
Effects of different conditions on feeding biofloc of red swamp crayfish (Procambarus clarkii ) juveniles
-
摘要: 生物絮凝技术可以通过在养殖水体中调节碳氮比而维持良好水质,促进养殖品种健康生长,产生的生物蛋白可实现蛋白的多级利用,节约经济成本。实验从摄食环境(曝气、无曝气、水草)、摄食时间以及生物絮凝质量浓度等角度,探讨了克氏原螯虾 (Procambarus clarkii) 幼虾摄食絮凝的适宜条件。结果显示,絮凝质量浓度为500~600 mg·L–1可满足幼虾的摄食需求,并在摄食4 h之后幼虾肠道达饱和状态。相同摄食时间,絮凝曝气组幼虾摄食量最大,显著高于水草组(P<0.05),但与无曝气组差异不显著(P>0.05),比较幼虾肠道的充塞度,可见曝气组在摄食4 h后肠道食糜较多,肠道更为饱满,优于其他各组。综合生物絮凝培养条件、水化指标以及幼虾的摄食情况,认为生物絮凝技术应用到克氏原螯虾的养殖具有可行性。Abstract: Biofloc technology can maintain good water quality by adjusting the ratio of carbon to nitrogen in the aquaculture water, ensure the health of the cultured species, promote the multilevel utilization of microbial protein and save the cost of aqua-feed. We determined the suitable biofloc conditions for the red swamp crayfish (Procambarusclarkii) juveniles feeding by studying the feeding environment (aeration, no aeration and aquatic plants), feeding duration and biofloc concentration. The results show that 500–600 mg·L–1 biofloc could meet the demand of juveniles, and the intestines of the fed juveniles were saturated after 4-hour feeding. With the same feeding duration, the juveniles from the group with aeration had the highest food intake, higher than the group with aquatic plants significantly (P<0.05), but no obvious difference was found in comparison with the non-aerated group (P>0.05). Comparing the fullness rank of intestine in each group, the group with aeration had more chyme after 4-hour feeding. Considering the culture conditions of biofloc, hydrochemical index as well as the feeding situation, it is feasible to apply biofloc technology to the culture of red swamp crayfish.
-
Keywords:
- Procambarus clarkii /
- biofloc technology /
- juvenile shrimp /
- feeding
-
海刀豆(Canavalia maritima)属于豆科刀豆属,在中国沿海滩涂有广泛分布,其主要成分为黄酮、黄烷类、甾醇类化合物[1-3]。总黄酮广泛存在于各种植物和大型真菌中,具有多种生物活性[4]。近年来,槲皮素、芦丁等物质用于临床使得总黄酮的研究日益活跃,研究发现相当一部分总黄酮具有显著的生理及药理活性,例如抗氧化、抗病毒、抗肿瘤、抗炎、抗抑郁、抗焦虑、调节血管渗透性、改善记忆、中枢抑制、神经保护等功能,这些生理和药理特性使其广泛应用于食品、医药等领域[5-6]。加速溶剂萃取是利用升高温度和压力来增加物质溶解度与溶质扩散速率,从而提高萃取率的一种样品前处理方法[7-8]。ASE具有比传统萃取方法耗时短、使用溶剂量低、自动化高的优势[9],现已成为较好的样品前处理方法之一,广泛应用于食品、环境、药物等行业。响应面分析法(response surface methodology,RSM)是以多元二次回归方程式的形式将多因子试验中因素与指标的相互关系近似拟合,依此对函数的响应面和等高线进行分析,研究因子与响应值之间、因子与因子之间的相互关系[10-11]。与过去广泛使用的“正交试验设计法”不同的是, RSM具有试验周期短,求得的回归方程精度高,能研究几种因素间交互作用等优点[12-13]。
目前,中国对海刀豆中总黄酮的提取工艺研究较少,其他原料中总黄酮的提取多采用机械振荡法[14]、微波辅助萃取法[15]、超声提取法[16]、索氏提取法[17]等。笔者研究采用加速溶剂萃取法提取海刀豆中总黄酮,并采用响应面分析法对其工艺条件进行了优化,为之后海刀豆中总黄酮的分离纯化及其性质的研究奠定了基础。
1. 材料与方法
1.1 材料与试剂
海刀豆(采于海南沿海滩涂);硅藻土,芦丁标准品(广州市奇云生物技术有限公司出品);甲醇为分析纯;试验用水均符合GB/T 6682一级水的标准。
1.2 仪器
ASE300加速溶剂萃取仪(美国Dionex公司出品);分析天平(瑞士Mettler-Toledo公司出品);紫外-可见分光光度计(上海元析仪器有限公司出品);超声波清洗仪(德国Elma公司出品);高速万能粉碎机(天津市泰斯特仪器有限公司出品)。
1.3 试验方法
