
Photophysiological responses of Gracilariopsis bailinae to temperature and light intensity
-
摘要: 大型海藻对温度和光照强度的适应存在种属差异。异枝江蓠 (Gracilariopsis bailinae) 是一种喜高温的大型海藻,为科学指导该藻在海水养殖、海洋生态修复等方面的应用,利用叶绿素荧光技术,结合藻体光合色素含量和生长率变化,探究了异枝江蓠对温度和光照强度的光合生理响应特征。测定了15、20、25、30、35 ℃和1 000、3 000、6 000、9 000 lx条件下,异枝江蓠的特定生长率 (SGR)、光合色素含量 [叶绿素a (Chl a)、类胡萝卜素 (Car)、藻胆蛋白] 及叶绿素荧光参数 [PSII最大光化学效率 (Fv/Fm)、实际光能转化效率 (ΦPSII)、电子传递速率 (ETR)、光化学淬灭 (qP)、非光化学淬灭 (NPQ)] 的变化。结果显示,温度和光照强度对上述相关指标 (除Car和Fv/Fm) 的影响具有极显著的交互作用 (P<0.01);温度升高显著增加了异枝江蓠的SGR、藻胆蛋白含量以及叶绿素荧光参数值 (P<0.05);高光照显著降低了异枝江蓠的光合色素含量以及Fv/Fm、ΦPSII、ETR和qP值,但NPQ和SGR却显著上升 (P<0.05)。结果表明,异枝江蓠是一种喜高温的大型海藻,通过增加藻胆蛋白含量可提高其在高温条件下的生存能力,同时,通过增加热耗散和减少光合色素的合成,可实现藻体在高光下的光保护。Abstract: There are species differences in the adaptation of macroalgae to temperature and light intensity. Gracilariopsis bailinae is a large alga that likes high temperature. In order to scientifically guide its application in mariculture and marine ecological restoration, we investigated the photosynthetic physiological responses characteristics of G. bailinae to temperature and light intensity by using chlorophyll fluorescence technology, combined with changes in photosynthetic pigment content and growth rate of algae. We measured various physiological parameters, including specific growth rate (SGR), photosynthetic pigments content [Chlorophyll a (Chl a), carotenoid (Car), and phycobiliprotein], and chlorophyll fluorescence parameters [Maximum photochemical efficiency of PSII (Fv/Fm), actual light conversion efficiency (ΦPSII), electron transfer rate (ETR), photochemical quenching (qP), and non-photochemical quenching (NPQ)] of G. bailinae cultured at five different temperatures (15, 20, 25, 30, 35 ℃) and four different light intensities (1 000, 3 000, 6 000, 9 000 lx). Results reveal that temperatures and light intensities had a significant interaction effect on the parameters measured, except for Car and Fv/Fm (P<0.01). The SGR, phycobiliprotein contents, and chlorophyll fluorescence parameters of G. bailinae increased significantly as temperature increased (P<0.05), whereas high light intensity treatment led to a significant decrease in photosynthetic pigments content and values of Fv/Fm, ΦPSII, ETR, and qP, but a significant increase in NPQ and SGR (P<0.05). These results indicate that G. bailinae can enhance its survival capacity under high temperature conditions by increasing the synthesis of phycobilin, and achieve photoprotection under high light conditions by increasing heat dissipation and decreasing photosynthetic pigment synthesis.
-
长体圆鲹(Decapterus macrosoma),又名长身圆鲹,隶属于鲈形目、鲹科、圆鲹属,主要分布于中国南海、印度尼西亚、澳洲和日本南部沿海等地[1]。长体圆鲹在中国南海分布较广,是南海灯光围网渔业的主要捕捞对象,具有较高的经济价值[2-5]。目前,国内外学者关于长体圆鲹的研究主要集中在生长繁殖[6]和资源评估[7-8]方面,与种群遗传和分子标记相关的研究报道较少。微卫星标记仅见翟云等[9]开发蓝圆鲹微卫星标记中获得5个跨物种通用标记可于用长体圆鲹,并无专门针对长体圆鲹开发的微卫星标记。种群遗传信息的匮乏,将大大影响对其资源的评估和长期有效的管理。
微卫星分子标记因是共显性标记,具有多态性高、变异性强、数据易统计等突出优点[10],广泛应用于海洋生物遗传结构及遗传多样性分析[11-12]。但由于微卫星标记通用性较差,常常具有极强的种属特异性。鱼类微卫星标记开发中多以二核苷酸重复为主[13-15],普遍认为它们具有较高的遗传变异[16],但是也有部分学者研究认为三、四核苷酸重复位点较二核苷酸重复具有更高的筛选效率和多态性[17-19]。
