
Effects of multiple environmental factors on molting death of Procambarus clarkii and countermeasures
-
摘要:
为探究克氏原螯虾 (Procambarus clarkii) 蜕壳死亡原因及预防对策,该研究以不同环境因子饲养和胁迫不同规格的克氏原螯虾,初步探讨其蜕壳死亡原因及预防对策。结果表明,水体中钙(Ca2+)质量浓度为30 mg·L−1、镁(Mg2+)质量浓度为15 mg·L−1时,克氏原螯虾的成活率和蜕壳率最高,显著高于对照组 (P<0.05),而蜕壳死亡率显著低于对照组 (P<0.05);水体中pH为9.23,或者溶解氧质量浓度为4.5 mg·L−1时,虾的成活率和蜕壳率显著低于对照组 (P<0.05),而蜕壳死亡率显著高于对照组 (P<0.05);盐度为21时的蜕壳死亡率最高 (P<0.05)。环境因子等诸多因素是造成克氏原螯虾蜕壳死亡的重要原因,而蜕壳死亡现象时有发生,需采取预防对策方可提高产量。
Abstract:In order to study the cause of molting death of Procambarus clarkii and its preventive measures, we raised and stressed the crayfish of different sizes by different environmental factors. The results show that the survival rate and decidua rate of P. clarkii were highest in groups of 30 mg·L−1 Ca2+ and 15 mg·L−1 Mg2+, significantly higher than those in the contol (P<0.05), but their decidua mortalities were significantly lower than those in the control (P<0.05). When the pH value was 9.23 or the dissolved oxygen concentration was 4.5 mg·L−1, the survival rate and decidua rate were significantly lower than those in the control (P<0.05), but the decidua mortality was significantly higher than that in the control (P<0.05). The molting mortality of P. clarkii was highest when the salinity was 21 (P<0.05). Environmental factors and many other factors are the important causes of molting death of P. clarkii.
-
拖网作为渔业中的重要捕捞方式,具有覆盖水域广泛、捕获物种多样等特点[1]。2022年全国拖网渔业总产量451 万吨,拖网渔船共计22 566 艘,总功率达516 万千瓦[2]。巨大的经济产值下,不可忽视的是成千上万吨的燃油消耗和有限的单船产量。现代单船拖网作业中,拖网网板的作用是扩大网口、提高捕捞效率,加速网具下沉、减少渔获逃逸[3]。实际生产中,不同阶段对网板的作用需求不同,如拖网布放和收起阶段中,应要减小网口扩张,减轻渔船机械负荷,而传统网板无法兼顾[4]。此外,控制网口深度需要依靠调整渔船速度和曳纲长度,这不但要求操作者经验丰富,还对渔船的机械素质要求较高。研究能够调节扩张力大小和变水层拖曳的网板是拖网渔业领域中的重点需求,备受国内外学者的关注[5]。
网板的设计与改进应以效益和效率为前提,需要平衡网板扩张力与阻力的关系,同时保证网板姿态稳定。网板的结构现已从矩形、V形、椭圆形、圆盾形等单叶类型,逐渐发展为带副翼的单叶类型网板和双叶类型网板等[6]。在这个过程中,最初的矩形网板发展成矩形曲面网板进行研究[7]。Liu等[8]对底拖网使用的椭圆双开缝网板,结果显示,当冲角达到15° 时,其升阻比达4.165,但椭圆形网板对海底环境影响较大。刘志强[9]研究的立式双曲面网板具有较好的稳定性,在数值模拟实验中升力系数为1.88。庄鑫[10]研究的高升力双叶型网板在30° 冲角下最大升力系数可达2.19。除了基础形状的设计和优化外,有学者针对网板的不同结构参数进行梯度研究,也取得了有效进展。Xu等[11]的数值模拟研究表明,矩形网板的最佳展弦比为0.5,且更高的展弦比具有更小的临界冲角。Chu等[12]研究分析得出垂直曲面V形网板基础结构参数状态下的最大升阻比为5.7。Fukuda等[13]通过水槽实验发现,展弦比和弯曲度均对网板水动力性能有明显影响,展弦比小于2.5 时,网板临界冲角偏大,弯曲度15%时产生最大升力的最佳冲角范围较大。王明彦等[14]利用风洞实验得出,影响立式 V 形曲面网板水动力性能的主要因素依次为板面折角、展弦比和后退角。刘健等[15]比较了2种立式曲面缝翼式网板的性能,发现合理调节导流翼偏角可以达到优化网板水动力学性能的目的。刘志强等[16]认为现有研究还应考虑网板发生倾斜时其水动力及周围流场特性的变化。