1.3.1 总黄酮提取率的测定
1) 配制芦丁标准溶液。准确称取充分干燥后的芦丁标准品5.00 mg,用30%的乙醇溶解,并定容至25 mL,摇匀,即得到质量浓度为0.2 mg ·mL-1的标准溶液。
2) 标准曲线的建立[18]。准确吸芦丁标准液0、1 mL、2 mL、3 mL、4 mL、5 mL和6 mL,分别置于25 mL容量瓶中,各加入5%亚硝酸钠(NaNO2)溶液1 mL,摇匀,放置6 min。加入10%硝酸铝[Al(NO3)3]溶液1 mL,摇匀,放置6 min,加入10 mL 5%氢氧化钠(NaOH)溶液,用30%乙醇稀释至刻度,摇匀放置10 min,以空白溶液为参比,用分光光度计在510 nm处测定吸光值(OD)。作芦丁浓度-吸光度标准曲线,得回归方程。
3) 海刀豆总黄酮含量的测定。经过单因素及响应面试验分析得出ASE提取海刀豆中总黄酮的最优条件为萃取压力10.3 MPa,萃取温度130 ℃,萃取时间16 min,甲醇体积分数66%,循环次数2次,且在该条件下多次提取后分别按照芦丁标准曲线测定法测定吸光值,直至吸光值稳定不变代表总黄酮提取完全,代入回归方程得出质量浓度,计算出各提取液中总黄酮的质量;多次提取出的总黄酮质量之和即为海刀豆中总黄酮的总质量M。
将各试验所得提取物用30%的乙醇溶解并定容于50 mL容量瓶中,取0.5 mL移入25 mL容量瓶中,按照芦丁标准曲线测定法测定吸光度,代入回归方程得出质量浓度C,计算出提取率[19]。
$$ \begin{aligned} & \;\;\;\;\text { 提取率 }(\%)=\frac{100 \times C\left(\mathrm{mg} \cdot \mathrm{~mL}^{-1}\right) \times 10^{-3}}{\text { 总黄酮质量 } M \mathrm{(~g})} \times \\ & 100 \end{aligned} $$ 1.3.2 ASE提取海刀豆中总黄酮
新采集到的海刀豆经过清洗,干燥后用粉碎机粉碎待用;分别精确称量5.00 g海刀豆粉末与硅藻土,混匀后装于底部放有玻璃纤维素滤纸片的34 mL的不锈钢萃取池中,在一定的萃取温度、萃取压力、萃取时间、甲醇体积分数和循环次数下,进行加速溶剂萃取。萃取液经过减压浓缩后按照上述方法测定吸光值,计算提取率。
1.3.3 单因素试验
影响总黄酮提取率的因素主要有萃取温度、萃取时间、甲醇体积分数、循环次数、萃取池体积。该试验分别对各个因素选取5个水平,进行单因素试验[20],来考察各因素对总黄酮提取率的影响。单因素试验因素水平表见表 1。
表 1 单因素试验因素与水平设计Table 1. Factors and levels for single factor analysis水平
level萃取温度/℃
temperature萃取时间/min
time甲醇体积分数/%
methanol concentration循环次数/次
cycle萃取池体积/mL
size of extraction pool1 50 5 50 1 10 2 70 10 60 2 34 3 90 15 70 3 66 4 110 20 80 4 5 130 25 90 5 1.3.4 响应面分析法试验设计
在单因素试验基础上,根据Box-Behnken的中心组合试验设计原理[21],选取对提取率有较显著影响的3个因素即萃取温度、萃取时间、甲醇体积分数,设计3因素3水平的响应面分析试验方案。响应面因素与水平表见表 2。
表 2 Box-Behnken设计试验因素与水平Table 2. Factors and levels for Box-Behnken design编码水平
code level萃取温度/℃
temperature (A)萃取时间/min
time (B)甲醇体积分数/%
methanol concentration (C)-1 90 10 60 0 110 15 70 1 130 20 80 2. 结果与分析
2.1 标准曲线的绘制
图 1是以芦丁为标准样品绘制的标准曲线,线性拟合方程式为y=9.700x+0.016,R2=0.992。测定样品的吸光度,从标准曲线上读取对应的浓度,计算出样品中总黄酮的提取率。
2.2 单因素试验
2.2.1 甲醇体积分数对总黄酮提取率的影响
分别精确称取5.00 g海刀豆粉末与硅藻土混匀,确定提取条件为萃取温度70 ℃,压力10.3 MPa,萃取时间10 min,循环1次,分别以50%、60%、70%、80%、90%和100%甲醇为萃取溶剂进行萃取,提取液减压浓缩后测定总黄酮含量,以3次试验平均值来表示提取率,结果见图 2。随着甲醇体积分数的升高,总黄酮提取率增加,当甲醇体积分数达到70%时提取率达到最大值,之后随着甲醇体积分数的增加,总黄酮提取率降低。因此,选取70%甲醇作为最佳萃取溶剂。
![]() 图 2 甲醇体积分数对总黄酮提取率的影响Figure 2. Effect of concentration of methanol on extraction rate of flavonoids
图 2 甲醇体积分数对总黄酮提取率的影响Figure 2. Effect of concentration of methanol on extraction rate of flavonoids2.2.2 萃取时间对总黄酮提取率的影响
分别精确称取5.00 g海刀豆粉末与硅藻土混匀,确定提取条件为萃取温度70 ℃,压力10.3 MPa,萃取溶剂70%甲醇,循环1次,分别静态萃取5 min、10 min、15 min、20 min和25 min,提取液干燥后测定总黄酮质量,以3次试验平均值来表示提取率,结果见图 3。静态萃取时间越长,提取率越高,然而,15 min与25 min对提取率的影响变化不显著(P>0.05),且考虑到提取效率以及提取试剂的用量,以15 min为最佳萃取时间。
2.2.3 萃取温度对总黄酮提取率的影响
分别精确称取5.00 g海刀豆粉末与硅藻土混匀,确定提取条件为压力10.3 MPa,萃取时间10 min,萃取溶剂70%甲醇,循环1次,分别在50 ℃、70 ℃、90 ℃、110 ℃和130 ℃下进行萃取,提取液浓干后测定总黄酮含量,以3次试验平均值来表示提取率,结果见图 4。随着萃取温度的升高总黄酮提取率依次升高,在升至110 ℃之后,110 ℃与130 ℃对提取率的影响变化不显著(P>0.05)。因此考虑到试验成本,最优的萃取温度以110 ℃为宜。
