
Effects of BDE3 on SOD, MDA and GSH of Perna viridis
-
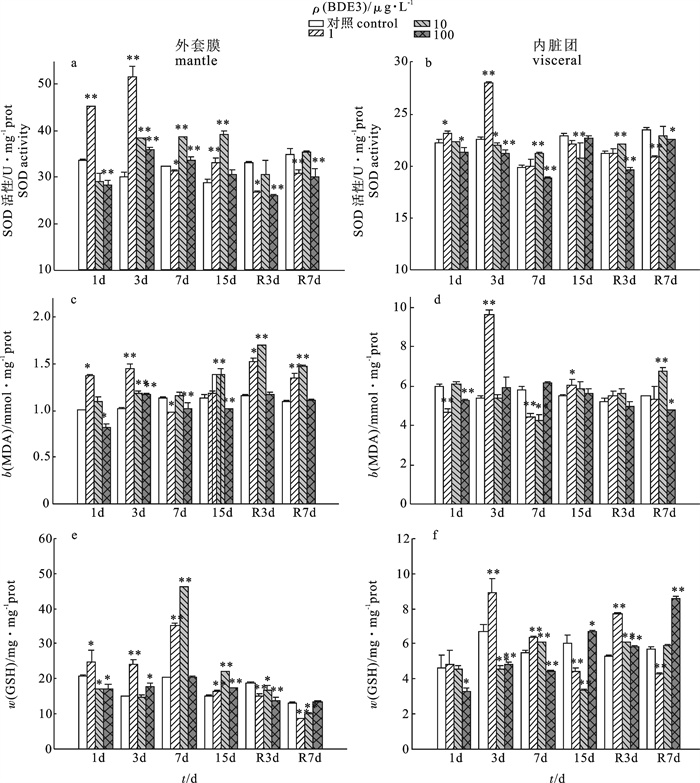
摘要:
采用半静态试验方法,研究了不同质量浓度的一溴联苯醚(BDE3)(1 μg · L-1、10 μg · L-1和100 μg · L-1)胁迫1 d、3 d、7 d和15 d且清洁海水释放3 d和7 d后,翡翠贻贝(Perna viridis)外套膜和内脏团超氧化物歧化酶(SOD)活性、丙二醛(MDA)质量摩尔浓度和谷胱甘肽(GSH)质量分数的变化规律。结果表明,BDE3胁迫后低浓度组翡翠贻贝两组织SOD活性均受到诱导(P < 0.01),诱导率随胁迫时间延长而下降,中、高浓度组外套膜对BDE3响应比内脏团灵敏;BDE3对翡翠贻贝外套膜b(MDA)的影响总体呈诱导后抑制趋势,对内脏团b(MDA)的影响表现抑制-诱导反复变化,其中低浓度组第3天时诱导率最高(26.04%),高浓度组第7天时抑制率最高(14.92%);翡翠贻贝外套膜w(GSH)在BDE3胁迫下,低浓度组一直被诱导(P < 0.01),中、高浓度组总体呈诱导后抑制作用,内脏团中高浓度组w(GSH)胁迫第1天受到显著抑制(P < 0.05),随暴露时间的延长,低浓度组w(GSH)显著增加(P < 0.01),中、高浓度组出现诱导抑制的不规律变化。释放阶段结束后仅个别浓度组恢复至对照组水平。
Abstract:To study the variation trends of superoxide dismutase (SOD) activity, malondialdehyde (MDA) level and glutathione (GSH) content in tissues of mantle and visceral mass of Perna viridis, we performed an experiment under semi-static conditions by exposing the samples to BDE3 (1 μg · L-1, 10 μg · L-1 and 100 μg · L-1) for 1 d, 3 d, 7 d and 15 d then releasing them by clean seawater for 3 d and 7 d. The results show that during the exposing period, SOD activities of the 2 tissues in 1 μg · L-1 group were induced very significantly (P < 0.01), and the induction rate decreased with exposing time; SOD activities of mantle in groups of 10 μg · L-1 and 100 μg · L-1 were more sensible than those of visceral mass. MDA level increased at first then inhibited later in mantle, while induction and inhibition alternated in visceral mass; for 1 μg · L-1 group, the highest induction rate reached 26.04% on 3rd day of exposing while the highest inhibitory rate reached 14.92% on 7th day. GSH content in mantle in 1 μg · L-1 group was induced significantly (P < 0.01) all the time, while it increased at first then inhibited later for groups of 10 μg · L-1 and 100 μg · L-1; in visceral mass, GSH content in 100 μg · L-1 group was induced significantly (P < 0.01) on 1st day, while that in 1 μg · L-1 group increased significantly with exposing time (P < 0.01);groups of 10 μg · L-1 and 100 μg · L-1 showed irregular trends of induction-inhibition. After the releasing period, only individual groups recovered to the same level in the control.