在研究方法上,Xu等[17]分析了计算流体动力学X 模式 (Computational fluid dynamics X, CFX) 和流体 (FLUENT) 2种仿真方法对V形网板的水动力结果实验影响发现,FLUENT在网板周围的速度场描绘方面更加精确。Xu等[18]还通过中心组合设计法 (Central composite design, CCD) 和响应面分析得出,矩形弧面网板最大相对弧度为15.5%、最大弧度位置为35.1%时,展弦比为2.0时的性能较好。氢气泡和数字粒子图像测速 (Digital particle image velocimetry, DPIV) 粒子等新技术的加入使得渔具周围流场特点直观地展示出来,用以验证计算机模拟流场结果[19-21]。Liu等[22]通过循环水槽实验结果总结了展弦比对网板性能的影响,随着展弦比的增大,最大升力系数与最大升阻比呈现先增后减的趋势。Wang等[23]通过风洞模型检验了一款双叶网板,在展弦比2.0、前翼弯度0.16、间隙比0.35时性能最适,但该方法在实验时对模型在流体中的雷诺数匹配程度要求高。
本研究基于数值模拟方法,研究叶片转向、叶片转角及冲角等参数对多叶可控网板水动力性能的影响,并通过流场可视化,分析不同叶片转角下网板周围流体的变化,以探究叶片转动对网板运动姿态的影响,以及多叶可控网板对网口状态的控制策略。
1. 材料与方法
1.1 网板结构参数
多叶可控网板模型整体翼弦 $ ( $C$ ) $ 宽300 mm,翼展 $ ( $L$ ) $ 长600 mm,展弦比 ($ { \lambda } $) 2.0。网板结构参数以及设计变量见图1。多叶可控网板主要分为上、下两部分,每部分由3个翼型叶片组成;叶片弦长100 mm,翼展200 mm,叶片中心面轮廓形状为P50-3翼型,该翼型最大厚度在翼弦的20.5%处,为9.7%;最大曲面在翼弦的46.8%处,为9.57%;其中,上、下两端的前叶与中叶可绕自身平行于翼展方向的重心轴转动,设定与网板冲角转动一致的方向为叶片转动的正方向,与网板冲角转动相反的方向为叶片转动的负方向,上端叶片转角为 $ {\theta }_{1} $,下端叶片转角为 $ {\theta}_{\text{2}} $,叶片转动范围为−40°~40°[24]。设置叶片闭合状态下的网板冲角范围为0°~45°,间隔5°,实验对照设置如表1所示。网板两端尾部的叶片为固定襟翼。
表 1 多叶可控网板冲角以及叶片转角设置情况Table 1 Multi-blade controllable otter board attack angle and blade rotation angle setting实验组别
Experimental group冲角
Angle of attack转动的
叶片位置
Position of
blade叶片转角
Blade rotation angle1 0°~45° 无 0° 2 0° 两端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°3 0° 上端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°4 0° 下端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°5 20° 两端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°6 20° 上端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°7 20° 下端 40°、30°、20°、10°、
−40°、−30°、−20°、−10°1.2 数值模拟
本研究使用Solidworks 2016三维绘图软件对多叶可控网板进行结构建模。采用数值模拟方法,通过计算流体动力学 (Computational fluid dynamics, CFD) 软件ANSYS 21.0对雷诺平均N-S方程进行求解,描述了不可压缩流体的动力学如图2所示。模型设定位于距离入水口5.0 m,距离水底高度1.0 m,网板中心轴距离两侧壁面2.0 m。为保证数值模拟的有效性,进行了网格无关性验证,分别对600、800、1 000和1 100万级网格数量的模拟结果进行测试。模拟出的升力与阻力结果的标准差均未超过0.04,结果表明网格密度的增加,对计算结果的影响有限,800万级网格数量足以保证模拟的精度[25]。计算域中最小单元格尺寸为5 mm,最大为100 mm,网板的局部单元格尺寸为5 mm (图3)。网板的边界处和计算域壁面设置了5级膨胀层,增长率为1.5 (1.07<y+<42.34)。对网板进行局部加密,得到非结构化网格。节点数量和元素总数分别达到了4.8×105和8.5×106。采用k-ε湍流模型 (Realizable) 求解流动模型的控制方程[26]。
流体设定为淡水,其密度为 $ \text{998.2kg∙}{\text{m}}^{{\text{−}}\text{3}} $,黏度为 $ \text{0.001 003kg∙}{\text{ms}}^{{\text{−}}\text{1}} $。设定计算域上流入口为速度入口边界,沿X轴方向均匀流动,湍流强度为5%,下流出口设置为充分发展的自由流边界 (0 Pa)。模型表面被认为是光滑且不移动条件,且设定其他壁面具有自由滑动条件的光滑壁面。求解方法选择压力-速度耦合方法。通过二阶迎风方程对压力、动量、湍流动能和湍流耗散率进行了空间离化[27]。其收敛残差精度为 $ {10}^{{\text{−}}3} $。
1.3 水动力系数公式
网板水动力性能相关参数有雷诺数 $ {R}_{\mathrm{e}} $、升力系数 $ {C}_{{\mathrm{l}}} $、阻力系数 $ {C}_{{\mathrm{d}}} $、升阻比K、Z轴分力系数 $ {C}_{{\mathrm{z}}} $ 以及X轴力矩系数 $ {C}_{\mathrm{m}\mathrm{x}} $,计算公式为:
