
Comparison of seven metallic elements in prismatic layer and nacreous layer of shells from four shell color strains in pearl oyster (Pinctada fucata)
-
摘要:
利用电感耦合等离子体发射光谱法(ICP-AES)对合浦珠母贝(Pinctada fucata)黑、金、红、白F5代选育群体贝壳棱柱层和珍珠质层的7种金属元素质量分数进行比较分析。结果显示, 合浦珠母贝贝壳棱柱层中铁(Fe)、钾(K)、镁(Mg)和锰(Mn)的质量分数明显高于珍珠质层, 而钠(Na)、Sr(锶)和锌(Zn)的质量分数则低于珍珠质层。4种壳色棱柱层和珍珠质层中的金属元素质量分数均存在差异: Mg、Sr、K、Zn在黑壳合浦珠母贝棱柱层中质量分数最低, Mn、Fe在金壳棱柱层中最低, 而Na却在红壳棱柱层中最低; 单因素方差分析显示, Fe、Mg、Zn可能分别会影响合浦珠母贝红壳、金壳、白壳的形成。白壳贝珍珠质层的6种金属元素(Mg除外)质量分数都最少, 更有可能培育出优质珍珠。
Abstract:We analyzed the contents of seven metallic elements in prismatic layer and nacreous layer of shells from four shell color strains [(B), gold (G), red (R) and white (W)] in pearl oyster (Pinctada fucata) by ICP-AES. The Fe, K, Mg and Mn contents in prismatic layer were obviously higher than those in nacreous layer. The elements of Mg, Sr, K and Zn in prismatic layer of black shell were the lowest. Similarly, the Mn and Fe contents in prismatic layer of gold shell were lower than those in the other color shells, but Na had minimum content in red shell. Fe, Mg and Zn might have influence on the formation of red, gold and white shells. Besides, the nacreous layer of white shell had the lowest level in the content of analytical elements except Mg, which indicates that the white shell in pearl oysters is more likely to produce pearls of better quality.
-
Keywords:
- Pinctada fucata /
- shell color /
- metallic elements /
- prismatic layer /
- nacreous layer
-
九孔鲍(Haliotis diversicolor supertexta)作为一种非常有营养价值与经济价值的海产贝类,在中国南方沿海被广泛养殖。为满足日益增长的市场需求,养殖规模不断扩大。但是由于长期的近亲繁殖和不注重亲鲍培育等原因导致种质退化、育苗成功率低、养殖时间延长且个体偏小等问题, 这严重制约了九孔鲍养殖业的发展。为了解决这些突出问题,国内外水产育种工作者都做了大量的研究工作,种内群体间杂交是贝类杂种优势利用与新品种培育的主要途经之一[1-5]。
杂种优势是指2个遗传结构不同的亲本群体杂交所产生的杂种群体在生长、繁殖、成活、抗逆产量和品质上比其亲本表现优越的现象。张国范等[6]率先利用不同地理群体的皱纹盘鲍(H.discus hannai)杂交培育出“大连1号”杂交鲍并通过国家新品种审定,极大促进中国鲍鱼养殖业的健康可持续发展。柯才焕和游伟伟[7]用杂色鲍(H.diversicolor aquatili)台湾群体(♀)×日本群体(♂)杂交并经过多代评估与选种获得新品种“东优1号”,其抗病力较原有养殖种有显著提高,增产效果显著,对重振南方杂色鲍养殖产业发挥了重要作用。YOU等[3]用3个九孔鲍群体完全双列杂交选出1个生长杂种优势(6.84%)和成活率杂种优势(69.09%)均较高的群体。杂交及杂种优势的利用已是鲍鱼高效养殖的核心关键技术。
杂交育种过程中,杂种表现的优劣主要取决于杂交双亲。配合力是亲本选育和组合选配的重要遗传参数。关于贝类不同群体间杂交的配合力分析,前人做过很多相关研究,邓岳文[8]通过3个不同群体皱纹盘鲍的完全双列杂交得出了随着养殖日龄的增加,一般配合力逐渐下降,特殊配合力增加,大部分组合的优势率增加的结论。谷龙春等[9]通过对不同地理群体的合浦珠母贝(Pinctada fucata)进行杂交并对配合力与杂种优势进行了分析,结果表明北海的亲本具有很好的杂种优势与配合力。王炳谦等[10]以不同遗传背景的虹鳟(Oncorhynchus mykiss)作为亲本进行杂交并分析亲本间配合力,有望培育出杂种优势率较高的优势组合。至于在杂种优势与配合力的关系研究方面,中国的研究结果不尽一致,如毕详等[11]的研究认为配合力与杂种优势呈极显著相关(R=0.959)。然而周国峰等[12]研究表明两者强弱并不一致。关于九孔鲍的配合力与杂种优势分析研究少见报道,该试验选用2个不同地理群体的九孔鲍通过完全双列杂交设计方法建立4个杂交组合,分析杂交组合杂种优势,并利用线性模型均数加权二乘分析法估算主要九孔鲍各发育阶段生长性状的配合力,为九孔鲍的新品种选育与杂种优势利用提供理论参考与基础数据。