2.2.4 循环次数对总黄酮提取率的影响
分别精确称取5.00 g海刀豆粉末与硅藻土混匀,确定提取条件为萃取温度110 ℃,压力10.3 MPa,静态萃取时间15 min,萃取溶剂70%甲醇,分别循环1、2、3、4和5次,提取液干燥后测定总黄酮提取率,以3次试验平均值来表示,结果见表 3。随着循环次数的增加,提取率依次增加;然而循环2、3、4和5次对提取率的影响差异不大,且循环次数越多耗费时间越长,使萃取效率大大降低,故循环次数以2次为宜。
表 3 循环次数对总黄酮提取率的影响Table 3. Effect of cycles on extraction rate of flavonoids循环次数/次cycle 1 2 3 4 5 提取率/% extraction rate 73.45±0.23a 76.34±0.67b 77.28±0.34b 78.31±0.59b 78.91±0.41b 注:同一行中拥有不同小写字母的表示有显著性差异(P < 0.05);后表同此
Note:Different lowercase letters represent significant difference among different treatments (P < 0.05);the same case in the following table.2.2.5 萃取池体积对总黄酮提取率的影响
分别精确称取5.00 g海刀豆粉末与硅藻土混匀,确定提取条件为萃取温度110 ℃,压力10.3 MPa,萃取时间15 min,萃取溶剂70%甲醇,分别用10 mL、34 mL和66 mL的萃取池进行萃取,提取液干燥后测定总黄酮提取率,结果见表 4。同样条件下34 mL萃取池能够得到较高的提取率,因此34 mL萃取池为最优选择。
表 4 萃取池体积对总黄酮提取率的影响Table 4. Effect of extraction pool volume on extraction rate of flavonoids萃取池体积/mL size of extraction pool 10 34 66 提取率/% extraction rate 38.72±0.71a 78.27±0.64b 64.59±0.58c 2.2.6 各因素与提取率的方差分析
利用SPSS 17.0软件对不同提取条件下总黄酮提取率进行方差分析,结果见表 5。萃取温度、萃取时间、甲醇体积分数、循环次数对总黄酮提取率均影响显著甚至极显著,循环次数对于总黄酮提取率的影响不如其他3个因素显著。
表 5 单因素试验的方差分析(F/P)Table 5. Single-factor analysis of variance因素
factor甲醇体积分数
concentration of methonal萃取时间
time萃取温度
temperature循环次数
cycleF 513.16 36.90 287.86 19.19 P <0.000 1 0.000 7 <0.000 1 0.007 7 注:P < 0.05为显著;P < 0.01为极显著
Note:significant at P < 0.05;very significant at P < 0.01.2.3 模型方程的建立与显著性分析
根据单因素试验将显著影响总黄酮提取率的3因素的适合区间列于表 2。采用Box-Behnken设计法,对海刀豆中总黄酮提取进行3因素3水平的优化试验,考察各因素之间的交叉影响;以3次试验所得总黄酮提取率的平均值为响应值,结果见表 6。17个试验点可分为2类:1)析因点。自变量取值在各因素所构成的三维顶点,共有12个析因点;2)零点。为区域的中心点,零点试验重复5次,用以估计试验误差。
表 6 试验设计及结果Table 6. Test design and results of Box-Behnken method试验序号
test No.萃取温度
temperature (A)萃取时间
time (B)甲醇体积分数
concentration of methonal (C)提取率/%
extration rate1 0 0 0 77.46 2 0 -1 1 67.47 3 0 0 0 78.37 4 1 0 -1 83.87 5 0 1 -1 65.62 6 0 0 0 82.96 7 0 -1 -1 65.41 8 1 1 0 82.39 9 -1 -1 0 54.49 10 0 0 0 84.52 11 -1 1 0 64.33 12 -1 0 1 76.97 13 1 -1 0 62.73 14 0 1 1 76.03 15 -1 0 -1 67.57 16 1 0 1 78.91 17 0 0 0 85.57 采用Design Expert 8.05软件对各因素进行方差分析[22-23],结果见表 7。回归模型极显著(P < 0.01),并且其失拟性不显著(P>0.05),说明回归模型能够较好地预测相应的响应变量;萃取温度与萃取时间对总黄酮提取率有显著影响,甲醇体积分数对总黄酮提取率的影响相对不显著,然而,单因素试验数据说明甲醇体积分数对于总黄酮提取率的影响是显著的,其原因可能在于3个因素之间对响应值有着交叉影响,且萃取温度和萃取时间对响应值的影响要比甲醇体积分数显著;即影响响应值因素的显著性顺序为萃取温度、萃取时间、甲醇体积分数。因此各个因素对响应值的影响不是简单的线性关系,经回归拟合后可用如下二次多项回归方程(响应值由y表示)表示:y=-310.98+3.25A+9.73B+3.17C+0.02AB-0.02AC+0.04BC-0.01A2-0.48B2-0.01C2
表 7 方差分析表Table 7. Analysis of variance方差来源
soruce of variation平方和
sum of squares自由度
df均方
mean squareF P 显著性