-
Keywords:
- BDE3 /
- Perna viridis /
- mantle /
- viscera mass /
- SOD /
- MDA /
- GSH
-
游泳能力对水生动物的生存能力有着巨大影响,而从力竭运动后状态恢复的能力反映了水生动物的持续极限运动能力。在野外环境中,力竭运动常出现在动物进行洄游、穿越激流、躲避天敌攻击和捕食等场景中;在人工养殖环境中,力竭运动常出现在动物抢食、逃避捕捞、同类相残、高密度暂养和运输等场景中[1-3]。对虾类具有较为出色的运动能力,其在低速游泳时使用游泳足摆动进行游泳,可适应较长距离的洄游活动;受到攻击或刺激时使用发达的腹部肌肉进行收缩,尾扇张开拨水,迅速向后弹跳逃离危险水域[4-5]。研究报道,鱼类和虾类在力竭运动时,无氧呼吸强度显著加强,无氧呼吸链的酶活性显著升高,有氧代谢活动被抑制[6-8];力竭运动结束后,肌肉中会积累乳酸,高浓度乳酸将会限制动物再次进行力竭运动的强度和持续运动的耐久力[9-12]。根据黑鲷 (Sparus macrocephlus) 幼鱼的血乳酸浓度恢复情况分析,黑鲷幼鱼在力竭运动后4 h即可再次进行力竭运动[13];斑马鱼 (Danio rerio) 在力竭运动后第8小时肌乳酸下降恢复至对照组水平,虽然肌糖原未恢复至对照组水平,但已能进行力竭运动[14],可见动物体内乳酸水平是限制动物重复进行力竭运动的关键因素。
对虾类在运动疲劳时,会出现肝、肌糖原浓度均显著下降、血糖和血乳酸浓度显著升高的现象[15-16],属于“乳酸释放型”动物。对虾类的内环境平衡比鱼类的更脆弱,失衡后恢复所需的时间亦更长[17-18]。凡纳滨对虾 (Litopenaeus vannamei) 和中国明对虾 (Fenneropenaeus chinensis) 虾苗在运动频率显著上升的背景下,出现生长率显著降低、免疫能力低下和死亡率升高的现象[19-20];凡纳滨对虾在弹跳运动中,体内蛋白质代谢供能、无氧和有氧代谢强度均显著提升[21],证明过量运动会对虾类的代谢健康造成强烈影响。斑节对虾 (Penaeus monodon) 隶属软甲纲、十足目、对虾科、对虾属,俗名草虾、黑虎虾等,具有适应性强、个体大、生长速度快等优点,是中国沿海重要的经济养殖和增殖放流品种[22]。在目前虾类增养殖业快速发展的背景下,虽然已经有关于虾类运动前后代谢变化的报道,但虾类力竭运动后的恢复代谢特征与调节机制尚不明晰。因此,本研究以斑节对虾为对象,在实验室水槽模拟斑节对虾幼虾力竭运动情况,探究其在力竭运动后肝胰腺与腹部肌肉中的代谢能力相关指标的恢复调节过程,以期为斑节对虾养殖生产和增殖放流提供科学参考。
1. 材料与方法
1.1 实验材料
实验使用的斑节对虾幼苗为中国水产科学研究院南海水产研究所深圳试验基地自行繁育的养殖虾苗。虾苗体长为0.5~0.9 cm时放入室外池塘中放养标粗3周,标粗后挑选体长为 (3.0±0.4) cm、体质量为 (0.2±0.07) g的约1 000尾规格相近的斑节对虾幼虾进行力竭运动胁迫。在力竭运动后的24 h内禁食,在之后的恢复阶段每日共投喂2.25 g (全部虾苗总质量的3%) 的斑节对虾配合0号饲料,分3次 (8:00、16:00、23:00) 投喂。力竭运动实验装置为自行设计的环形水道 (外径为1 004 mm,内置隔离装置直径为 604 mm,内置隔离装置高为395 mm,容量为200 L)。
1.2 实验方法
1.2.1 力竭状态幼虾组织样品收集
从育肥池中随机挑选30尾幼虾为一组,设3组平行,进行力竭运动胁迫。将幼虾放入环形水道中进行胁迫实验,期间水道中流速控制在12 cm·s−1 (在预实验时测得幼虾在该流速下可顶水游泳1.5 h),用小抄网从幼虾游泳方向前方捕捞攻击幼虾,使幼虾持续进行跳跃逃避运动,根据已有报道,当幼虾对刺激无法进行反应、游泳足摆动频率下降、无法再逆水流游泳的时候即进入力竭状态[21]。捕捞刺激持续约1 h,当大部分幼虾陷入力竭状态时,在其中所有进入力竭状态的幼虾中随机挑选12尾置于冰冷的取样盘中取样,收集其肝胰腺与腹部肌肉进行之后的指标测定,为力竭运动后第0小时的样品。取样后立即置入液氮中急冻,之后于−80 ℃保存。
1.2.2 恢复过程中幼虾组织收集
按恢复时间点 (第1、第4、第8、第12、第24、第48、第72小时) 设立7个处理组,每组设3组平行,每组约30尾。以1.2.1中方法使每组幼虾陷入力竭状态,将每组全部存活的力竭幼虾放入室内恢复池的网箱中,同时开始计算恢复时间,在设立的8个时间点取样,收集幼虾的肝胰腺与肌肉。实验水体信息见表1。
表 1 实验水体的物理化学参数Table 1. Physical and chemical parameters of experimental water body水体
Water body温度
Temperature/℃盐度
SalinitypH 溶解氧
Dissolved oxygen/(mg·L−1)育肥池 Fattening pond 25±1 23±2 8.0±0.3 4.2±0.5 环形水道 Circular waterway 25 23 7.8 4.5 恢复池 Recovering pond 25±1 23 7.8±0.3 5.6±0.5 1.2.3 对照组、空白组样品收集
力竭实验前从育肥池中随机挑选12尾幼虾,收集幼虾肝胰腺与肌肉组织作为空白组样品。在8个恢复时间点收集处理组样品前,在育肥池中随机选择12尾幼虾以收集其肝胰腺与肌肉组织,作为各个取样时间点的对照组样品。
1.2.4 样品生化指标测定