1. 材料与方法
1.1 亲本来源
试验用九孔鲍养殖群体取自汕尾多代养殖群体,遗传背景为中国九孔鲍养殖群体与日本野生群体杂交子代自繁子二代(引自福建省东山县),平均壳长为5.46 cm;野生群体来源于深圳附近海域,平均壳长为7.63 cm,选择健康、性腺发育良好的亲本来进行试验。
1.2 试验设计
利用九孔鲍养殖群体(BD)与深圳野生群体(YS)通过完全双列杂交设计4个组合的杂交与自繁试验组,每个试验组合设3个平行组,共得到12组F1代,即杂交组为BD♀×YS♂(正交),YS♀×BD♂(反交),自繁组为YS♀×YS♂,BD♀×BD♂。12个组合分别养殖于12个小池内(100 cm×100 cm×80 cm)并置于同一个大水泥池中(7 m×3 m×1.35 m),保证养殖条件一致,降低环境因子影响,保证试验的准确性。
1.3 催产与授精
2011年6月,自繁组与杂交组亲本分别诱导产卵,综合运用阴干,紫外线照射海水与变温刺激方法,过程为本底温度23.5 ℃阴干2~2.5 h,放入新鲜海水中适应1 h,再置于经紫外线照射过的海水进行刺激,温度变化为常温26.5 ℃经过1.5 h降温至22 ℃,再经过1 h升温至25.5 ℃,最后经0.5 h升至28 ℃,若亲鲍还未排卵(精),则按上述步骤重复1次,接着将鲍卵收集到一塑料容器中,加入新鲜精液缓慢搅匀静置授精,30~40 min洗卵1次,经3~5次洗卵后放入培苗池中,根据受精卵密度每个小池布置受精卵3×104个左右,按照传统的鲍鱼育苗方式培育。
1.4 生长和存活性状测量
幼体发育阶段持续充氧,稚鲍培育期用人工饵料进行喂养,每天换水100%,冲洗保持水质清洁。40 d左右稚鲍剥离至四角砖下,平均每平方米稚鲍为6 000个左右,当幼鲍达到2.5 cm左右时分别装笼,每笼放25~30个,投喂嫩江蓠、海带等。每个杂交组合有严格标记。待分别养殖至120日龄、150日龄、210日龄和270日龄时每个组合随机取九孔鲍幼鲍30个测定壳长、壳宽与体质量生长数据用于杂种优势与配合力分析。
1.5 数据处理与分析
1.5.1 杂种优势H的计算
参照PEDRO和IBARRA[13]的试验方法,用下面2个公式来计算杂种优势(heterosis):
$$ \mathrm{MP}(\%)=\frac{F_1-P_0}{P_0} \times 100 $$ $$ \operatorname{BP}(\%)=\frac{F_1-P_B}{P_B} \times 100 $$ 式中MP代表中亲优势,F1为杂交组均值,P0为相应双亲均值,BP代表超亲优势,PB为杂交组较好的亲本均值。
1.5.2 配合力分析模型
利用SPSS 17.0软件对生长性状数据进行初步整理,当差异显著时,进行下一步的配合力分析。参照王炳谦等[10]的试验方法,配合力分析统计模型为:
$$ Y_{i j k}=u+S+D_j+S D_{i j}+E_{i j k} $$ 其中Yijk是第i个父本群体与第j个母本群体的第k个杂种个体的表型观测值,u是总体均数,Si为第i个父本一般配合力效应,Dj是第j个母本一般配合力效应,SDij为第i个父本与第j个母本的互作效应即正交特殊配合力效应,Eijk为随机误差效应,假设它独立地服从N(0,σe2)分布。整个群体的变异可以剖分为一般配合力(GCA)、特殊配合力(SCA)、随机误差等组成部分。
模型采用二因素有互作的均数加权最小二乘分析法进行配合力分析。其正规方程为:
$$ \left(X^{\prime} X\right) b=X^{\prime} Y $$ 其约束条件为:
$$ \sum\limits_i=\sum\limits_j D=\sum\limits_i D=\sum\limits_j D=\sum \sum\limits_{i j} D=\mathbf{0} $$ 2. 结果与分析
2.1 不同日龄生长和存活性状的统计性描述量
试验测量了4个不同日龄阶段,每次统计360只九孔鲍数据。单因素方差分析多重比较显示,各交配组合壳长、壳宽、体质量性状和成活率差异均达到显著水平(P < 0.05)或极显著水平(P < 0.01)(表 1)。综合120~270日龄来看,反交组(YS♀×BD♂)的壳长、壳宽和体质量等生长性状数据一直高于正交组和自繁组,而群体的成活率性状正交组最高。各组的变异系数为0.04~0.47。其中体质量变异系数较大(0.33~0.47),说明此性状有很大的改良潜力;而不同生长阶段各生长性状的变异系数变化不大,说明这些性状的遗传性能比较稳定。
表 1 九孔鲍杂交群体生长和存活性状的描述性统计量Table 1. Descriptive statistics of growth and survival traits of hybrid populations of H.diversicolor supertexta性状
trait日龄
dph养殖×野生
BD♀×YS♂变异系数
CV野生×养殖
YS♀×BD♂变异系数
CV野生×野生
YS♀×YS♂变异系数
CV养殖×养殖
BD♀×BD♂变异系数
