
Influence of mariculure on tempo-spatial distribution of nitrogen and phosphorus in subtropical zone: a case study of Shen'ao Bay
-
摘要:
文章对典型的亚热带养殖海湾——深澳湾海水中无机氮(DIN)、磷酸盐(PO4-P)浓度的时空变化特征进行了分析,研究了鱼类网箱和贝藻筏式等规模化养殖活动对营养盐时空分布特征的影响,并对营养盐的潜在限制性进行了探讨。结果显示,深澳湾DIN和PO4-P浓度及分布呈明显的季节变化:DIN在秋季最高,夏季最低;PO4-P在冬季最高,夏季最低。春季网箱区的DIN浓度和氮磷比(N/P)低于贝藻养殖区和对照区,而其他3个季节,网箱区的DIN和PO4-P浓度以及N/P均高于贝藻养殖区和对照区。贝藻养殖区和对照区之间在各个季节,氮、磷营养盐和N/P之间均无显著差异。各个季节DIN和PO4-P浓度均高于理论上浮游植物生长的营养盐阈值,不存在营养盐的绝对限制。夏、冬季的N/P分别为13.6、13.1,低于Redfield值,说明存在N的潜在限制;春、秋季的N/P分别为16.6、19.0,说明P的潜在限制性较强。深澳湾的年均N/P为14.3,全湾受N潜在限制性较强。除夏季外,硝酸盐 (NO3-N)是DIN的主要组成,比例介于51.7%~92.7%,其次为NH4-N (5.2%~43.8%),亚硝酸盐 (NO2-N)比例最低(2.1%~27.2%),说明深澳湾的氮营养盐达到了热力学平衡状态。与2001年相比,深澳湾海区的DIN和PO4-P浓度均有下降,由中度营养型转变成贫营养型,年平均N/P更接近Redfield值,说明深澳湾的生产力水平依然受氮限制,营养盐的时空分布特征一定程度上体现了规模化贝藻养殖的影响。
Abstract:We investigated the tempo-spatial variation of disovled inorganic nitrogen (DIN) and phosphate (PO4-P) in Shen'ao Bay that is a typical subtropical aquaculture bay. Besides, we studied the effects of large-scale aquaculture activities such as fish cage and shellfish raft on the tempo-spatial distribution of nutrients, and evaluated the potential limiting factors of the nutrients. The concentration and distribution of DIN and PO4-P showed obvious seasonal variation (DIN and PO4-P reached their maximum values in autumn and winter, respectively, and both reached their minimum values in summer). The DIN nutrients concentrations and N/P ratios in fish cage area in spring were lower than those in oyster-macroalgae culture area and the control area. However, in the other three seasons, the DIN and PO4-P concentrations and N/P ratios in fish cage area were higher than those in the other areas. There was no significant difference in the nitrogen and phosphorus concentrations as well as N/P ratios between oyster-macroalgae culture area and the control area in each season. Both DIN and PO4-P were higher than the theoretical threshold for phytoplankton growth, indicating that there was no absolute nutrient limitation. The N/P ratios in summer and winter were 13.6 and 13.1, respectively, which were lower than the Redfield ratio, indicating that there was potential N limitation during these seasons. The N/P ratios in spring and autumn were 16.6 and 19.0, respectively, indicating potential P limitation. The average seasonal ratio was 14.3, indicating that N was probably the most limiting nutrient in Shen'ao Bay. DIN was dominated by nitrate nitrogen (NO3-N) except for summer with percentages of 51.7%−92.7%, followed by ammonia nitrogen (NH4-N) with percentages of 5.2%−43.8%, and nitrite nitrogen (NO2-N) was lowest with percentages of 2.1%−27.2%, indicating that the thermodynamic equilibrium of N was reached. Compared with 2001, the concentrations of both DIN and PO4-P decreased, with the trophic level changing into poor from moderate. It is shown that the productivity level of Shen'ao Bay is still controlled by nitrogen, and the tempo-spatial distribution of nutrients reflects the impact of large-scale shellfish culture to a certain extent.
-
Keywords:
- Shen'ao Bay /
- nitrogen /
- phosphate /
- nutrient /
- tempo-spatial variation /
- nutrient limitation
-
ABC转运子(ATP-binding cassette transporter)广泛存在于真核和原核生物中,是细胞内物质运输系统的重要组成部分,主要功能为利用ATP水解产生的能量,对底物进行逆浓度梯度的跨膜运输,转运各种细菌生长所需的营养物质,如糖类、多肽、脂质、无机离子或抗生素等,并参与许多重要生理生化过程,如摄入营养、信号转导及抗原呈递等[1-6]。