本研究通过RAD-Seq高通量测序方法开发长体圆鲹二、三核苷酸微卫星分子标记,并对测试群体进行多样性分析,同时比较二、三核苷酸的筛选效率和多态性差异,旨在为长体圆鲹种群遗传结构及遗传多样性分析提供技术基础,并为该资源的评估和管理提供帮助。
1. 材料与方法
1.1 样品采集与基因组DNA提取
长体圆鲹样品采集于中国南海中沙群岛东部海域,共35尾。剪取部分肌肉样品加入无水乙醇保存。每个样品剪取少量肌肉组织,使用海洋动物组织基因组DNA提取试剂盒(天根,北京) 提取基因组DNA,0.8%的琼脂糖凝胶电泳检测DNA提取质量,−20 ℃保存备用。
1.2 高通量测序与引物合成
使用HiSeq2000高通量测序仪(Illumina,USA) 对长体圆鲹基因组DNA进行RAD-seq (测序服务由广州基迪奥生物科技有限公司提供),经生物信息学搜索出微卫星位点[20]。使用Premier 5.0软件在重复单元侧翼序列上选择性设计出112条引物,主要参数为:G-C含量为40%~60%,引物长度为18~25 bp,退火温度为45~60 ℃,预期产物长度180~320 bp。送上海英潍捷基贸易有限公司合成引物。
1.3 引物筛选与分型检测
选取3个样本混合成的基因组DNA为模板,优化PCR反应条件,对引物进行首轮筛选,琼脂糖电泳检测是否能扩增出稳定且均一的目的片段。之后选取8尾个体的基因组DNA作为模板,使用三引物法[21],利用M13荧光接头引物进行PCR扩增,扩增产物送华大基因公司经毛细管电泳进行等位基因分型,检测引物是否具有多态性。PCR反应体系为15 μL,其中包括10×PCR Buffer 1.5 μL,2.5 mmol·L–1 MgCL2 1.2 μL,2 mmol·L–1 dNTPs 2 μL,M13正向引物(10 μmol·L–1) 0.2 μL,M13反向引物(10 μmol·L–1) 0.6 μL,M13通用荧光引物(10 μmol·L–1) 0.5 μL,Taq酶(5 U·μL–1) 0.15 μL,DNA模版1 μL,加双蒸水至15 μL。PCR扩增程序为:94 ℃预变性5 min;94 ℃变性45 s,55~60 ℃退火45 s,72 ℃延伸45 s,35个循环;94 ℃变性45 s,53 ℃退火45 s,72 ℃延伸45 s,8个循环;72 ℃延伸30 min。
1.4 长体圆鲹群体遗传学评价
使用35尾长体圆鲹个体的基因组DNA为模板,对通过筛选的微卫星标记的种群遗传学特征进行评价。PCR反应体系和条件、等位基因分型方法如上。使用软件Genepop 4.0[22]对每个标记的种群遗传学特征值进行计算,包括等位基因数(Na)、表观杂合度(Ho)和期望杂合度(He),进行“哈迪-温伯格”平衡(HWE)检验和连锁不平衡检测,并对P值进行Bonferroni校正。使用Cervus 3.0.7[23]软件计算多态信息含量(PIC)。
2. 结果
2.1 高通量测序结果与微卫星位点分析
RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,Q30达93.05%。说明测序结果质量较好,可用于后续分析。搜索后共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个,其中二核苷酸重复微卫星位点最多(37 646个),占总数的53.39% (表1),说明二核苷酸重复为主要的微卫星类型。二核苷酸重复微卫星位点共有4种重复类型,4种类型重复微卫星数量相差较大,AC/GT类有29 754个,占二核苷酸重复的68.4%;AG/CT类有6 487个,占17.2%;AT/TA有1 340个,占3.6%;GC/CG仅有65个,占0.17%。
表 1 长体圆鲹基因组中不同类型SSR统计Table 1. Different types of SSR statistics in D. macrosoma genome重复单元
repeat unit微卫星数量/个
number of microsatellite占比/%
ratio一核苷酸 mono-nucleotide 8 184 11.61 二核苷酸 di-nucleotide 37 646 53.39 三核苷酸 tri-nucleotide 13 960 19.80 四核苷酸 tetra-nucleotide 7 741 10.98 五核苷酸 penta-nucleotide 2 255 3.20 六核苷酸 hexa-nucleotide 722 1.02 合计 total 70 508 100.00 2.2 PCR引物设计和筛选
选取112条二、三核苷酸重复序列设计引物,其中二核苷酸重复为81对,三核苷酸重复为31对。经过筛选后,共有27对引物通过筛选(表2),27对引物扩增的序列中18个位点为二核苷酸重复,重复次数为9~14次;8个位点为三核苷酸重复,重复次数为6~10次。二核苷酸重复位点检出效率为22.2%,三核苷酸重复位点检出效率为25.8%。
表 2 27对长体圆鲹微卫星引物信息Table 2. Information of 27 pairs of primers in D. macrosoma位点
locus引物序列 (5'−3')
primer sequence重复单元
repeat motif退火温度/℃
annealing temperature期望长度/bp
allele sizeDma03 F:CCACGCCTATTGAGTTACAGA (CA)9 60 186 R:GAGCCAGTGGATGAACAGAGT Dma07 F:GCCCCTGTGGGTGTGTGA (CA)9 60 225 R:GGGTGGTGGGTTCGGTTT Dma12 F:GAACCAGTGCCTACAATAGA (AC)9 60 243 R:CTGCTCACGGTAAGTCCA Dma15 F:ACAGGAAGGAACAGGACAG (TG)10 55 254 R:TATTGAAGTGAAAAAGCCG Dma22 F:CGCTGTTGAAATGAAGAAGA (GT)10 60 317 R:AGTGATGTCGCCTCATAAAT Dma23 F:AAACTGAGGGCGAGATAGAGG (AC)10 55 190 R:CCACAGGCTGAGTAAACCAAC Dma26 F:ATCCCATTCACCGACATAG (TG)10 58 258 R:CTGTGGTATCGTTCCCTGT Dma28 F:TGATTGGCTTCTACTCTGC (AC)10 55 281 R:AGTGGCTTGTTTGACTCTTAT Dma36 F:GGATGTAGTGAAGAGGGGAG (GT)11 55 239 R:CACAATCAGTGTTATGGCAG Dma38 F:GCCAATAAAGGCAAACAGT (CA)11 60 227 R:ATCCGAGACAAAGACATACAA