
Characteristics of fish community structure and its relationship with environmental factors in marine ranching zone in southern area of Yintan in Guangxi
-
摘要:
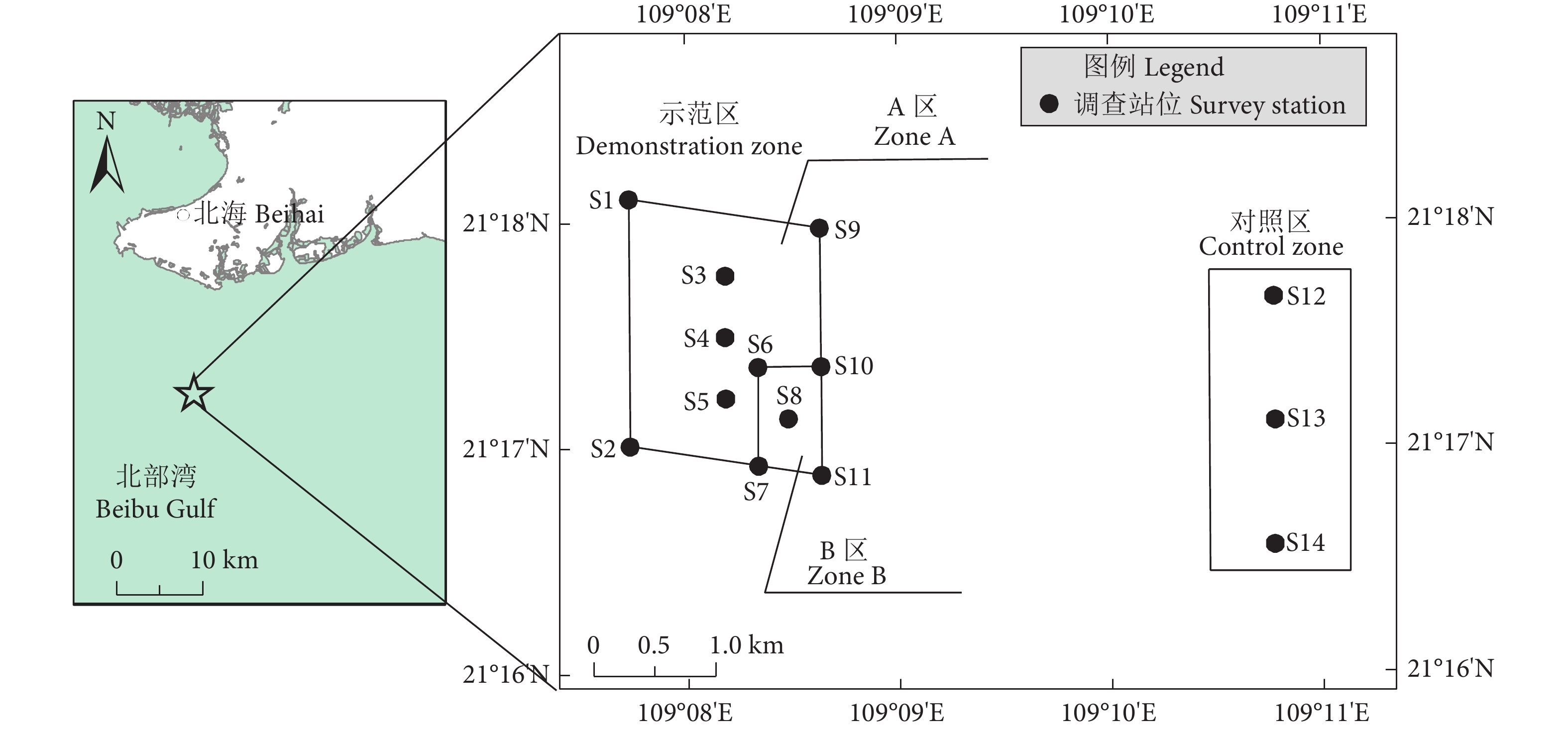
适时评价国家级海洋牧场示范区的鱼类资源状况,对推进海洋牧场建设具有重要意义。基于2022年4月广西银滩南部海域海洋牧场渔业资源调查数据,分析了鱼类资源特征、群落结构及其与环境因子的相关性,以为北部湾海洋牧场生态效应研究提供参考。结果显示,示范区鱼类数量密度、生物量密度和种类数均高于对照区。示范区和对照区鱼类物种组成在种级水平上为中等不相似,且2个区域鱼类群落均受到一定程度的干扰。研究区鱼类群落可划分为4个具有显著性差异的类群 (p<0.05),分别位于示范区B区和对照区南部站位 (类群I)、对照区北部站位 (类群II)、示范区A区和B区交界处 (类群III) 和示范区A区 (类群IV)。类群I和III的平均相异性指数为53.88%,贡献率排前3位的分歧种为吉打副叶鲹 (Alepes djedaba)、蓝圆鲹 (Decapterus maruadsi)和竹荚鱼 (Trachurus japonicus);类群III和IV的平均相异性指数为48.85%,贡献率排前3位的分歧种为竹荚鱼、短吻鲾 (Leiognathus brevirostris) 和花斑蛇鲻 (Saurida undosquamis)。水温和化学需氧量是显著影响研究区鱼类群落结构的主要环境因子,溶解氧和浮游植物密度的影响稍弱。
Abstract:Timely assessment of fish resources in state-level marine ranching demonstration zone is important in promoting the construction of marine ranching. Based on the fishery resources survey data in the southern area of Guangxi Yintan in April 2022, we analyzed the characteristics of fish resources, community stability, fish community structure and their correlation with environmental factors to provide references for research on the ecological effects of marine ranching in Beibu Gulf. The results show that the fish individual density, biomass density and number of species in the demonstration zone were higher than those in the control zone. The fish species composition in the demonstration and control zones was moderately dissimilar at species level. In addition, fish communities in both zones were somewhat disturbed. The fish community in the survey area was divided into four significantly different clusters (p<0.05), which were located in the southern station of the demonstration zone B and the control zone (Cluster I), the northern station of the control zone (Cluster II), the junction of the demonstration zone A and B (Cluster III), and the demonstration zone A (Cluster IV). The average dissimilarity index of Cluster I and III was 53.88%, and the divergent species that ranked in the top three in terms of their contribution were Alepes djedaba, Decapterus marauds and Trachurus japonicus, while the average dissimilarity index of Cluster III and IV was 48.85%, with the top three divergent species (T. japonicus, Leiognathus brevirostri and Saurida undosquamis) contributing to the dissimilarity index. Water temperature and chemical oxygen demand were the main environmental factors significantly influencing the structure of the fish community in the survey area, while dissolved oxygen and phytoplankton density had a slightly weaker effect.