significanceModel 1 265.80 9 140.64 7.97 0.006 1 ** A 247.98 1 247.98 14.06 0.007 2 ** B 183.07 1 183.07 10.38 0.014 6 * C 35.74 1 35.74 2.03 0.197 6 AB 24.11 1 24.11 1.37 0.280 6 AC 51.55 1 51.55 2.92 0.131 1 BC 17.43 1 17.43 0.99 0.353 3 A2 60.70 1 60.70 3.44 0.106 0 B2 605.73 1 605.73 34.35 0.000 6 ** C2 5.56 1 5.56 0.32 0.591 9 残差residual 123.45 7 17.64 失拟量lack of fit 69.90 3 23.30 1.74 0.296 7 纯误差pure error 53.55 4 13.39 总离差total 1 389.25 16 注:*. 显著(P < 0.05);* *. 极显著(P < 0.01);不显著(P >0.1)
Note:*. significant (P < 0.05);* *. very significant (P < 0.01);not significant (P>0.1)2.4 响应面分析对总黄酮提取率的影响
3个因素之中两两因素对响应值的交叉影响见图 5、图 6和图 7。在3D响应面图中,响应值随着因素数值的增加变化增大,即斜面的倾斜度越大,说明该因素对响应值(提取率)的影响越显著。在等高线中,两两因素构成的等高线越像椭圆形,说明这两种因素对于响应值有着交叉影响。因此,响应面呈规则的凸起形,表明在试验因素水平范围内存在极大值,即响应面的最高点;萃取温度与萃取时间的等高线呈椭圆形,即两者之间存在交叉影响(图 5);响应值随着萃取时间的增加变化显著,而随甲醇体积分数增加变化不明显,萃取温度与萃取时间的等高线呈椭圆形,即说明两者之间存在着交叉影响且萃取时间比甲醇体积分数对响应值影响显著(图 6);响应值随着萃取温度与甲醇体积分数的增加变化均显著,而萃取温度与甲醇体积分数的等高线呈线条状,说明两者之间无交叉影响(图 7)。
![]() 图 5 萃取温度和萃取时间对总黄酮提取率影响的响应面与等高线Figure 5. Response surface and contour for effects of temperature and time on extraction rate of flavonoids
图 5 萃取温度和萃取时间对总黄酮提取率影响的响应面与等高线Figure 5. Response surface and contour for effects of temperature and time on extraction rate of flavonoids![]() 图 6 萃取时间和甲醇体积分数对总黄酮提取率影响的响应面与等高线Figure 6. Response surface and contour for effects of time and concentration of methanol on extraction rate of flavonoids
图 6 萃取时间和甲醇体积分数对总黄酮提取率影响的响应面与等高线Figure 6. Response surface and contour for effects of time and concentration of methanol on extraction rate of flavonoids![]() 图 7 萃取温度和甲醇体积分数对总黄酮提取率影响的响应面与等高线Figure 7. Response surface and contour for effects of temperature and concentration of methanol on extraction rate of flavonoids
图 7 萃取温度和甲醇体积分数对总黄酮提取率影响的响应面与等高线Figure 7. Response surface and contour for effects of temperature and concentration of methanol on extraction rate of flavonoids2.5 最佳预测条件
由响应面软件Design Expert分析得到的海刀豆中总黄酮的最佳提取条件见表 8,由于这2组条件比较接近,所以选取第一组条件进行验证,为了操作方便将条件修正为萃取温度130 ℃,萃取时间16 min,甲醇体积分数66%,循环2次,3次平行试验所得总黄酮提取率分别为82.40%、84.61%和83.27%,平均值为83.43%,与预测值相对偏差-1.63%,说明该模型具有一定的适用性。
表 8 总黄酮提取率的最佳预测条件Table 8. Optimum of extraction rate of flavonoids序号
No.萃取温度/℃
temperature萃取时间/min
time甲醇体积分数/%
concentration of methanol提取率/%
extraction rate理想值
ideal value1 130 16.33 65.99 84.808 1 0.666 2 129.99 16.34 66.08 84.808 0 0.666 3. 结论
通过单因素试验优化条件,根据Box-Behnken设计法进行试验得出数据,最后经过响应面分析得出加速溶剂萃取技术提取海刀豆中总黄酮的最佳工艺条件为萃取温度130 ℃,萃取时间16 min,甲醇体积分数为66%,循环次数2次,得到的理论提取率为84.81%,经验证实际提取率为83.43%。通过响应面可知各单因素对海刀豆中总黄酮提取率的影响显著性顺序为萃取温度>萃取时间>甲醇体积分数>循环次数。
-
![]()
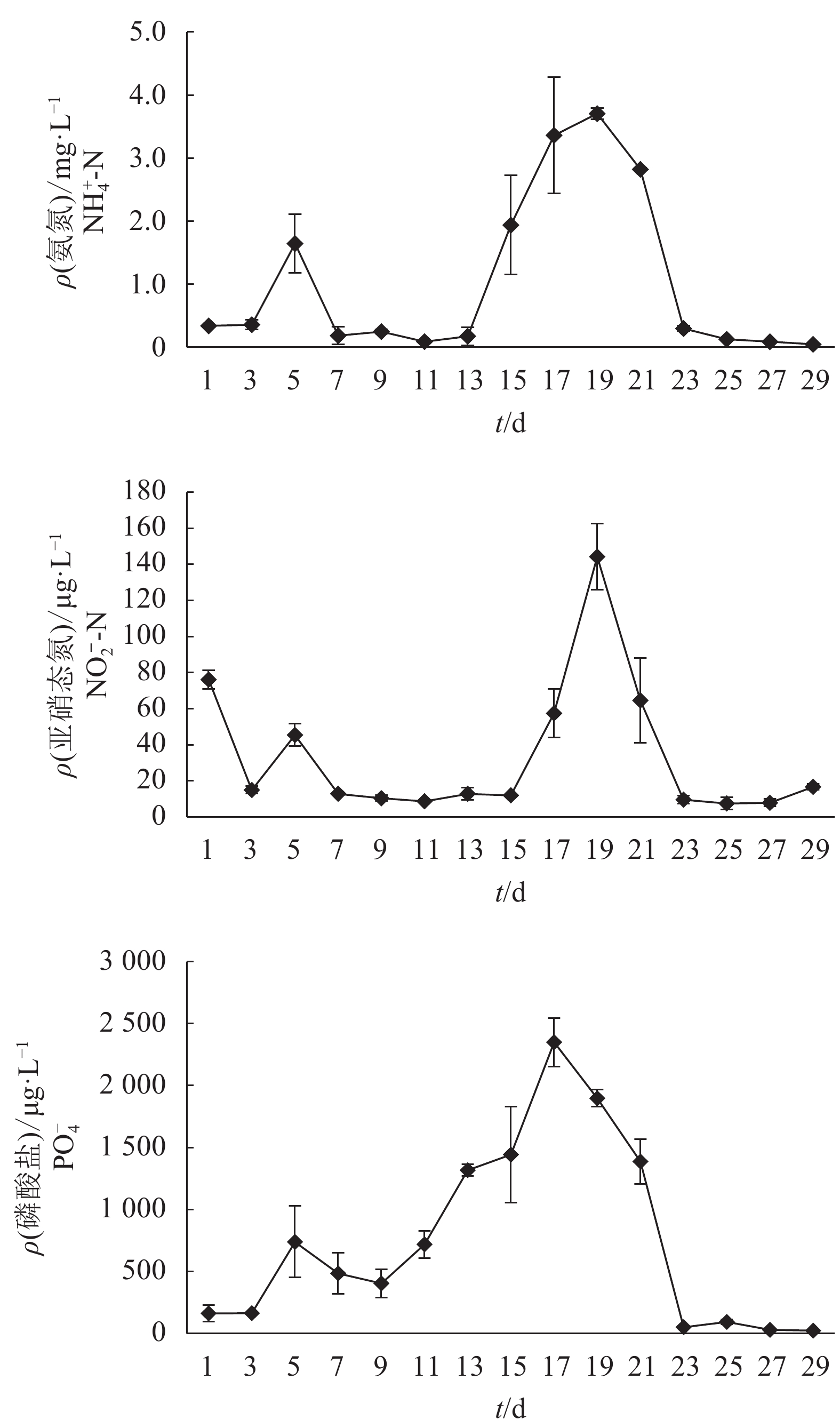
图 1 培养稳定生物絮凝体1个月期间水体水化指标
图中数据为平均数±标准差(n=3),后图同此
Figure 1. Hydrochemical indices of stable biofloc cultivation during one month
The data are average values and standard deviation of three replicates. The same case in the following figures.