Dma39 F:AGTGTGCTGACTTTTCTCTG (CA)11 55 241 R:TTATTGTTTGTTGTCTGGGT Dma45 F:CTCCTTTTTCTTCTTCCTCT (CA)11 60 281 R:CTACCTGCTCTTCAACTCAT Dma51 F:TGACAGCCTCCACTACTCC (GA)12 55 225 R:GCTAACCAGACACGCAAA Dma54 F:AAAGCCCATCTGTCTCGT (GT)12 60 202 R:TGTTTCAGTCCGTTCCTG Dma58 F:TCAAGAGGGAGTGGGAGC (AC)12 58 279 R:TCAAATGGGTGTTTAGCG Dma64 F:GCTCAGACTGCGTGGACA (TG)13 55 314 R:GCTGGTGAACAACAGGACA Dma72 F:TTCCGCAGGCATAAAAAC (CT)13 58 301 R:CCAAGGTCCGCTACACTA Dma76 F:TTCTCGCTGACCTGCTTG (TG)14 55 253 R:GCGTCCTCGTCGTCTTTC Dma81 F:GAGACACGGTCAGAAAACA (TGC)6 60 216 R:GGAAGTAGGACTCTAGGGG Dma82 F:CTGTCACTCCATTCCTATTCC (GTT)6 58 244 R:CCTACATTTGTGCTTTTGTTC Dma83 F:CTCTAAAGCCGACCTAACC (CTT)6 58 239 R:TGTCTCAACACAGCGAAAC Dma84 F:AAACTAACTCATCACCAG (TGT)6 55 283 R:AAACGACAGGAACTCAAT Dma85 F:CTCACTTTGACCCAACCAG (AGG)6 55 256 R:CCTTTCACCGAGACACCAG Dma131 F:TGCGGATGGGTGGTAGTGT (GGT)8 55 208 R:ATTGCTGGTAGTCGGTGGC Dma132 F:CCCAGTGAGACCAGAACCA (GCT)8 55 268 R:GACCCGTAGACAGGAGAGT Dma135 F:GTTGTTGTTTTTTTCCTT (GCA)9 55 301 R:CATCAGTCTGGCTTTATA Dma145 F:ACGATACAGCAGCCGAAG (TCA)10 60 197 R:AGTGATGTCGCCTCATAAAT 2.3 微卫星标记的种群遗传学评价
使用1个采集自南海东南部海域的长体圆鲹群体对筛选合格的微卫星标记进行种群遗传学评价。所有27个标记在测试群体中共检测到285个等位基因,等位基因数为5~17,Ho为0.342 9~0.857 1,平均为0.631 7;He为0.538 3~0.911 8,平均为0.7968。PIC为0.497~0.886,平均为0.780 9 (表3),表明开发的微卫星位点具有较高的多态性。共有19个标记等位基因频率符合“哈迪-温伯格”平衡。连锁不平衡检测表明各位点间无连锁不平衡现象。
表 3 长体圆鲹微卫星标记的种群遗传学特征Table 3. Characteristics of microsatellite loci in D. macrosoma位点
locusN Na Ho He PHWE PIC Dma03 35 8 0.857 1 0.790 9 0.042 0 0.746 0 Dma07 35 9 0.771 4 0.855 9 0.553 4 0.825 0 Dma12 34 11 0.685 7 0.816 6 0.106 4 0.809 0 Dma15* 35 14 0.542 9 0.911 8 0.000 0 0.890 0 Dma22* 32 14 0.485 7 0.813 2 0.000 0 0.864 0 Dma23 34 17 0.714 3 0.864 8 0.003 3 0.868 0 Dma26* 34 14 0.428 6 0.869 1 0.000 0 0.870 0 Dma28 35 11 0.742 9 0.837 7 0.035 3 0.803 0 Dma36 35 9 0.685 7 0.786 3 0.054 5 0.743 0 Dma38* 34 12 0.628 6 0.851 2 0.000 0 0.849 0 Dma39* 33 12 0.514 3 0.782 0 0.000 0 0.793 0 Dma45 34 12 0.771 4 0.847 8 0.139 2 0.844 0 Dma51* 31 12 0.485 7 0.784 5 0.000 0 0.859 0 Dma54 35 15 0.771 4 0.900 6 0.139 0 0.878 0 Dma58 34 12 0.771 4 0.845 6 0.449 7 0.843 0 Dma64* 31 13 0.428 6 0.788 3 0.000 0 0.865 0 Dma72 35 8 0.628 6 0.713 9 0.155 7 0.659 0 Dma76 35 12 0.685 7 0.908 1 0.003 6 0.886 0 Dma81 35 8 0.628 6 0.795 0 0.045 7 0.752 0 Dma82 35 5 0.485 7 0.538 3 0.023 6 0.497 0 Dma83* 34 8 0.342 9 0.705 8 0.000 0 0.674 0 Dma84 35 7 0.542 9 0.704 8 0.005 9 0.649 0 Dma85 35 8 0.828 6 0.747 8 0.876 9 0.704 0 Dma131 35 9 0.628 6 0.717 6 0.646 6 0.661 0 Dma132 35 8 0.657 1 0.746 6 0.365 5 0.692 0 Dma135 34 10 0.742 9 0.840 5 0.241 8 0.835 0 Dma145 34 7 0.600 0 0.749 3 0.077 4 0.725 0 注:N. 有效样品数;Na. 等位基因数;Ho. 表观杂合度;He. 期望杂合度;PHWE. “哈迪-温伯格”平衡显著性检验P值;PIC. 多态信息含量;*. 经Bonferroni校正后显著背离“哈迪-温伯格”平衡 (校正P<0.001 85) Note: N. effective number of samples; Na. number of alleles; Ho. observed heterozygosity; He. expected heterozygosity; PHWE. Hardy–Weinberg probability test; PIC. polymorphism information content; *. significant deviation from HWE after Bonferroni's correction (adjusted P-value<0.001 85) 3. 讨论