-
Keywords:
- Marine ranching /
- Fish community structure /
- Environmental factor /
- Yintan in Guangxi
-
氨氮 (NH4 +-N) 和亚硝酸盐 (NO2 −-N) 等无机氮是水产养殖主要的氮素污染物,对养殖动物具有较强的毒性,研究显示氨氮能明显抑制养殖动物的生理和免疫系统,而亚硝酸根离子 (NO2 −) 能够抑制血液载氧能力,通常在较低的浓度下危害水生动物健康,甚至导致死亡[1-5]。随着养殖密度不断提高,水体氨氮和亚硝酸盐浓度的快速累积造成了潜在危险。传统方法依靠大量换水降低氨氮和亚硝酸盐的浓度,会给周边环境造成污染,引起水体富营养化。近年来,随着国家对环境保护要求日趋严格,直排直灌的养殖方式已经不可持续,如何环保、高效地去除水体有害氮素是促进水产养殖成功的关键。
微生物在生态链中扮演分解者角色,是物质和能量循环的重要组成,利用有益微生物调控养殖水质是近年的研究热点[6-8]。胡晓娟等[9]发现菌株NB5对水体氨氮具有显著的去除效果,信艳杰等[10]利用光合细菌吸收水体氮 (N)、磷 (P)。不同的细菌具有不同的生态功能,在循环养殖系统中多用硝化细菌来消除氨氮和亚硝酸盐,但自养硝化细菌生长缓慢,且容易受环境影响,目前还未在水产养殖中大范围推广应用[11-12]。红球菌作为一类重要的模式微生物,在自然环境中分布广泛且菌株种类多样。由于红球菌具有降解有机污染物、生物转化和降解毒素等作用,在环境治理、生物合成与转化以及医药领域具有潜在的应用价值,其中赤红球菌 (Rhodococcus ruber) 生长快且易于培养,具有消除环境中氨氮和亚硝酸盐的潜力[13-14]。本实验室前期筛选到一株赤红球菌HDRR2Y,初步研究发现其可去除水体氨氮和亚硝酸盐[15]。细菌的功能与环境因素往往密切相关,为了更好地应用于水产养殖生产中,本文研究了赤红球菌HDRR2Y发酵液去除氨氮和亚硝酸盐的效果,并探讨了细菌浓度、温度、底物浓度、转速和盐度等5种常见的环境因子对HDRR2Y发酵液去除氨氮和亚硝酸盐功能的影响,以期为菌株HDRR2Y在水产养殖中的应用提供理论依据。
1. 材料与方法
1.1 菌株来源
赤红球菌HDRR2Y由中国水产科学研究院南海水产研究所提供,该菌种保藏于中国典型培养物保藏中心 (CCTCC NO: M2019009),已申请发明专利 (申请号:202010159584.2)。
1.2 菌株发酵
采用德国贝朗生物系统公司Biostat B型体积为2 L的发酵罐进行发酵培养,培养基为(g·L−1):三水合醋酸钠 [C2H3NaO2·3(H2O)] 3.0,酵母膏1.8,硫酸镁 (MgSO4) 0.2,磷酸二氢钾 (KH2PO4) 0.5,氯化钙 (CaCl2) 0.5,氯化钠 (NaCl) 9.0,硫酸锰 (MnSO4) 0.025,硫酸铁 (FeSO4) 0.05,谷氨酸 (C5H9NO4) 0.002。发酵条件为:29 ℃,36 h,pH 7.0,装液量1.2 L (60%),初始接种菌量1%,通气量3 L·min−1,搅拌转速400 r·min−1。
1.3 发酵过程中菌株浓度测定与形态观察
在菌株发酵第0、第18、第24、第30、第36小时通过50 mL注射器针筒以抽气、排气的方式获取发酵液,取样口在取样前后需用酒精灯灼烧,非取样时间用无菌锡箔纸包裹取样口。通过梯度稀释法将发酵原液制成十倍系列稀释匀液,取3个最适稀释度,每个稀释浓度设置3个平行组,然后进行平板涂布,置于培养箱内,在30 ℃、湿度90%的条件下培养24 h,待菌落形成后进行菌落计数。同时通过显微镜将菌株细胞放大1 000倍进行观察,记录下不同时间点采集的发酵液中细菌的细胞形态。
1.4 菌株培养液去除氨氮和亚硝酸盐效果测定
菌株发酵完成后,在无菌条件下取适量培养液加入对虾养殖池塘水 (来自陆丰某养殖场),至水体赤红球菌初始浓度为5×104 CFU·mL−1 (加菌组,GH),水体总氨氮和亚硝酸盐的初始质量浓度均为15 mg·L−1,实验条件为水温30 ℃、溶解氧质量浓度5 mg·L−1、pH 8.0,在实验第0、第3和第7天取适量水体,分别依照GB 17378.4—2007中靛酚蓝分光光度法和盐酸萘乙二胺分光光度法测定水体氨氮和亚硝酸盐浓度。对照组取相同体积的池塘水,不加入赤红球菌,其他操作与GH组一致 (对照组,GC)。
1.5 5种环境因子对赤红球菌去除底物效果的影响
取赤红球菌培养液加入对虾养殖水体中,然后按照温度、转速、盐度、接种菌量、底物(氨氮和亚硝酸盐) 浓度等5种因子进行Plackett-Burman实验设计。实验条件为溶解氧质量浓度5 mg·L−1、pH 8.0,其他条件依据表1和表2,实验周期为7 d,7 d后检测氨氮和亚硝酸盐浓度,并计算两者的去除率。