![]()
图 2 培养稳定生物絮凝体一个月期间生物沉降体积和水体总悬浮颗粒物质量浓度
Figure 2. BFV and TSS mass concentration of stable biofloc cultivation during one month
![]()
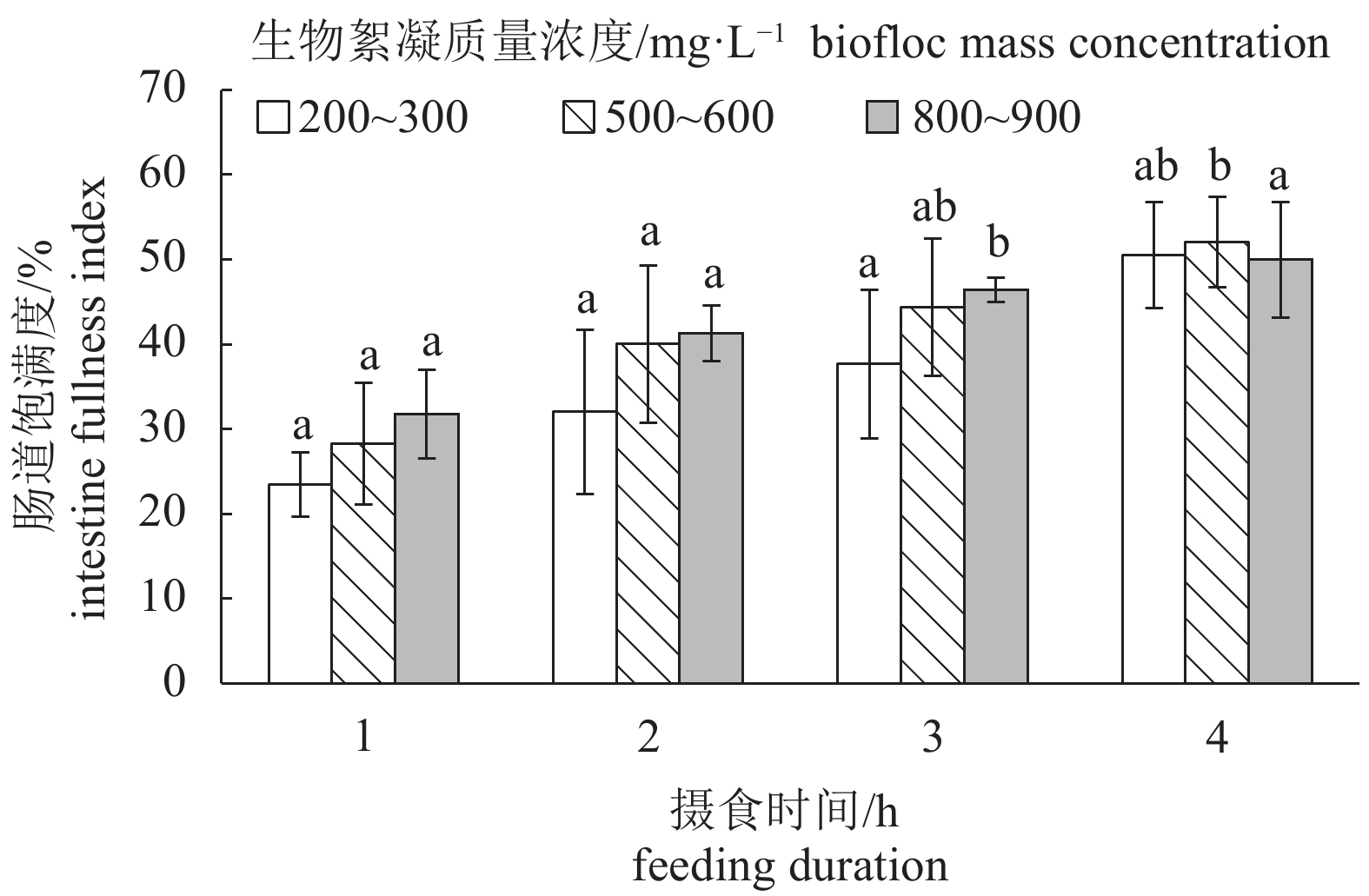
图 3 幼虾摄食相同质量浓度生物絮凝不同摄食时间肠道饱满指数变化 (n=10)
图中柱形图数值为平均值,方柱上不同字母表示存在显著性差异(P<0.05),后图同此
Figure 3. Intestine fullness index of juveniles fed at same biofloc mass concentration with different feeding durations
The data are average values. Values with different small letters are different significantly (P<0.05). The same case in the following figures.