核糖转运系统对于核糖的高亲和吸收至关重要,大肠杆菌的核糖转运系统由rbs操纵子(rbsA,-B,-C,-D,-K,-R)编码的6种蛋白质组成[7]。rbs操纵子转运核糖的途径如下,首先核糖通过外膜孔蛋白扩散到革兰氏阴性菌的周质中,游离核糖被RbsB结合,随后将核糖传递到由RbsAC形成的内膜复合物中,RbsAC再以ATP依赖的方式将底物跨膜运输至细胞质[8-10],核糖被激酶RbsK磷酸化后螯合在细胞中[11]。RbsR是结合到rbs操纵子转录起始位点的阻遏物,RbsD位于RbsK上游,可催化核糖改变构型,由吡喃构象转化为呋喃构象,从而发挥其功能[12]。RbsB主要功能为结合核糖,并将其转运至细胞中。除与核糖结合相关的功能外,近年来有研究发现RbsB蛋白还可介导细菌种间通讯及参与细菌生长调控。在伴放线聚集杆菌(Aggregatibacter actinomycetemcomitans)中,RbsB被视作AI-2信号分子的受体,参与对密度感应系统(QS系统)的调控,胸膜肺炎放线杆菌的RbsB2还可结合哈维弧菌BB170培养物上清液中的AI-2信号分子。此外,rbsB基因与生物被膜的形成和在铁限制条件下生长的能力也有一定的关系[13-16]。
近年来,哈维弧菌(Vibrio harveyi)是中国南方沿海地区水产养殖业中最主要的病原菌之一,感染对象广泛,包括海洋鱼类[17-19]、甲壳类[20]、软体动物[21]和珊瑚[22]等,给沿海养殖户带来重大经济损失。为探究RbsB在哈维弧菌中的功能,本研究以课题组前期测序得到的哈维弧菌rbsB片段序列为基础,克隆哈维弧菌rbsB基因全长,并对其氨基酸序列进行预测分析结构功能,多序列比对比较与其他弧菌RbsB氨基酸差异,构建原核表达载体并对其进行诱导表达获得重组蛋白,为进一步研究其致病性及疫苗制备奠定基础。
1. 材料与方法
1.1 菌株来源与培养
哈维弧菌354菌株为中国水产科学研究院南海水产研究所渔业生物病害防治研究室于2013年10月从深圳南澳患病珍珠龙趸 [鞍带石斑鱼(Epinephelus lanceolatus)♂×棕点石斑鱼(E.fuscoguttatus)♀] 体内分离保种[23]。接种于LBS液体培养基(酵母提取粉0.5%,胰蛋白胨1%,NaCl 3%),28 ℃、140 r·min–1振荡培养16 h。
1.2 rbsB基因的同源扩增
根据前期对哈维弧菌354菌株进行全基因组测序所得的rbsB基因序列,设计1对引物,引物两端分别加限制性酶BamH I和Xho I酶切位点(由上海英潍捷基合成),正向引物:5′ GATCTGGTTCCGCGTGGATCCATGAAAAAACTCGCAACTCTTATC 3′,反向引物:5′ GTCACGATGCGGCCGCTCGAGCTTAGTCACGACTTTAAGCGGAA 3′。PCR反应为50 μL体系,反应条件为95 ℃预变性5 min;95 ℃变性30 s,55 ℃退火40 s,72 ℃延伸1 min,共35个循环;72 ℃延伸10 min,4 ℃保存。扩增产物经1.2%琼脂糖凝胶电泳检测后,切胶回收并连接转化,挑取阳性菌送上海英潍捷基生物技术有限公司测序。
1.3 rbsB基因序列分析
用软件ApE分析哈维弧菌rbsB基因序列及其开放阅读框,并将翻译后的氨基酸序列应用NCBI中的BLAST程序(https://blast.ncbi.nlm.nih.gov/)进行同源性检索;应用SMART及ExPaSy-Prosite对RbsB蛋白进行功能位点和功能域的预测;在线工具SignalP 4.0 Server及TMHMM 2.0预测蛋白的信号肽和跨膜结构域;在线(http://www.cbs.dtu.dk/services/NetPhos/)对RbsB氨基酸序列进行磷酸化位点分析;应用ExPASy的ProtParam程序对RbsB氨基酸序列进行氨基酸数量、分子量、等电点等基本理化性质分析;ExPaSy-SOPMA分析蛋白质的二级结构,对蛋白质三级结构采用SWISS-MODEL进行预测。用Clustal W 2.0进行多重序列比对,用MAGA 7.0构建NJ系统进化树。
1.4 重组质粒的构建
按照质粒DNA提取试剂盒说明提取rbsB-DH5α和pGEX-4t-1两种质粒,构建pGEX-rbsB重组质粒,随后将重组质粒转化入大肠杆菌BL21(DE3)感受态中,阳性菌以BamH I、Xho I双酶切后进行PCR反应,经1.2%琼脂糖凝胶电泳检测后,送上海英潍捷基生物技术有限公司测序以验证插入序列的正确性。
1.5 重组蛋白表达条件优化
将测序验证结果正确的阳性菌菌液按1∶100的比例接种到含100 μg·mL–1Amp的LB液体培养基中,37 ℃ 、180 r·min–1振荡培养至OD600为0.4~0.6时,加入异丙基硫代半乳糖甘(IPTG)诱导表达,IPTG的浓度分别为0、0.2 mmol·L–1、0.4 mmol·L–1、0.6 mmol·L–1、0.8 mmol·L–1和1 mmol·L–1,在30 ℃和37 ℃ 2个温度下180 r·min–1继续诱导培养8 h,分别收集诱导(0 h,2 h,4 h,6 h,8 h)菌体2 mL,菌液4 ℃离心5 min后弃上清,加入50 μL 5×SDS-PAGE Loading buffer和200 μL 1×PBS悬浮菌体,100 ℃煮沸10 min,4 ℃离心3 min后进行SDS-PAGE电泳,考马斯亮蓝R-250过夜染色后进行分析。同时进行Western blot,以Anti GST-Tag Rabbit HRP为抗体,最后用HRP-DAB (辣根过氧化物酶)底物显色试剂盒进行显色处理。
1.6 铁离子对rbs基因表达的影响
将哈维弧菌354接种于T1N1 (胰蛋白胨1%,酵母提取粉0.5%,NaCl 1%)培养基中140 r·min–1振荡过夜培养,按1∶1 000的比例重新接种于T1N1培养基中,实验组额外加入终浓度为7 mmol·L–1的柠檬酸铁,以加入菌液后的时间记为第0小时,分别培养4 h、8 h、12 h、24 h、48 h后按照RNA提取试剂盒说明提取RNA,根据逆转录试剂盒的说明合成cDNA用于qRT-PCR。采用qRT-PCR检测基因表达,设计1对qRT-PCR引物(正向引物:5′ GTGCAGAAGCGAAAGCGAAA 3′,反向引物:5′ GCACCACGGTCTAGTGTCAA 3′),以哈维弧菌16S rDNA作为内参基因(正向引物:5′ CCTACGGGAGGCAGCAG 3′,反向引物:5′ ATTACCGCGGCTGCTGG 3′)。反应体系为10 μL,反应条件为94 ℃预变性1 min;94 ℃变性10 s,60 ℃退火延伸40 s,共40个循环。通过熔解曲线和琼脂糖凝胶电泳共同验证PCR产物的特异性。同时扩增目的基因rbsB与内参基因16S rDNA,以各时间段的16S rDNA的表达量做△Ct校正样,以对照组培养第4小时的Ct值为△△Ct校正样,第0小时重新接种后的培养液数据作0处理,设为坐标原点,使用2–△△Ct的方法[24]计算rbsB基因不同时间段的相对拷贝数,作图分析。
2. 结果
2.1 rbsB全长核苷酸序列及其推导的氨基酸序列分析
克隆得到rbsB基因(GenBank序列号MF797015)测序后经DNAstar分析,得到开放阅读框(ORF)大小为879 bp (图1),G+C含量为49%。该序列可编码292个氨基酸,分子量为30.7 kD,理论等电点为5.05,在1~23 aa处有信号区域,N-末端为Met。
![]() 图 1 哈维弧菌rbsB基因序列及氨基酸序列起始密码子(ATG)和终止密码子(TAA)大写标明,磷酸位点加粗并用下划线标出;以“<”和“>”标出Peripla_BP_4结构域的起始和终止位置Figure 1. Gene and amino acid sequence of rbsB gene from V.harveyiThe start and stop codons are capitalized; phosphate sites are bold and underlined; the start and end positions of the Peripla_BP_4 domain are identified by "<" and ">".