表 1 Plackett-Burman实验因素及与水平设计Table 1. Factors and levels design of Plackett-Burman因素 Factor 水平 Level −1 1 A:菌浓度Bacterial concentration/(CFU·mL−1) 104 107 B:底物 (氨氮/亚硝酸盐) 质量浓度Substrate concentration (Ammonia nitrogen/Nitrite)/(mg·L−1) 15 30 C:盐度 Salinity/‰ 15 30 D:转速 Rotating speed/(r·min−1) 150 225 E:温度 Temperature/℃ 20 30 表 2 Plackett-Burman设计Table 2. Plackett-Burman experimental design序号 No. A:菌浓度Bacterial concentration B:底物 (氨氮/亚硝酸盐) 浓度Substrateconcentration (Ammonia nitrogen/Nitrite) C:盐度Salinity D:转速Rotating speed E:温度Temperature 1 1 −1 −1 −1 1 2 1 −1 1 1 1 3 1 −1 1 1 −1 4 −1 1 −1 1 1 5 −1 1 1 −1 1 6 −1 −1 −1 1 −1 7 −1 −1 1 −1 1 8 1 1 −1 −1 −1 9 −1 −1 −1 −1 −1 10 −1 1 1 1 −1 11 1 1 −1 1 1 12 1 1 1 −1 −1 1.6 数据统计与分析
底物浓度去除率的计算公式为:R=(C0− Ct)/ C0×100%。式中:R为去除率;C0为初始浓度;Ct实验后的浓度。采用Design-Expert 11软件对实验进行设计与分析,以OriginPro 2016软件作图,通过SPSS 20.0软件进行单因素方差分析 (One-way ANOVA) ,比较各组数据的差异显著性,显著水平设定为P<0.05。
2. 结果
2.1 赤红球菌发酵过程中生长情况
如图1所示,在0~24 h菌株HDRR2Y数量快速增加,为生长对数期;在24~36 h菌株生长逐渐进入稳定期。发酵结束后,菌浓度达4.08×109 CFU·mL−1。
2.2 菌株形态变化情况
菌株HDRR2Y在发酵第0、第18、第24、第30、第36小时的细胞形态变化见图2。在第0小时,菌株细胞呈球状或短杆状,多由1或2个细胞构成;第18小时,菌株细胞变长呈弯曲杆状,处于分裂增殖期;第24小时,菌株细胞多为弯曲杆状,进一步增殖;第30小时,菌株长细胞分裂为短细胞,呈弯曲杆状;第36小时,菌株细胞进一步变短,开始出现球状 (图2)。
2.3 菌株HDRR2Y发酵液对氨氮和亚硝酸盐的去除效果
如图3所示,添加菌株HDRR2Y发酵液后第3天,养殖水体的氨氮和亚硝酸盐浓度明显降低,氨氮质量浓度从第0天的15 mg·L−1降至5.56 mg·L−1,去除率为62.96%;亚硝酸盐质量浓度从15 mg·L−1降至6.95 mg·L−1,去除率为59.37%。第7天,水体氨氮和亚硝酸盐浓度略有上升,但与第3天比较无显著性差异 (P>0.5)。实验过程中,对照组水体的氨氮和亚硝酸盐浓度基本保持不变,与添加菌组相比较差异极显著 (P<0.01) (表3)。
![]() 图 3 赤红球菌HDRR2Y对养殖水体氨氮和亚硝酸盐作用效果Figure 3. Effect of HDRR2Y on ammonia and nitrite nitrogen in aquaculture water表 3 养殖水体中氨氮和亚硝酸盐去除率Table 3. Removal rate of ammonia nitrogen and nitrite in aquaculture water
图 3 赤红球菌HDRR2Y对养殖水体氨氮和亚硝酸盐作用效果Figure 3. Effect of HDRR2Y on ammonia and nitrite nitrogen in aquaculture water表 3 养殖水体中氨氮和亚硝酸盐去除率Table 3. Removal rate of ammonia nitrogen and nitrite in aquaculture water被测指标Measured index 时间Time 去除率 Removal rate/% 对照组 GC 加菌组 GH 氨氮 NH4 +-N 第3天 −5.16±6.87a 62.92±1.53b 第7天 −4.06±7.31a 57.50±2.16b 亚硝酸盐 NO2 −-N 第3天 −1.43±2.10a 59.37±2.49b 第7天 −4.92±1.41a 56.36±1.67b 注:同行数据的不同字母代表对照组和加菌组差异显著 (P<0.05)。 Note: Values with different letters within the same column indicate significant difference (P<0.05). 2.4 5种因素对赤红球菌去除氨氮和亚硝酸效果的影响