![]()
图 4 幼虾相同摄食时间不同浓度生物絮凝肠道饱满指数变化
Figure 4. Intestine fullness index of juveniles fed at different biofloc mass concentrations with same feeding duration
![]()
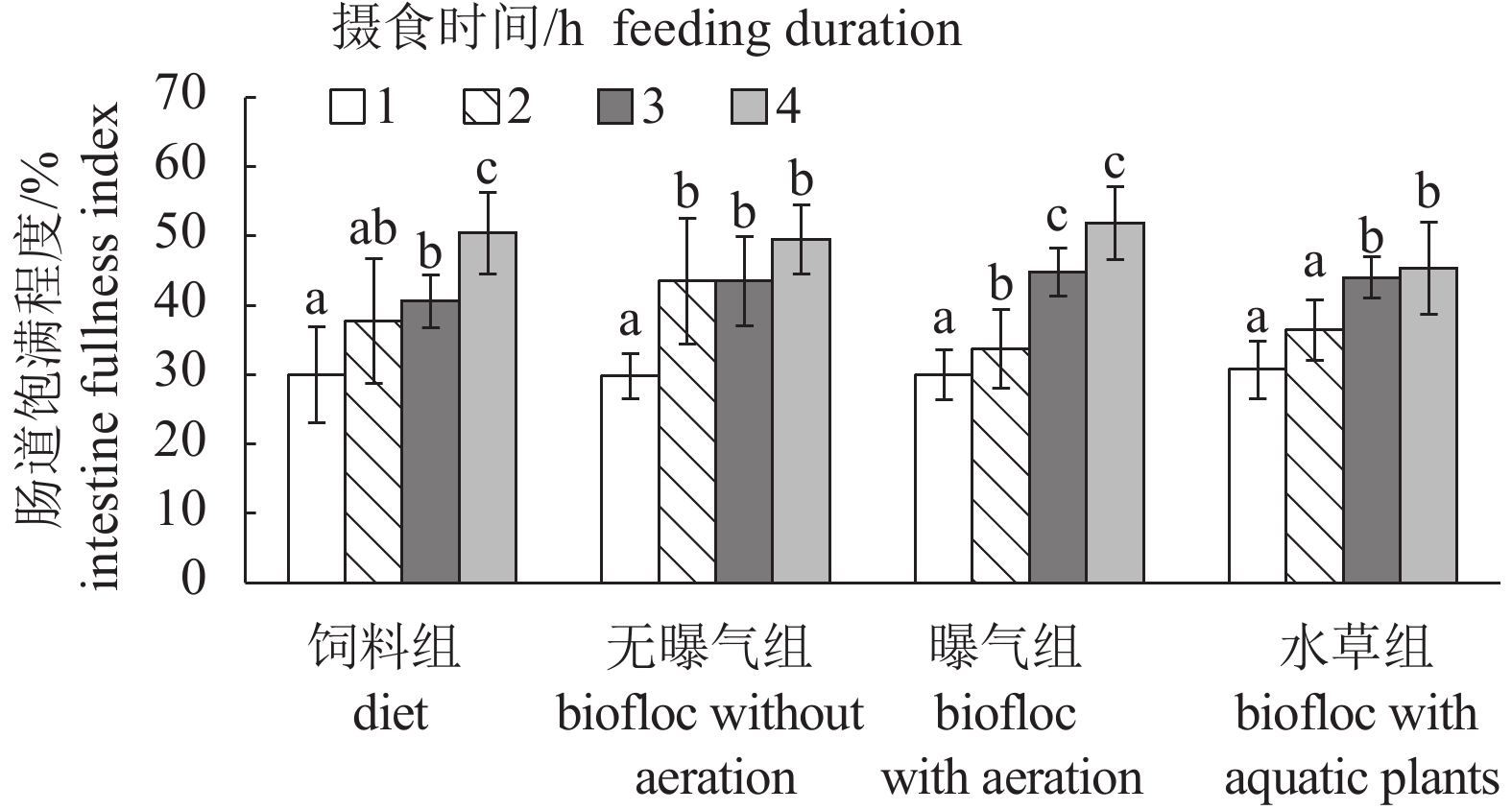
图 5 对照组与絮凝组的肠道饱满指数 随摄食时间的变化 (n=8)
Figure 5. Change of intestine fullness index of the control and biofloc groups with different feeding durations
![]()
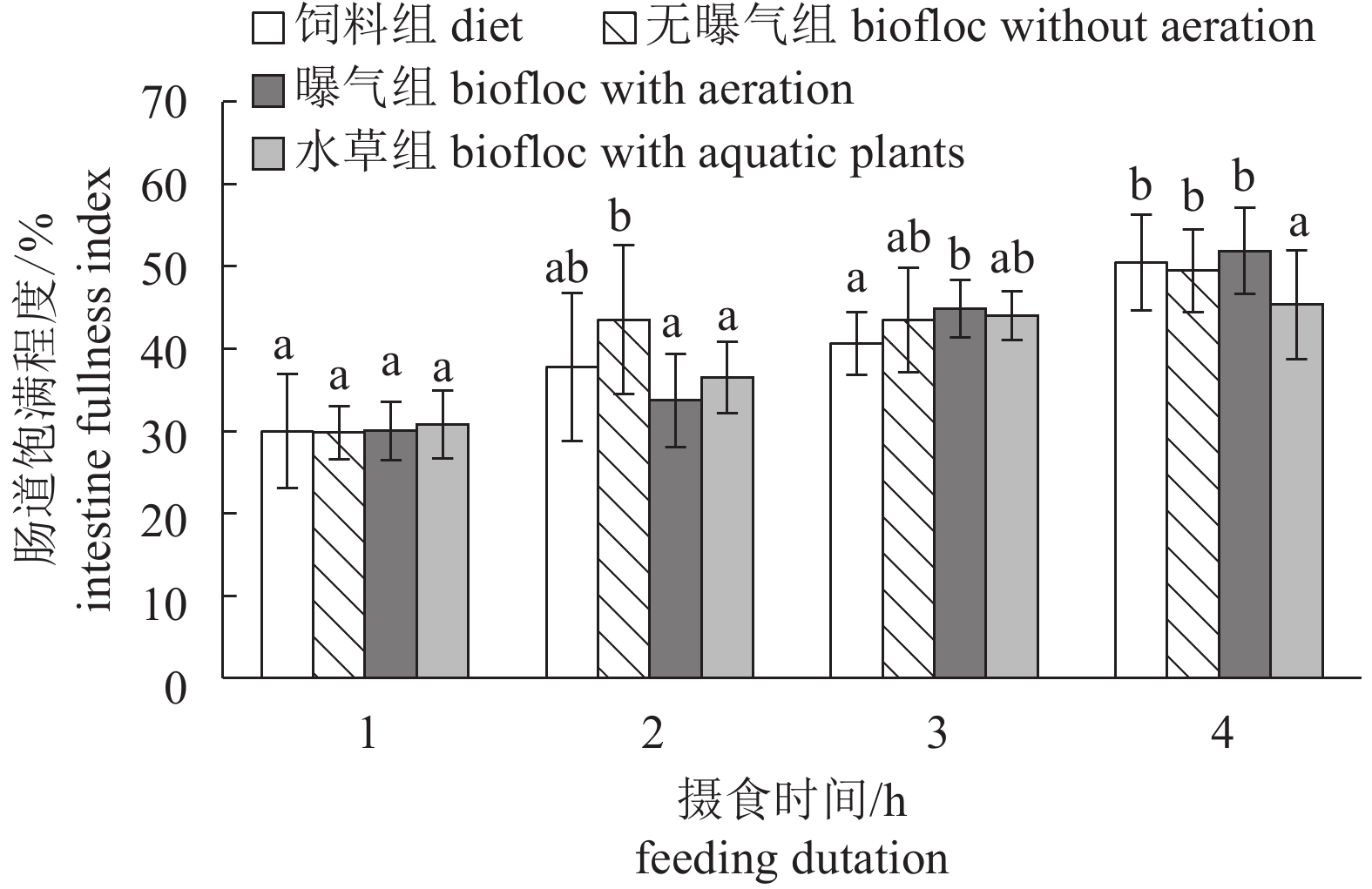
图 6 相同时间内对照组与不同摄食条件絮凝组的肠道饱满指数的变化
Figure 6. Change of intestine fullness index of the control and biofloc groups under different feeding conditions and with same feeding duration
表 1 不同生物絮凝浓度下幼虾的肠道充塞度
Table 1 Fullness rank of intestine of juveniles fed at different biofloc mass concentrations
生物絮凝质量浓度/mg·L–1
biofloc mass concentration摄食时间/h
feeding duration1 2 3 4 200~300 0~1级 1~2级 3级 3~4级 500~600 0~1级 2~3级 3级 4级 800~900 1级 2~3级 3~4级 4级  下载: 导出CSV
下载: 导出CSV
表 2 不同摄食条件下幼虾的肠道充塞度
Table 2 Fullness of intestines of juveniles under different feeding conditions
不同条件组
group摄食时间/h
feeding duration1 2 3 4 饲料组 diet 0~1级 1级 2级 3~4级 无曝气组 biofloc without aeration 0~1级 2级 2~3级 3~4级 曝气组 biofloc with areation 1级 1~2级 2~3级 4级 水草组 biofloc with aquatic plants 0~1级 1~2级 2~3级 3~4级