图 1 哈维弧菌rbsB基因序列及氨基酸序列起始密码子(ATG)和终止密码子(TAA)大写标明,磷酸位点加粗并用下划线标出;以“<”和“>”标出Peripla_BP_4结构域的起始和终止位置Figure 1. Gene and amino acid sequence of rbsB gene from V.harveyiThe start and stop codons are capitalized; phosphate sites are bold and underlined; the start and end positions of the Peripla_BP_4 domain are identified by "<" and ">".2.2 RbsB蛋白理化性质分析
RbsB原子总数为4 362个,分子式为C1338H2215N371O428S10,不稳定系数为19.36,属于稳定蛋白。脂溶系数为102.26,亲水性系数为0.043。分析氨基酸组成可知RbsB蛋白含酸性氨基酸38个,碱性氨基酸30个,极性氨基酸224个。
2.3 RbsB结构分析和功能预测
TargetP 1.1 Server预测结果表明RbsB蛋白无线粒体目标肽,但有一分泌途径信号肽。采用Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)中的Gram-negative protein程序进行亚细胞定位,推测RbsB蛋白最可能定位于细胞质中,为周质蛋白。
在Swiss-Prot和TrEMB的模式序列进行搜索没有得到与已知序列匹配的功能位点。InterProscan结合Smart分析RbsB蛋白的结构功能域,结果表明RbsB蛋白中27~278 aa存在Peripla_BP_4结构域(图1)。采用TMHMM 2.0 Server分析RbsB无跨膜区域,为非跨膜蛋白。
RbsB蛋白共含有23个蛋白质磷酸化位点,包括9个苏氨酸位点、12个丝氨酸位点、2个酪氨酸位点(图1)。RbsB蛋白的二级结构主要由4种形式组成,其中α-螺旋占43.84%,无规则卷曲占22.60%,延伸链占21.58%,β-转角占11.99%,三维结构见图2。由此推测,RbsB蛋白主要在运载结合中发挥作用。
2.4 RbsB多重序列比对和系统进化树构建
将获得的哈维弧菌RbsB氨基酸序列与其他弧菌RbsB氨基酸序列(表1)应用Clustal W 2.0进行多重序列比对(图3)。根据比对情况利用MEGA 7.0构建NJ系统进化树(图4),结果显示哈维弧菌和欧文弧菌(V.owensii CAIM 1854)关系最近,与远青弧菌(V.azureus NBRC 104587)关系较远。
表 1 哈维弧菌354菌株的RbsB与其他细菌的氨基酸同源性分析Table 1. Homology analysis of amino acids of RbsB from V.harveyi strain 354 and other bacterial species菌种
species同源性/%
homology注册号
accession No.哈维弧菌 (V.harveyi ATCC 33843) 100 AIV07307.1 欧文弧菌 (V. owensii CAIM 1854) 100 KIF46393.1 坎氏弧菌 (V.campbellii ATCC BAA-1116) 98 AGU97393.1 轮虫弧菌 (V.rotiferianus HM-10) 98 OHY94262.1 溶藻弧菌 (V.alginolyticus ATCC 17749) 97 GAD69766.1 副溶血弧菌 (V.parahaemolyticus ATCC 17802) 97 KKI09273.1 需钠弧菌 (V.natriegens ATCC 14048) 97 EPM40141.1 贻贝弧菌 (V.mytili CAIM 528) 96 KIN10841.1 创伤弧菌 (V.vulnificus ATCC 33147) 92 KGK71697.1 远青弧菌 (V.azureus NBRC 104587) 92 GAD74752.1 ![]() 图 3 哈维弧菌RbsB氨基酸序列与其他已知菌种的比对分析“*”表示所有序列中都一致的氨基酸;“:”表示保守的氨基酸;“.”表示半保守的氨基酸,方框内为Peripla_BP_4结构域Figure 3. Sequence alignment of predicted RbsB amino acid from V.harveyi and other known bacterial species“*” means that the residues or nucleotides are identical in all sequences in the alignment; “:” indicates conserved substitution; “.” indicates semi-conserved substitution; Peripla_BP_4 domain is in box.