$$ {R}_{{\mathrm{e}}}{\text{=}}\frac{vc\rho }{\mu } $$ (1) $$ {C}_{{\mathrm{l}}}{\text{=}}\frac{2{F}_{{\mathrm{l}}}}{\rho s{v}^{2}} $$ (2) $$ {C}_{{\mathrm{d}}}{\text{=}}\frac{2{F}_{{\mathrm{d}}}}{\rho s{v}^{2}} $$ (3) $$ K=\frac{{C}_{{\mathrm{l}}}}{{C}_{{\mathrm{d}}}} $$ (4) $$ {C}_{{\mathrm{z}}}{\text{=}}\frac{2{F}_{{\mathrm{z}}}}{\rho s{v}^{2}} $$ (5) $$ {C}_{{\mathrm{mx}}}{\text{=}}\frac{2{M}_{{\mathrm{x}}}}{\rho bs{v}^{2}} $$ (6) 式中:v为流速 (m·s−1);c为网板翼弦 (m);ρ为流体密度 (kg·m−3);μ为流体动力黏性系数 (kg∙ms−1);Fl为升力 (N);Fd为阻力 (N);Fz为Z轴分力 (N);Mx为X轴力矩 (N∙m);b为网板翼展 (m)。
2. 结果
2.1 叶片闭合时网板水动力系数
数值模拟结果如图4所示,1号组模型的阻力系数随冲角的增大而上升。升力系数随冲角的增大出现先上升后下降的趋势,并在20° 冲角时达到最高值0.88,随后下降,在45° 冲角时与阻力系数接近。升阻比方面,该状态下网板的升阻比最大值为8.85,冲角为5°,随后冲角继续增大,升阻比下降明显,在45° 冲角时仅为1.02。网板在Z轴方向上的分力系数受冲角变化的影响较小,在20° 冲角时,该力方向与Z轴负半轴方向一致,系数仅为1.85×10−3。同样,X轴的力矩系数受冲角变化影响也较小,最大值在15° 冲角时出现,为1.03×10−3。
![]() 图 4 叶片闭合时网板水动力系数Fig. 4 Hydrodynamic coefficient of otter board when blades were closed
图 4 叶片闭合时网板水动力系数Fig. 4 Hydrodynamic coefficient of otter board when blades were closed2.2 叶片打开时网板水动力系数
2.2.1 0° 冲角下叶片转动对网板水动力特性的影响
2号、3号、4号组模型仿真结果如图5所示,2号组叶片朝正反两方向转动均会导致网板的阻力系数上升,且朝正方向转动时影响更大。而3号和4号组的阻力系数均低于2号组;此外,3组模型实验结果均显示,与负方向对比,叶片朝正方向转动时,阻力系数上升更快。
![]() 图 5 0° 冲角下叶片转角对网板水动力特性的影响Fig. 5 Influence of blade rotation angle of 0° attack angle on hydrodynamic characteristics otter board
图 5 0° 冲角下叶片转角对网板水动力特性的影响Fig. 5 Influence of blade rotation angle of 0° attack angle on hydrodynamic characteristics otter board升力系数方面,2号组叶片转动对升力系数的影响高于3号和4号组。3组模型叶片朝正方向转动时,升力系数先增加后减小,均在 $\theta \text{=20°} $ 时达到升力系数最大值;叶片朝负方向转动时,升力系数均逐渐下降,且2号组模型的叶片转动到 −20°时,升力方向转为收缩方向,增加叶片转角,收缩趋势增加,而3号和4号组的升力系数逐渐趋于0。
升阻比方面,2号组模型的叶片向两个方向转动均会导致网板的升阻比下降,且叶片朝负方向转动,升阻比下降明显;当2号组两端叶片转角达到 −20° 时,升力方向改变,升阻比为 −0.99,随后转角增大,升阻比下降。当 $\theta \text{=40°} $ 时,3号和4号组的升阻比为1.74,2号组的升阻比为1.14。
X轴的力矩方面,2号组的旋转力矩绕0值波动,变化幅值小,在 $\theta \text{=30°} $ 时,$ C_{\rm{mx}} $ 仅为 $ -\text{2.04×}{\text{10}}^{{\text{−}}\text{3}} $。当3号组模型叶片朝负方向转动时,该力矩系数先增加后减小,在 $\theta \text{=}{\text{−}}\text{30°} $ 时,$ C_{\rm{mx}} $为 $ \text{1.53×}{\text{10}}^{{\text{−}}\text{2}} $;当3号组模型叶片朝正方向转动时,力矩系数同样先增加后减小,在 $\theta \text{=20°} $ 时,$ C_{\rm{mx}} $= $ \text{1.38×}{\text{10}}^{{\text{−}}\text{2}} $。4号组模型叶片朝负方向转动时,该力矩系数不断增加,在 $\theta \text{=}{\text{−}}\text{40°} $ 时,$ C_{\rm{mx}}\text{=1.53×}{\text{10}}^{{\text{−}}\text{2}} $;当4号组模型叶片朝正方向转动时,力矩系数先增后减,在 $\theta \text{=20°} $ 时,$ C_{\rm{mx}}\text{=1.43×}{\text{10}}^{{\text{−}}\text{2}} $。
Z方向上的分力变化情况,2号组模型Z方向上的分力绕0上下波动,$\theta \text{=40°} $ 时,该系数仅为 $ \text{5.3×}{\text{10}}^{{\text{−}}\text{4}} $,且力的方向与Z轴负方向一致。当3号组叶片朝负方向转动时,Z轴上的分力方向与Z轴正方向一致,力的系数均未超过 $ \text{8.0×}{\text{10}}^{{\text{−}}\text{4}} $;当4号组叶片朝正方向转动时,分力方向不改变,分力系数先增加后缓慢下降,在 $\theta \text{=30°} $ 时达到最大值 $ \text{2.83×}{\text{10}}^{{\text{−}}\text{3}} $。当4号组叶片朝负方向转动时,Z轴上的分力方向与Z轴负方向一致,力的系数未超过 $ \text{9.0×}{\text{10}}^{{\text{−}}\text{4}} $;当4号组叶片朝正方向转动时,分力方向不改变,分力系数先增加后缓慢下降,在 $\theta \text{=30°} $时达到最大值 $ \text{2.63×}{\text{10}}^{{\text{−}}\text{3}} $。