Plackett-Burman实验因素水平与回归方程的方差分析结果显示,5种因素的回归方程为,氨氮:Y1=45.74+1.11A+3.43B −0.075C+1.84D+ 5.74E。式中:Y1表示氨氮去除率;A表示菌浓度;B为氨氮质量浓度;C为盐度;D为转速;E为温度。该模型的显著性达到极显著水平 (P<0.001),且判定系数R2为0.893 7 (R2>0.8),说明该模型可信度高,拟合度可靠;亚硝酸盐:Y2=45.42+0.77A+0.82B−0.97C+3.57D+3.67E。式中:Y2表示亚硝酸盐去除率;A表示菌浓度;B为亚硝酸盐质量浓度;C为盐度;D为转速;E为温度。此模型的可信度同样较高 (P<0.01),拟合度可靠 (R2=0.805 3>0.8)。结果显示赤红球菌HDRR2Y能显著去除水体中的氨氮 (P<0.05),最高去除率为59.3%,氨氮质量浓度从30.00 mg·L−1降至12.21 mg·L−1 (表4)。5种因子中,温度、氨氮浓度可显著影响赤红球菌HDRR2Y去除氨氮效果 (P<0.05),转速、菌量和盐度对赤红球菌HDRR2Y去除氨氮效果的影响不显著 (P>0.05),对权重的影响程度依次为:温度>氨氮浓度>转速>菌量>盐度 (表5)。
表 4 Plackett-Burman实验底物去除率Table 4. Removal rate of substrate in Plackett-Burman experiment序号No. Y1:氨氮去除率Removal rate of ammonia nitrogen/% Y2:亚硝酸盐去除率 Removal rate of nitrite/% 1 48.4 42.5 2 51.3 36.6 3 36.2 40.2 4 52.4 47.1 5 53.6 36.8 6 42.5 42.9 7 43.9 39.9 8 40.7 32.5 9 31.6 43.7 10 43.8 38.7 11 59.3 53.4 12 45.2 30.7 表 5 Plackett-Burman实验因素水平与回归方程方差分析 (底物为氨氮)Table 5. Plackett-Burman's experimental factor level and ANOVA of regression equation (Substrate is ammonia nitrogen)因素Factor 平方和Sum of squares 自由度df 均方Mean square F P>F 显著性Significance 重要性排序Importance ranking 模型 Model 591.88 5 118.38 10.09 0.007 0 ** A:菌浓度Bacterial concentration/(CFU·mL−1) 14.74 1 14.74 1.26 0.305 2 4 B:氨氮质量浓度 Ammonia nitrogen mass concentration/ (mg·L−1) 140.77 1 140.77 12.00 0.013 4 * 2 C:盐度 Salinity/‰ 0.0675 1 0.067 5 0.005 8 0.942 0 5 D:转速 Rotating speed/(r·min−1) 40.70 1 40.70 3.47 0.111 9 3 E:温度 Temperature/℃ 395.60 1 395.60 33.71 0.001 1 * 1 残留误差 Residual error 70.41 6 11.74 总差 Total difference 662.29 11 注:判定系数R2=0.893 7;*. 差异显著 (P<0.05);**. 差异极显著( P<0.001)。 Note: Determination coefficient (R2)= 0.893 7; *. Significant difference (P<0.05); **. Very significant difference (P<0.001). 赤红球菌HDRR2Y同样能显著去除水体中的亚硝酸盐 (P<0.05),最高去除率为53.4%,亚硝酸盐质量浓度从30.00 mg·L−1降至13.98 mg·L−1 (表4)。在养殖水体中,5种因子影响赤红球菌HDRR2Y去除亚硝酸盐效果的权重程度依次为:温度>转速>盐度>亚硝酸盐浓度>菌量。其中,温度和转速显著影响赤红球菌HDRR2Y对亚硝酸盐的去除效果 (P<0.05),亚硝酸盐浓度、菌量和盐度对赤红球菌HDRR2Y去除亚硝酸盐效果的影响不显著 (P>0.05) (表6)。