图 3 哈维弧菌RbsB氨基酸序列与其他已知菌种的比对分析“*”表示所有序列中都一致的氨基酸;“:”表示保守的氨基酸;“.”表示半保守的氨基酸,方框内为Peripla_BP_4结构域Figure 3. Sequence alignment of predicted RbsB amino acid from V.harveyi and other known bacterial species“*” means that the residues or nucleotides are identical in all sequences in the alignment; “:” indicates conserved substitution; “.” indicates semi-conserved substitution; Peripla_BP_4 domain is in box.![]() 图 4 利用Clustal W 2.0和MEGA 7.0构建的rbsB基因的氨基酸序列系统进化树Figure 4. Phylogenetic tree of amino sequence of rbsB gene by Clustal W 2.0 and MEGA 7.0
图 4 利用Clustal W 2.0和MEGA 7.0构建的rbsB基因的氨基酸序列系统进化树Figure 4. Phylogenetic tree of amino sequence of rbsB gene by Clustal W 2.0 and MEGA 7.02.5 重组表达载体的构建与鉴定
将PCR扩增产物和原核表达pGEX-4t-1质粒经XhoI、BamH I双酶切后构建重组质粒pGEX-4t-1-rbsB,并将其转化到大肠埃希菌BL21 (DE3)感受态中,经IPTG诱导后进行SDS-PAGE电泳,观察到诱导组在59 kD处有一条带。Western bolt结果表明重组蛋白诱导表达成功(图5)。37 ℃条件下诱导8 h表达量最高(图6),IPTG浓度对蛋白的表达量没有影响。对载体pGEX-4t-1-rbsB菌液测序后,与NCBI中V.harveyi CAIM 1792的rbsB基因序列进行比对,序列一致。
![]() 图 6 温度及时间对诱导RbsB蛋白的表达量的影响M. Marker;1. pGEX-4t-1菌液;2. 诱导0 h后菌液;3~5. 30 ℃诱导4 h、6 h、8 h后菌液;6~8. 37 ℃诱导4 h、6 h、8 h后菌液Figure 6. Effect of temperature and time on expression of RbsB proteinM. Marker; 1. pGEX-4t-1 plasmid; 2. 0 h after IPTG induction; 3–5. 4 h, 6 h and 8 h after IPTG induction at 30 ℃, respectively; 6–8. 4 h, 6 h and 8 h after IPTG induction at 37 ℃, respectively
图 6 温度及时间对诱导RbsB蛋白的表达量的影响M. Marker;1. pGEX-4t-1菌液;2. 诱导0 h后菌液;3~5. 30 ℃诱导4 h、6 h、8 h后菌液;6~8. 37 ℃诱导4 h、6 h、8 h后菌液Figure 6. Effect of temperature and time on expression of RbsB proteinM. Marker; 1. pGEX-4t-1 plasmid; 2. 0 h after IPTG induction; 3–5. 4 h, 6 h and 8 h after IPTG induction at 30 ℃, respectively; 6–8. 4 h, 6 h and 8 h after IPTG induction at 37 ℃, respectively2.6 铁离子对rbsB基因表达的影响
用qRT-PCR对rbsB在富铁培养条件下的表达量进行分析(图7),结果显示rbsB的表达具有明显的差异性。在限铁培养条件下,rbsB表达量在培养第4小时达到最高(6.22),随着培养时间的增加表达量逐步降低,第48小时为0.35;富铁培养条件下则相反,第4小时表达量仅为0.45,培养第48小时rbsB的表达量为5.48。
3. 讨论
在革兰氏阴性菌中,周质结合蛋白溶解在细胞周质中,底物进入周质后与周质结合蛋白可逆性结合,从而通过构象上的改变,进一步与膜结合蛋白相互作用,最终使底物进入胞浆发挥功能作用[25]。RbsB作为周质结合蛋白,被认为可能参与细菌种间通讯及细菌生长调控,在细菌毒力因子的表达中发挥重要作用。