2.2.2 20° 冲角下叶片转动对网板水动力特性的影响
多叶可控网板整体的升力系数在20° 冲角下达到最大,该冲角下叶片转动对网板水动力性能的影响分析如图6。
![]() 图 6 20° 冲角下叶片转角对网板水动力特性的影响Fig. 6 Influence of blade rotation angle of 20° attack angle on hydrodynamic characteristics otter board
图 6 20° 冲角下叶片转角对网板水动力特性的影响Fig. 6 Influence of blade rotation angle of 20° attack angle on hydrodynamic characteristics otter board首先,当网板的冲角增加至20° 时,叶片转动后的阻力系数也增大。5号组模型网板的阻力系数先下降后上升,在 $\theta \text{=}{\text{−}}\text{30°} $ 时,$ C_{{{\mathrm{d}}}}\text{=0.147} $;叶片朝正方向转动,阻力系数缓慢上升。6号和7号组的趋势相近,叶片朝正、负方向均会导致阻力系数下降。
其次,升力系数方面,叶片朝负方向转动时,5号、6号、7号3组模型的升力系数均先上升后下降,最大升力系数均出现在 $\theta \text{=}{\text{−}}\text{10°} $ 时,其中5号组的最大升力系数为1.05,6号和7号组分别为1.04和0.93。而当这3组模型的叶片朝正方向转动,升力系数均出现下降。
升阻比方面,该冲角下,叶片朝正方向转动后的升阻比均低于0° 冲角时对应叶片转角下的升阻比。而当3组模型的叶片朝负方向转动超过 10° 时,5号、6号、7号3组模型的升阻比反超2号、3号、4号组模型,并在 $\theta \text{=}{\text{−}}\text{20°} $ 时,达到最大升阻比,分别为5.25、4.25、3.96。
最后X轴力矩方面,5号组的力矩系数围绕0值波动范围小。6号和7号组的叶片朝负方向转动时,力矩系数变化幅度小,均在 $\theta \text{=}{\text{−}}\text{40°} $ 时,力矩改变方向;叶片朝正方向转动时,6号与7号组的力矩方向相反,但均随转角的增大而变大。
Z轴方向上的分力变化情况显示,20° 冲角下6号与7号组的叶片朝负方向转动,转角增大,分离系数绕0值波动,当 $\theta \text{=}{\text{−}}\text{40°} $ 时,分力改变方向。6号组模型叶片朝正方向转动,转角增大,分力方向与Z轴负半轴方向一致,且系数继续增大,当叶片朝正方向转动角度超过20° 后,Z轴上分力系数逐渐平稳;7号组该系数的变化形式与6号组相似,分力方向相反。
2.3 多叶可控网板压力流场分布及对比
2.3.1 1号组网板模型压力流场分布及对比
1号组在0° 冲角下的压力流场显示,3组叶片的前缘上部的正压区,越靠近前端的叶片面积越大,压力也越大;当冲角增大到20° 时,网板的凸面正压区转移至另一侧,同时凸面形成面积较大的负压区,从前中后依次向后,负压区的面积逐渐减小,涡流脱落位置靠近前叶,正压区主要集中在叶片的凹面中央,其中前叶的压力区域最集中;当冲角继续增大到30° 时,负压区中心向后移动,负压区逐渐均匀分布至网板凸面,涡流脱落位置靠后,另一侧凹面的正压区持续增长 (图7)。
![]() 图 7 1号组网板模型中心面压力流场分布Fig. 7 Pressure flow field of Group 1 on center surface of otter board model
图 7 1号组网板模型中心面压力流场分布Fig. 7 Pressure flow field of Group 1 on center surface of otter board model2.3.2 2号组网板模型压力流场分布及对比
2号组的上、下两端前叶和中叶朝正方向旋转10°,中叶与固定襟翼凸面前缘生成正压区,且固定襟翼正压区压力区域大于中叶,压力数值更高 (图8)。叶片间的水流通过受阻;叶片转角继续增加,前叶凹面正压区域面积增加,压力增大,背部负压区域面积增加,压力减小;当叶片转角超过30° 时,前叶背部的负压尾流覆盖中叶与固定襟翼区域。叶片朝负方向转动,3组叶片的凸面前缘产生正压区,且随转角增大,前叶正压区域面积与压力数值继续增大,中叶与固定襟翼的正压区逐渐减小至消失;冲角增大至 −30° 时,叶片间隙的流速增加,该状态下,所有叶片开合,网板间隙增大,流速加快,网板出现向收缩方向运动的趋势,与系数结果一致。
![]() 图 8 2号组网板模型中心面压力流场分布Fig. 8 Pressure flow field of Group 2 on center surface of otter board model
图 8 2号组网板模型中心面压力流场分布Fig. 8 Pressure flow field of Group 2 on center surface of otter board model2.3.3 3号和4号组网板模型压力流场分布及对比
3号组叶片朝正方向旋转10°,前叶凹面,中叶以及固定襟翼的凸面前缘产生正压区,前缘背部产生负压区;随转角增大,前叶背部的负压区域面积逐渐覆盖尾部区域,固定襟翼的前缘正压区面积先变大后减小至消失。叶片朝负方向转动,3组叶片均在背部前缘以及凹面尾部出现正压区,随转角增大,前叶正压区持续扩大,前叶负压区的影响范围覆盖后者 (图9);此状态下,上端叶片开合,网板间隙增大,流速加快,上端叶片周围压力减小,而另一端叶片未转动,周围流体压力大于上端,网板产生Z轴向上的分力,与系数结果一致。