3.1 高通量测序发掘微卫星序列的技术优势
传统微卫星标记开发方法耗时长、花费高、技术难度大。以磁珠富集法为例,标记开发过程中基因组DNA浓度、接头连接效率、富集过程中的杂交温度以及洗脱条件的控制等因素都会影响微卫星筛选的效率[24-25],且最终获得的有效微卫星序列仅几百条[26-27]。相比较而言,高通量测序技术开发微卫星标记,省略了建库、克隆、筛选等,只需提取基因组DNA测序,利用生物信息学手段可直接获取微卫星序列,通常是传统方法获得微卫星序列数目的几百倍[15,28-29],具有高效、便捷、准确的特点,能够满足短时间内大批量微卫星位点的开发需求,比如连锁图谱构建、QTL定位等[30-31]。
本次RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,测序质量Q30达93.05%;共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个。说明测序质量稳定高效,并获得了数量庞大、类型丰富的长体圆鲹微卫星序列,可用于后续长体圆鲹微卫星标记的大规模开发和相关遗传学研究。
3.2 不同核苷酸重复微卫星位点比较
本次高通量测序结果表明在长体圆鲹微卫星位点中二核苷酸重复为主要重复类型,AC/GT类重复数量最为丰富,GC/CG重复较为少见。此结果与大量水产动物微卫星位点研究结果相一致[32-34],差异仅在于比例多少,以及除二核苷酸重复占主要地位外其他核苷酸重复的含量差异。熊良伟等[33]对中华鳑鲏(Rhodeus sinensis)微卫星的分析中,二核苷酸占总微卫星位点的53.59%,其次为单核苷酸重复,二核苷酸重复中AC/GT类占60.63%,GC/CG仅占0.32%。在裸体异鳔鳅鮀 (Xenophysogobio nudicorpa)中[32],二核苷酸重复占总微卫星位点比例高达83.15%,AC/GT类重复占49.36%,GC/CG重复仅有4个。
多数鱼类开发的微卫星标记以二核苷酸重复为主,但研究表明,在人类基因组中三核苷酸重复序列与遗传疾病的发生有关,并且具有较高的多态性和遗传稳定性[35]。部分学者对三、四核苷酸重复微卫星标记的研究结果存在差异。房祖业等[28]对大刺鳅 (Mastacembelus armatus) 二、三、四核苷酸重复微卫星标记的筛选发现二核苷酸重复较三、四核苷酸重复具有更高的筛选效率和多态性;鲁翠云等[17]、谭照君等[18]、李文升等[19]的研究认为三、四核苷酸具有更高的多态性和分型效果。长体圆鲹二、三核苷酸的筛选效率分别为22.2%和29.0%,PIC分别为0.827 4和0.687 7 (表4)。就筛选效率而言,三核苷酸重复略高于二核苷酸重复,但二者相差不大。PIC为衡量种群遗传变异程度的重要指标[36],二核苷酸重复多态性明显高于三核苷酸重复。本文中长体圆鲹二核苷酸重复筛选效率低于三核苷酸重复,但多态性二核苷酸重复明显高于三核苷酸重复。因此,筛选效率和多态性的差异可能由种属差异或其他多种因素导致。
表 4 长体圆鲹二、三核苷酸重复微卫星标记的比较Table 4. Comparison on di- and trinucleotide-repeated microsatellite loci in D. macrosoma序列
sequence引物数
primer number重复次数
repeat number筛选效率
efficiencyPIC 二核苷酸重复
di-nucleotide-repeated18 9~14 22.2% 0.827 4 三核苷酸重复
tri-nucleotide-repeated9 6~10 29.0% 0.687 7 通过筛选的27对引物中18个位点为二核苷酸重复,重复次数为9~14次不等;9个位点为三核苷酸重复,重复次数为6~10次不等,符合Ellegren[37]提出的真核生物微卫星位点重复大部分在30次重复以下。但Ellegren[37]认为二核苷酸重复以15~19次为主,本文中高通量测序获得的二核苷酸重复主要在6~15次。基于Weber[38]的研究结果,重复次数高的微卫星在种群中表现出的多态性较高,龚小玲等[39]对澳洲鳗鲡 (Anguilla australis) 进行标记开发时发现,微卫星重复序列的重复次数过高会影响PCR效果,应选择居中的重复次数为宜。长体圆鲹二核苷酸重复PIC为0.827 4,具有较高多态性,表明选择6~15次的二核苷酸重复是合适的。
3.3 微卫星标记的种群遗传学特征
群体杂合度的高低反映了群体在多个基因座上的遗传变异及群体遗传多样性丰富度[19]。本研究中长体圆鲹中沙群体的平均Ho为0.631 7,平均He为0.796 8,说明长体圆鲹该群体的遗传多样性较高。平均Ho和He存在差异,说明存在杂合子缺失或者纯合子过剩的情况。PIC也是衡量群体遗传多样性的重要指数,Botstein等[36]认为基因标记PIC>0.5为高度多态位点,0.25<PIC<0.5为中度多态位点,PIC<0.25为低度多态性位点,通常不作为遗传多样性分析。本文中长体圆鲹位点除1个为中度多态外,其他位点均为高度多态位点。表明开发所得的长体圆鲹微卫星标记在中沙群体中具有较好的遗传稳定性和丰富的遗传多样性。
在所有27个位点中有8个位点偏离了“哈迪-温伯格”平衡,这些位点不适合进一步的遗传分析。近亲杂交、无效等位基因、种群退化和自然选择等因素皆可能导致微卫星位点偏离HWE[15]。
-
![]()
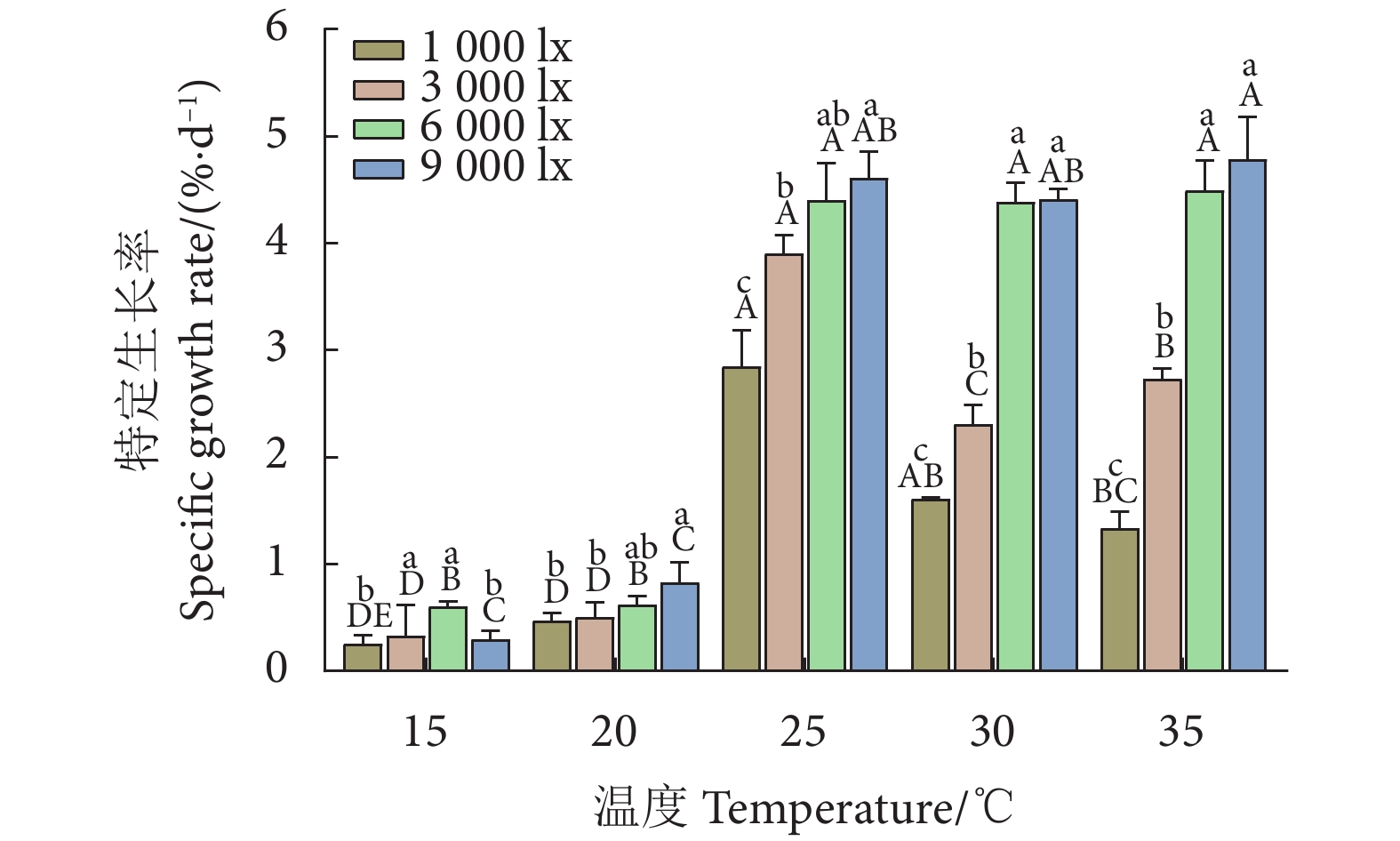
图 1 不同温度和光照强度下的异枝江蓠特定生长率
注:不同小写字母表示在相同温度条件下不同光照强度处理间差异显著 (P<0.05),不同大写字母表示在相同光照强度条件下不同温度处理间差异显著 (P<0.05);后图同此。
Figure 1. Specific growth rate of G. bailinae at different temperatures and light intensities
Note: Different lowercase letters indicate significant differences between different light intensity treatments under the same temperature condition (P<0.05), while different uppercase letters indicate significant differences between different temperature treatments under the same light intensity condition (P<0.05). The same case in the following figures.