下载: 导出CSV
-
[1] 王顺昌. 克氏螯虾的生物学和生态养殖模式[J]. 淡水渔业, 2003, 33(4): 59-61. [2] 田娟, 许巧情, 田罗, 等. 洞庭湖克氏原螯虾肌肉成分分析及品质特性分析[J]. 水生生物学报, 2017, 41(4): 870-877. [3] 农业部渔业政策管理局. 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2017: 24-25. [4] CRAB R, AVNIMELECH Y, DEFOIRDT T, et al. Nitrogen removal techniques in aquaculture for a sustainable production[J]. Aquaculture, 2007, 270(1/2/3/4): 1-14.
[5] 罗国芝, 朱泽闻, 潘云峰, 等. 生物絮凝技术在水产养殖中的应用[J]. 中国水产, 2010(2): 62-63. [6] 任信林, 凌武海, 纪翠萍. 环境因子对克氏原螯虾养殖的影响[J]. 水产科学, 2009, 28(11): 710-712. [7] 宋亮, 张建平, 韩晓磊, 等. 克氏原螯虾养殖现状及对策[J]. 常熟理工学院学报, 2011, 25(2): 85-87, 91. [8] AZIM M E, LITTLE D C. The biofloc technology (BFT) in indoor tanks: water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus)[J]. Aquaculture, 2008, 283(1/2/3/4): 29-35.
[9] QIU X, DAVIS D A. Effects of dietary phytase supplementation on growth performance and apparent digestibility coefficients of Pacific white shrimp Litopenaeus vannamei[J]. Aquacult Nutr, 2016, 23(5): 942-951.
[10] AVNIMELECH Y. Carbon nitrogen ratio as a control element in aquaculture systems[J]. Aquaculture, 1999, 176(3/4): 227-235.