表 6 Plackett-Burman实验因素水平与回归方程方差分析 (底物为亚硝酸盐)Table 6. Plackett-Burman's experimental factor level and ANOVA of regression equation (Substrate is nitrite)因素Factor 平方和Sum of squares 自由度df 均方Mean square F P>F 显著性Significance 重要性排序Importance ranking 模型 Model 340.26 5 68.05 4.96 0.038 2 * A:菌浓度 Bacterial concentration/(CFU·mL−1) 7.05 1 7.05 0.514 5 0.500 2 5 B:氨氮质量浓度Ammonia nitrogen mass concentration/(mg·L−1) 8.00 1 8.00 0.583 8 0.473 8 4 C:盐度 Salinity/‰ 11.21 1 11.21 0.817 9 0.400 6 3 D:转速 Rotating speed/(r·min−1) 152.65 1 152.65 11.13 0.015 7 * 2 E:温度 Temperature/℃ 161.33 1 161.33 11.77 0.014 0 * 1 残留误差 Residual error 82.26 6 13.71 总差 Total difference 422.52 11 注:判定系数R2=0.8053;*. 差异显著 (P<0.05)。 Note: Determination coefficient (R2)= 0.8053; *. Significant difference (P<0.05). 3. 讨论
赤红球菌属于化能异养细菌,需通过氧化有机物获得生长的能量和碳源,可利用无机氮或有机氮作为氮源。本研究发现,采用前期响应面优化的发酵培养参数,在以三水合醋酸钠为碳源、酵母膏为氮源的培养基中能快速生长,培养液细菌浓度在24 h内从104 CFU·mL−1增至109 CFU·mL−1,之后快速进入稳定期,发酵效果良好。与简单的摇瓶培养相比,发酵罐培养中的通气、控制pH能使微生物生长代谢更加旺盛[16-18]。发酵培养中持续的通气能使发酵液中的溶解氧一直处于较高水平,本研究中的赤红球菌HDRR2Y为好氧细菌,在氧气充足的前提下能快速分裂增殖,从而提高菌量。从细菌的形态同样可以观察到,第18和第24小时的细胞形态绝大多数呈长条形,是细胞分裂增殖期的特点,第30小时之后的细胞又逐渐回归至球形或短杆形,说明细菌生长已经处于稳定期。细菌的细胞形态是细菌种类鉴定及活力评判的重要指标之一,如葡萄球菌 (Staphyloccocus) 一般呈球形,乳酸菌 (Lactobacillus) 多呈短杆状。此外,细胞形态一定程度上反映了细菌的生长状态,毛青钟和胡金凤[19]发现细菌形态不正常的发酵醪其酸度升高较快,容易引起发酵醪的酸败,周丽英等[20]发现假单胞菌 (Pseudomonas) 在质量浓度为100 mg·L−1的镉 (Cd) 环境中,细胞表面出现丝状物及不同程度的凹陷,推测可能与重金属离子所起的胁迫密切相关。
赤红球菌喜欢生长在有机质丰富的环境中,目前多应用于生活污水或工业废水的生物处理[21-23]。陈静和龚艳华[14]应用赤红球菌处理污水、废水,可有效去除水体氨氮。罗固源等[13]发现一株赤红球菌在好氧的条件下可去除废水中的亚硝酸盐。本研究中,将赤红球菌HDRR2Y发酵液添加至富含氨氮和亚硝酸盐的养殖水体后,第3天氨氮和亚硝酸盐浓度去除率约达60%,去除效果显著 (P<0.05),但第7天的氨氮、亚硝酸盐浓度比第3天略有上升,推测可能是水体营养不足,限制了赤红球菌的生长。
温度能够显著影响赤红球菌HDRR2Y降解氨氮和亚硝酸盐效果,主要是因为温度影响微生物的生长代谢,从而影响微生物对外界物质的吸收效率。高温使细菌细胞内蛋白质变性、酶失活,使细菌灭亡,低温会抑制微生物生长,微生物体内酶的活性在最适温度下可提高1~2倍[24]。多数研究显示,硝化菌在约30 ℃时硝化功能得到最大发挥,而在20 ℃及以下时硝化作用受到极大抑制[25-26]。本研究显示在30 ℃时,菌株HDRR2Y对氨氮和亚硝酸盐降解效果最好,与多数研究结果相符。因此菌株HDRR2Y适合在高温养殖季使用。
底物浓度是影响微生物代谢活性的重要因子,底物浓度越高,微生物代谢活性越高。本研究发现,底物亚硝酸盐的浓度不是影响菌株降解亚硝酸盐效果的显著因素,而底物氨氮的浓度则显著影响菌株降解氨氮的效果,说明菌株HDRR2Y更易利用氨氮。王晓明和王杰[27]发现随着进水氨氮浓度的不断升高,硝化系统对氨氮的去除率随之升高,但过高的氨氮浓度会抑制硝化反应效率[28]。本研究结果表明,菌株HDRR2Y更适合用于高浓度氨氮的去除。