在所有样品采集完成后于实验室 (26 ℃) 测定肝胰腺和肌肉的储能物质含量:总蛋白质 (TP)、乳酸 (LA)、糖原 (GLY);呼吸代谢酶活指标:琥珀酸脱氢酶 (SDH) 活性、乳酸脱氢酶 (LDH) 活性;氨基酸代谢酶活指标:谷草转氨酶 (AST) 活性和谷丙转氨酶 (ALT) 活性。以上指标的测定均使用南京建成生物工程研究所的试剂盒。
1.3 数据处理
实验数据经Excel 2016软件处理后,使用SPSS 21.0软件进行数据分析,以力竭运动后的恢复时间为自变量、以取样时间序列排序的各项生理指标为因变量进行单因素方差分析 (One-Way ANOVA),若组间出现极显著 (P<0.01) 或显著差异 (P<0.05) 时,则用Duncan法进行组间差异显著性的多重比较,数据以“平均值±标准差 (
$\overline X \pm {\rm{SD}} $ )”表示。2. 结果
2.1 力竭运动后不同恢复时间对斑节对虾幼虾代谢能力相关指标的影响
根据表2,斑节对虾幼虾在力竭运动后,随恢复时间的延长,腹部肌肉中GLY、LA和LDH活性存在显著差异 (P<0.05),其中LDH活性差异极显著 (P<0.01)。斑节对虾幼虾在力竭运动后,随恢复时间的延长,肝胰腺中LA差异显著 (P<0.05);SDH、LDH、ALT和AST活性均有极显著差异 (P<0.01)。
表 2 斑节对虾幼虾在力竭运动后8个恢复时间点和空白组腹部肌肉、肝胰腺中代谢指标单因素分析Table 2. Analysis of metabolic capacity in abdominal muscles (M) and heaptopancreas (H) of juvenile P. monodon following strenuous activity by One-Way ANOVA因变量
Dependent variable自由度
df均方
MSF P 肌乳酸质量浓度 LA mass concentraion (M) 8 0.000 19 2.613 0.043 肝乳酸质量浓度 LA mass concentraion (H) 8 0.003 3.082 0.023 肌总蛋白质量分数 TP mass fraction (M) 8 3.152 1.294 0.307 肝总蛋白质量分数 TP mass fraction (H) 8 9.508 12.283 0.000 肌糖原质量分数 GLY mass fraction (M) 8 10.057 2.326 0.045 肝糖原质量分数 GLY mass fraction\ (H) 8 0.582 1.305 0.302 肌SDH活性 SDH activity (M) 8 0.007 0.491 0.847 肝SDH活性 SDH activity (H) 8 21.175 22.601 0.000 肌LDH活性 LDH activity (M) 8 0.232 6.318 0.001 肝LDH活性 LDH activity (H) 8 0.000 29 8.599 0.000 肌ALT活性 ALT activity (M) 8 0.937 2.151 0.085 肝ALT活性 ALT activity (H) 8 0.502 6.380 0.001 肌AST活性 AST activity (M) 8 0.560 0.943 0.507 肝AST活性 AST activity (H) 8 75.490 227.247 0.000 2.2 力竭运动后幼虾的代谢指标随恢复时间延长的变化
2.2.1 TP变化
斑节对虾幼虾肝胰腺TP水平在力竭运动前后无显著差异,随恢复时间延长,肝胰腺中TP水平在第4小时达到顶峰,显著高于对照组 (P<0.05),第4至第12小时间显著下降至低于对照组水平 (P<0.05),第48小时恢复至对照组水平 (图1-A)。腹部肌肉中的TP水平在力竭运动前后和之后的恢复中无显著变化 (图1-B)。
![]() 图 1 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中总蛋白质量分数变化不同字母代表各恢复时间点实验组间存在显著差异(P<0.05);“*”表示实验组和对照组在胁迫后同一时间点差异显著(P<0.05),后图同此Figure 1. Total protein mass fractions in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activityDifferent letters represent significant differences between the experimental groups at different recovery time (P<0.05); *. Significant differences between experimental group and the control group at the same time (P<0.05). The same case in the following figures