![]()
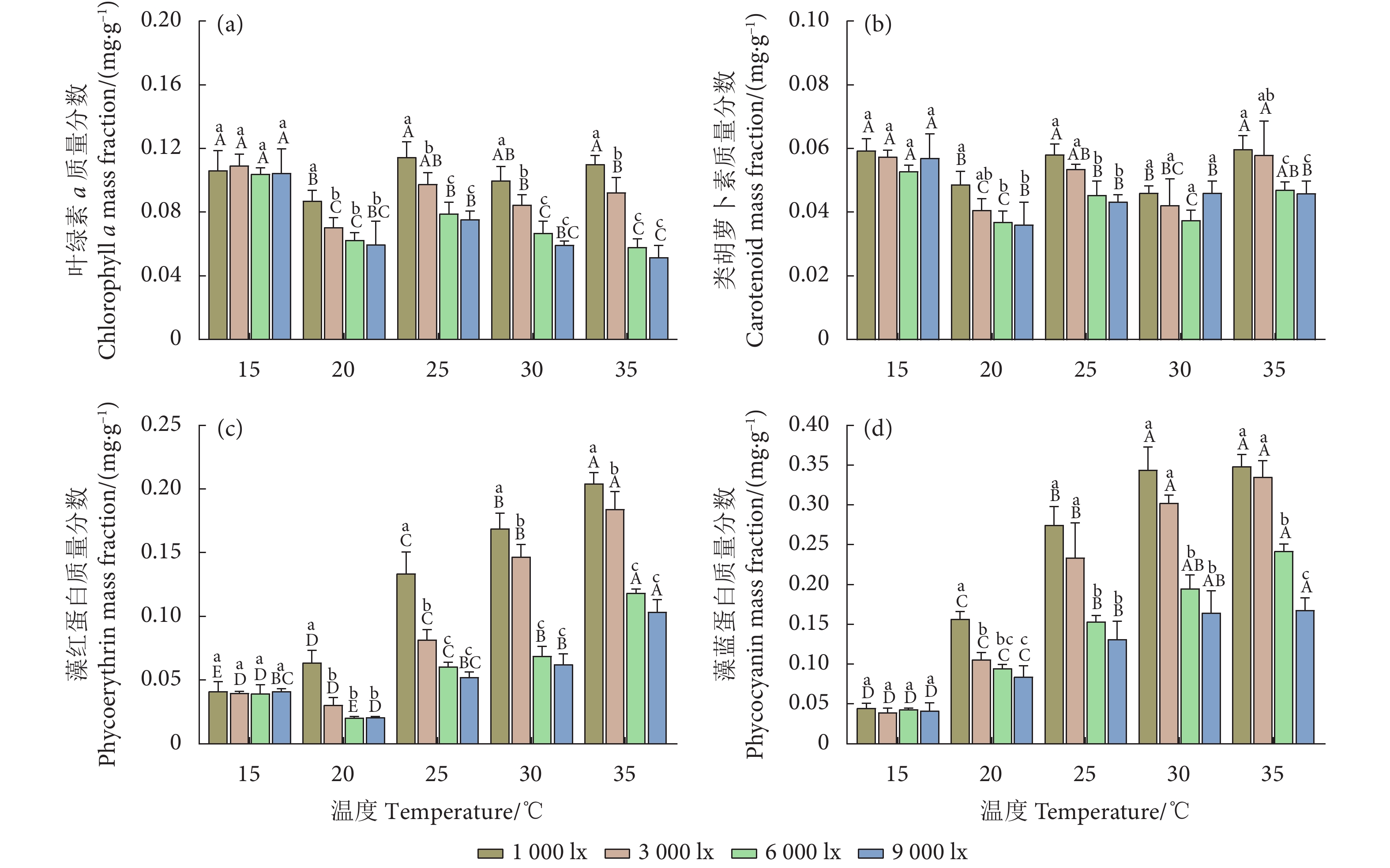
图 2 不同温度和光照强度下的异枝江蓠色素质量分数
Figure 2. Pigment mass fraction of G. bailinae at different temperatures and light intensities
![]()
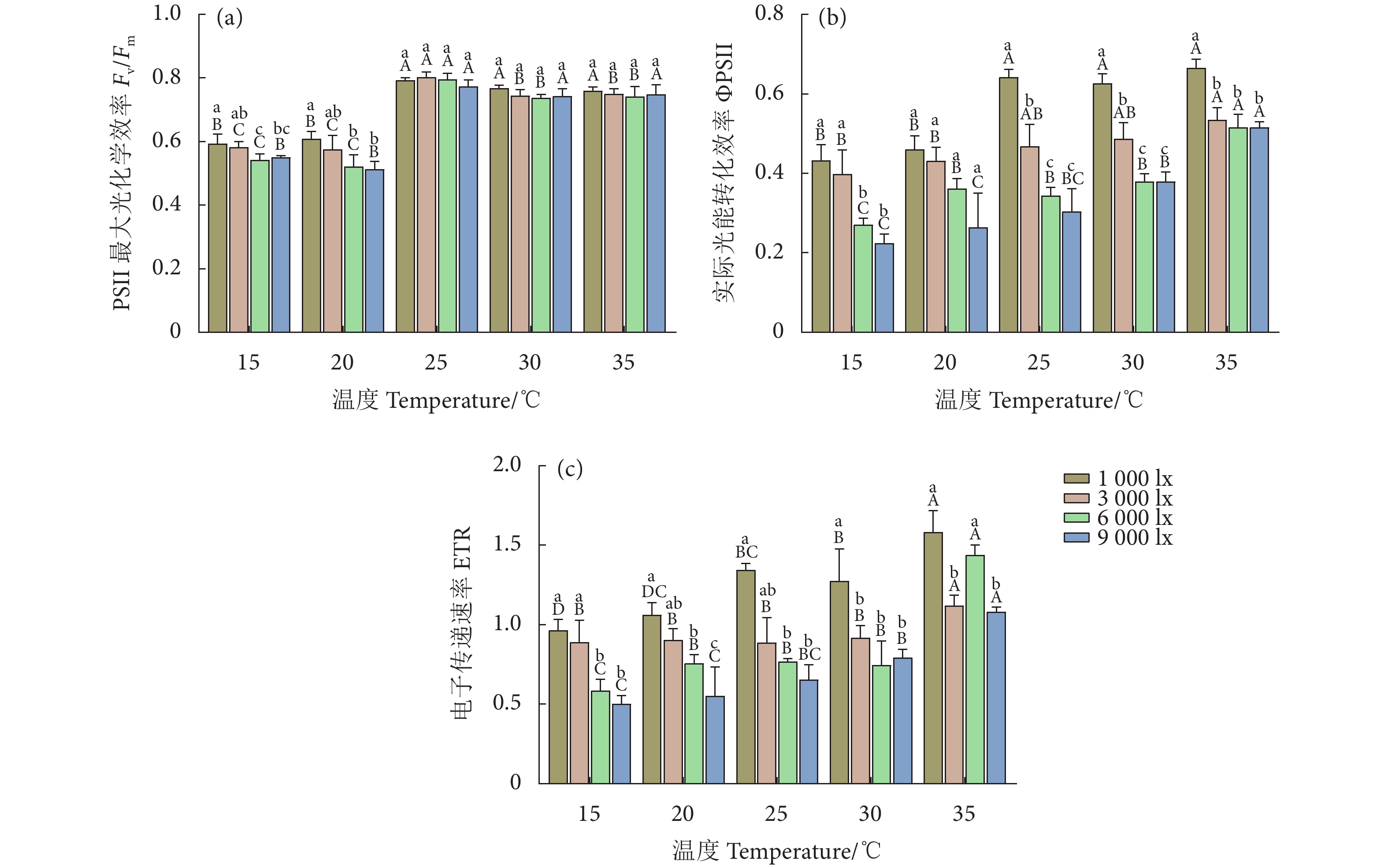
图 3 不同温度和光照强度下的异枝江蓠光化学效率参数
Figure 3. Photochemical efficiency parameters of G. bailinae at different temperatures and light intensities
![]()
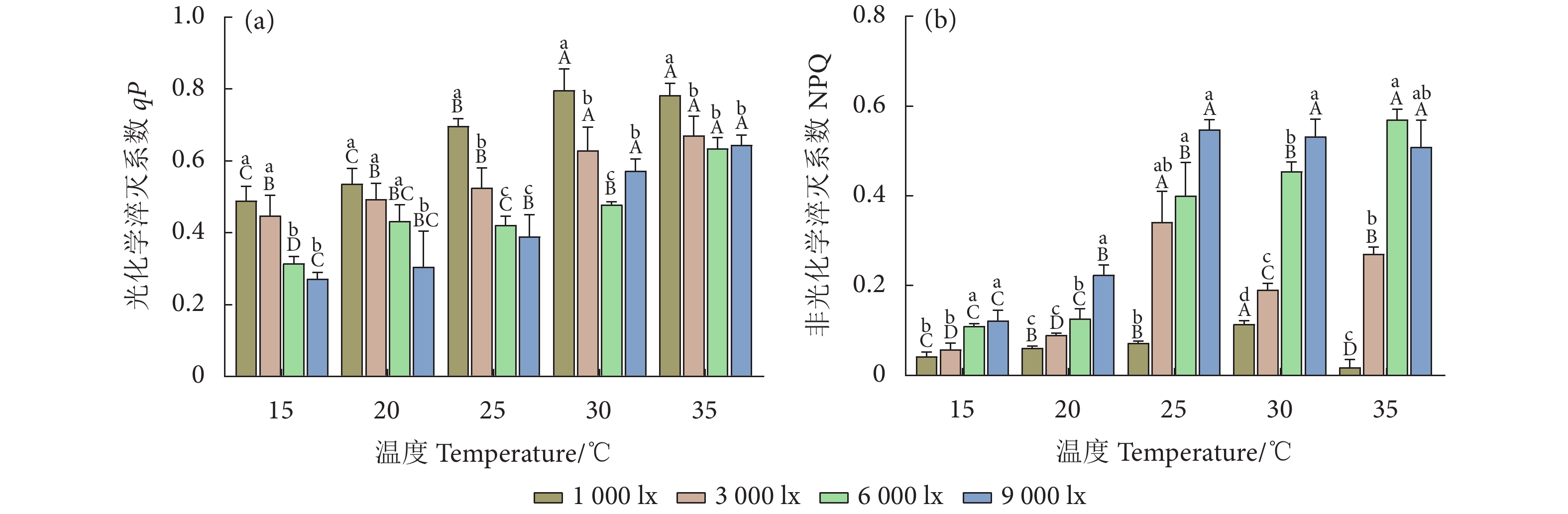
图 4 不同温度和光照强度下的异枝江蓠荧光淬灭参数
Figure 4. Fluorescence quenching parameters of G. bailinae at different temperatures and light intensities
表 1 温度、光照强度与异枝江蓠生长、光合色素及叶绿素荧光参数的相关性分析
Table 1 Correlation analysis between temperature and light intensity and growth, photosynthetic pigments and chlorophyll fluorescence parameters of G. bailinae
项目
Item光照强度 Light intensity/lx 温度 Temperature/℃ 1 000 3 000 6 000 9 000 15 20 25 30 35 特定生长率 SGR 0.501 0.668** 0.873** 0.889** 0.197 0.766** 0.898** 0.938** 0.958** 叶绿素 a Chl a 0.242 −0.195 −0.704** −0.727** −0.130 −0.782** −0.910** −0.935** −0.950** 类胡萝卜素 Car −0.042 0.041 −0.188 −0.215 −0.310 −0.736** −0.910** −0.101 −0.730** 藻红蛋白 PE 0.978** 0.948** 0.886** 0.842** 0.011 −0.849** −0.910** −0.937** −0.950** 藻蓝蛋白 PC 0.951** 0.969** 0.992** 0.923** −0.130 −0.882** −0.920** −0.947** −0.950** PSII最大光化学效率 Fv/Fm 0.804** 0.745** 0.752** 0.792** −0.710** −0.799** −0.420 −0.532 −0.230 实际光能转化效率 ΦPSII 0.885** 0.776** 0.878** 0.895** −0.910** −0.853** −0.940** −0.913** −0.790** 电子传递速率 ETR 0.859** 0.537* 0.784** 0.890** −0.910** −0.902** −0.900** −0.771** −0.600* 光化学淬灭系数 qP 0.918** 0.866** 0.912** 0.929** −0.920** −0.835** −0.920** −0.755** −0.760** 非光化学淬灭系数 NPQ 0.007 0.679** 0.952** 0.848** 0.912** 0.936** 0.937** 0.966** 0.902** 注:** 表示在0.01水平 (双尾) 极显著相关,* 表示在0.05水平 (双尾) 显著相关。 Note: **. Extremely significant correlation at 0.01 level (Two-tailed); *. A significant correlation at 0.05 level (Two-tailed).  下载: 导出CSV
下载: 导出CSV
表 2 温度和光照强度对异枝江特定生长率的双因素方差分析