转速是另一个影响赤红球菌去除氨氮和亚硝酸盐效果的重要因子。本研究发现,较高的转速有助于赤红球菌HDRR2Y去除氨氮和亚硝酸盐,且转速对于赤红球菌代谢亚硝酸盐的影响大于氨氮代谢。已有研究发现水体流速可显著影响硝化及反硝化的效率[29-30],搅动的主要作用是让微生物与水体中的氨氮和亚硝酸盐更充分的接触,同时还能增加水体的溶解氧,通常在一定范围内,转速与溶解氧浓度成正比[31-32]。赤红球菌是好氧细菌,氧气不足时代谢会受到抑制,氧气充足时,新陈代谢速度达到最快,底物利用率达到最高。刘玉沛等[33]研究赤红球菌对苯酚的降解效果时,氧气充足条件下降解率为61.87%,而氧气不足条件下降解率显著降低 (50.44%)。因此,在养殖过程中使用赤红球菌,要充分利用水车等增氧设备,提高水体流速和溶解氧浓度,发挥菌株最大的功效。
微生物细胞浓度与其生态功能密切相关,利用自养硝化细菌去除环境中的氨氮、亚硝酸盐往往起效时间较长,因为硝化细菌生长缓慢,一般需要富集5~8周才能发挥明显的硝化效应[34-35]。本研究结果显示,赤红球菌的浓度对氨氮和亚硝酸盐的去除无显著影响,可能是由于低浓度组的赤红球菌浓度已经达到发挥效应的水平。此外,赤红球菌属于异养菌,在适宜的环境中能迅速生长,这也是异养硝化细菌的优势。水体盐度也会影响硝化反应,目前多数研究认为,盐分对硝化反应存在抑制作用,且对亚硝酸盐氧化反应的抑制要大于对好氧氨氧化反应的抑制作用[36-37]。但经过盐分驯化培养后,在较高的盐分浓度下仍然维持较好的硝化效果[38]。本研究发现在盐度为15‰和30‰时,赤红球菌去除氨氮和亚硝酸盐的效率并未受到太大的影响,这可能与该株菌自身广盐性有关[15]。因此,该株菌在高盐度或低盐度的水域中均具有推广使用的潜力,拥有广泛的应用前景。
4. 结论
赤红球菌HDRR2Y在装液量60% (V/V)、29 ℃、pH 7.0、机械搅拌转速400 r·min−1、接种量1%、通气量3 L·min−1的条件下培养36 h,发酵完成后的其质量浓度达4.08×109 CFU·mL−1。发酵后的菌株可在72 h内显著降低养殖水体中的氨氮和亚硝酸盐,说明该菌株具有作为优良渔业微生物菌剂的应用前景。此外,温度、底物浓度及水体流速是对菌株代谢氨氮和亚硝酸盐效率影响最显著的3个环境因子,因此在实际应用当中需要注意水体的温度和流速,以确保菌株使用效果的最大化。
-
![]()
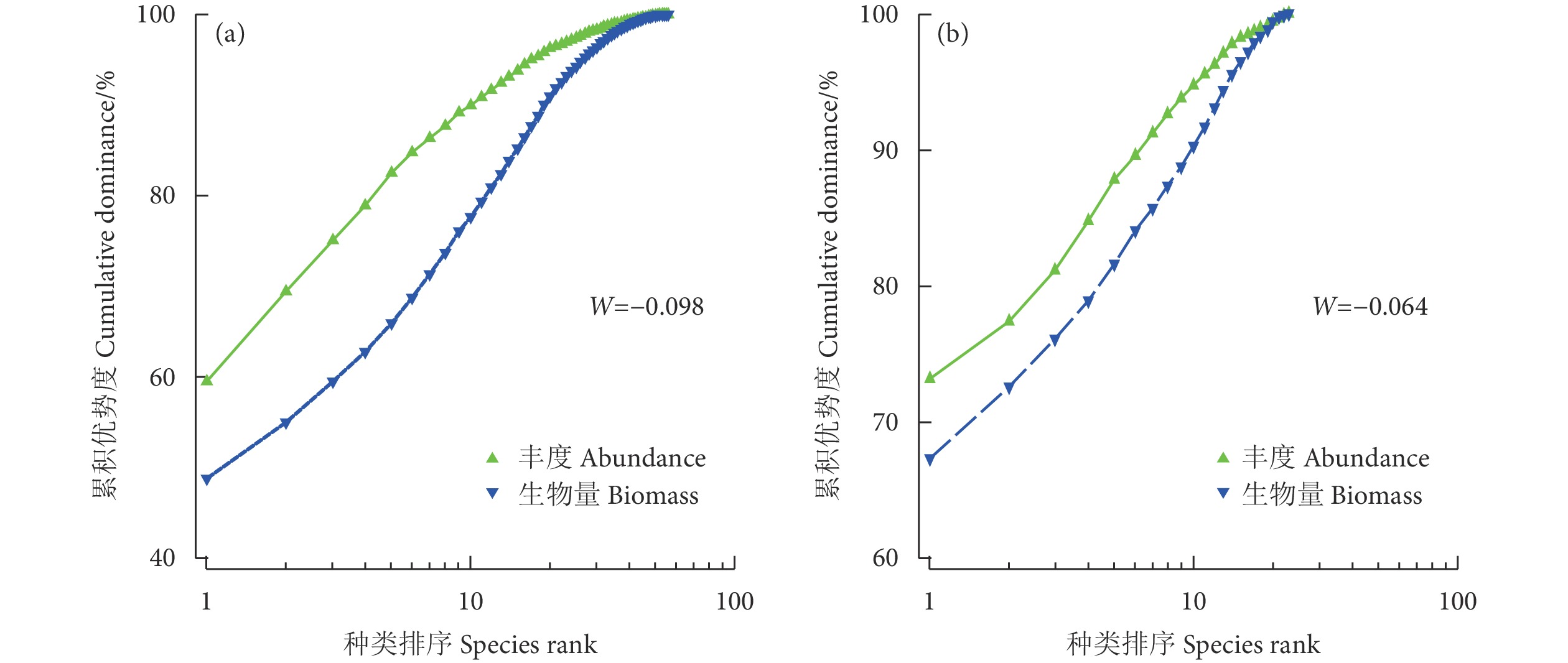
图 2 示范区 (a) 和对照区 (b) 鱼类群落丰度-生物量比较曲线
Figure 2. Abundance biomass comparison curve of fish community in demonstration zone (a) and control zone (b)
![]()
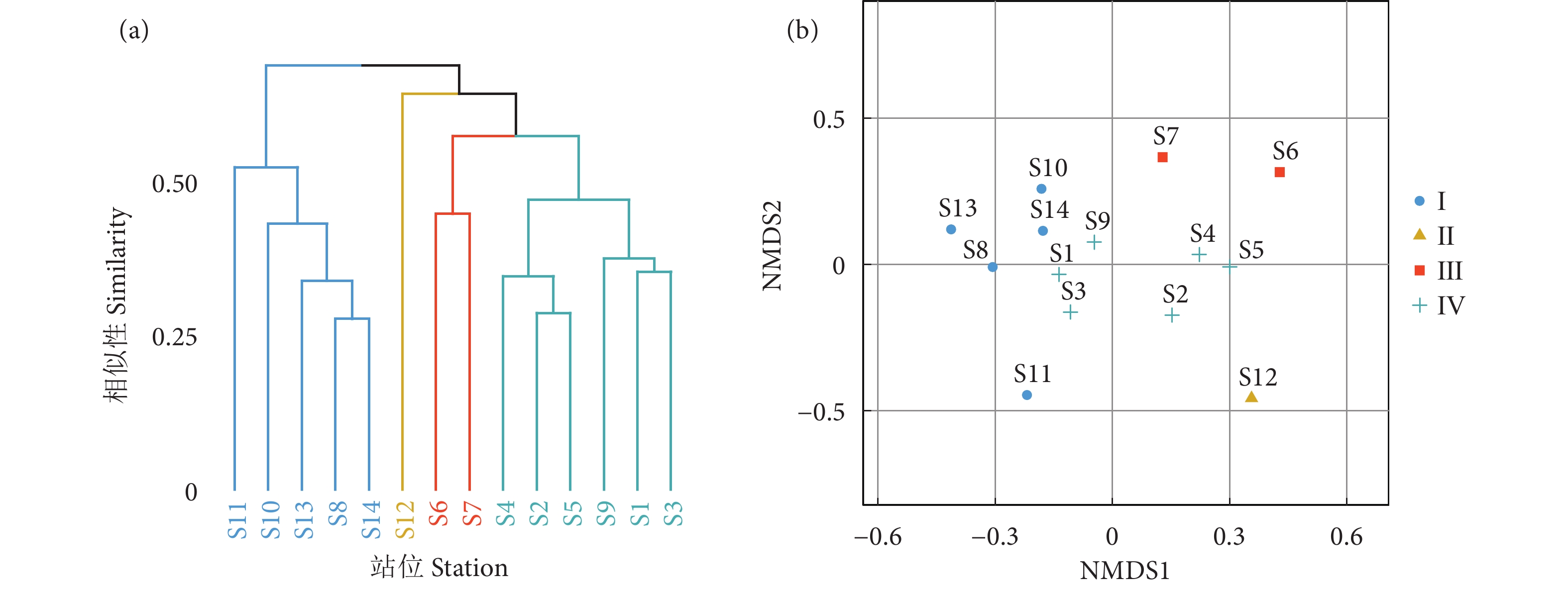
图 3 银滩南示范区鱼类聚类分析 (a) 和非度量多维标度分析 (b)
Figure 3. Cluster analysis (a) and NMDS (b) of fish community in southern station of demonstration zone in Yintan
![]()
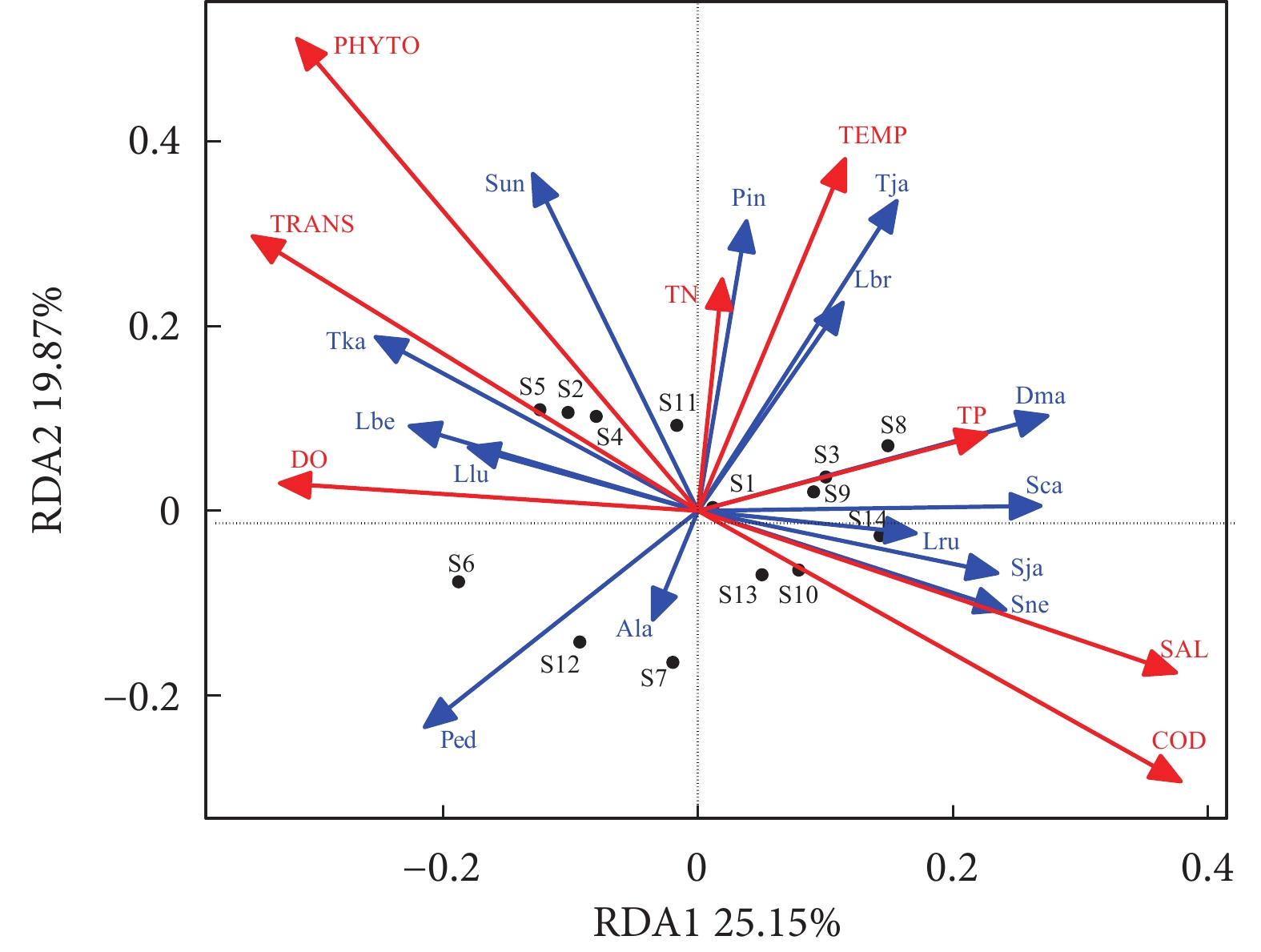
图 4 银滩南示范区主要鱼种与环境因子的RDA分析
注:TEMP. 水温;SAL. 盐度;TRANS. 透明度;DO. 溶解氧;COD. 化学需氧量;TN. 总氮;TP. 总磷;PHYTO. 浮游植物密度;Ala. 侧带天竺鲷;Dma. 蓝圆鲹;Lbe. 细纹鲾;Lbr. 短吻鲾;Llu. 月尾兔头鲀;Lru. 鹿斑仰口鲾;Ped. 二长棘鲷;Pin. 鲬;Sca. 黄斑篮子鱼;Sja. 少鳞鱚;Sne. 魔拟鲉;Sun. 花斑蛇鲻;Tja. 竹荚鱼;Tka. 赤鼻棱鳀。
Figure 4. Redundancy analysis between fish species and environmental factors in southern station of demonstration zone in Yintan
Note: TEMP. Temperature; SAL. Salinity; TRANS. Transparency; DO. Dissolved oxygen; COD. Chemical oxygen demand; TN. Total nitrogen; TP. Total phosphate; PHYTO. Phytoplankton density; Al. A. lateralis; Dma. D. maruadsi; Lbe. L. berbis; Lbr. L. brevirostris; Llu. L. lunaris; Lru. L. ruconius; Ped. P. edita; Pin. P. indicus; Sca. S. canaliculatus; Sja. S. japonica; Sne. S. neglecta; Sun. S. undosquamis; Tja: T. japonicas; Tka. T. kammalensis.