图 1 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中总蛋白质量分数变化不同字母代表各恢复时间点实验组间存在显著差异(P<0.05);“*”表示实验组和对照组在胁迫后同一时间点差异显著(P<0.05),后图同此Figure 1. Total protein mass fractions in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activityDifferent letters represent significant differences between the experimental groups at different recovery time (P<0.05); *. Significant differences between experimental group and the control group at the same time (P<0.05). The same case in the following figures2.2.2 GLY变化
斑节对虾幼虾肝胰腺GLY水平在整个实验过程中并未出现显著差异(图2-A)。腹部肌肉GLY水平在力竭运动后出现显著下降 (P<0.05),第0至第4小时保持在低于对照组水平。腹部肌肉GLY水平在第1小时后开始恢复,保持上升,第8小时恢复至对照组水平,第72小时显著高于对照组水平 (P<0.05,图2-B)。
![]() 图 2 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中糖原质量分数变化Figure 2. Glycogen mass fractions in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 2 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中糖原质量分数变化Figure 2. Glycogen mass fractions in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity2.2.3 LA变化
力竭运动后斑节对虾幼虾肝胰腺中LA水平无显著变化,腹部肌肉LA水平显著上升 (P<0.05)。肝胰腺LA水平随恢复时间延长逐渐上升,第12小时显著高于对照组并达到顶峰 (P<0.05),第12至第24小时内显著回落,第72小时恢复至对照组水平 (图3-A)。腹部肌肉LA水平在第8小时恢复至对照组水平,但在第8至第12小时再次显著升高 (P<0.05),第24小时恢复至对照组水平 (图3-B)。
![]() 图 3 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中乳酸质量摩尔浓度变化Figure 3. Lactic acid molar concentrations in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 3 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中乳酸质量摩尔浓度变化Figure 3. Lactic acid molar concentrations in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity2.2.4 AST活性变化
斑节对虾幼虾肝胰腺中AST活性力竭运动前后无显著差异,第4至第12小时内显著上升 (P<0.05),第12至第72小时下降至对照组水平(图4-A)。腹部肌肉中的AST活性在力竭运动前后无显著差异,第0至第48小时内维持在对照组水平,但在第48至第72小时显著下降 (P<0.05,图4-B)。
![]() 图 4 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中谷草转氨酶活性变化Figure 4. Aspartate aminotransferase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 4 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中谷草转氨酶活性变化Figure 4. Aspartate aminotransferase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity2.2.5 ALT活性变化
斑节对虾幼虾肝胰腺中ALT活性力竭运动前后无显著差异,第4至第12小时内显著上升 (P<0.01),第12至第48小时下降至对照组水平(图5-A)。腹部肌肉中ALT活性在力竭运动后持续下降,第12小时时显著低于对照组水平 (P<0.05),在之后的恢复过程中保持较低水平,第72小时时仍显著低于对照组水平 (图5-B)。
![]() 图 5 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中谷丙转氨酶活性变化Figure 5. Alanine aminotransferase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 5 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中谷丙转氨酶活性变化Figure 5. Alanine aminotransferase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity2.2.6 LDH活性变化
斑节对虾幼虾肝胰腺中LDH活性在力竭运动前后无显著差异,第2至第12小时显著上升 (P<0.05),第12至第72小时降低至对照组水平 (P<0.05,图6-A)。腹部肌肉的LDH活性在力竭运动后出现显著升高 (P<0.05),第0至第1小时短暂下降后在第1至第12小时内继续显著升高,并在第12至第72小时内仍保持在较高水平 (P<0.05,图6-B)。