本研究首次克隆出哈维弧菌rbsB基因全长,二级结构预测含有α-螺旋、无规则卷曲、延伸链、β-转角4种结构。SDS-PAGE和Western blot检测结果表明,Vh-RbsB蛋白与已报道的豌豆根瘤菌(Rhizobium leguminosarum)[16]、伴放线聚集杆菌[26]的RbsB蛋白分子量相似,为非分泌型蛋白,均含有N-端信号肽,亚细胞定位为周质蛋白。具核梭杆菌(Fusobacterium nucleatum)中另一种AI-2信号分子D-半乳糖结合蛋白Gbp也显示出与RbsB序列的相似性[27]。然而豌豆根瘤菌RbsB蛋白与哈维弧菌LuxP蛋白共享结构域,其N-端信号肽可被I型分泌系统所识别,研究比对哈维弧菌354株RbsB与LuxP的结构域后,并未发现如上特征,这可能是不同种属间RbsB氨基酸序列差异所导致的功能差异。
结合Pfam蛋白数据库和SMART分析发现Vh-RbsB蛋白包含Peripla_BP_4结构域,Peripla_BP_4结构域所属的第2周质结合蛋白家族簇中包含目前为止发现的2种具有AI-2结合能力的周质结合蛋白AI-2受体,分别是哈维弧菌的LuxP型受体和鼠伤寒沙门菌的LsrB型受体[28-29]。在非流行性流感嗜血杆菌(Nontypeable Haemophilus influenzae,NTHI)中,rbsB编码的蛋白对NTHI AI-2信号系统起着至关重要的作用,可促进体外生物膜的形成及慢性感染,rbsB的失活显著降低NTHI对AI-2和AI-2前体二羟基戊二酮(dihydroxypentanedione,DPD)的吸收,被视作AI-2的调节基因[30]。胸膜肺炎放线杆菌中RbsB与哈维弧菌的LuxP有27%相似性,能够结合哈维弧菌AI-2分子[15],James等[14]、Shao等[26]实验也表明在限铁培养条件下伴放线聚集杆菌中rbsB缺失株与luxS缺失株表型类似。以上结果显示RbsB蛋白在部分细菌中起到结合AI-2的功能。因此推测在哈维弧菌354株中,RbsB蛋白可能与密度感应系统有关,为一种潜在的AI-2受体,与LuxP协同或竞争性结合AI-2信号分子,参与信号结合及对密度感应系统的调控。
RbsB蛋白同AI-2的活性密切相关,在铁限制培养条件下rbsB基因表达量的变化同细菌的生长及AI-2的活性有明显的联系。本文通过qRT-PCR分析哈维弧菌354株在富铁与限铁条件下rbsB基因表达量的变化,发现限铁条件下rbsB表达量在对数生长早期就达到最高值,而后持续下降,在培养第24小时达到最低;富铁条件下rbsB表达量在对数生长后期(第12小时)达到最低,而后显著上升,在培养第48小时达到最高。James等[14]证实在限铁条件下,伴放线聚集杆菌rbsB缺失株生长水平仅为野生株的30%,rbsB的失活影响着铁吸收基因的表达。鳗弧菌与嗜水气单胞菌在对数生长前期6 h就表现出明显AI-2活性[31-32]。这些研究结果意味着Vh-RbsB在限铁条件下可能在生长早期就发挥其生理学功能,富铁条件下Vh-RbsB在高细胞密度下即在生长的稳定期发挥功能。由于本研究并未同步构建rbsB基因缺失株及回补株,并对其他铁摄取和代谢相关的基因进行表达差异的分析,结果具有部分局限性。
对于一些细菌而言,通过在培养液中添加核糖,RbsB介导抑制哈维弧菌发光是可逆的,说明核糖与AI-2分子共同竞争RbsB结合位点,核糖可影响AI-2的活性及生物膜的形成[15,26];然而对于某些细菌,AI-2前体DPD的摄取并未受到核糖或其他戊糖的竞争性抑制[30]。本研究成功构建pGEX-4t-1-rbsB重组质粒并转化至大肠埃希菌BL21 (DE3)中,得到重组蛋白的相对分子量约为59 kD,IPTG浓度不影响重组蛋白的表达。本研究可为研制哈维弧菌亚单位疫苗提供理论基础。
-
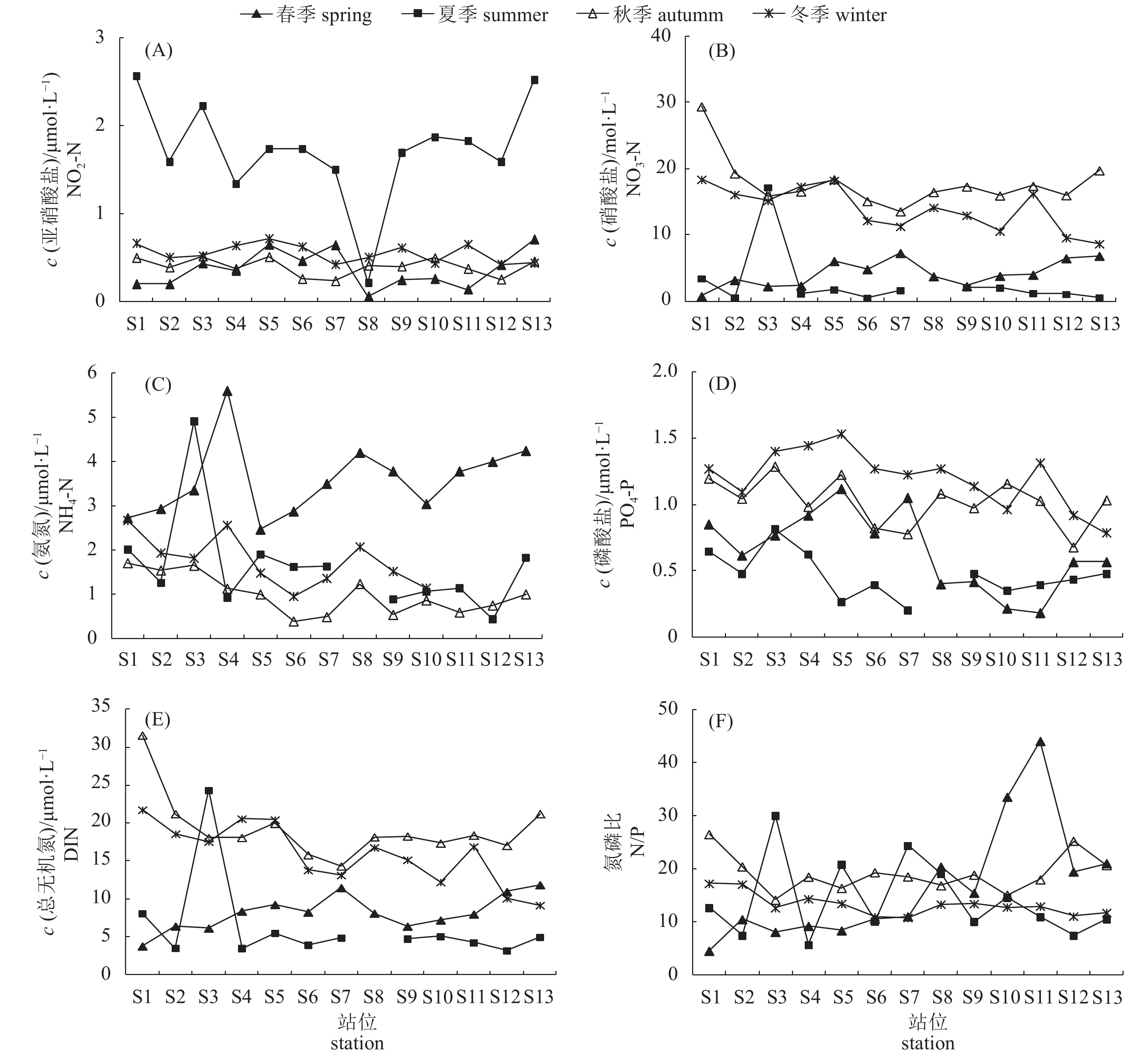
![]()
图 2 深澳湾氮、磷营养盐浓度时空变化
Figure 2. Tempo-spatial variation of nitrogen and phosphate nutrients in Shen'ao Bay
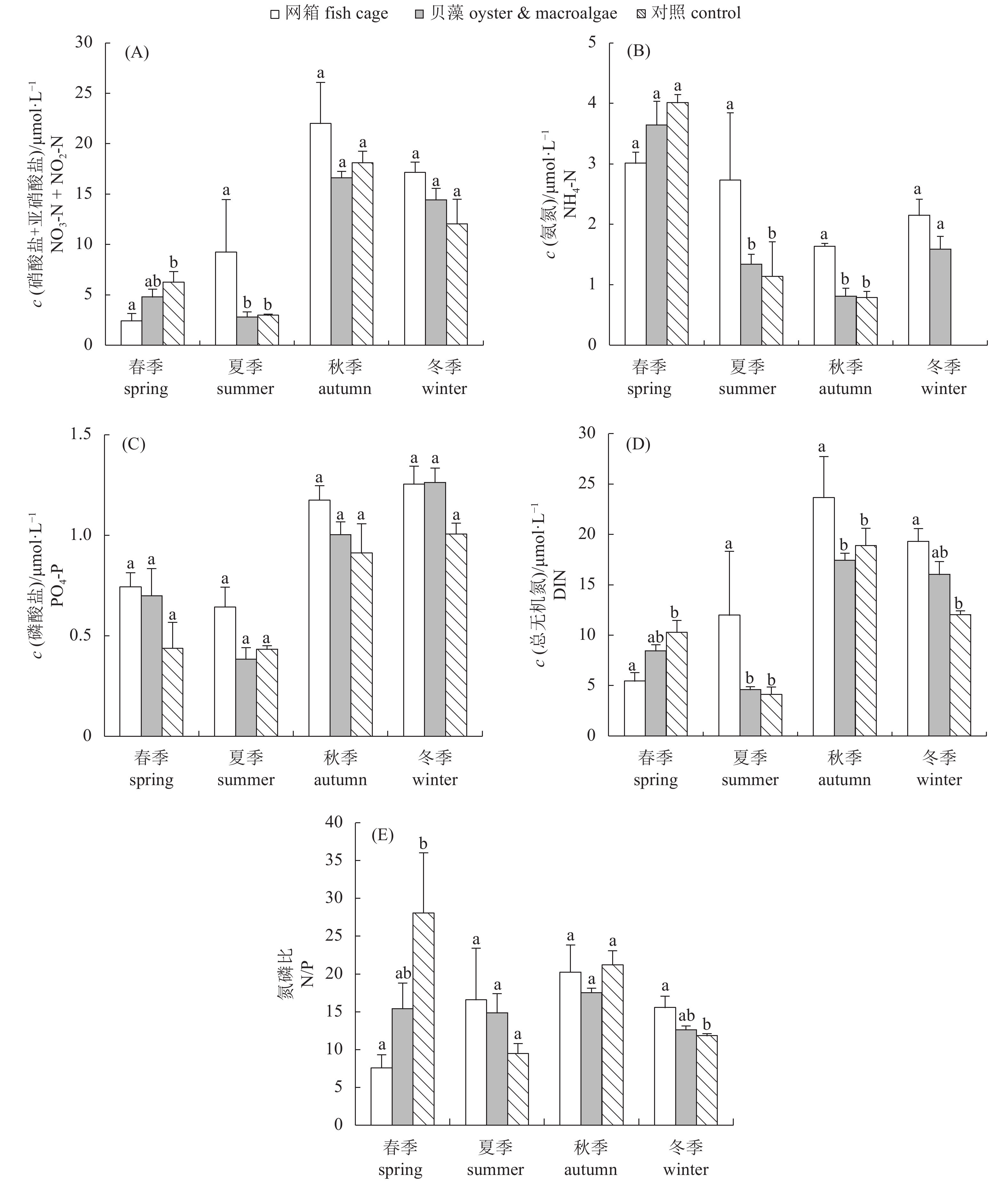
![]()
图 3 不同区域氮磷营养盐浓度和氮磷比的季节变化
同一季节内相同字母表示差异不显著(P>0.05)
Figure 3. Seasonal variation of nitrogen and phosphate nutrients in Shen'ao Bay
Values with the same superscript letters indicate no significant difference (P>0.05).