CV壳长/cm shell length 120 1.23±0.26Aa 0.21 1.30±0.23Aa 0.18 1.19±0.17ABa 0.14 0.98±0.15Bb 0.15 150 1.91±0.43Bb 0.23 2.16±0.45Aa 0.21 1.83±0.31Bb 0.17 1.65±0.32Cc 0.21 210 2.38±0.53Bc 0.22 2.69±0.54Aa 0.20 2.56±0.42ABb 0.16 2.26±0.44Bc 0.20 270 2.86±0.48Cc 0.17 3.45±0.69Aa 0.20 3.10±0.52Bb 0.17 2.72±0.53Cc 0.20 壳宽/cm shell width 120 0.80±0.14BbC 0.18 1.01±0.16Aa 0.16 0.85±0.12Bb 0.14 0.72±0.10Cc 0.14 150 1.29±0.21ABb 0.16 1.44±0.34Aa 0.24 1.28±0.22Bb 0.17 1.14±0.27ABc 0.24 210 1.47±0.36Bb 0.24 1.71±0.32Aa 0.19 1.61±0.24ABb 0.15 1.30±0.21Cc 0.16 270 1.81±0.48Bbc 0.27 2.07±0.40Aa 0.20 1.93±0.26ABab 0.13 1.72±0.26Bc 0.15 体质量/g body weight 150 1.46±0.57Ab 0.40 1.39±0.64Aab 0.46 1.60±0.66Aa 0.41 0.95±0.44Bc 0.46 210 2.41±0.94Aa 0.40 2.50±1.15ABa 0.46 2.56±1.07Aa 0.42 2.08±0.97Bb 0.47 270 3.05±1.01Aa 0.33 3.22±1.12ABa 0.35 3.37±1.33Aa 0.40 2.57±1.21Ab 0.47 成活率/% survival 120 41.96±1.41Aa 0.04 34.64±4.07ABab 0.12 34.05±6.04Bb 0.18 28.44±3.64Bb 0.13 150 37.66±1.78Aa 0.05 30.69±3.10ABab 0.10 27.60±6.95Bb 0.25 25.16±3.36Bb 0.13 210 34.82±1.81Aa 0.05 28.68±2.65ABab 0.10 25.01±5.76Bb 0.23 24.05±3.03Bb 0.13 270 33.34±1.62Aa 0.05 27.38±3.67ABb 0.13 22.26±3.43Bbc 0.15 20.20±2.85Bc 0.14 注:变异系数=标准差/平均值;同一行各组合均值上标注不同小写字母为差异显著(P < 0.05),标注不同大写字母为差异极显著(P < 0.01)
Note:coefficient of variation=standard deviation/mean;the values within the same line with different lowercase and uppercase letters are significantly and very significantly different at the levels of 0.01 and 0.05,respectively.2.2 不同日龄生长和存活性状杂种优势分析
九孔鲍不同日龄壳长、壳宽、体质量和存活性状杂种优势分析结果见表 2。壳长、壳宽、体质量和成活率的中亲优势和超亲优势不同生长阶段存在较大差异。壳长性状MP和BP的变化范围是-1.7% ~19.8%和-7.7% ~18.0%;壳宽性状MP和BP的变化范围是-0.8% ~28.7%和-8.7% ~18.8%;体质量性状MP和BP的变化范围是2.7% ~14.5%和-13.1% ~ -2.3%;存活性状MP和BP的变化范围是10.9% ~57.0%和1.7% ~49.8%。随着日龄的增加,壳长、壳宽和体质量性状的MP和BP值总体呈减小趋势,反交组的MP和BP均高于正交组。然而正交组存活性状的MP和BP较高。体质量性状并没有体现出超亲优势,均为负值。
表 2 九孔鲍群体杂交生长和存活性状的中亲优势与超亲优势Table 2. Heterosis and heterobeltiosis of growth and survival traits of mid-parent H.diversicolor supertexta性状
trait日龄
dph中亲优势 MP 超亲优势 BP 正交
BD♀×YS♂反交
YS♀×BD♂均值
mean正交
BD♀×YS♂反交
YS♀×BD♂均值
mean壳长/cm shell length 120 13.4 19.8 16.6 3.4 9.2 6.3 150 9.8 24.1 17.0 4.4 18.0 11.2 210 -1.2 11.6 5.2 -7.0 5.1 -1.0 270 -1.7 18.6 8.5 -7.7 11.3 1.8 壳宽/cm shell width 120 1.9 28.7 15.3 -5.9 18.8 6.5 150 6.6 19.0 12.8 0.8 12.5 6.6 210 1.0 17.5 9.3 -8.7 6.2 -1.2 270 -0.8 13.4 6.3 -6.2 7.3 0.5 体质量/g body weight 150 14.5 9.0 11.8 -8.8 -13.1 -10.9 210 3.9 7.8 5.9 -5.9 -2.3 -4.1 270 2.7 8.4 5.6 -9.5 -4.5 -7.0 成活率/% survival 120 34.3 10.9 22.6 23.2 1.7 12.5 150 42.8 16.3 29.6 36.4 11.2 23.8 210 41.9 16.9 29.4 39.2 14.7 26.9 270 57.0 29.0 43.0 49.8 23.0 36.4 2.3 一般配合力与特殊配合力效应值分析