![]() 图 6 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中乳酸脱氢酶活性变化Figure 6. Lactate dehydrogenase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 6 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中乳酸脱氢酶活性变化Figure 6. Lactate dehydrogenase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity2.2.7 SDH活性变化
斑节对虾幼虾肝胰腺中SDH活性在力竭运动后升高,但无显著差异,第4小时显著低于第0小时水平 (P<0.05),第4至第12小时间显著升高,第24小时恢复至对照组水平(图7-A)。虾腹部肌肉力竭运动后的SDH活性显著降低,低于对照组水平,第12小时恢复至对照组水平,之后再次下降,第72小时仍显著低于对照组水平 (图7-B)。
![]() 图 7 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中琥珀酸脱氢酶活性变化Figure 7. Succinate dehydrogenase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity
图 7 斑节对虾幼虾力竭运动后肝胰腺 (A) 与肌肉 (B) 中琥珀酸脱氢酶活性变化Figure 7. Succinate dehydrogenase activities in hepatopancreas (A) and muscle (B) of juvenile P. monodon following strenuous activity3. 讨论
3.1 斑节对虾幼虾力竭运动后体内乳酸和糖原的变化
肌肉对乳酸的耐受性与糖原储备量被认为是限制动物进行力竭运动的关键因素,而乳酸代谢能力与糖原恢复能力则是限制动物重复力竭运动的关键因素[1]。具有出色游泳能力的虹鳟 (Oncorhynchus mykiss) 恢复时间短于瓦氏黄颡鱼(Pelteobagrus vachelli)等不喜游泳的鱼类[23],虹鳟和斑马鱼 (Danio rerio) 对乳酸的耐受能力也显著高于不喜游泳的瓦氏黄颡鱼和鲇 (Silurus asotus)[14, 24],可见乳酸清除和糖原恢复时间越短的水生动物游泳能力越强[24]。虾类力竭运动性能显著低于鱼类[19],在比较前人对不同体型对虾跳跃能力的研究中发现,体质量越大、体型越大的虾类跳跃距离越远[19],可认为体型大的成虾逃逸时所需的跳跃次数显著少于体型小的幼虾。
本研究中幼虾腹肌GLY、LA在力竭运动后第8小时即恢复至对照组水平,且肌GLY在恢复后第72小时仍显著高于对照组,力竭后的肌LA上升幅度、清除时间和GLY均高于体型较大的日本囊对虾 (Penaeus japonicus) 和凡纳滨对虾[5, 19],因此可认为幼虾耐受力竭的能力强于成虾,从力竭状态中恢复所需时间亦短于成虾。推测由于幼虾跳跃距离较短,需要更多次跳跃以逃离危险,因此腹部肌肉的糖原储备、乳酸耐受力适应性高于成虾。
研究表明,动物乳酸清除代谢有彻底氧化供能、合成葡萄糖和合成肌糖原3种途径,其中氧化供能的场所在肝胰腺,合成糖原和葡萄糖则在肌肉[25-26]。对虾属于乳酸释放型,力竭运动后通过血淋巴转运肌乳酸到其他组织进一步氧化分解[9, 21]。前人发现肝胰腺是水生动物清除体内乳酸的主要场所[27-28],本研究中幼虾肝LA水平在恢复阶段先升后降,说明幼虾肝胰腺承担清除乳酸的工作,保证机体在力竭运动后的内环境稳定;而肌LA在恢复后第8至第12小时再次上升,可能是肌肉组织将血乳酸重新摄取以合成糖原[29],但虾类体内是否存在此代谢途径还有待验证。
3.2 斑节对虾幼虾力竭运动后组织内转氨酶活性与蛋白质含量变化
谷丙转氨酶 (ALT) 和谷草转氨酶 (AST) 广泛存在于虾类的肝胰腺与心肌中,其活性可作为指示氨基酸代谢强度和肝功能是否健全的生理指标。两种酶联合介导的氨基酸转氨作用是细胞内糖异生作用的关键,在能源物质匮乏的情况下起到填补能量缺口的功能[30-31]。蛋白质作为生命活动的承担者,参与机体的绝大部分活动。
本研究中肌AST、ALT活性和TP水平在力竭运动前后并无显著变化,在恢复过程中AST、ALT活性持续降低,第72小时显著低于对照组水平,说明肌肉中氨基酸代谢被显著抑制,氨基酸并没有被作为能量物质而被分解供能。在恢复前4 h幼虾肝TP显著上升,转氨酶活性较低,可认为此阶段肝胰腺主要工作是合成蛋白。在恢复后第4小时转氨酶活性显著上升,推测是由于未补充外源营养,机体激活氨基酸代谢通路,通过脱氨作用产生丙酮酸和草酰乙酸供能或进入糖异生通路,进行代谢工作或补充能量缺口,与饥饿状态时的能量动员机制相似[31]。
虾类陷入缺氧状态时肝胰腺会合成并释放大量血蓝蛋白进入血淋巴中,以提高机体氧容量[32-34]。推测释放血蓝蛋白与糖异生作用是肝TP在第4至第8小时内显著下降的主要原因。第24小时开始摄入外源营养后,肝TP逐渐恢复到对照组水平。本研究中幼虾在恢复阶段的前24 h禁食,模拟幼虾在放流时遇到的极端情况,在幼虾进行剧烈运动后若无足够的能量摄入,其肝胰腺可在约24 h内通过氨基酸代谢供能以应对代谢能量不足的困境。