表 1 营养级的划分原则
Table 1 Classification of trophic level
级别
grade营养级
trophic levelc (总无机氮)/μmol·L–1
DINc (磷酸盐)/μmol·L–1
PO4-P氮磷比
N/PⅠ 贫营养 <14.28 <0.97 8~30 Ⅱ 中度营养 14.28~21.41 0.97~1.45 8~30 Ⅲ 富营养 >21.41 >1.45 8~30 ⅣP 磷限制中度营养 14.28~21.41 − >30 ⅤP 磷中等限制潜在性富营养 >21.41 − 30~60 ⅥP 磷限制潜在性富营养 >21.41 − >60 ⅣN 氮限制中度营养 − 0.97~1.45 <8 ⅤN 氮中等限制潜在性富营养 − >1.45 4~8 ⅥN 氮限制潜在性富营养 − >1.45 <4  下载: 导出CSV
下载: 导出CSV
表 2 深澳湾海水环境因子季节变化
Table 2 Seasonal variation in environmental parameters in Shen'ao Bay
$\overline {{\mathit{\boldsymbol{X}}}} \pm {\bf{SD}} $ 季节
season温度/℃
temperature盐度
salinityρ(溶解氧)/mg·L−1
DOρ(叶绿素a)/μg·L−1
Chl apH 春季 spring 22.0±0.8 (20.8~23.3) 30.3±0.6 (28.8~30.9) 11.6±2.5 (8.9~11.5) 2.01±0.33 (1.62~2.69) 8.3±0.1 (8.2~8.4) 夏季 summer 28.2±0.5 (27.6~29.1) 32.2±0.4 (31.5~32.7) 5.22±0.6 (3.9~5.9) 3.06±0.64 (1.95~4.16) − 秋季 autumn 20.7±0.3 (20.4~21.2) 32.7±0.1 (32.6~32.9) 7.0±0.2 (6.7~7.3) 1.33±0.21 (1.01~1.63) 8.0±0.1 (7.8~8.1) 冬季 winter 14.9±0.1 (14.8~15.0) 32.5±0.0 (32.4~32.5) 7.9±0.1 (7.7~8.3) 0.99±0.21 (0.75~1.47) 7.8±0.1 (7.6~7.9) 注:−. 没有采集到数据 Note: −. no data
下载: 导出CSV
表 3 深澳湾海水中氮磷营养盐浓度、氮磷比及营养级
Table 3 Seasonal variation in nitrogen and phosphate concentrations, N/P ratio and trophic level of seawater in Shen'ao Bay
$ \overline {\mathit{\boldsymbol{X }}} {\mathit{\boldsymbol{\pm}}} {\bf SD}$ 季节
seasonc(亚硝酸盐)/μmol·L–1
NO2-Nc(硝酸盐)/μmol·L–1
NO3-Nc(氨氮)/μmol·L–1
NH4-Nc(总无机氮)/μmol·L–1
DINc(磷酸盐)/μmol·L–1
PO4-P氮磷比
N/P营养级
trophic level春季 spring 0.37±0.06 4.22±0.56 3.58±0.23 8.17±0.64 0.65±0.08 16.6±3.1 Ⅰ 夏季 summer 1.72±0.17 2.83±1.33 1.64±0.33 6.32±1.68 0.46±0.05 13.6±2.2 Ⅰ 秋季 autumn 0.40±0.03 17.81±1.07 1.00±0.12 19.21±1.16 1.02±0.05 19.0±1.0 Ⅱ 冬季 winter 0.55±0.03 13.96±0.91 1.76±0.16 15.86±1.12 1.20±0.06 13.1±0.6 Ⅱ 平均值 mean 0.87±0.08 8.92±0.74 2.36±0.15 12.39±0.95 0.88±0.04 14.3±0.7 Ⅰ
下载: 导出CSV
表 4 深澳湾不同形态氮占总无机氮的比例
Table 4 Seasonal variation of dissolved inorganic nitrogen composition in Shen'ao Bay
% 季节
season亚硝酸盐
NO2-N硝酸盐
NO3-N氨氮
NH4-N春季 spring 4.5 51.7 43.8 夏季 summer 27.2 44.8 25.9 秋季 autumn 2.1 92.7 5.2 冬季 winter 3.5 88.0 11.1 平均值 mean 9.3 69.3 21.5
下载: 导出CSV
-
[1] ZEHR J P, KUDELA R M. Nitrogen cycle of the open ocean: from genes to ecosystems[J]. Ann Rev Mar Sci, 2010, 3(1): 197.
[2] STEVENS C J. Nitrogen in the environment[J]. Science, 2019, 363(6427): 578-580. doi: 10.1126/science.aav8215
[3] REDFIELD A C. The biological control of chemical factors in the environment[J]. Sci Prog, 1960, 11: 150-170.
[4] LAGUS A, SUOMELA J, WEITHOFF G, et al. Species-specific differences in phytoplankton responses to N and P enrichments and the N: P ratio in the Archipelago Sea, northern Baltic Sea[J]. J Plankton Res, 2004, 33(5/6): 529-551.
[5] GIBBS M T. Sustainability performance indicators for suspended bivalve aquaculture activities[J]. Ecol Indic, 2007, 7(1): 94-107. doi: 10.1016/j.ecolind.2005.10.004
[6] 周毅频, 李绪录, 夏华永. 大鹏湾海水中氮和磷的分布变化[J]. 广东海洋大学学报, 2011, 31(3): 50-55. doi: 10.3969/j.issn.1673-9159.2011.03.009 [7] 李凡. 海岸带陆海相互作用(LOICZ)研究及我们的策略[J]. 地球科学进展, 1996, 11(1): 19-23. doi: 10.3321/j.issn:1001-8166.1996.01.006 [8] 刘瑞玉, 胡敦欣. 中国的海岸带陆海相互作用(LOICZ)研究[J]. 地学前缘, 1997(Z1): 198. [9] BOUWMAN L, GOLDEWIJK K K, van der HOEK K W, et al. Exploring global changes in nitrogen and phosphorus cycles in agriculture induced by livestock production over the 1900−2050 period[J]. Proc Natl Acad Sci USA, 2013, 110(52): 20882-20887. doi: 10.1073/pnas.1012878108
[10] VANNI M J, MCINTYRE P B. Predicting nutrient excretion of aquatic animals with metabolic ecology and ecological stoichiometry: a global synthesis[J]. Ecology, 2016, 97(12): 3460-3471. doi: 10.1002/ecy.1582
[11] VERDEGEM M C. Nutrient discharge from aquaculture operations in function of system design and production environment[J]. Rev Aquacult, 2013, 5(3): 158-171.