Table 2 Two-way ANOVA analysis for effects of temperature and light intensity on specific growth rate of G. bailinae
变异来源
Source of variation自由度
DFF 显著性
Sig.温度
Temperature4 802.800 <0.000 1 光照强度
Light intensity3 233.200 <0.000 1 温度×光照强度
Temperature×Light intensity12 37.810 <0.000 1 残差
Residual40
下载: 导出CSV
表 3 温度和光照强度对异枝江蓠光合色素的双因素方差分析
Table 3 Two-way ANOVA analysis for effects of temperature and light intensity on photosynthetic pigments of G. bailinae
光合色素 Photosynthetic pigment 变异来源 Source of variation 自由度 DF F 显著性 Sig. 叶绿素 a Chl a 温度 Temperature 4 36.650 <0.000 1 光照强度 Light intensity 3 53.480 <0.000 1 温度×光照强度 Temperature×Light intensity 12 4.415 0.000 2 残差 Residual 40 类胡萝卜素 Car 温度 Temperature 4 23.850 <0.000 1 光照强度 Light intensity 3 14.830 <0.000 1 温度×光照强度 Temperature×Light intensity 12 1.490 0.168 4 残差 Residual 40 藻红蛋白 PE 温度 Temperature 4 448.300 <0.000 1 光照强度 Light intensity 3 219.400 <0.000 1 温度×光照强度 Temperature×Light intensity 12 25.370 <0.000 1 残差 Residual 40 藻蓝蛋白 PC 温度 Temperature 4 356.000 <0.000 1 光照强度 Light intensity 3 131.500 <0.000 1 温度×光照强度 Temperature×Light intensity 12 14.400 <0.000 1 残差 Residual 40
下载: 导出CSV
表 4 温度和光照强度对异枝江蓠 PSII 光化学效率参数的双因素方差分析
Table 4 Two-way ANOVA analysis for effects of temperature and light intensity on photochemical efficiency parameters of PSII of G. bailinae
光化学效率参数
Photochemical efficiency parameter变异来源
Source of variation自由度
DFF 显著性
Sig.PSII 最大光化学效率 Fv/Fm 温度 Temperature 4 290.800 <0.000 1 光照强度 Light intensity 3 10.380 <0.000 1 温度×光照强度 Temperature×Light intensity 12 1.864 0.069 9 残差 Residual 40 实际光能转化效率 ΦPSII 温度 Temperature 4 61.560 <0.000 1 光照强度 Light intensity 3 106.000 <0.000 1 温度×光照强度 Temperature×Light intensity 12 4.071 0.000 4 残差 Residual 40 电子传递速率 ETR 温度 Temperature 4 53.820 <0.000 1 光照强度 Light intensity 3 70.460 <0.000 1 温度×光照强度 Temperature×Light intensity 12 4.001 0.000 4 残差 Residual 40
下载: 导出CSV
表 5 温度和光照强度对异枝江蓠荧光淬灭参数的双因素方差分析