3.3 斑节对虾幼虾力竭运动后组织内呼吸代谢变化
呼吸作用是生物消耗能源物质,提供生命活动所需能量的唯一途径,各种代谢相关酶活性反映了呼吸代谢的强度[1-2]。LDH与SDH分别为无氧和有氧呼吸的关键酶,其活性标志了该组织进行无氧与有氧呼吸强度的高低,在氧充足环境中,LDH还能将乳酸逆转化为丙酮酸参与有氧呼吸,实现乳酸彻底分解[35-36]。鱼虾类在力竭运动或较长时间的持续运动后,肌肉中的无氧呼吸强度和能量动员能力显著升高,而生长率和储能物质则显著下降[37-38]。本研究中幼虾腹肌在陷入力竭时LDH活性显著升高,SDH活性显著降低,说明幼虾剧烈运动时肌肉中产生能量较快的无氧呼吸显著增强,而有氧呼吸作用被抑制。肝胰腺LDH与SDH活性在力竭运动前后无显著变化,但SDH活性在第1至第4小时内显著下降,LDH活性则显著上升,出现了与已有报道一致的对虾缺氧时的代谢特征[32]。
但本研究所有过程中幼虾均处于富氧环境,但幼虾在恢复前8 h内仍有陷入缺氧状态的迹象。Savant等[39]研究发现,持续的低pH胁迫会使泥蟹 (Ilyoplax gangetica) 的耗氧率下降;陶易凡等[40]研究发现,克氏原螯虾 (Procambarus clarkii) 在外源性低pH胁迫时,鳃细胞氧交换效率下降,导致机体陷入缺氧状态。推测由于对虾属于乳酸释放型动物[9-10],肌肉在运动后将乳酸释放至血淋巴,导致血淋巴pH下降,摄氧能力下降,使得幼虾在恢复的前4 h内氧交换能力下降并进入缺氧状态。在此阶段中,幼虾机体的能量主要由无氧呼吸提供,代谢平衡极其脆弱,再次出现胁迫或将导致幼虾猝死。
恢复后第8小时,幼虾肌肉中的LDH活性显著高于力竭运动后的水平,SDH活性在前12 h恢复至对照组水平时不再升高,之后下降。可能是由于肌肉组织将乳酸转化为丙酮酸重新利用,同时抑制有氧呼吸,使肌糖原过量恢复。肝胰腺在前24 h内SDH、LDH活性均显著升高,但糖原并无减少。说明幼虾肝胰腺通过转化乳酸、氨基酸参与三羧酸循环以完成清除力竭运动代谢产物的目的,与凡纳滨对虾经历无水运输后的恢复规律吻合[32]。
4. 小结
在不同恢复时间点上,斑节对虾幼虾肝胰腺中各因子指标均差异显著,而肌肉中仅呼吸相关指标差异显著。幼虾机体在力竭运动后恢复的前4 h内,由于内源性pH变化,导致机体摄氧能力降低,在肝胰腺中有氧呼吸强度显著升高的情况下,致使肌肉有氧呼吸被抑制。幼虾为了适应下次力竭运动作出调节:增强无氧、抑制有氧呼吸抑制肌糖原消耗,并通过糖异生作用进行储能,证明力竭运动训练可提高幼虾力竭刺激的生理适应性,但是否能提高其游泳能力还有待验证。
力竭运动会对虾类机体产生久远影响,其产生的刺激在幼虾经历72 h后仍使机体代谢平衡存在较显著差异。在斑节对虾的增养殖工作中,在经济条件允许的范围内可将幼虾尽量培养至较大体型,并对其进行一定强度的运动训练,以48~72 h为周期进行力竭训练,可有效提高幼虾对力竭刺激的生理适应性,有利于提高其养殖及野外的成活率;在进行操作前应加强幼虾的营养尤其是维生素B6和抗应激剂等补剂,使其体内有足够的能量储备和抗氧化能力以应对刺激。
-
[1] SJODIN A, PATTERSON D G Jr., BERGMAN A. A review on human exposure to brominated flame retardants-particularly polybrominated diphenylethers[J]. Environ Int, 2003, 29(6): 829-839. doi: 10.1016/S0160-4120(03)00108-9
[2] ANDERSSON O, BLOMKVIST G. Polybrominated aromatic pollutants found in fish in Sweden[J]. Chemosphere, 1981, 10(9): 1051-1060. doi: 10.1016/0045-6535(81)90216-2
[3] 马新东. 海洋沉积物和生物中多溴联苯醚的方法研究及应用[D]. 大连: 大连海事大学, 2007. 10.7666/d.y1037142 MA Xindong. The method research and application of polybrominated diphenyl ethers in marine sediment and organism[D]. Dalian: Dalian Maritime University, 2007. (in Chinese) 10.7666/d.y1037142
[4] BETTS K. Does a key PBDE break down in the environment?[J]. Environ Sci Technol, 2008, 42(18): 6781. doi: 10.1021/es8018463
[5] BRANCHI I, CAPONE F, VITALONE A, et al. Early developmental exposure to BDE 99 or Aroclor 1254 affects neurobehavioural profile: interference from the administration route[J]. Neuro Toxicol, 2005, 26(2): 183-192. doi: 10.1016/j.neuro.2004.11.005