表 1 示范区和对照区环境因子和渔获参数的均值、最小值和最大值
Table 1 Mean, minimum and maximum values of environmental factor and catch parameters in demonstration area and control area
环境因子
Environment factor示范区
Demonstration zone对照区
Control zone显著性
Significance均值 (最小值~最大值)
Mean (Minimum–Maximum)均值 (最小值~最大值)
Mean (Minimum–Maximum)水深 Water depth/m 11.69 (10.70~13.00) 11.87 (11.20~12.70) n.s. 透明度 Transparency/m 3.76 (3.50~4.20) 3.83 (3.80~3.90) n.s. 水温 Water temperature/℃ 21.33 (20.90~21.80) 20.47 (20.40~20.60) ** 盐度Salinity/‰ 32.34 (32.09~32.56) 32.35 (32.30~32.41) n.s. 溶解氧Dissolved oxygen/(mg·L−1) 8.52 (8.45~8.62) 8.29 (8.10~8.50) * 化学需氧量Chemical oxygen demand/(mg·L−1) 1.69 (0.60~3.00) 2.85 (2.68~2.96) * 总磷Total phosphate/(mmol·L−1) 0.67 (0.27~3.31) 0.42 (0.35~0.49) n.s. 总氮Total nitrogen/(mmol·L−1) 38.64 (32.00~40.40) 41.40 (39.00~43.60) n.s. 浮游植物密度Phytoplankton density/(103 个·m−3) 595 (250~977) 241 (215~266) * 数量密度 Individual density/(尾·km−2) 19 137 (14 839~32 997) 16 852 (14 439~20 038) n.s. 生物量密度Biomass density/(kg·km−2) 1 897 (1 504~2 630) 1 244 (1 006~1 493) * 种类数Number of species 16 (11~21) 12 (11~13) * 注:根据Mann-Whitney U检验,n.s.. 无显著性差异;*. p<0.05;**. p<0.01。 Note: According to Mann-Whitney U test: n.s.. Insignificant difference; *. p<0.05; **. p<0.01.  下载: 导出CSV
下载: 导出CSV
表 2 鱼类不同类群组间差异和主要分歧种及其贡献率 (>4%)
Table 2 Differences in community structure and contribution rates of different species (>4%)
类群
Cluster平均相异性
Average dissimilarity/%种类
Species数量密度变化
Change in individual density贡献率
Contribution rate/%类群I & III
Cluster I & III53.88 吉打副叶鲹 Alepes djedaba 增加 6.8 蓝圆鲹 Decapterus maruadsi 减少 6.1 竹荚鱼 Trachurus japonicus 减少 5.6 魔拟鲉 Scorpaenopsis neglecta 增加 5.5 黄斑篮子鱼 Siganus canaliculatus 减少 5.2 短吻鲾 Leiognathus brevirostris 减少 4.9 鹿斑仰口鲾 Secutor ruconius 增加 4.4 纤羊舌鲆 Arnoglossus tenuis 增加 4.4 细纹鲾 Leiognathus berbis 增加 4.3 毒拟鲉 Scorpaenopsis diabolus 减少 4.3 侧带天竺鲷 Apogon lateralis 增加 4.1 斑鰶 Konosirus punctatus 减少 4.0 类群III & IV
Cluster III & IV48.85 竹荚鱼 Trachurus japonicus 增加 8.7 短吻鲾 Leiognathus brevirostris 增加 6.6 花斑蛇鲻 Saurida undosquamis 增加 5.8 蓝圆鲹 Decapterus maruadsi 增加 5.4 魔拟鲉 Scorpaenopsis neglecta 减少 5.2 吉打副叶鲹 Alepes djedaba 减少 5.1 鲬 Platycephalus indicus 增加 4.9 细纹鲾 Leiognathus berbis 减少 4.7 少鳞鱚 Sillago japonica 增加 4.4 纤羊舌鲆 Arnoglossus tenuis 减少 4.4 鹿斑仰口鲾 Secutor ruconius 增加 4.1 赤鼻棱鳀 Thryssa kammalensis 增加 4.0
下载: 导出CSV
表 3 RDA分析结果显著性检验
Table 3 Significance test of redundancy analysis results
环境因子
Environment factorF p 水温 Water temperature 3.336 3 0.002** 化学需氧量 Chemical oxygen demand 2.806 3 0.005** 溶解氧 Dissolved oxygen 2.173 1 0.032* 浮游植物密度 Phytoplankton density 2.149 1 0.041* 总磷 Total phosphate 1.925 8 0.060 盐度 Salinity 1.325 2 0.225 透明度 Transparency 1.282 9 0.252 总氮 Total nitrogen 1.186 3 0.307 注:**. 在0.01水平上显著相关;*. 在0.05水平上显著相关。 Note: **. Significant correlation at 0.01 level; *. Significant correlation at 0.05 level.
下载: 导出CSV
-
[1] 陈丕茂, 舒黎明, 袁华荣, 等. 国内外海洋牧场发展历程与定义分类概述[J]. 水产学报, 2019, 43(9): 1981-1869. [2] 杨红生, 章守宇, 张秀梅, 等. 中国现代化海洋牧场建设的战略思考[J]. 水产学报, 2019, 43(4): 1255-1262. [3] 罗惠桂, 汪佳仪, 谢珍玉, 等. 三亚蜈支洲岛毗邻海域鱼类物种多样性及群落结构特征[J]. 海洋科学, 2023, 47(7): 74-86. [4] 陈丕茂, 袁华荣, 贾晓平, 等. 大亚湾杨梅坑人工鱼礁区渔业资源变动初步研究[J]. 南方水产科学, 2013, 9(5): 100-108. doi: 10.3969/j.issn.2095-0780.2013.05.016 [5] HU C, SHUI Y, TIAN K, et al. Functional group classification and niche identification of major fish species in the Qixing Islands Marine Reserve, Zhejiang Province[J]. Biodiv Sci, 2016, 24(2): 175-184. doi: 10.17520/biods.2015225
[6] 何倩, 刘淑德, 唐衍力, 等. 山东琵琶岛海域人工鱼礁区鱼类群落物种及功能多样性[J]. 中国水产科学, 2023, 30(12): 1479-1495. doi: 10.12264/JFSC2023-0195 [7] 冯雪, 戴小杰, 袁华荣, 等. 外伶仃海洋牧场附近海域游泳生物群落结构及其与环境因子的关系[J]. 上海海洋大学学报, 2024, 33(1): 186-201. doi: 10.12024/jsou.20230304115 [8] 张亚洲, 蒋日进, 梁君. 嵊泗马鞍列岛海域国家级海洋牧场渔业资源增殖养护效果评析[J]. 浙江海洋大学学报 (自然科学版), 2022, 41(5): 466-472. [9] 李寇军, 邱永松, 王跃中, 等. 自然环境变动对北部湾渔业资源的影响[J]. 南方水产, 2007, 3(1): 7-13. [10] 陈国宝, 李永振, 陈丕茂, 等. 南海北部陆架区海域蓝圆产卵场的研究[J]. 热带海洋学报, 2003, 22(6): 22-28. doi: 10.3969/j.issn.1009-5470.2003.06.004 [11] 张公俊, 杨长平, 孙典荣, 等. 北部湾中北部海域鱼类群落的季节变化特征[J]. 南方农业学报, 2021, 52(10): 2861-2871. doi: 10.3969/j.issn.2095-1191.2021.10.027 [12] 张曼, 王雪辉, 蔡研聪, 等. 北部湾带鱼空间聚散变化特征[J]. 中国水产科学, 2022, 29(11): 1647-1658. doi: 10.12264/JFSC2021-0609 [13] 侯刚, 冯钰婷 , 陈妍颖, 等. 北部湾二长棘犁齿鲷时空分布及其与环境因子的关系[J]. 广东海洋大学学报, 2021, 41(4): 8-16. [14] 黎树式, 戴志军, 葛振鹏, 等. 北部湾北部生态环境灾害变化研究[J]. 灾害学, 2014, 29(4): 43-47. doi: 10.3969/j.issn.1000-811X.2014.04.009 [15] JACCARD P. The distribution of flora in the alpine zone[J]. New Phytol, 1912, 11: 37-50. doi: 10.1111/j.1469-8137.1912.tb05611.x
[16] WARWICK R M. A new method for detecting pollution effects on marine macrobenthic communities[J]. Mar Biol, 1986, 92: 557-562. doi: 10.1007/BF00392515
[17] 裴精花, 陈清华, 范金金, 等. 珠江口海域游泳动物群落结构及多样性特征[J]. 南方农业学报, 2023, 54(12): 3727-3738. doi: 10.3969/j.issn.2095-1191.2023.12.027 [18] 王雪辉, 林昭进, 王跃中, 等. 南海西北部陆架区鱼类的种类组成与群落格局[J]. 生态学报, 2013, 33(7): 2225-2235. [19] 郭彪, 于莹, 张博伦, 等. 天津大神堂海域人工鱼礁区游泳动物群落特征变化[J]. 海洋渔业, 2015, 37(5): 409-418. doi: 10.3969/j.issn.1004-2490.2015.05.003 [20] 徐浩, 曾晓起, 顾炎斌, 等. 人工鱼礁对山东莱州朱旺港海区游泳动物的群落结构及季节变化的影响[J]. 中国海洋大学学报, 2012, 42(5): 47-54. [21] 罗峥力, 杨长平, 王良明, 等. 北部湾北部沿岸海域鱼类资源时空分布特征及多样性变化[J]. 南方农业学报, 2023, 54(6): 1847-1857. doi: 10.3969/j.issn.2095-1191.2023.06.026 [22] 冯雪, 范江涛, 孙晓, 等. 珠海外伶仃人工鱼礁对鱼类资源养护效果初步评估[J]. 南方农业学报, 2021, 52(12): 3228-3236. doi: 10.3969/j.issn.2095-1191.2021.12.005 [23] 印瑞, 周永东, 梁君, 等. 中街山列岛海洋牧场大黄鱼时空分布与环境因子的关系[J]. 浙江海洋大学学报 (自然科学版), 2022, 41(6): 483-489. [24] 韩业越, 殷蕊, 孙桂清, 等. 北戴河国家级海洋牧场示范区人工鱼礁建设效果评价[J]. 河北农业, 2020, 316(4): 24-28. [25] 王欢欢, 毕福洋, 曹敏, 等. 獐子岛海洋牧场秋季渔业资源声学调查与评估[J]. 大连海洋大学学报, 2018, 33(6): 802-807. [26] 崔晨, 张云岭, 张秀文, 等. 唐山祥云湾海洋牧场渔业资源增殖效果评估[J]. 河北农业, 2021(1): 25-31. [27] 李建都, 赵祺, 刘晋冀, 等. 黄渤海不同人工鱼礁区渔业生物群落结构特征及生物增量影响要素[J]. 中国水产科学, 2023, 30(3): 371-383. doi: 10.12264/JFSC2022-0406 [28] HYKEMA A, DEBROT A O, OSINGA R, et al. Fish assemblages of three common artificial reef designs during early colonization[J]. Ecol Eng, 2020, 157: 105994.