壳长、壳宽与体质量3个性状在不同杂交组合中均达到显著性水平(P < 0.05),可以进一步进行配合力方差分析。BD群体各生长阶段壳长、壳宽和体质量性状的GCA效应值均为负值,范围分别为-0.19~-0.09、-0.11~-0.07和-0.40~-0.24;YS群体各生长阶段壳长、壳宽和体质量性状的GCA效应值均为正值,范围分别为0.09~0.19、0.07~0.11和0.24~0.40(表 3)。YS群体与BD群体各生长阶段壳长、壳宽和体质量性状的SCA效应值分别为0.01~0.14、0~0.07和0.02~0.07。综合3个生长性状来看,各发育阶段均存在母本效应,反交效应不明显。BD群体的一般配合力效应值、特殊配合力和母本效应均小于YS群体,因此野生群体亲本对杂交组的生长影响较大。随着日龄的增加,各生长性状反交效应值呈上升趋势。
表 3 九孔鲍野生群体与养殖群体杂交生长性状配合力效应值分析Table 3. Combining ability of growth traits of cultured and wild stocks of H.diversicolor supertexta性状
trait日龄 dph 壳长/cm shell length 壳宽/cm shell width 体质量/g body weight 100 160 220 280 100 160 220 280 100 220 280 一般配合力 general combining ability BD父本 -0.04 0.02 0 0.05 0.02 0 -0.02 0.01 -0.18 -0.10 -0.16 YS父本 0.04 -0.02 0 -0.05 -0.02 0 0.02 -0.01 0.18 0.10 0.16 BD母本 -0.07 -0.11 -0.15 -0.24 -0.09 -0.07 -0.14 -0.12 -0.15 -0.14 -0.24 YS母本 0.07 0.11 0.15 0.24 0.09 0.07 0.14 0.12 0.15 0.14 0.24 BD群体 -0.11 -0.09 -0.15 -0.19 -0.07 -0.07 -0.16 -0.11 -0.33 -0.24 -0.40 YS群体 0.11 0.09 0.15 0.19 0.07 0.07 0.16 0.11 0.33 0.24 0.40 母本效应 maternal effect BD群体 -0.04 -0.13 -0.16 -0.30 -0.11 -0.08 -0.12 -0.13 0.04 -0.05 -0.09 YS群体 0.04 0.13 0.16 0.30 0.11 0.08 0.12 0.13 -0.04 0.05 0.09 特殊配合力 specific combining ability 正交组 0.01 0.02 0.03 0.06 0 0.03 0.06 0.05 0.06 0.03 0.02 反交组 0.01 0.26 0.06 0.04 0 0.10 0.07 0.03 0.07 0.02 0.02 均值 0.01 0.14 0.05 0.05 0 0.07 0.07 0.04 0.07 0.03 0.02 反交效应 reciprocal effect 0 -0.24 -0.03 0.02 0 -0.07 -0.01 0.02 -0.01 0.01 0 2.4 九孔鲍群体壳长、壳宽和体质量的配合力方差组分
表 4中显示杂交组合配合力方差分量。综合各日龄阶段,壳长、壳宽和体质量性状的GCA和SCA方差组分差异显著。壳长、壳宽和体质量性状SCA方差组分较大,占表型方差组分的2.19%~44.78%。其中120日龄壳宽性状GCA方差组分占表型方差组的44.78%。壳长、壳宽和体质量性状SCA方差组分较小,如120日龄壳长性状的SCA方差组分仅占表型方差组分的0.29%。
表 4 九孔鲍群体壳长、壳宽和体质量性状配合力的方差组分Table 4. Variance components of combining ability of shell length, shell width and body weight of H.diversicolor supertexta性状
trait日龄
dph方差 variance 一般配合力方差
σGCA2特殊配合力方差
σSCA2残差
σE2表型方差
σp2一般配合力方差/表型方差/%
σGCA2/σp2特殊配合力方差/表型方差/%
σSCA2/σp2壳长/cm shell length 120 0.042 0.000 4 0.096 0.138 30.43 0.29 150 0.004 0.043 0 0.140 0.183 2.19 23.50 210 0.042 0.006 0 0.235 0.283 14.84 2.12 270 0.058 0.013 0 0.399 0.470 12.34 2.77 壳宽/cm shell width 120 0.030 0.000 2 0.037 0.067 44.78 0.30 150 0.010 0.008 0 0.054 0.072 13.89 11.11 210 0.034 0.009 0 0.081 0.124 27.42 7.26 270 0.080 0.007 0 0.139 0.226 35.40 3.10 体质量/g body weight 150 0.085 0.018 0 0.348 0.451 18.85 3.99 210 0.042 0.006 0 1.084 1.132 3.71 0.53 270 0.142 0.006 0 1.783 1.931 7.35 0.31 3. 讨论