[6] DARNERUD P O, RISBERG S. Tissue localisation of tetra-and pentabromodiphenyl ether congeners (BDE-47, -85, and-99) in perinatal and adult C57BL mice[J]. Chemosphere, 2006, 62(3): 485-493. doi: 10.1016/j.chemosphere.2005.04.004
[7] DARNERUD P O, WONG J, BERGMAN A, et al. Common viral infection affects pentabrominated diphenyl ether(PBDE) distribution and metabolic and hormonal activities in mice[J]. Toxicology, 2005, 210(2/3): 159-167. doi: 10.1016/j.tox.2005.01.015
[8] 任金亮, 王平. 多溴联苯醚环境行为的特征与研究进展[J]. 化工进展, 2006, 25(10): 1152-1157. doi: 10.3321/j.issn:1000-6613.2006.10.006 REN Jinliang, WANG Ping. Progress and characteristics of environmental behavior of polybrominated diphenyl ethers[J]. Chem Ind Eng Prog, 2006, 25(10): 1152-1157. (in Chinese) doi: 10.3321/j.issn:1000-6613.2006.10.006
[9] 孟范平, 李卓娜. 多溴联苯醚(PBDEs)在海洋环境中的行为研究进展[J]. 中国海洋大学学报, 2006, 34(5): 25-27. doi: 10.3969/j.issn.1672-5174.2009.02.036 MENG Fanping, LI Zhuona. Research progress on the behavior of polybrominated diphenyl ethers (PBDEs) in marine environment[J]. Periodic Ocean Univ China, 2006, 34(5): 25-27. (in Chinese) doi: 10.3969/j.issn.1672-5174.2009.02.036
[10] 唐学玺, 柯可, 王悠. 多溴联苯醚在海洋环境中的分布及其对海洋生物的影响研究[J]. 海洋湖沼通报, 2010(1): 36-46. https://www.doc88.com/p-9975156623532.html TANG Xuexi, KE Ke, WANG You. Effects of anthracene on activity of superoxide dismutase in Sebastodes fuscescens[J]. Trans Oceanol Limnol, 2010(1): 36-46. (in Chinese) https://www.doc88.com/p-9975156623532.html
[11] STEBBING A R D. Tolerance and hormesis-increased resistance to copper in hydroids linked to hormesis[J]. Mar Environ Res, 2002, 54(3/4/5): 805- 809. doi: 10.1016/S0141-1136(02)00119-8
[12] 王辅明, 朱祥伟, 马永鹏, 等. 低浓度五氯酚暴露对稀有鲫体内SOD活性、GSH和HSP70含量的影响[J]. 生态毒理学报, 2009, 4(3): 415-421. https://cstj.cqvip.com/Qikan/Article/Detail?id=31139018&from=Qikan_Article_Detail WANG Fuming, ZHU Xiangwei, MA Yongpeng, et al. Effects of low concentration of pentachlorophenol exposure on SOD activity, GSH and HSP70 content in rare minnow[J]. Asian J Ecotoxicol, 2009, 4(3): 415-421. (in Chinese) https://cstj.cqvip.com/Qikan/Article/Detail?id=31139018&from=Qikan_Article_Detail
[13] 秦洁芳, 陈海刚, 蔡文贵, 等. 邻苯二甲酸二丁酯对翡翠贻贝抗氧化酶及脂质过氧化水平的影响[J]. 应用生态学报, 2011, 22(7): 1878-1884. https://www.cjae.net/CN/Y2011/V22/I07/1878 QIN Jiefang, CHEN Haigang, CAI Wengui, et al. Effects of di-n-butyl phthalate on the antioxidant enzyme activities and lipid peroxidation level of Perna viridis[J]. Chin J Appl Ecol, 2011, 22(7): 1878-1884. (in Chinese) https://www.cjae.net/CN/Y2011/V22/I07/1878
