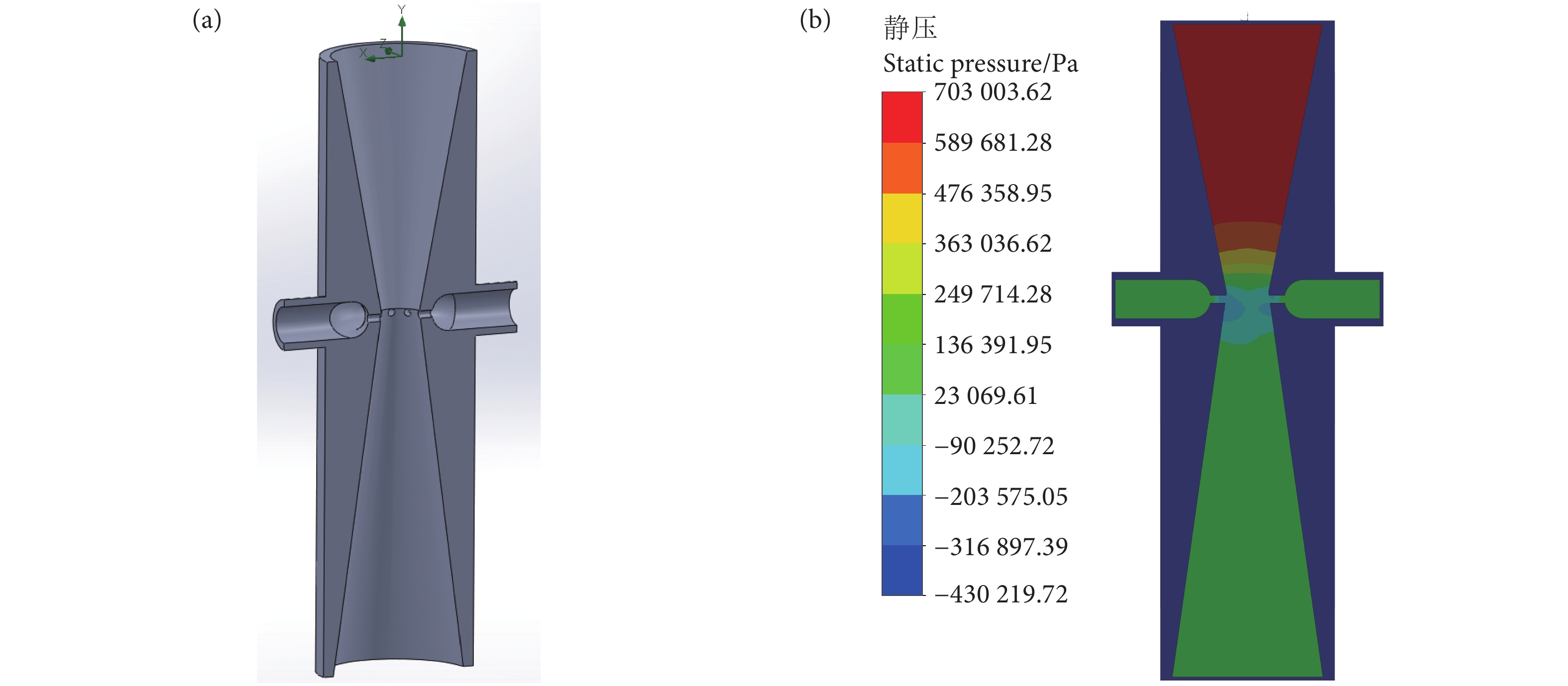
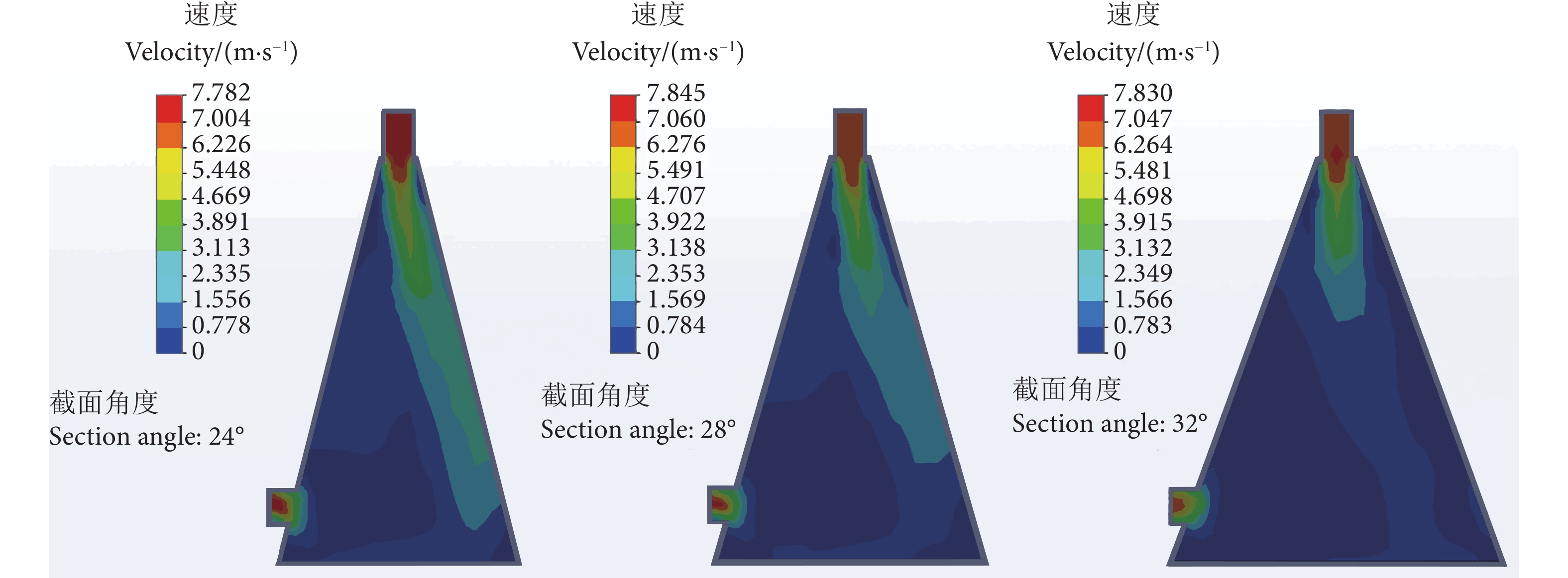
图 1 24°、28°、32° 截面角度的流体仿真分析图
Figure 1. Analysis diagram of fluid simulation at 24°, 28° and 32° section angles
![]()
图 2 增氧锥发生的偏流、回流现象和实验模拟过程中发生的偏流
Figure 2. Partial flow and reflux in speece cone and biased flow occurred during experimental simulation
![]()
图 3 微孔曝气射流器截面图和流体仿真图
Figure 3. Cross-sectional view and fluid simulation diagram of microporous aeration jet
![]()
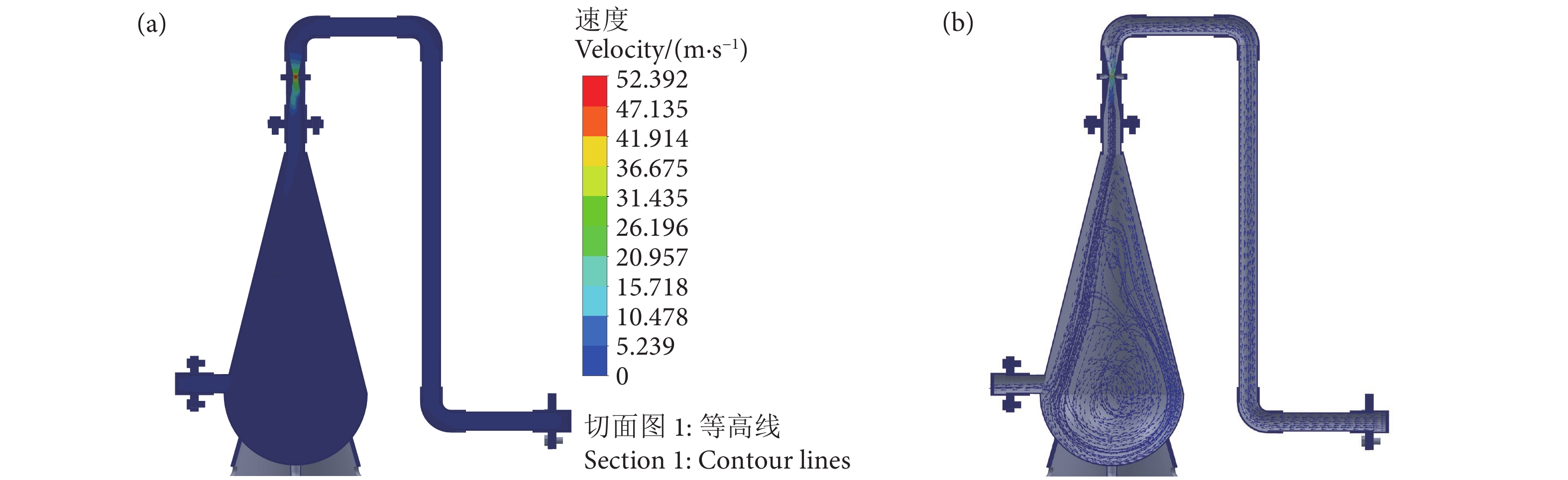
图 4 增氧锥整体速度分布图和内液体流动轨迹图
Figure 4. Overall velocity distribution plot and diagram of liquid flow trajectories in speece cone
![]()
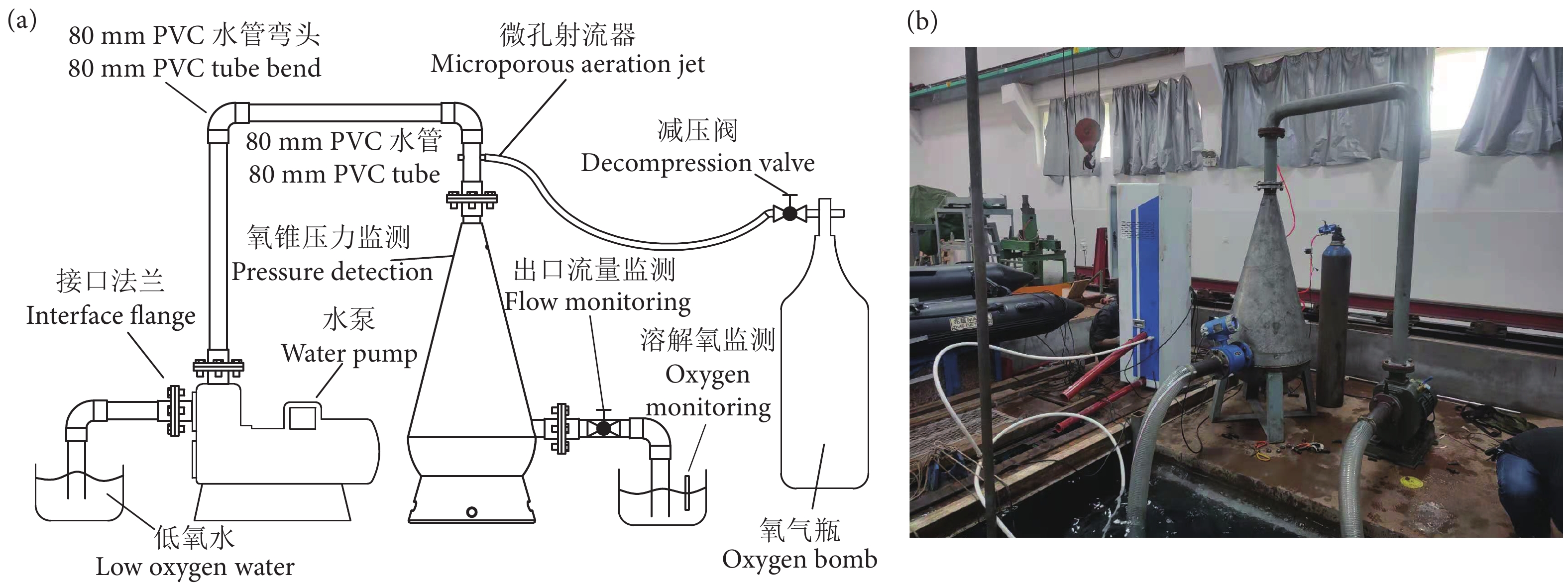
图 5 增氧锥增氧系统整体流程图和现场试验图
Figure 5. Overall flow chart and physical diagram of speece cone aeration system