[12] BERMAN T, BRONK D. Dissolved organic nitrogen: a dynamic participant in aquatic ecosystems[J]. Aquat Microbial Ecol, 2003, 31(3): 279-305.
[13] 魏皓, 赵亮, 原野, 等. 桑沟湾水动力特征及其对养殖容量影响的研究——观测与模型[J]. 渔业科学进展, 2010, 31(4): 65-71. doi: 10.3969/j.issn.1000-7075.2010.04.009 [14] 史洁, 魏皓. 半封闭高密度筏式养殖海域水动力场的数值模拟[J]. 中国海洋大学学报(自然科学版), 2009, 39(6): 1181-1187. [15] 杜虹, 郑兵, 陈伟洲, 等. 深澳湾海水养殖区水化因子的动态变化与水质量评价[J]. 海洋与湖沼, 2010, 41(6): 816-823. [16] 彭璇, 马胜伟, 陈海刚, 等. 夏季柘林湾-南澳岛海洋牧场营养盐的空间分布及其评价[J]. 南方水产科学, 2014, 10(6): 27-35. doi: 10.3969/j.issn.2095-0780.2014.06.004 [17] 王亮根, 杜飞雁, 陈丕茂, 等. 南澳岛北部海域浮游动物生态学特征及水团影响[J]. 南方水产科学, 2016, 12(5): 23-33. doi: 10.3969/j.issn.2095-0780.2016.05.004 [18] 郭卫东, 章小明. 中国近岸海域潜在性富营养化程度的评价[J]. 应用海洋学学报, 1998(1): 64-70. [19] WANG W L, MOORE J K, MARTINY A C, et al. Convergent estimates of marine nitrogen fixation[J]. Nature, 2019, 566(7743): 205. doi: 10.1038/s41586-019-0911-2
[20] MOORE C M, MILLS M M, ARRIGO K R, et al. Processes and patterns of oceanic nutrient limitation[J]. Nat Geosci, 2013, 6(9): 701-710. doi: 10.1038/ngeo1765
[21] KRISHNAMURTHY A, MOORE J K, MAHOWALD N, et al. Impacts of atmospheric nutrient inputs on marine biogeochemistry[J]. J Geophys Res Biogeosci, 2010, 115(G1): 93-101.
[22] HOEGH-GULDBERG O, BRUNO J F. The impact of climate change on the world's marine ecosystems[J]. Science, 2010, 328(5985): 1523-1528. doi: 10.1126/science.1189930
[23] 马媛, 魏巍, 夏华永, 等. 珠江口伶仃洋海域营养盐的历史变化及影响因素研究[J]. 海洋学报(中文版), 2009, 31(2): 69-77. doi: 10.3321/j.issn:0253-4193.2009.02.010 [24] MICHELI F, HALPERN B S, WALBRIDGE S A, et al. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: assessing current pressures and opportunities[J]. PLoS One, 2013, 8(12): e79889. doi: 10.1371/journal.pone.0079889
[25] CERCO C F, NOEL M R. Can oyster restoration reverse cultural eutrophication in Chesapeake Bay?[J]. Estuar Coasts, 2007, 30(2): 331-343. doi: 10.1007/BF02700175
[26] SHU T, WEN Y, YONGSEN L U, et al. Research on material balance of N and P in cage culture: take example for cage culture in Yaling Bay, Guangdong[J]. Acta Scientiae Circumstantiae, 2004, 24(6): 1046-1052.
[27] KELLOGG M L, SMYTH A R, LUCKENBACH M W, et al. Use of oysters to mitigate eutrophication in coastal waters[J]. Estuar Coast Shelf Sci, 2014, 151(5): 156-168.
[28] 毛玉泽, 李加琦, 薛素燕, 等. 海带养殖在桑沟湾多营养层次综合养殖系统中的生态功能[J]. 生态学报, 2018, 38(9): 1-7. doi: 10.3969/j.issn.1673-1182.2018.09.001 [29] AJJABI L C, ABAAB M, SEGNI R. The red macroalga Gracilaria verrucosa in co-culture with the Mediterranean mussels Mytilus galloprovincialis: productivity and nutrient removal performance[J]. Aquacult Int, 2018, 26(1): 253-266. doi: 10.1007/s10499-017-0206-2
[30] GRANT J B C. A numerical model of flow modification induced by suspended aquaculture in a Chinese bay[J]. Can J Fish Aquat Sci, 2001, 58(5): 1003-1011. doi: 10.1139/f01-027
[31] 刘慧, 方建光, 董双林, 等. 莱州湾和桑沟湾养殖海区主要营养盐的周年变动及限制因子[J]. 中国水产科学, 2003, 10(3): 227-234. doi: 10.3321/j.issn:1005-8737.2003.03.010 [32] 张继红, 蒋增杰, 王巍, 等. 桑沟湾营养盐时空分布及营养盐限制分析[J]. 渔业科学进展, 2010, 31(4): 16-25. doi: 10.3969/j.issn.1000-7075.2010.04.003 [33] 毛玉泽, 杨红生, 周毅, 等. 龙须菜(Gracilaria lemaneiformis)的生长、光合作用及其对扇贝排泄氮磷的吸收[J]. 生态学报, 2006, 26(10): 3225-3231. doi: 10.3321/j.issn:1000-0933.2006.10.009 [34] ZHOU Y, YANG H S, HU H Y, et al. Bioremediation potential of the macroalga Gracilaria lemaneiformis (Rhodophyta) integrated into fed fish culture in coastal waters of North China[J]. Aquaculture, 2006, 252(2/3/4): 264-276.
[35] XU Y J, FANG J, WEI W. Application of Gracilaria lichenoides (Rhodophyta) for alleviating excess nutrients in aquaculture[J]. J Appl Phycol, 2008, 20(2): 199-203. doi: 10.1007/s10811-007-9219-y
[36] XU Z G, ZOU D H, GAO K S. Effects of elevated CO2 and phosphorus supply on growth, photosynthesis and nutrient uptake in the marine macroalga Gracilaria lemaneiformis (Rhodophyta)[J]. Botanica Marina, 2010, 53(2): 123-129.