[11] WANG C, PAN L, ZHANG K, et al. Effects of different carbon sources addition on nutrition composition and extracellular enzymes activity of bioflocs, and digestive enzymes activity and growth performance of Litopenaeus vannamei in zero-exchange culture tanks[J]. Aquacult Res, 2016, 47(10): 3307-3318.
[12] AHMAD I, RANI A, VERMA A K, et al. Biofloc technology: an emerging avenue in aquatic animal healthcare and nutrition[J]. Aquacult Int, 2017, 25(3): 1215-1226.
[13] CUZON G, LAWRENCE A, GAXIOLA G, et al. Nutrition of Litopenaeus vannamei, reared in tanks or in ponds[J]. Aquaculture, 2004, 235(1): 513-551.
[14] 徐增洪, 周鑫, 水燕. 克氏原螯虾的食物选择性及其摄食节律[J]. 大连海洋大学学报, 2012, 27(2): 166-170. [15] 母红霞, 孙宝柱, 曹文宣, 等. 张氏鳖的食性分析[J]. 水生生物学报, 2011, 35(3): 373-378. [16] LONG L A, YANG J, LI Y, et al. Effect of biofloc technology on growth, digestive enzyme activity, hematology, and immune response of genetically improved farmed tilapia (Oreochromis niloticus)[J]. Aquaculture, 2015, 448: 135-141.
[17] WANG G J, YUA E, XIE J, et al. Effect of C/N ratio on water quality in zero-water exchange tanks and the biofloc supplementation in feed on the growth performance of crucian carp, Carassius auratus[J]. Aquaculture, 2015, 443: 98-104.
[18] 徐武杰. 生物絮团在对虾零水交换养殖系统中功能效应的研究与应用[D]. 青岛: 中国海洋大学, 2014: 9-10. [19] 方金龙, 王元, 房文红, 等. 氨氮胁迫下白斑综合征病毒对凡纳滨对虾的致病性[J]. 南方水产科学, 2017, 13(4): 52-58. [20] 洪美玲. 水中亚硝酸盐和氨氮对中华绒螯蟹幼体的毒性效应及维生素E的营养调节[D]. 上海: 华东师范大学, 2007: 29. [21] MOULLAC G L, HAFFNER P. Environmental factors affecting immune responses in Crustacea[J]. Aquaculture, 2000, 191(1/2/3): 121-131.
[22] 孙国铭, 汤建华, 仲霞铭. 氨氮和亚硝酸氮对南美白对虾的毒性研究[J]. 水产养殖, 2002(1): 22-24. [23] 钟君伟, 朱永安, 孟庆磊, 等. 氨氮对2种规格克氏原螯虾的急性毒性研究[J]. 长江大学学报(自科版), 2013, 10(8): 55-59. [24] 钟君伟, 朱永安, 付佩胜, 等. 亚硝酸盐氮对克氏原螯虾成虾的毒性及其抗病因子的影响[J]. 长江大学学报(自科版), 2014, 11(2): 36-40. [25] 杨万喜, 赖伟, 堵南山. 日本沼虾行为研究[J]. 动物学杂志, 1997, 32(3): 51-54. [26] 丁张妮, 徐永健, 林建华, 等. 盐度对三疣梭子蟹摄食行为及生长特性的影响[J]. 生态科学, 2014, 33(5): 899-903. [27] SCHVEITZER R, ARANTES R, COSTODIO P F, et al. Effect of different biofloc levels on microbial activity, water quality and performance of Litopenaeus vannamei in a tank system operated with no water exchange[J]. Aquacult Eng, 2013, 56(3): 59-70.
[28] RAY A J, LEWIS B L, BROWDY C L, et al. Suspended solids removal to improve shrimp (Litopenaeus vannamei) production and an evaluation of a plant-based feed in minimal-exchange, superintensive culture systems[J]. Aquaculture, 2010, 299(1/2/3/4): 89-98.
[29] 陈勇, 李朝晖, 唐宁, 等. 饲料形状对克氏原螯虾摄食率和生长的影响[J]. 湖北农业科学, 2011, 50(9): 1859-1860. [30] 李泽健. 低氧胁迫对中华绒螯蟹能量代谢、呼吸代谢及抗氧化代谢的影响[D]. 保定: 河北大学, 2012: 2-3. [31] 谢文星, 董方勇, 谢山. 克氏原螯虾的食性、繁殖和栖息习性研究[J]. 水生态学杂志, 2008, 28(4): 63-65. [32] 但丽, 张世萍, 羊茜, 等. 克氏原螯虾食性和摄食活动的研究[J]. 湖北农业科学, 2007, 46(3): 436-438. [33] 叶奕佐, 韩育章, 王苹萍, 等. 沼虾、河蟹对栖息隐蔽物的选择性[J]. 水产科技情报, 1995, 22(6): 243-249. [34] 孙中勇, 李嘉尧, 杨筱珍, 等. 不同条件对中华绒螯蟹扣蟹摄食生物絮团的影响[J]. 生物学杂志, 2017, 34(6): 32-36. -
期刊类型引用(3)
1. 李晨露,李京昊,成永旭,涂纹,吴乐,奚业文,朱传忠,李嘉尧. 不同饵料对克氏原螯虾仔虾养殖水质、生长、消化酶和非特异性免疫酶的影响. 上海海洋大学学报. 2022(04): 883-892 .  百度学术
百度学术
2. 刘栋梁,黄建华,杨丽诗,谭理琦. 多种环境因子对克氏原螯虾蜕壳死亡的影响及对策研究. 南方水产科学. 2020(02): 29-35 . 本站查看
3. 骆磊,黄建华,刘栋梁,谭理琦,江世贵,周发林,姜松,杨其彬,杨丽诗. 克氏原螯虾对2种材质洞穴的选择和空间分布研究. 南方水产科学. 2019(04): 76-81 . 本站查看
其他类型引用(8)









计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 11
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