![]()
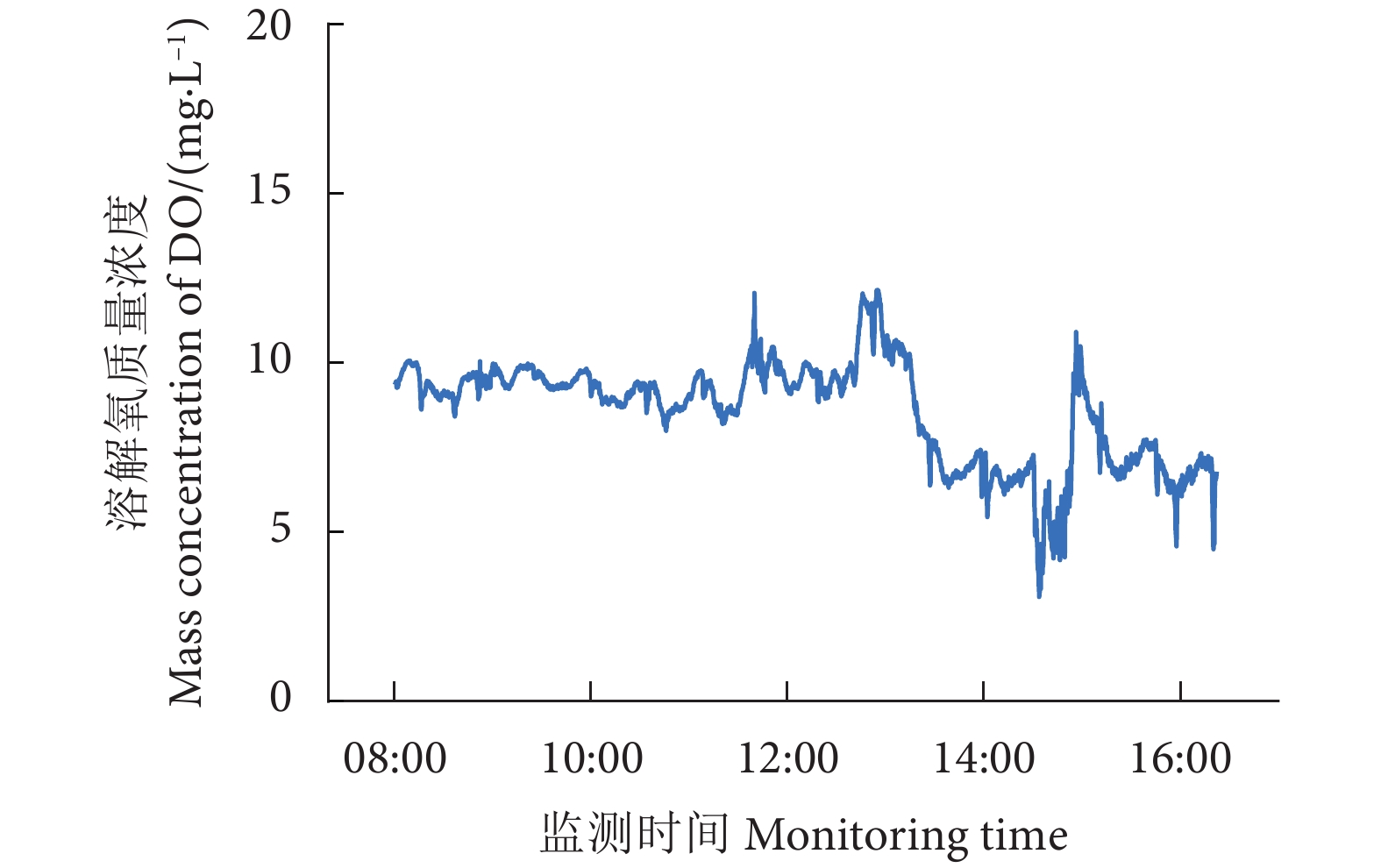
图 6 某养殖工船养殖舱内溶解氧时段变化
Figure 6. Variation in dissolved oxygen in cabin of an aquaculture ship
![]()
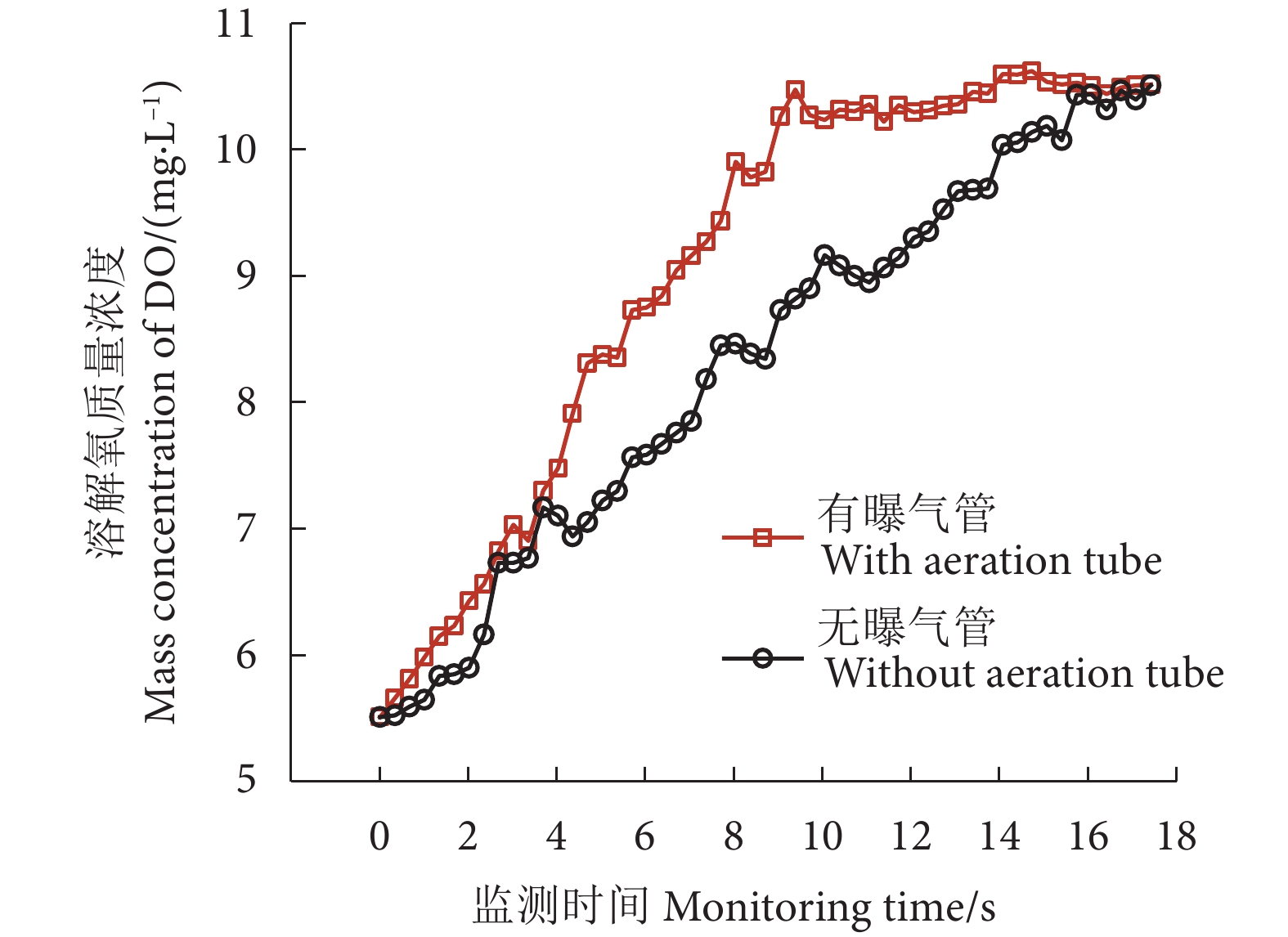
图 7 增氧锥内溶解氧提升速度对比
Figure 7. Comparison of dissolved oxygen enhancement rates in speece cone
表 1 管径、速度、流量对照表
Table 1 Check list of caliber, velocity and rate of flow
管径 (DN)
Caliber/mm流量 Flow rate/(m3·h−1) 1.2 m·s−1 1.4 m·s−1 1.6 m·s−1 1.8 m·s−1 2.0 m·s−1 2.2 m·s−1 2.4 m·s−1 2.6 m·s−1 2.8 m·s−1 3.0 m·s−1 40 5.4 6.3 7.2 8.1 9.0 10.0 10.9 11.8 12.7 13.6 50 8.5 9.9 11.3 12.7 12.7 15.6 17.0 18.4 19.8 21.2 65 14.3 16.7 19.1 21.5 21.5 26.3 28.7 31.1 33.4 35.8 80 21.7 25.3 29.0 32.6 32.6 39.8 43.4 47.0 50.7 54.3 100 