[29] 王宇, 侯纯强, 汪笑宇, 等. 天津近岸海域人工鱼礁区渔业资源增殖效果初步评估[J]. 海洋湖沼通报, 2018(2): 137-145. [30] 张荣良, 刘辉, 孙东洋, 等. 烟台近岸人工鱼礁与自然岩礁底层渔业生物群落特征对比分析[J]. 海洋与湖沼, 2021, 52(3): 697-707. doi: 10.11693/hyhz20201000296 [31] 王新萌, 唐衍力, 孙晓梅, 等. 威海小石岛人工鱼礁海域渔获物群落结构特征及其与环境因子相关性[J]. 海洋科学, 2016, 40(11): 34-39. doi: 10.11759/hykx20151123001 [32] ROSAS-ALAYOLA J, HEMANDES-HERRERA A, GALVAN-MAGANA F, et al. Diet composition of sailfish (Istiophorus platypterus) from the southern Gulf of California, Mexico[J]. Fish Res, 2002, 57: 185-195. doi: 10.1016/S0165-7836(01)00344-7
[33] KADYE W T, MAGADZA C H D, MOYO N A G, et al. Stream fish assemblages in relation to environmental factors on a montane plateau (Nyika, Malawi)[J]. Enriron Biol Fish, 2008, 83(4): 417-428. doi: 10.1007/s10641-008-9364-4
[34] SANDER R M J, CHANDLER C R, LANDRY A M J. Hydrological, diel and lunar factors affecting fishes on artificial reefs off Panama City, Florida[J]. Bull Mar Sci, 1985, 37(1): 318-328.
[35] OROPEZE O A, BALART E F. Community structure of reef fish in several habitats of a rocky reef in the Gulf of California[J]. Mar Ecol, 2001, 22(4): 283-305.
[36] 薛燕妮, 王洁英, 别又才, 等. 清远市南部淡水养殖鱼塘水质抽样调查与污染评价[J]. 黑龙江环境通报, 2024, 37(4): 40-42. doi: 10.3969/j.issn.1674-263X.2024.04.014 [37] 杨斌, 钟秋平, 鲁栋梁, 等. 钦州湾海域COD时空分布及对富营养化贡献分析[J]. 海洋科学, 2014, 38(3): 20-25. [38] 刘子萌, 冯民权, 杨锐婧, 等. 漳泽水库浮游植物群落结构特征及其影响因子[J]. 黑龙江大学工程学报, 2024, 15(1): 76-84. [39] 王奕杨, 毛志刚, 谷孝鸿, 等. 高宝邵伯湖鱼类群落结构及其多样性特征[J/OL]. 生态学杂志. https://link.cnki.net/urlid/ 21.1148.Q.20240205.1147.002 [40] MAES J, STEVENS M, BREINE J. Modelling the migration opportunities of diadromous fish species along a gradient of dissolved oxygen concentration in a European tidal watershed[J]. Estuar Coast Shelf Sci, 2007, 75(1/2): 151-162.
[41] 张雷敏, 梁化亮, 米海峰, 等. 低氧胁迫对鱼类的影响以及营养调控策略的研究进展[J]. 动物营养学报, 2023, 35(10): 6207-6219. doi: 10.12418/CJAN2023.570 [42] 曾嘉维, 林坤, 王学锋, 等. 雷州湾附近海域鱼类群落结构及其与环境因子的关系[J]. 中国水产科学, 2019, 26(1): 108-117. [43] 曾雷, 陈国宝, 李纯厚, 等. 大亚湾湾口游泳生物群落季节异质特征与生态效应分析[J]. 南方水产科学, 2019, 15(3): 22-32. doi: 10.12131/20180246 [44] 胡翠林, 张洪亮, 张亚洲, 等. 带鱼保护区春秋季鱼类群落特征及与环境因子的关系[J]. 水产学报, 2018, 42(5): 694-703. [45] 唐广隆, 刘永, 吴鹏, 等. 珠江口万山群岛海域春季渔业资源群落结构特征及其与环境因子的关系[J]. 中国水产科学, 2022, 29(8): 1198-1209. [46] 张雪, 徐晓甫, 戴媛媛, 等. 天津近岸人工鱼礁海域浮游植物群落及其变化特征[J]. 渔业科学进展, 2018, 39(6): 1-10. [47] 谢斌, 张硕, 李莉, 等. 海州湾海洋牧场浮游植物群落结构特征及其与水质参数的关系[J]. 环境科学学报, 2017, 37(1): 121-129. [48] 刘懂, 陈晨, 王莉, 等. 象山港海洋牧场示范区浮游植物落特征及其与环境因子的关系[J]. 海洋与湖沼, 2016, 47(5): 1024-1032. doi: 10.11693/hyhz20160500096 -
期刊类型引用(1)
1. 刘睿,梁颖,殷豆豆,宋超东,莫祖琴,肖骏峰,易晗,张红岩,张彬,申乃坤. 降解亚硝酸盐菌株Bacillus velezensis GXMZU-B1的筛选、条件优化及初步应用. 南方水产科学. 2024(04): 133-143 .  本站查看
本站查看
其他类型引用(2)



计量
- 文章访问数: 1211
- HTML全文浏览量: 34
- PDF下载量: 92
- 被引次数: 3
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