[37] DUAN Y, YANG N, HU M, et al. Growth and nutrient uptake of Gracilaria lemaneiformis under different nutrient conditions with implications for ecosystem services: a case study in the laboratory and in an enclosed mariculture area in the East China Sea[J]. Aquat Botany, 2019, 153: 73-80. doi: 10.1016/j.aquabot.2018.11.012
[38] 谢琳萍, 蒲新明, 孙霞, 等. 荣成湾营养盐的时空分布特征及其影响因素分析[J]. 海洋通报, 2013, 32(1): 19-27. [39] MAO Y, ZHOU Y, YANG H, et al. Seasonal variation in metabolism of cultured Pacific oyster, Crassostrea gigas, in Sanggou Bay, China[J]. Aquaculture, 2006, 253(1): 322-333.
[40] 贾后磊, 温琰茂, 舒廷飞. 哑铃湾网箱养殖水环境中的营养盐含量及特征[J]. 海洋环境科学, 2003, 22(3): 12-15. doi: 10.3969/j.issn.1007-6336.2003.03.003 [41] 黄显兵, 杜虹, 黄洪辉, 等. 深澳湾表层底质沉积物中凯氏氮和总磷的时空分布及与周边地区的比较[J]. 中国农学通报, 2011, 27(6): 333-337. [42] JUSTIC D, RABALAIS N N, TURNER R E. Stoichiometric nutrient balance and origin of coastal eutrophication[J]. Mar Pollut Bull, 1995, 30(1): 41-46. doi: 10.1016/0025-326X(94)00105-I
[43] VITOUSEK P M, HOWARTH R W. Nitrogen limitation on land and in the sea: how can it occur?[J]. Biogeochemistry, 1991, 13(2): 87-115.
[44] 郝林华, 孙丕喜, 郝建民, 等. 桑沟湾海域叶绿素a的时空分布特征及其影响因素研究[J]. 生态环境学报, 2012, 21(2): 338-345. doi: 10.3969/j.issn.1674-5906.2012.02.024 [45] REDFIELD A C, KETCHUM B H, RICHARDS F A. The influence of organisms on the composition of the sea water[J]. Intersci Publishers, 1963, 24(6): 26-27.
[46] WEBER T S, DEUTSCH C. Ocean nutrient ratios governed by plankton biogeography[J]. Nature, 2010, 467(7315): 550-554. doi: 10.1038/nature09403
[47] SCHINDLER D W. Eutrophication and recovery in experimental lakes: implications for lake management[J]. Science, 1974, 184(4139): 897-899. doi: 10.1126/science.184.4139.897
[48] ZEHR J P. Nitrogen fixation by marine cyanobacteria[J]. Trends Microbiol, 2011, 19(4): 162-173. doi: 10.1016/j.tim.2010.12.004
[49] 彭欣, 宁修仁, 孙军, 等. 南海北部浮游植物生长对营养盐的响应[J]. 生态学报, 2006, 26(12): 3959-3968. doi: 10.3321/j.issn:1000-0933.2006.12.006 [50] FREDE T T, LI Z U, FEREIDOUN R. P limitation of heterotrophic bacteria and phytoplankton in the northwest Mediterranean[J]. Limnol Oceanogr, 1998, 43(1): 88-94.
[51] ZOHARY T, ROBARTS R D. Experimental study of microbial P limitation in the eastern Mediterranean[J]. Limnol Oceanogr, 1998, 43(3): 387-395. doi: 10.4319/lo.1998.43.3.0387
[52] KIRKKALA T, HELMINEN H, ERKKILA A. Variability of nutrient limitation in the Archipelago Sea, SW Finland[J]. Hydrobiologia, 1998, 363(1/2/3): 117-126.
[53] ALLEN J I, BLACKFORD J C, RADFORD P J. An 1-D vertically resolved modelling study of the ecosystem dynamics of the middle and southern Adriatic Sea[J]. J Mar Syst, 1998, 18(1/2/3): 265-286.
[54] 李纯厚, 徐姗楠, 杜飞雁, 等. 大亚湾生态系统对人类活动的响应及健康评价[J]. 中国渔业质量与标准, 2015, 5(1): 1-10. [55] 徐东会, 陈碧鹃, 江涛, 等. 桑沟湾养殖海域营养盐时空分布特征及富营养化评价[J]. 渔业科学进展, 2017, 38(3): 1-11. [56] 周凯. 2000~2001年粤东大规模增养殖区柘林湾和深澳湾的生态学研究[D]. 广州: 暨南大学, 2002: 96. [57] JONES A B, PRESTON N P, DENNISON W C. The efficiency and condition of oysters and macroalgae used as biological filters of shrimp pond effluent[J]. Aquacult Res, 2015, 33(1): 1-19.
[58] STADMARK J, CONLEY D J. Mussel farming as a nutrient reduction measure in the Baltic Sea: consideration of nutrient biogeochemical cycles[J]. Mar Pollut Bull, 2011, 62(7): 1385-1388. doi: 10.1016/j.marpolbul.2011.05.001
[59] 丘耀文. 珠江口水体的三氮特征[J]. 热带海洋学报, 1992(3): 84-88. [60] 郑爱榕, 沈海维, 刘景欣, 等. 大亚湾海域低营养盐维持高生产力的机制探讨Ⅰ[J]. 海洋科学, 2001, 25(11): 48-52. doi: 10.3969/j.issn.1000-3096.2001.11.014 [61] 舒廷飞, 温琰茂, 汤叶涛. 养殖水环境中氮的循环与平衡[J]. 水产科学, 2002, 21(2): 30-34. doi: 10.3969/j.issn.1004-2091.2002.02.015 [62] 周毅, 杨红生, 何义朝, 等. 四十里湾几种双壳贝类及污损动物的氮、磷排泄及其生态效应[J]. 海洋与湖沼, 2002, 33(4): 424-431. doi: 10.3321/j.issn:0029-814X.2002.04.012 [63] GLIBERT P M, WILKERSON F P, DUGDALE R C, et al. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions[J]. Limnol Oceanogr, 2016, 61(1): 165-197. doi: 10.1002/lno.10203
-
期刊类型引用(1)
1. 薛尊,石建高,张文阳. 中国重力式深水网箱研究进展和展望. 渔业研究. 2024(04): 393-402 .  百度学术
百度学术
其他类型引用(1)







计量
- 文章访问数: 4969
- HTML全文浏览量: 2312
- PDF下载量: 69
- 被引次数: 2
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