双列杂交指一定数量不同基因型亲本轮换与其他亲本交配,使其包括所有的可能的杂交组合[14]。该杂交技术在20世纪70年代已经得到广泛应用,在水产动物育种中发挥重大作用,并取得客观的经济效益和社会效益[15-16]。双列杂交交配设计能够提供关于组合材料变异来源、遗传成分、遗传方差等方面的本质信息,预测杂交后代的实际表现能力,因而能有效地评估亲本及发现优异组合材料。此外,群体间的基因频率差异越大、遗传距离越远杂交产生的杂种优势就会越大[17]。该试验野生群体和养殖群体遗传背景不同,两者又完全地理隔离没有基因交流,因此利用九孔鲍野生群体和养殖群体双列杂交获得数量性状改良的杂种优势后代具备理论基础。
杂种优势是一种普遍的重要的生物学现象,该试验探讨了杂种优势在九孔鲍不同生长阶段的表达规律,发现随着养殖日龄的增加,在生长方面壳长、壳宽和体质量性状的中亲优势值呈逐渐减少趋势,在成活方面中亲优势值呈逐渐增长趋势;各生长性状的超亲优势并不明显。这与林红军等[18]报道的杂种优势表达规律是一致的,而与YOU等[3]研究的结果不同。笔者认为当中存在原因可能是2方面:1)该试验选用群体间存在较大遗传差异,遗传背景复杂,基因表达的时间、空间上存在较大差异;2)与游伟伟等[5]所选用九孔鲍材料不同,不同群体杂交选育效果不同,致使交配结果不同。
从数量遗传学角度上分析,一般配合力主要是由于基因的加性效应所致; 特殊配合力则是双亲杂交后通过互作表现出的基因的显性、超显性和上位效应,即非加性效应[19]。因此对九孔鲍养殖群体和野生群体一般配合力效应值分析,可初步预测其选育潜力。综合不同生长阶段各交配组合数据显示,野生群体壳长、壳宽和体质量性状GCA效应值均表现为正效应(0.07~0.4),该群体内存在可稳定遗传的加性遗传变异较为丰富。大量文献表明,遗传力越低,受基因非加性效应影响作用越大[9],所以通过配合力效应值可进一步分析和指导群体杂交选育。在水产动物育种中,王炳辉等[10]对虹鳟(Oncorhynchus mykiss),ZHENG等[20]对海湾扇贝(Argopectens irradias),MALLET和HALEY[21]对野生牡蛎(Concha ostreae)的研究均表明存在着较强的生长非加性效应。该试验综合分析正反交组合生长性状的特殊配合力效应值发现,120日龄至270日龄反交组群体壳长、壳宽和体质量性状SCA效应值正向优势较明显,说明反交组群体内存在较强的生长非加性效应,杂交优势明显,可参考作为留种群体。关于一般配合力与特殊配合力是否相关已有报道[22-23]。通过分析3个性状的配合力表明,各性状亲本一般配合力与杂交组合的特殊配合力之间相互独立,没有显著相关关系,这为选育一般配合力和特殊配合力高的强优势组合提供可能。此外,九孔鲍群体配合力方差组分分析显示,一般配合力方差组分和特殊配合力方差组分在表型方差中均占有一定的分量,一般配合力方差组分大于特殊配合力方差组分,说明加性效应和非加性效应共同影响九孔鲍生长性状的表现,且加性效应对生长性状表达的影响较大。
笔者通过对九孔鲍野生群体和养殖群体完全双列杂交设计,利用线性模型均数加权二乘分析法统计分析其生长性状配合力和杂种优势得出,增加野生群体在交配亲本中的数量可有效提高九孔鲍生长性能,可用于参考解决中国南方九孔鲍种质资源近交衰退、群体多样性下降的难题。
-
![]()
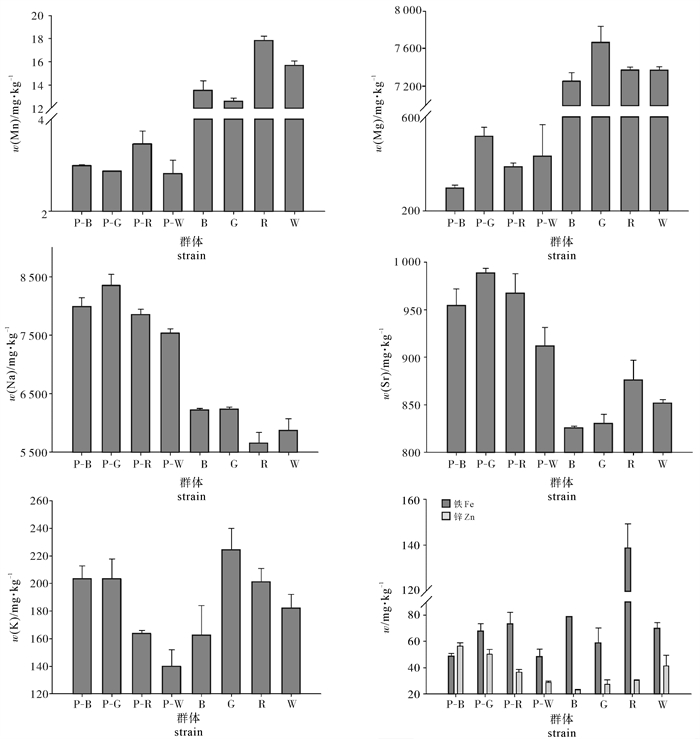
图 1 4个壳色合浦珠母贝珍珠质层和棱柱层的金属元素质量分数分布
Figure 1. Distribution of each metallic element in prismatic layer and nacreous layer of shells from four shell color strains of P.fucata
表 1 4种壳色合浦珠母贝贝壳棱柱层金属元素质量分数
Table 1 Metallic element contents in prismatic layer of shells from four shell color strains of P.fucata
mg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) 黑壳(B) 78.32±0.54a 162.57±21.47a 7 259.69±87.27a 13.45±0.81a 6 226.10±20.93a 825.75±1.86a 23.07±0.47a 金壳(G) 58.72±11.51a 224.38±15.44b 7 674.63±165.29b 12.57±0.28a 6 233.30±35.18a 830.27±9.66a 27.33±3.28ab 红壳(R) 138.66±10.56b 200.92±9.97b 7 378.63±31.03a 17.85±0.36b 5 657.00±179.97b 876.11±20.84b 30.31±0.47bc 白壳(W) 69.99±4.31a 182.07±10.02c 7 374.81±36.14a 15.67±0.40c 5 872.20±195.92b 851.69±3.82c 41.28±8.27d 平均值average 86.42±6.73 192.49±14.23 7 421.94±79.93 14.91±0.46 5 997.15±108.00 845.96±9.05 30.50±3.12 注:同列不同小写字母表示差异显著( P<0.05)
Note:Values with different letters in the same column have significant difference (P<0.05). 下载: 导出CSV
下载: 导出CSV
表 2 4种壳色合浦珠母贝贝壳珍珠质层金属元素质量分数