33.9 39.6 45.2 50.9 56.5 62.2 67.9 73.5 79.2 84.8 125 53.0 61.9 70.7 79.5 88.4 97.2 106.0 114.9 123.7 132.5 150 76.3 89.1 101.8 114.5 127.2 140.0 152.7 165.1 178.1 190.9  下载: 导出CSV
下载: 导出CSV
-
[1] 张宝龙, 赵子续, 曲木, 等. 工厂化水产养殖现状分析[J]. 养殖与饲料, 2020(1): 31-34. [2] 周小燕, 倪琦, 徐皓, 等. 2021年中国水产养殖全程机械化发展报告[J]. 中国农机化学报, 2022, 43(12): 1-4. [3] 赵文涓. 水产生态养殖与新养殖模式发展战略探析[J]. 畜牧兽医科技信息, 2022(4): 203-205. [4] 王海姮, 侯昊晨, 刘鹰. 循环水养殖系统的研究进展及发展趋势[J]. 水产科学, 2023, 42(4): 735-741. [5] 农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2023中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2023: II-III. [6] 王祖峰, 刘晓军, 赵文武. 传统水产养殖场 (区) 生态化、景观化、休闲化改造的初步思考[J]. 淡水渔业, 2020, 50(4): 108-112. [7] 徐皓, 陈家勇, 方辉, 等. 中国海洋渔业转型与深蓝渔业战略性新兴产业[J]. 