![]() 图 9 3号组网板模型中心面压力流场分布Fig. 9 Pressure flow field of Group 3 on center surface of otter board model
图 9 3号组网板模型中心面压力流场分布Fig. 9 Pressure flow field of Group 3 on center surface of otter board model4号组网板模型压力流场分布与3号组大致相同,但由于开合叶片位置呈对称关系,因此,叶片朝负方向转动,下端叶片周围压力减小,4号组模型产生Z轴向下的分力,与系数结果一致 (图10)。
![]() 图 10 4号组网板模型中心面压力流场分布Fig. 10 Pressure flow field of Group 4 on center surface of otter board model
图 10 4号组网板模型中心面压力流场分布Fig. 10 Pressure flow field of Group 4 on center surface of otter board model2.3.4 5号组网板模型压力流场分布及对比
20° 冲角下,5号组模型叶片朝正方向旋转10°,前叶、中叶以及固定襟翼凹面产生的正压区依次减小;与0° 冲角下的压力流场相比,20° 冲角下叶片间的水流速度减小 (图11)。叶片转角增加,前叶凹面正压区域面积增加,压力增大,背部负压区域面积增加,压力减小;当叶片转角超过30°,前叶背部的负压尾流覆盖中叶与固定襟翼区域。叶片朝负方向转动,3组叶片的凸面前缘产生负压区,且随转角增大,前叶与中叶的负压区压力逐渐上升并转变为正压区;而固定襟翼的凹面正压区压强随叶片转角增大不断增加。冲角增大至 −30° 时,叶片间隙的流速增加,前叶凸面产生的正压区与固定襟翼凹面的正压区共同导致阻力上升,与系数结果一致。
![]() 图 11 5号组网板模型中心面压力流场分布Fig. 11 Pressure flow field of Group 5 on center surface of otter board model
图 11 5号组网板模型中心面压力流场分布Fig. 11 Pressure flow field of Group 5 on center surface of otter board model2.3.5 6号组和7号组网板模型压力流场分布及对比
6号组叶片朝正方向旋转10°,前叶凹面,中叶以及固定襟翼的凸面前缘均产生正压区;随着转角继续增大,中叶的正压区由凹面中央向尾部转移,最终脱落消失。叶片朝负方向转动,3组叶片均在背部前缘以及凹面尾部出现负压区,随着转角的增大,前叶负压区的压力数值不断增大 (图12);此状态下,上端叶片开合方向与叶片转动方向相反,上端叶片周围压力增大,而另一端叶片未转动,周围流体压力小于上端,网板产生Z轴向下的分力,与系数结果一致。
![]() 图 12 6号组网板模型中心面压力流场分布Fig. 12 Pressure flow field of Group 6 on center surface of otter board model
图 12 6号组网板模型中心面压力流场分布Fig. 12 Pressure flow field of Group 6 on center surface of otter board model7号组网板模型压力流场分布与6号组大致相同,但由于开合叶片位置呈对称关系,因此,当叶片朝负方向转动,下端叶片周围压力减小,7号组模型产生Z轴向下的分力,与系数结果一致 (图13)。
![]() 图 13 7号组网板模型中心面压力流场分布Fig. 13 Pressure flow field of Group 7 on center surface of otter board model
图 13 7号组网板模型中心面压力流场分布Fig. 13 Pressure flow field of Group 7 on center surface of otter board model3. 讨论
3.1 网板控制策略设计
目前,网板研究的主要热点方向是增强水动力性能,进而提高经济效益。自数值仿真和水槽渔具模型实验等技术手段的兴起与应用以来,对传统形态的网板,如矩形网板、椭圆型网板、双曲面网板以及立式V形曲面网板等,在结构上的调整优化已经非常充分[28]。在降低网板阻力系数的研究上,Zhuang等[29]通过在双曲面网板面板展向上开缝,有效降低了网板的阻力系数。Leifsson等[30]基于局部代理模型,模拟网板调整多个结构参数以优化水动力的行为,减少了计算次数,并达到设计效果。Lee等[24]也曾尝试利用疏水材料覆盖网板表面,实验发现网板的阻力下降了21.3%。对于扩张力的提升研究,Shen等[31]在立式曲面网板基础上设计的带翼端板的高升力网板,发现弯曲度为25%,0.10倍翼弦宽度的翼端板的模型升力系数达到2.48。You等[32]在此基础上设计了双叶型高升力网板,在22° 冲角下,最大升力系数为1.78,提高升力的同时不失稳定性。而学者们以增大拖网网口扩张程度为研究目的,在提高捕捞效率的同时往往也带来新的问题。无法调节扩张力的网口,除了增加兼捕,误捕的风险,在收网和放网两个阶段中,扩大网口也意味着拖网需要过滤更多水体,增加阻力和网具的磨损消耗。同时对于拖网捕捞过程中,释放的曳纲长度往往是固定的,且作业过程中几乎不会调整曳纲、手纲与网板的固结点位置,因此传统网板在拖网系统沉浮状态中只扮演配重的角色,增加下沉速度和深度,无法提供上升的力[33]。