Table 2 Metallic element contents in nacreous layer of shells from four shell color strains of P.fucata
mg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) P-B 48.56±2.36a 203.36±9.28a 297.33±12.22a 2.99±0.015a 7 992.04±150.69a 954.75±17.16a 56.44±2.62a P-G 67.77±5.68b 203.16±14.69a 516.95±38.18b 2.88±0.002b 8 354.21±181.37a 988.45±5.01b 50.22±3.65a P-R 73.41±8.80c 163.62±2.44b 387.18±16.01c 3.47±0.268c 7 853.86±88.11b 967.71±20.24ab 36.73±1.73b P-W 48.26±5.92a 139.99±11.94c 432.85±133.36abc 2.81±0.295ab 7 542.58±66.52b 911.93±19.58c 28.90±0.87c 平均值average 59.50±5.69 177.53±9.59 408.58±49.94 3.04±0.145 7 935.67±121.67 955.71±15.50 43.07±2.22 注:P-B、P-G、P-R和P-W分别代表合浦珠母贝黑壳、金壳、红壳和白壳珍珠质层。同列不同小写字母表示差异显著(P<0.05)
Note:P-B,P-G,P-R and P-W represent nacreous layers of black,gold,red,white shells in pearl oyster,respectively. Values with different letters in the same column have significant difference (P<0.05).
下载: 导出CSV
-
[1] 龙敏明, 黄桂菊, 邹记兴, 等. 育珠对合浦珠母贝N 19和Prismalin-14基因表达水平的影响[J]. 南方水产科学, 2013, 9(5): 58-63. doi: 10.3969/j.issn.2095-0780.2013.05.010 [2] 张根芳, 许式见, 方爱萍. 三角帆蚌(Hyriopsis cumingii)外套膜无核珍珠颜色成因的育珠实验[J]. 海洋与湖沼, 2013, 44(2): 531-536. doi: 10.11693/hyhz201302042042 [3] 符韶, 谢绍河, 邓岳文, 等. 利用马氏珠母贝4个壳色系F3培育厚层优质珍珠[J]. 水产学报, 2012, 36(9): 1418-1424. doi: 10.3724/SP.J.1231.2012.27905 [4] 王庆恒, 逯云召, 邓岳文, 等. 马氏珠母贝生长性状与珍珠质量和珍珠层厚度的相关分析[J]. 广东海洋大学学报, 2013, 33(3): 18-21. doi: 10.3969/j.issn.1673-9159.2013.03.003 [5] TAYALE A, GUEGUEN Y, TREGUIER C, et al. Evidence of donor effect on cultured pearl quality from a duplicated grafting experiment on Pinctada margaritifera using wild donors[J]. Aquat Living Resour, 2012, 25(3): 269-280. doi: 10.1051/alr/2012034
[6] 王海. 珍珠与马氏珠母贝表型性状相关性以及矿化基因在珍珠囊和外套膜中的定量表达研究[D]. 海口: 海南大学, 2013. [7] KY C L, BLAY C, SHAM-KOUA M, et al. Family effect on cultured pearl quality in black-lipped pearl oyster Pinctada margaritifera and insights for genetic improvement[J]. Aquat Living Resour, 2013, 26(2): 133-145. doi: 10.1051/alr/2013055
[8] KY C L, BLAY C, SHAM-KOUA M, et al. Indirect improvement of pearl grade and shape in farmed Pinctada margaritifera by donor "oyster" selection for green pearls[J]. Aquaculture, 2014, 432(S1): 154-162. https://www.sciencedirect.com/science/article/pii/S0044848614002178
[9] ACOSTA-SALMÓN H, MARTÍNEZ-FERNÁNDEZ E, SOUTHGATE P C. Use of relaxants to obtain saibo tissue from the blacklip pearl oyster (Pinctada margaritifera) and the Akoya pearl oyster (Pinctada fucata)[J]. Aquaculture, 2005, 245(1/2/3/4): 167-172. https://www.sciencedirect.com/science/article/pii/S0044848604007264