[14] 杨涛, 陈海刚, 蔡文贵, 等. 菲和苯并(b)荧蒽曝露对翡翠贻贝外套膜的氧化胁迫及损伤[J]. 南方水产科学, 2011, 7(4): 24-29. doi: 10.3969/j.issn.2095-0780.2011.04.004 YANG Tao, CHEN Haigang, CAI Wengui, et al. Oxidative stress and damage of by exposure to phenanthrene and benzo (b) fluoranthene[J]. South China Fish Sci, 2011, 7(4): 24-29. (in Chinese) doi: 10.3969/j.issn.2095-0780.2011.04.004
[15] DARNERUD P O, ERIKSEN G S. Polybrominated diphenyl ethers: occurrence, dietary exposure, and toxicology[J]. Environ Health Persp, 2001, 109(Suppl 1): 49-68. doi: 10.1289/ehp.01109s149
[16] 陈海刚, 马胜伟, 林钦, 等. 氯化三丁基锡对黑鲷鳃和肝组织SOD、MDA和GPx的影响[J]. 南方水产, 2009, 5(2): 23-27. doi: 10.3969/j.issn.1673-2227.2009.02.004 CHEN Haigang, MA Shengwei, LIN Qin, et al. Effects of tributyltin chloride (TBTCl) on SOD activities, MDA contents and GPx activities in gill and liver of the black porgy (Sparus macrocephalus)[J]. South China Fish Sci, 2009, 5(2): 23-27. (in Chinese) doi: 10.3969/j.issn.1673-2227.2009.02.004
[17] 向金梅, 郭定宗, 杨世锦. N-乙酰半胱氨酸对砷致小鼠脂质过氧化损伤的保护作用研究[D]. 武汉: 华中农业大学, 2006. https://www.doc88.com/p-1092203371611.html XIANG Jinmei, GUO Dingzong, YANG Shijin. Study on the protective effect of N-acetyl-cysteine and VitE on lipid peroxidation damage induced by arsenic in rat[D]. Wuhan: Huazhong Agricultural University, 2006. (in Chinese) https://www.doc88.com/p-1092203371611.html
[18] 冯涛, 郑微云. 苯并(a)芘对大弹涂鱼肝脏还原型谷胱甘肽含量的影响[J]. 厦门大学学报: 自然科学版, 2001, 40(5): 1095-1099. doi: 10.3321/j.issn:0438-0479.2001.05.017 FENG Tao, ZHENG Weiyun. Effect of benzo(a) pyrene on content of reduced-glutathione in the liver of Boleophthalmus pectinirostris[J]. J Xiamen Univ: Natural Science, 2001, 40(5): 1095-1099. (in Chinese) doi: 10.3321/j.issn:0438-0479.2001.05.017
[19] 吕杨, 王立宁, 黄俊, 等. 海河渤海湾地区沉积物、鱼体样品中多溴联苯醚的水平与分布[J]. 环境污染与防治, 2007, 29(9): 652-660. doi: 10.3969/j.issn.1001-3865.2007.09.004 LV Yang, WANG Lining, HUANG Jun, et al. PBDEs in sediments and crucians of Haihe River and Bohai Bay[J]. Environ Pollut & Control, 2007, 29(9): 652-660. (in Chinese) doi: 10.3969/j.issn.1001-3865.2007.09.004
-
期刊类型引用(1)
1. 高杰,郭华阳,刘明鉴,刘宝锁,朱克诚,张楠,郭梁,张殿昌. 无乳链球菌感染对卵形鲳鲹脾脏的损伤及细胞凋亡相关基因表达的影响. 南方水产科学. 2022(06): 44-51 .  本站查看
本站查看
其他类型引用(2)
 下载:
下载:








计量
- 文章访问数: 3663
- HTML全文浏览量: 153
- PDF下载量: 2396
- 被引次数: 3
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