渔业现代化, 2020, 47(3): 1-9. [8] 何皛磊, 张海文. “深海渔场”的应用前景[J]. 船舶, 2018(2): 1-6. [9] 徐杰, 韩立民, 张莹. 我国深远海养殖的产业特征及其政策支持[J]. 中国渔业经济, 2021, 39(1): 98-107 [10] 徐琰斐, 徐皓, 刘晃. 中国深远海养殖发展方式研究[J]. 渔业现代化, 2021, 48(1): 9-15. [11] 闫国琦, 倪小辉, 莫嘉嗣. 深远海养殖装备技术研究现状与发展趋势[J]. 大连海洋大学学报, 2018, 33(1): 123-129. [12] 纪毓昭, 王志勇. 我国深远海养殖装备发展现状及趋势分析[J]. 船舶工程, 2020, 42(S2): 1-4, 82. [13] 胡方珍, 盛伟群, 王体涛. 深远海养殖装备技术现状及标准化工作建议[J]. 船舶标准化工程师, 2021, 54(5): 6-12. [14] 黄小华, 庞国良, 袁太平, 等. 我国深远海网箱养殖工程与装备技术研究综述[J]. 渔业科学进展, 2022, 43(6): 121-131. [15] 张琳桓, 张青亮, 孟广玮. 基于可移动式养殖工船的新型深远海养殖产业链分析[J]. 船舶工程, 2020(增2): 40-44. [16] 崔铭超, 金娇辉, 黄温赟. 养殖工船系统构建与总体技术探讨[J]. 渔业现代化, 2019, 46(2): 61-66. [17] ASHLEY K, MAVINIC D, HALL K. Oxygenation performance of a laboratory-scale speece cone hypolimnetic aerator: preliminary assessment[J]. Can J Civil Engin, 2008, 35(7): 663-675. doi: 10.1139/L08-011