[10] 王照旗, 韩学凯, 白志毅, 等. 三角帆蚌紫色选育系1龄阶段内壳色及生长性状的遗传参数估计[J]. 水产学报, 2014, 38(5): 644-650. https://www.cnki.com.cn/Article/CJFDTotal-SCKX201405004.htm [11] 张文府. 马氏珠母贝四种壳色选育系中育珠贝家系和小片贝家系的选育研究[D]. 湛江: 广东海洋大学, 2013. [12] 侯巍巍, 白金泉, 吴翠敏. 卟啉及金属卟啉化合物的应用研究新进展[J]. 广东化工, 2014, 41(2): 44-45. doi: 10.3969/j.issn.1007-1865.2014.02.023 [13] 张根芳, 叶容晖, 方爱萍. 珍珠颜色和贝壳珍珠层颜色研究进展[J]. 动物学杂志, 2014, 49(1): 137-144. https://www.cnki.com.cn/Article/CJFDTotal-BIRD201401024.htm [14] 张蕴韬. 卟啉及金属卟啉对珍珠颜色的贡献及致色机理研究[D]. 北京: 中国地质大学, 2006. [15] 严俊, 胡仙超, 王巨安, 等. 不同颜色的淡水养殖珍珠呈色机理研究[J]. 岩矿测试, 2013, 32(2): 263-268. doi: 10.3969/j.issn.0254-5357.2013.02.014 [16] 杨磊, 欧阳茜茜, 李思东, 等. 中国海水珍珠Mg、Fe、Zn和Mn含量变化及其对珍珠微观结构的影响[J]. 矿物学报, 2014, 34(3): 416-420. https://www.cnki.com.cn/Article/CJFDTotal-KWXB201403019.htm [17] 隋欣, 吴海铭, 王宝辉, 等. ICP-AES应用进展及展望[J]. 牡丹江师范学院学报: 自然科学版, 2014(1): 25-27. doi: 10.3969/j.issn.1003-6180.2014.01.013 [18] 王茂林, 张秀梅, 高天翔, 等. 水体钙离子质量浓度对褐牙鲆幼鱼鱼体和耳石元素成分的影响[J]. 南方水产科学, 2013, 9(3): 31-38. doi: 10.3969/j.issn.2095-0780.2013.03.006 [19] 木士春, 马红艳. 养殖珍珠微量元素特征及其对珍珠生长环境的指示意义[J]. 矿物学报, 2001, 21(3) : 551-553. doi: 10.3321/j.issn:1000-4734.2001.03.069 [20] 林静瑜, 谢莉萍. EDTA对合浦珠母贝碱性磷酸酶活性的影响[J]. 集美大学学报, 2004, 9(4): 313. doi: 10.3969/j.issn.1007-7405.2004.04.006 [21] 高玲, 曹洪玉, 刘阁. 4种金属卟啉配合物与血清白蛋白作用的光谱研究[J]. 化学试剂, 2012, 34(2): 108-112. doi: 10.3969/j.issn.0258-3283.2012.02.004 [22] MICHIO S, HIROMICHI N. Mollusk shell structures and their formation mechanism[J]. Can J Zool, 2013, 91(6): 349-366. doi: 10.1139/cjz-2012-0333
[23] JACOB D E, SOLDATI A L, WIRTH R, et al. Nanostructure, composition and mechanisms of bivalve shell growth[J]. Geochimicaet Cpsmochimica Acta, 2008, 72(22): 5401-5415. doi: 10.1016/j.gca.2008.08.019
[24] 王慧, 封余军, 郑碧华, 等. 等离子体发射光谱分析珍珠贝贝壳结晶的微量元素[J]. 湖北农学院学报, 1998, 18(2): 142-144. https://www.cqvip.com/qk/91582X/199802/3080694.html [25] 夏静芬, 钱国英, 陈亮, 等. 珍珠粉和贝壳粉的化学成分和结构特征分析[J]. 化学研究与应用, 2010, 22(11): 1467-1471. doi: 10.3969/j.issn.1004-1656.2010.11.025 [26] 孙维萍, 潘建明, 刘小涯, 等. 浙江沿海贝类体内重金属元素含量水平与评价[J]. 海洋学研究, 2010, 28(4): 43-49. doi: 10.3969/j.issn.1001-909X.2010.04.006 [27] 逯云召. 金属离子对马氏珠母贝珍珠层颜色和金属硫蛋白基因表达的影响[D]. 湛江: 广东海洋大学, 2012. [28] MCGINTY E L, EVANS B S, TAYLOR T J, et al. Xenografts and pearl product ion in two pearl oyster species, P. maxima and P. margaritifera: effect on pearl quality and a key to understanding genetic contribution[J]. Aquaculture, 2010, 302(3/4): 175-181.