Table 5 Two-way ANOVA analysis for effects of temperature and light intensity on fluorescence quenching parameters of G. bailinae
荧光淬灭参数
Fluorescence quenching parameter变异来源
Source of variation自由度
DFF 显著性
Sig.光化学淬灭系数 qP 温度 Temperature 4 86.340 <0.000 1 光照强度 Light intensity 3 73.660 <0.000 1 温度×光照强度 Temperature×Light intensity 12 3.471 0.001 5 残差 Residual 40 非光化学淬灭系数 NPQ 温度 Temperature 4 204.600 <0.000 1 光照强度 Light intensity 3 338.400 <0.000 1 温度×光照强度 Temperature×Light intensity 12 33.670 <0.000 1 残差 Residual 40
下载: 导出CSV
-
[1] 刘榆莎, 王东, 徐晓婷, 等. 温度和盐度对浒苔生长和光合生理特性的影响[J]. 水生生物学报, 2016, 40(6): 1227-1233. doi: 10.7541/2016.160 [2] 史彦江, 罗青红, 宋锋惠, 等. 高温胁迫对新疆榛光合参数和叶绿素荧光特性的影响[J]. 应用生态学报, 2012, 23(9): 2477-2482. doi: 10.13287/j.1001-9332.2012.0342 [3] 张宝, 徐燕, 许凯, 等. 坛紫菜响应高光胁迫的分子机制[J]. 水产学报, 2022, 46(11): 2066-2075. [4] 杨宇峰, 罗洪添, 王庆, 等. 大型海藻规模栽培是增加海洋碳汇和解决近海环境问题的有效途径[J]. 中国科学院院刊, 2021, 36(3): 259-269. doi: 10.16418/j.issn.1000-3045.20210217103 [5] 付倩倩, 李航霄, 吴海龙, 等. 光强对缘管浒苔 (Ulva linza) 光合生理特性和短期温度效应的影响[J]. 海洋与湖沼, 2018, 49(5): 967-974. doi: 10.11693/hyhz20180400074 [6] 王晓艳. 不同温度和光照强度对裙带菜 (Undaria pinnatifida) 幼孢子体叶绿素荧光参数和抗氧化系统的影响[D]. 青岛: 中国海洋大学, 2015, 1-54. [7] 程晓鹏, 章守宇, 林军, 等. 海带孢子体光合活性对不同温度和光照的响应[J]. 水产学报, 2020, 44(2): 234-244. [8] 钟逸云, 杨蕴琪, 郜晓峰, 等. 盐度、温度和光照强度对针叶蕨藻的生长及光合活性的影响[J]. 热带亚热带植物学报, 2021, 29(6): 626-633. doi: 10.11926/jtsb.4378 [9] 陈伟洲, 吴文婷, 许俊宾, 等. 不同生态因子对皱紫菜生长及生理组分的影响[J]. 南方水产科学, 2013, 9(2): 14-19. doi: 10.3969/j.issn.2095-0780.2013.02.003 [10] 胡凡光, 郭萍萍, 王娟, 等. 水温、盐度、pH和光照度对龙须菜生长的影响[J]. 渔业现代化, 2013, 40(4): 23-27. doi: 10.3969/j.issn.1007-9580.2013.04.005 [11] ENDOMA L F, NUÑAL S N, TRAIFALGAR R F M, et al. Photo-bleached agar extracts from Gracilariopsis heteroclada[J]. Bot Mar, 2020, 63(6): 559-569. doi: 10.1515/bot-2020-0028
[12] ELLE B J, CORRE JR V, FELARCA K G, et al. Potential of Gracilariopsis bailiniae and Oreochromis mossambicus in improving water quality in intensive Litopenaeus vannamei tank culture[J]. AACL Bioflux, 2017, 10(5): 1309-1318.