[18] 庄礼贤. 流体力学[M]. 合肥: 中国科学技术大学出版社, 1997: 105-175. [19] 房燕, 曹广斌, 韩世成, 等. 基于Fluent的工厂化水产养殖增氧锥的数值模拟及结构优化设计[J]. 江苏农业科学, 2013, 41(4): 355-358. [20] ASHLEY K, FATTAH K, MAVINIC D, et al. Analysis of design factors influencing the oxygen transfer of a pilot-scale speece cone hypolimnetic aerator[J]. J Environ Engin, 2014, 140(3): 04013011. doi: 10.1061/(ASCE)EE.1943-7870.0000789
[21] 陈有光, 段登选, 陈秀丽, 等. 工厂化养鱼中氧气锥的增氧规律[J]. 渔业现代化, 2009, 36(3): 26-30. [22] 农宏亮, 曾伯胜, 莫建霖, 等. 基于SolidWorks Flow Simulation的甘蔗收割机排杂装置内部流场模拟[J]. 农业工程, 2017, 7(4): 133-137, 177. [23] 阎昌琪. 气液两相流 [M]. 哈尔滨: 哈尔滨工程大学出版社, 2010: 2-5. [24] 颜攀. 固定床鼓泡反应器中微气泡的形成演化规律 [D]杭州: 浙江大学, 2017: 51-52. [25] AGO K, NAGASAWA K, TAKITA J, et al. Development of an aerobic cultivation system by using a microbubble aeration technology[J]. J Chem Engin Jap, 2005, 38(9): 757-762. doi: 10.1252/jcej.38.757
[26] 王淼, 黄兴法, 李光永. 文丘里施肥器性能数值模拟研究[J]. 农业工程学报, 2006, 22(7): 27-31. [27] LI G, YANG X G, DAI G. CFD simulation of effects of the configuration of gas distributors on gas-liquid flow and mixing in a bubble column[J]. Chem Engin Sci, 2009, 64(24): 5104-5116. doi: 10.1016/j.ces.2009.08.016
[28] 蒋建明, 朱正伟, 李正明, 等. 水产养殖中复合精确自动增氧技术研究[J]. 农业机械学报, 2017, 48(12): 334-339. [29] 周红标. 基于自组织模糊神经网络的污水处理过程溶解氧控制[J]. 化工学报, 2017, 68(4): 1516-1524. [30] 史兵. 河蟹池塘养殖智能支持系统关键技术研究[D]. 镇江: 江苏大学, 2013: 40-47. [31] 马相鹏, 高海波, 李程, 等. 基于鲸鱼算法的养殖工船水泵模糊控制优化研究[J]. 渔业现代化, 2023, 50(3): 48-55. [32] 房燕. 工厂化水产养殖纯氧增氧锥优化设计与性能测试[D]. 上海: 上海海洋大学, 2013: 34-35. -