此外,Sala等[34]设计的Clark-Y型网板的升阻比为3.6,优于母型矩形V形网板,并进行了海上测试。而黄小双等[35]和孔祥洪等[36]基于美国国家航空咨询委员会 (National advisory committee for aeronautics, NACA) 翼型设计的低阻力仿生江豚为水下渔具设计提供了新的思路。本研究中,与传统网板姿态控制策略不同[37],多叶可控网板以不改变纲缆与网板的固结点为前提,通过组合转动叶片的位置和转动角度实现对网口状态的调整。多叶可控网板的P50-3翼型剖面具有良好的流体形状优势,在不同的叶片状态下均能表现出一定水动力性能。可针对作业流程和目标渔获的特点,为拖网提供如图14所示的,包括扩张、收缩、上升以及下沉4种控制状态[24]。当网口深度达到捕捞水层而处于待扩张状态,以20° 冲角下的多叶可控网板为例,此时可设定两端叶片朝负方向转动10°,以达到增加网板扩张力,增大网口扩张范围。在目标鱼完全进入网口后,继续保持扩口扩张程度会增加燃油的消耗,此时拖网处于待收缩状态,可继续增加叶片在负方向上的转角以降低阻力系数和升力系数,达到减小阻力和收缩网口的效果,减少渔获逃逸,提升捕捞效率。当网口扩张程度足够大,而作业水层需要调整时,拖网处于待上升或待下沉状态,可按6号和7号组模型状态设定叶片转角,以实现拖网上浮和下沉的运动控制。
3.2 不足之处
本研究设计的多叶可控网板,是对网板调整自身结构以改变水动力的初步探索。不同于传统网板调整姿态的方式,多叶可控网板在不改变结固点的前提下,通过两端叶片的转动以达到运动趋势的变化。在这个过程中,对于叶片转动方向的分析讨论是其他可变翼襟网板研究中所欠缺的[38];值得一提的是,不同冲角下,叶片受水流作用的部位也有所改变,本实验结果显示两组冲角下,相同叶片转角对网板的水动力影响有较大差异,而对于更多不同冲角下的状态分析是本研究没有涉及到的,后续研究中应加入更多组冲角以拓展分析广度,还应加入用户自定义程序 (User defined functions, UDF) 以分析叶片转动的动态变化过程以揭示水动力变化形势[30]。此外,本研究对于网板控制网口扩张程度与捕捞深度的探讨只基于网板自身的运动趋势,暂未加入曳纲与手纲对其作用关系分析,实际情况应更为复杂,后续的研究将加入水槽实验,进一步测试,并合理规划控制策略。
4. 结论与展望
本研究通过数值模拟仿真,利用翼型的流体优势[39-40],设计了多叶可控网板,对其水动力性能相关参数进行了计算分析,将其周围压力流场进行可视化,并探究了多叶可控网板对网口状态的控制策略的可行性。主要结论如下:
1) 1号组模型在20° 冲角下出现最大升力系数0.88,最大升阻比出现在5° 冲角时,为8.85;并且该状态下网板在竖直方向上的分力和X轴的旋转力矩受冲角影响程度小。
2) 2号、3号和4号组网板模型叶片朝负方向转动会导致升力系数减小,叶片朝正方向转动,升力系数先增后减,在20° 冲角时达到最大值0.32;网板叶片朝正方向转动,3号组网板模型会产生竖直向上的分力,4号组网板模型则产生竖直向下的分力。
3) 与0° 冲角状态下不同,5号、6号和7号组网板模型叶片朝正方向转动会导致网板升力不断减小,叶片朝负方向转动,升力系数先增后减,在 −10° 转角时达到最大值1.05;网板叶片朝正方向转动,6号组网板会产生竖直向下的分力,7号组网板会产生竖直向上的分力。
4) 20° 冲角下,多叶可控网板的叶片朝负方向转动,不仅能够减小阻力系数,一定范围内还能提高升力系数,有效提升网板的升阻比;上端或者下端叶片朝正方向转动,网板能够获得垂直方向上的运动趋势,以达到调整作业水层的目的。
多叶可控网板的设计思路符合现代捕捞装备智能化的发展方向。根据作业阶段特征改变阻力和扩张力以有效降低渔船的燃油消耗;垂直方向上的移动控制不但能够提高捕捞的概率和效率,更能在关键时刻避免拖网系统对海底的接触。因此,本研究通过改善拖网网板的生态友好性,有利于远洋渔业经济和环境的可持续发展。
-
![]()
图 1 蜕壳死亡的克氏原螯虾 (左) 和蜕壳后的虾皮 (右)
Figure 1. Molting dead crayfish (left) and molting crayfish shell (right)
![]()
图 2 不同钙、镁离子浓度对克氏原螯虾蜕壳死亡率的影响
同时期数据中上标不同字母者之间差异显著 (P<0.05),后图同此
Figure 2. Effects of different Ca2+ and Mg2+ concentrations on molting mortality of P. clarkii
Means with different superscript letters are significantly different (P<0.05). The same case in the following figures
![]()
图 3 不同pH和溶解氧对克氏原螯虾蜕壳死亡率的影响
Figure 3. Effects of different pH values and dissolved oxygen concentrations on molting mortality of P. clarkii
![]()
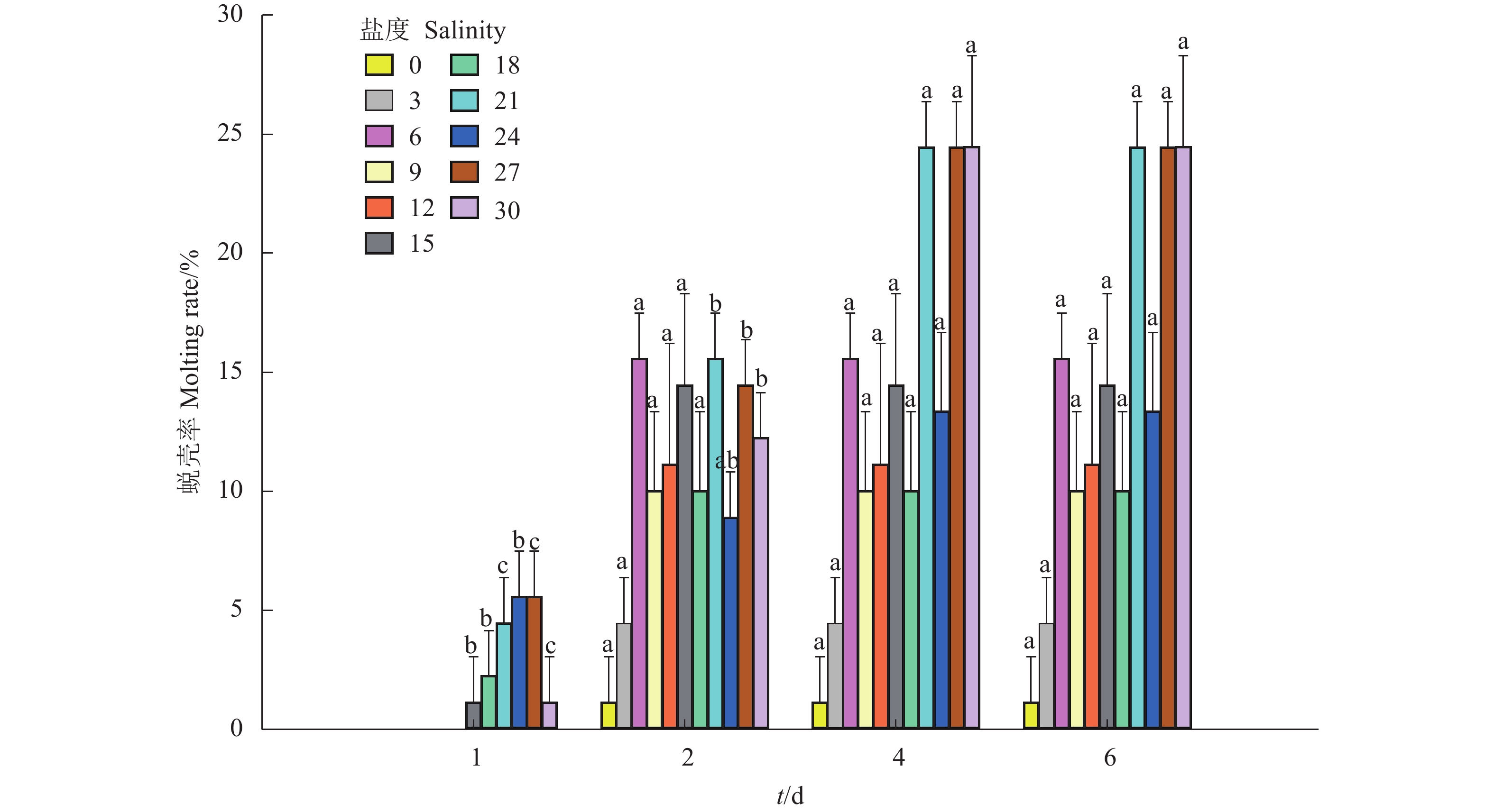
图 4 不同盐度对克氏原螯虾蜕壳率的影响
Figure 4. Effects of different salinities on molting rate of P. clarkii
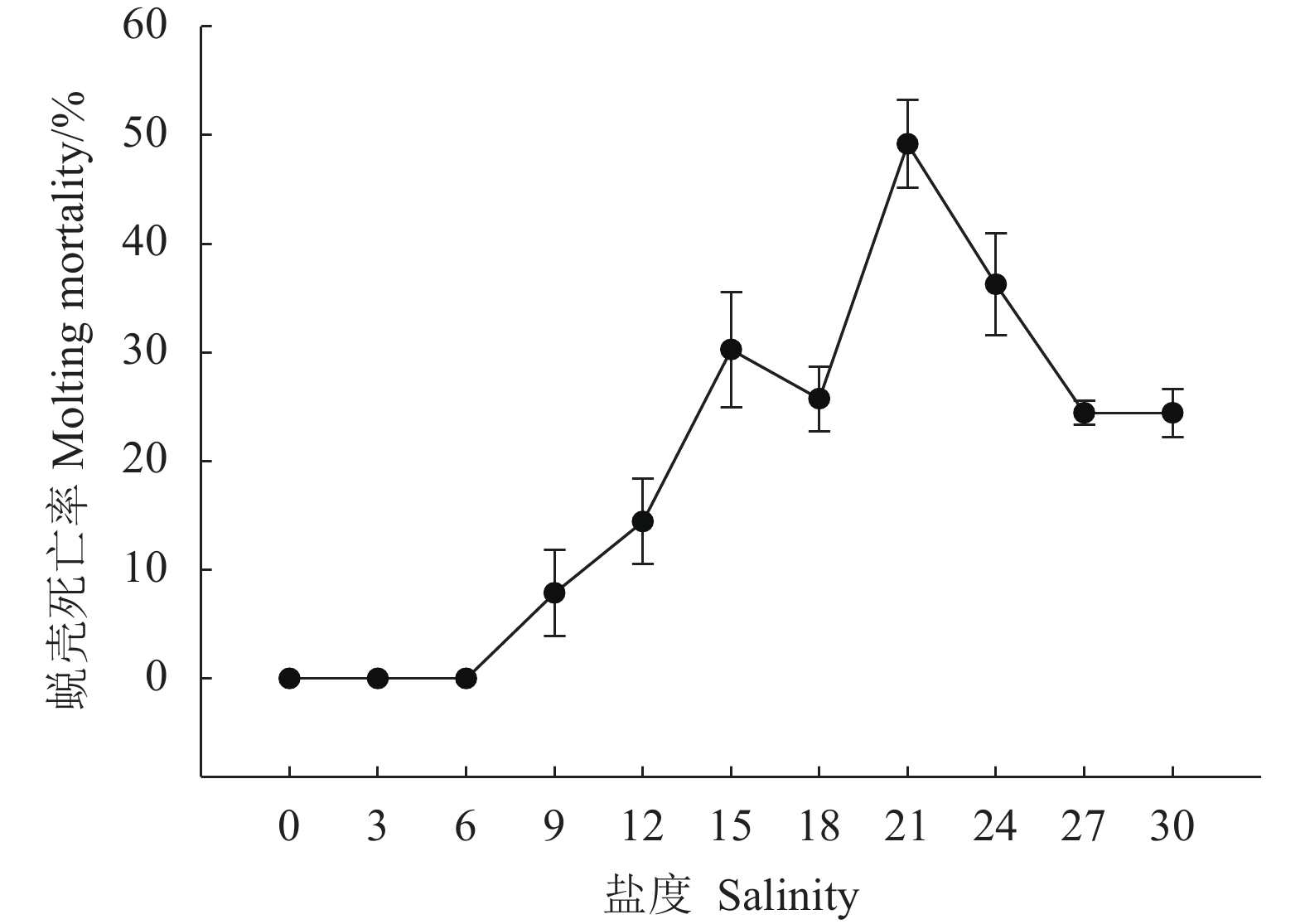
![]()
图 5 不同盐度对克氏原螯虾实验末期的蜕壳死亡率的影响
Figure 5. Effects of different salinities on molting mortality rate of P. clarkii at end of experiment
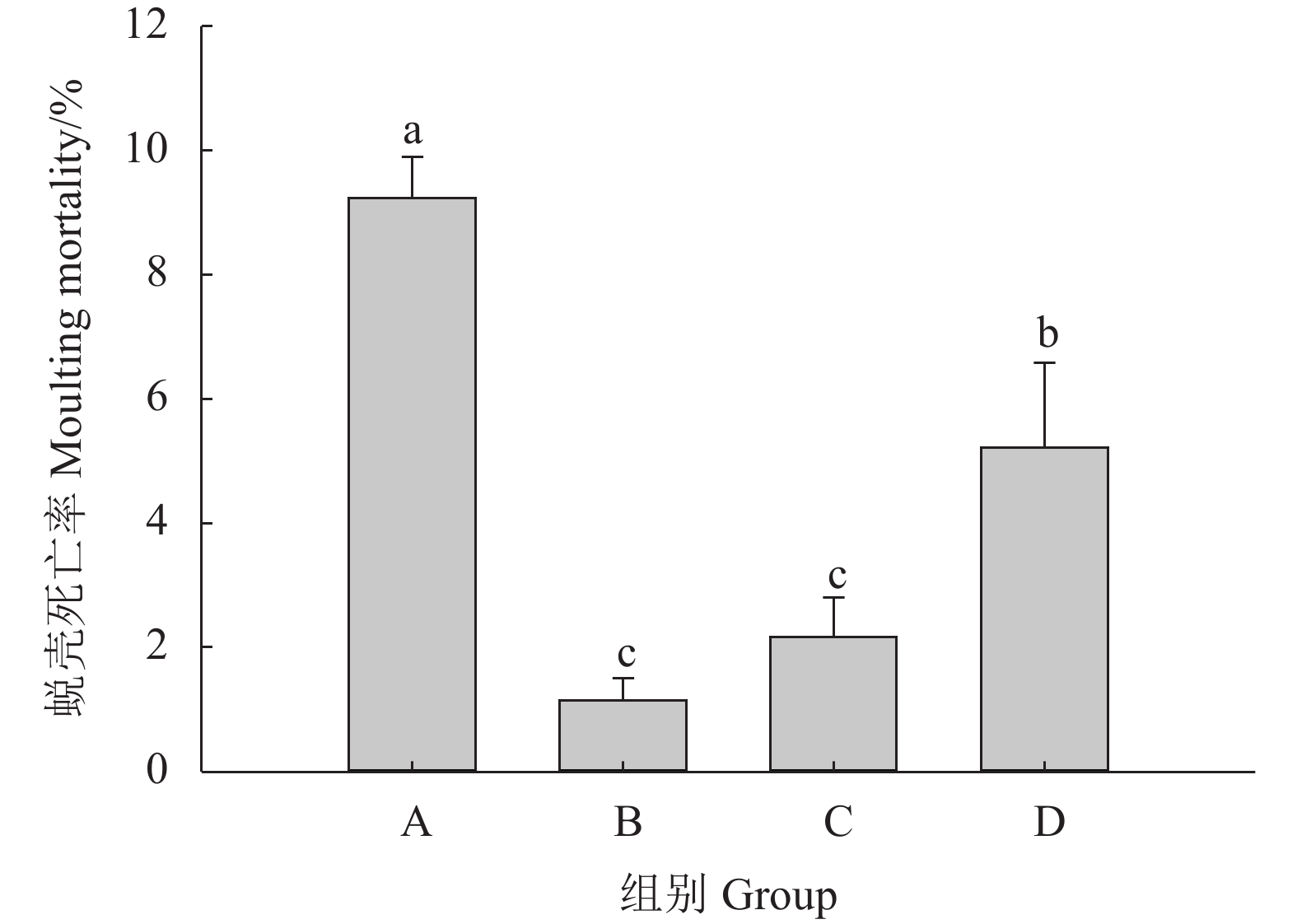
表 1 不同钙、镁离子浓度对克氏原螯虾生长及蜕壳的影响