[13] HUANG B W, CUI J J, CHEN X Y, et al. Mechanism of the allelopathic effect of macroalgae Gracilaria bailiniae on Nitzschia closterium[J]. Ecotoxicol Environ Saf, 2022, 241: 113767. doi: 10.1016/j.ecoenv.2022.113767
[14] 徐聪, 徐日升, 黄博文, 等. 异枝江蓠与凡纳滨对虾室内零换水混养[J]. 广东海洋大学学报, 2021, 41(3): 131-137. doi: 10.3969/j.issn.1673-9159.2021.03.017 [15] 夏邦美. 中国海藻志第二卷: 红藻门. 第五册: 伊谷藻目 杉藻目 红皮藻目[M]. 北京: 科学出版社, 1999, 31-34. [16] 钟志海, 黄中坚, 陈伟洲. 不同环境因子对异枝江蓠的生长及生化组分的影响[J]. 渔业科学进展, 2014, 35(3): 98-104. doi: 10.11758/yykxjz.20140314 [17] YONG Y S, YONG W T L, ANTON A. Analysis of formulae for determination of seaweed growth rate[J]. J Appl Phycol, 2013, 25(6): 1831-1834. doi: 10.1007/s10811-013-0022-7
[18] WELLBURN, ALAN R. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution[J]. J Plant Physiol, 1994, 144(3): 307-313. doi: 10.1016/S0176-1617(11)81192-2
[19] BEER S, ESHEL A. Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae[J]. Mar Freshw Res, 1985, 36(6): 785-792. doi: 10.1071/MF9850785
[20] 姜宏波, 田相利, 董双林, 等. 温度和光照强度对鼠尾藻生长和生化组成的影响[J]. 应用生态学报, 2009, 20(1): 185-189. doi: 10.13287/j.1001-9332.2009.0015 [21] 郜晓峰, 刘炜, 钟逸云, 等. 不同温度对大叶藻生长与光合生理的影响[J]. 应用与环境生物学报, 2022, 28(1): 175-181. [22] 王义婧, 徐胜, 何兴元, 等. 美国薄荷 (Monarda didyma L.) 对大气增温的生理生态响应[J]. 生态环境学报, 2018, 27(12): 2217-2224. [23] MURCHIE E H, LAWSON T. Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications[J]. J Exp Bot, 2013, 64(13): 3983-3998. doi: 10.1093/jxb/ert208
[24] 张卫强, 黄芳芳, 甘先华, 等. 遮阴和盐分对银叶树幼苗光合特性与叶绿素荧光参数的影响[J]. 生态环境学报, 2020, 29(3): 438-446. [25] GOLTSEV V, KALAJI H, PAUNOV M, et al. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus[J]. Russ J Plant Physiol, 2016, 63(6): 869-893. doi: 10.1134/S1021443716050058
[26] MAXWELL K, JOHNSON G N. Chlorophyll fluorescence: a practical guide[J]. J Exp Bot, 2000, 51(345): 659-668. doi: 10.1093/jexbot/51.345.659
[27] MISHRA A N. Chlorophyll fluorescence: a practical approach to study ecophysiology of green plants[M]//SÁNCHEZ-MOREIRAS A M, REIGOSA M J. Advances in plant ecophysiology techniques. Cham: Springer International Publishing AG, 2018: 77-97. https://doi.org/10.1007/978-3-319-93233-05.
[28] 薛娴, 许会敏, 吴鸿洋, 等. 植物光合作用循环电子传递的研究进展[J]. 植物生理学报, 2017, 53(2): 145-158. doi: 10.13592/j.cnki.ppj.2016.0432 [29] 周伟, 武卉, 黄晶晶, 等. 高浓度CO2和光周期对浒苔幼苗生长和光合生理的影响[J]. 南方水产科学, 2022, 18(5): 30-38. doi: 10.12131/20210278 [30] 陈敏, 王宁, 杨多利, 等. 隐藻藻胆蛋白的结构与能量传递功能[J]. 植物生理学报, 2015, 51(12): 2070-2082. doi: 10.13592/j.cnki.ppj.2015.0382 [31] 张文文, 郭永坚, 李俊伟, 等. 营养盐对海萝生长和藻体生化成分的影响[J]. 南方水产科学, 2016, 12(2): 30-35. doi: 10.3969/j.issn.2095-0780.2016.02.005 [32] 丁兰平, 孙国栋, 黄冰心, 等. 温度和盐度对刺枝鱼栖苔 (Acanthophora spicifera) (红藻门,松节藻科) 生长及其几种光合色素的影响[J]. 海洋与湖沼, 2013, 44(4): 913-918. -
期刊类型引用(4)
1. 袁太平,廖宇琦,黄小华,胡昱,陶启友,李根,庞国良. 深水网箱网衣清洗装备关键部件设计及试验. 南方水产科学. 2024(01): 25-33 .  本站查看
本站查看
2. 魏盛军,蔡文鸿,丁兰,张哲,张艺,潘柏霖,陈思源,杨章武,魏观渊,林元俊,郑国富. 福建海上养殖设施污损生物处理方式现状调查. 渔业研究. 2023(02): 147-154 . 百度学术
3. 黄小华,庞国良,袁太平,胡昱,王绍敏,郭根喜,陶启友. 我国深远海网箱养殖工程与装备技术研究综述. 渔业科学进展. 2022(06): 121-131 . 百度学术
4. 廖宇琦,袁太平,胡昱,王绍敏,陶启友,黄小华. 转盘流道结构对网箱清洗装备喷嘴流场特性的影响. 渔业现代化. 2021(06): 9-18 . 百度学术
其他类型引用(3)
计量
- 文章访问数: 331
- HTML全文浏览量: 106
- PDF下载量: 45
- 被引次数: 7
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