[29] 张根芳, 许式见, 方爱萍. 组织小片对三角帆蚌外套膜无核珍珠颜色成因的影响[J]. 水生生物学报, 2013, 37(3): 581-587. doi: 10.7541/2013.66 [30] 郝之奎, 王嫣, 顾志峰, 等. 珍珠颜色的研究进展[J]. 安徽农业科学, 2007, 35(11): 3260-3261. doi: 10.3969/j.issn.0517-6611.2007.11.061 [31] 刘娟花, 马孝甜, 苗东亮, 等. 珍珠致色因素研究概况[J]. 江苏农业科学, 2011(1): 289-292. doi: 10.3969/j.issn.1002-1302.2011.01.105 [32] 高珊. 广西马氏贝珍珠颜色特征研究和染色实验[D]. 北京: 中国地质大学, 2013. [33] 严俊, 邓小琼, 胡丹静, 等. 淡水养殖珍珠棱柱层的微结构形貌特征[J]. 南方水产科学, 2013, 9(1): 48-52. doi: 10.3969/j.issn.2095-0780.2013.01.008 -
期刊类型引用(12)
1. 王伦,王崇懿,刘建勇,傅学丽. 凡纳滨对虾多代选育群体生长和耐综合胁迫性状的配合力及杂种优势分析. 海洋学报. 2022(04): 65-73 .  百度学术
百度学术
2. 李金龙,刘越,车宗豪,田园,刘杰,闫喜武,霍忠明. 菲律宾蛤仔中国莱州群体与朝鲜新义州群体杂交子代早期生长发育. 大连海洋大学学报. 2020(02): 190-196 . 百度学术
3. 孙静,陈明强,魏海军,邓正华,李有宁,金朝焕,王继金,王雨,石承开,黄超旬. 不同地理群体合浦珠母贝双列杂交子代数量性状比较及其相关和通径分析. 南方农业学报. 2020(06): 1451-1461 . 百度学术
4. 栗志民,钱佳慧,刘建勇,艾加林,檀克勤. 九孔鲍α-淀粉酶基因克隆、表达分析及与生长性状相关性. 南方水产科学. 2017(06): 14-21 . 本站查看
5. 郑言鑫,赵春暖,于涛,蔡忠强,林建国,秦搏. 4个不同地理群体刺参杂交的杂种优势及配合力分析. 上海海洋大学学报. 2017(01): 57-63 . 百度学术
6. 江宗冰,戴习林,明磊,蒋飞,王海洋,袁新程,李玉锋,丁福江. 罗氏沼虾生长性状的种内杂交优势及遗传力与遗传相关分析. 上海海洋大学学报. 2017(02): 189-196 . 百度学术
7. 杨耀宗,卞友庆,姚新年,钱晓燕,姚颖,曹正锋,顾天天,徐琪,陈国宏. 鹅不同杂交组合生长性能比较及杂种优势分析. 中国家禽. 2017(16): 53-56 . 百度学术
8. 张伟杰,韩奋杰,陈顺,刘奎,周秘,王宝锋,王中,刘明泰,湛垚垚,经晨晨,常亚青. 中间球海胆与马粪海胆杂交家系早期生长比较和亲本配合力估计. 中国水产科学. 2017(05): 1035-1044 . 百度学术
9. 王佩佩,丁严冬,于兴达,贾秀琪,张国松,尹绍武,陈树桥,周国勤. 河川沙塘鳢3个不同地理群体杂交F_1代生长性能及遗传多样性. 水产科学. 2016(05): 528-534 . 百度学术
10. 胡志国,刘建勇,袁瑞鹏,张嘉晨. 3个凡纳滨对虾引进群体对温度和盐度耐受力的配合力分析. 海洋科学. 2016(01): 25-31 . 百度学术
11. 余勇,李琪,于红,孔令锋,于瑞海. 长牡蛎中国群体和日本群体杂交子代的杂种优势分析. 中国海洋大学学报(自然科学版). 2016(02): 35-41 . 百度学术
12. 李艳红,李宁,蒋丽,谷伟,张晓慧,杨润清,王炳谦. 虹鳟完全双列杂交试验生长性状遗传分析. 中国水产科学. 2015(06): 1095-1104 . 百度学术
其他类型引用(10)
计量
- 文章访问数: 3319
- HTML全文浏览量: 146
- PDF下载量: 2000
- 被引次数: 22
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