Table 1 Effects of different Ca2+ and Mg2+ concentrations on decidua of P. clarkii
指标
IndexA组
Group AB组
Group BC组
Group CD组
Group D末体质量 Final body mass/g 1.28±0.04b 1.99±0.16a 2.00±0.27a 1.83±0.19a 成活率 Survival rate/% 68.89±3.85b 86.67±3.34a 85.56±5.09a 74.45±3.85b 蜕壳率 Molting rate/% 56.67±3.34b 82.22±1.92a 78.89±1.92a 68.89±1.92b 注:同行数据 (平均值±标准差) 上标字母不同者之间表示存在显著差异 (P<0.05)
Note: Values (${\overline{X}\pm \rm{SD} } $) within the same row with different superscript letters are signiifcantly different (P<0.05). 下载: 导出CSV
下载: 导出CSV
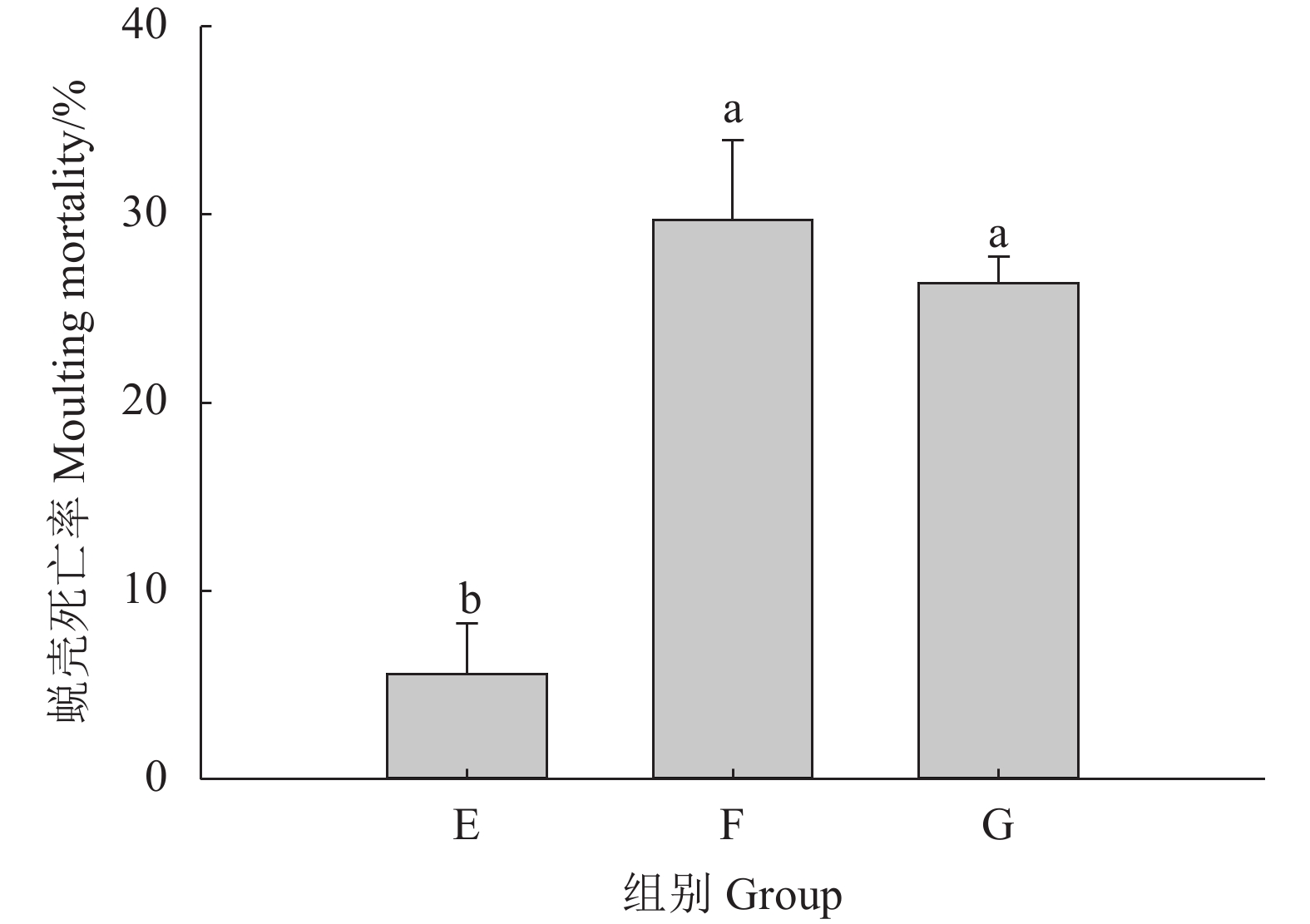
表 2 不同pH和溶解氧对克氏原螯虾生长及蜕壳的影响
Table 2 Effects of different pH values and dissolved oxygen concentrations on decidua of P. clarkii
指标
IndexE组
Group EF组
Group FG组
Group G末体质量
Final body mass/g42.37±1.10a 35.25±1.31b 33.44±0.99b 成活率
Survival rate/%85.56±5.09a 73.33±3.34b 71.11±5.09b 蜕壳率
Molting rate/%81.11±3.84a 58.89±5.09b 44.44±5.09c 注:同行数据 (平均值±标准差) 上标字母不同者之间表示存在显著差异 (P<0.05)Note: Values (${\overline{X}\pm \rm{SD} } $) within the same row with different superscript letters are signiifcantly different (P<0.05).
下载: 导出CSV
表 3 不同盐度对克氏原螯虾死亡率的影响
Table 3 Effects of different salinities on mortality rate of P. clarkii
% 盐度
Salnity1天
1 d2天
2 d4天
4 d6天
6 d8天
8 d10天
10 d12天
12 d0 0.00±0.00 1.11±1.11CD 2.22±1.11F 2.22±1.11E 2.22±1.11F 2.22±1.11H 2.22±1.11H 3 0.00±0.00c 1.11±1.11cCD 5.56±2.94cEF 13.33±1.93bD 23.33±1.95aCD 25.57±2.94aF 25.57±2.94aF 6 0.00±0.00c 0.00±0.00cD 8.89±1.11bDE 8.89±1.11bD 16.67±3.34aE 16.67±1.93aG 16.67±1.93aG 9 0.00±0.00d 0.00±0.00dD 3.33±1.93cdEF 7.78±1.11cDE 20.00±3.33cDE 26.67±1.93aF 26.67±1.94aEF 12 0.00±0.00d 0.00±0.00dD 2.22±1.11dF 8.89±1.11cD 25.56±1.11bCD 31.11±1.11aEF 31.11±1.11aDE 15 0.00±0.00c 0.00±0.00cD 4.44±1.11cEF 12.22±3.85bD 25.56±2.94aCD 30.00±1.92aEF 30.00±1.92aDEF 18 0.00±0.00e 1.11±1.11deCD 4.44±1.11dEF 12.22±3.85cD 20.00±1.92bDE 34.44±1.11aDE 34.44±1.11aCD 21 0.00±0.00f 7.78±1.11eAB 17.78±1.11dC 20.00±3.33dC 30.00±1.92cC 40.00±1.92bC 50.00±1.92aB 24 0.00±0.00e 5.57±2.94eB 13.33±1.93dCD 21.11±5.09cC 28.89±1.11bC 36.67±1.93aCD 36.67±1.93aC 27 0.00±0.00g 11.11±1.11fA 24.44±1.11eB 38.89±1.92dB 54.44±1.11cB 90.00±1.92bB 100.00±0.00aA 30 1.11±1.11e 4.44±1.11eBC 35.56±2.94bA 68.89±5.09cA 86.67±3.85bA 100.00±0.00aA 100.00±0.00aA 注:上标小写字母不同者表示同行数据 (平均值±标准差) 之间存在显著差异 (P<0.05),上标大写字母不同者表示同列数据 (平均值±标准差) 之间存在显著差异 (P<0.05)Note: Values (${\overline{X}\pm \rm{SD} } $) with different lowercase letters within the same row are signiifcantly different (P<0.05), while those with different uppercase letters within the same column are signiifcantly different (P<0.05).