期刊类型引用(15)
1. 周志希,唐汇娟,柯志新,刘甲星,周伟华. 基于形态学和高通量测序的春季南澳海域浮游植物群落特征及其与环境因子关系. 热带海洋学报. 2025(01): 53-65 .  百度学术
百度学术
2. 柳原,柯志新,李开枝,谭烨辉,梁竣策,周伟华. 人类活动和沿岸流影响下的粤东近海浮游动物群落特征. 热带海洋学报. 2024(04): 98-111 . 百度学术
3. 薛宇,于浩林,周广军,赵伟,唐衍力. 渤海小竹山岛海洋牧场区营养盐分布特征及增养殖适宜性探究. 渔业科学进展. 2023(01): 1-12 . 百度学术
4. 黎素菊,洪捷娴,陈树鹏. 柘林湾养殖区氮、磷季节分布特征及富营养化评价. 江西水产科技. 2022(04): 45-47+51 . 百度学术
5. 匡翠萍,王丹,赵钒,刘会欣,朱磊. 金梦海湾及其邻近海域水环境因子时空分布特征. 同济大学学报(自然科学版). 2021(06): 880-890 . 百度学术
6. 任玉正,柯志新,谭烨辉,李开枝. 广东省南澳岛东部海域浮游动物群落结构及其影响因素. 热带海洋学报. 2020(02): 65-76 . 百度学术
7. 杨文超,黄道建,陈继鑫,陈晓燕,王宇珊. 大亚湾海域2009—2015年氮、磷营养盐时空分布及富营养化评价. 南方水产科学. 2020(02): 54-61 . 本站查看
8. 陈丹婷,柯志新,谭烨辉,刘甲星. 汕头南澳—东山海域营养盐季节分布特征及其对浮游植物生长的潜在性限制. 生态科学. 2020(04): 41-50 . 百度学术
9. 马欢,秦传新,陈丕茂,林会洁,段丁毓. 柘林湾海洋牧场生态系统服务价值评估. 南方水产科学. 2019(01): 10-19 . 本站查看
10. 王言丰,胡启伟,余景,陈丕茂,舒黎明. 粤东柘林湾海洋牧场渔业资源增殖效果评估. 南方水产科学. 2019(02): 12-19 . 本站查看
11. 徐淑敏,齐占会,史荣君,刘永,韩婷婷,黄洪辉. 水产养殖对亚热带海湾氮磷营养盐时空分布的影响——以深澳湾为例. 南方水产科学. 2019(04): 29-38 . 本站查看
12. 王文杰,陈丕茂,袁华荣,冯雪,张露,龙鑫玲,陈文静,李丹丹. 粤东柘林湾甲壳类群落结构季节变化分析. 南方水产科学. 2018(03): 29-39 . 本站查看
13. 佟飞,秦传新,余景,陈丕茂. 粤东柘林湾溜牛人工鱼礁建设选址生态基础评价. 南方水产科学. 2016(06): 25-32 . 本站查看
14. 舒黎明,陈丕茂,秦传新,黎小国,周艳波,冯雪,于杰,袁华荣,李国迎. 柘林湾-南澳岛潮间带冬夏两季大型底栖动物种类组成及优势种. 生态学杂志. 2016(02): 423-430 . 百度学术
15. 舒黎明,陈丕茂,黎小国,秦传新,于杰,周艳波,袁华荣. 柘林湾附近海域大型底栖动物物种多样性. 中国水产科学. 2015(03): 501-516 . 百度学术
其他类型引用(5)





计量
- 文章访问数: 207
- HTML全文浏览量: 72
- PDF下载量: 56
- 被引次数: 20
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