下载: 导出CSV
-
[1] SMART A C, HARPER D M, MALAISSE F, et al. Feeding of the exotic Louisiana red swamp crayfish, Procambarus clarkii (Crustacea, Decapoda), in an African tropical lake: Lake Naivasha, Kenya[J]. Hydrobiologia, 2002, 488(1/2/3): 129-142.
[2] 许星鸿. 关于几种经济虾类的养殖技术之一: 克氏螯虾的生物学特性及人工养殖技术[J]. 中国水产, 2001(11): 38-39. doi: 10.3969/j.issn.1002-6681.2001.10.022 [3] 彭刚, 殷悦, 周春霞. 克氏原螯虾池塘主养技术[J]. 水产养殖, 2009, 30(1): 30-31. doi: 10.3969/j.issn.1004-2091.2009.01.016 [4] 唐建清, 宋胜磊, 潘建林, 等. 克氏原螯虾对几种人工洞穴的选择性[J]. 水产科学, 2004, 23(5): 26-28. doi: 10.3969/j.issn.1003-1111.2004.05.008 [5] DONG C, BAI S, DU L. Temperature regulates circadian rhythms of immune responses in red swamp crayfish Procambarus clarkii[J]. Fish Shellfish Immunol, 2015, 45(2): 641-647. doi: 10.1016/j.fsi.2015.05.025
[6] 安振华, 王柳富, 李晴. 虾青素对微囊藻毒素MC-LR胁迫下克氏原螯虾的生长、繁殖及免疫力的影响[J]. 水产养殖, 2017, 38(1): 30-34. doi: 10.3969/j.issn.1004-2091.2017.01.007 [7] 肖鸣鹤, 肖英平, 吴志强, 等. 养殖密度对克氏原螯虾幼虾生长、消化酶活力和生理生化指标的影响[J]. 水产学报, 2012, 36(7): 1088-1093. [8] MATTHEW L, HANS L, ELLEN H. Control of methyl farnesoate synthesis in the mandibular organ of the crayfish Procambarus clarkia: evidence for peptide neurohormones with dual functions[J]. Int J Invertebr Reprod, 2011, 16(1/2/3): 165-168.
[9] MANFRIN C, TOM M, MORO G D, et al. The eyestalk transcriptome of red swamp crayfish Procambarus clarkii[J]. Gene, 2015, 557(1): 28-34. doi: 10.1016/j.gene.2014.12.001
[10] 赵朝阳, 周鑫, 邴旭文, 等. 饥饿对克氏原螯虾亲虾消化酶活性及部分免疫指标的影响[J]. 大连海洋大学学报, 2010, 25(1): 85-87. doi: 10.3969/j.issn.1000-9957.2010.01.018 [11] 李京昊, 成永旭, 王海锋, 等. 不同条件对克氏原螯虾幼虾摄食生物絮凝的影响[J]. 南方水产科学, 2018, 14(3): 58-64. doi: 10.3969/j.issn.2095-0780.2018.03.007 [12] 董君焰. 蜕壳气味对克氏原螯虾行为及单胺类含量的影响[D]. 扬州: 扬州大学, 2014: 1-13. [13] 唐宁, 何晓瑾, 何晓冬, 等. 复方中草药对克氏原螯虾生长和脱壳的影响[J]. 淡水渔业, 2010, 40(1): 70-73. doi: 10.3969/j.issn.1000-6907.2010.01.013 [14] 方春林, 邓勇辉, 余智杰, 等. 克氏原螯虾生物学特性的研究[J]. 江西水产科技, 2010(3): 18-20. doi: 10.3969/j.issn.1006-3188.2010.03.006 [15] 冯依才. 浅谈对虾的生长与两大类影响因素[J]. 当代水产, 2017, 42(12): 58-59. doi: 10.3969/j.issn.1674-9049.2017.12.016 [16] 章爱如, 董福健, 朱云清, 等. 虾蟹养殖要点: 补钙固壳抗应激[J]. 当代水产, 2016, 41(10): 79-79. [17] 刘金生, 王军, 岳武成, 等. 水体pH对中华绒螯蟹幼蟹蜕壳生长及其相关基因表达的影响[J]. 淡水渔业, 2016, 46(4): 96-100. doi: 10.3969/j.issn.1000-6907.2016.04.016 [18] 黄海涛. 温度、盐度、溶解氧、氨氮、亚硝酸盐氮对拟穴青蟹蜕壳的影响[D]. 湛江: 广东海洋大学, 2011: 1-13. [19] MUGNIER C, SOYEZ C. Response of the blue shrimp Litopenaeus stylirostris to temperature decrease and hypoxia in relation to molt stage[J]. Aquaculture, 2005, 244(1/2/3/4): 315-322.
[20] 刘其根, 李应森, 陈蓝荪. 克氏原螯虾的生物学[J]. 水产科技情报, 2008, 35(1): 21-23. doi: 10.3969/j.issn.1001-1994.2008.01.011 [21] 刘存歧, 刘丽静, 王军霞, 等. 盐碱地渗水钙镁离子对凡纳滨对虾酶活力的影响[J]. 动物学杂志, 2007, 42(2): 129-133. doi: 10.3969/j.issn.0250-3263.2007.02.025 [22] 岳彩锋. 生长环境和食物对克氏原螯虾幼体生长发育的影响[D]. 武汉: 华中师范大学, 2009: 2-15. [23] 郭春雨, 管越强, 刘波兰, 等. 温度、pH和盐度对克氏原螯虾鳃Na+-K+-ATPase活性的影响[J]. 动物学杂志, 2007, 42(6): 96-102. doi: 10.3969/j.issn.0250-3263.2007.06.016 [24] BONVILLAIN C P, RUTHERFORD D A, KELSO W E. Effects of environmental hypoxia on population characteristics of red swamp crayfish Procambarus clarkiiin the Atchafalaya River Basin, Louisiana[J]. Hydrobiologia, 2015, 743(1): 309-319. doi: 10.1007/s10750-014-2049-4
[25] 慕峰. 水体盐度及Ca浓度对克氏原螯虾代谢、生长及蜕壳的影响研究[D]. 上海: 上海海洋大学, 2007: 4-51. [26] 陶易凡, 强俊, 王辉, 等. 高pH胁迫对克氏原螯虾的急性毒性和鳃、肝胰腺中酶活性及组织结构的影响[J]. 水产学报, 2016, 40(11): 1694-1704. [27] 任信林, 凌武海, 纪翠萍. 环境因子对克氏原螯虾养殖的影响[J]. 水产科学, 2009, 28(11): 710-712. doi: 10.3969/j.issn.1003-1111.2009.11.024 [28] 黄桂香. 水产养殖池塘中溶解氧的变化及调控[J]. 现代农业科技, 2014(17): 295-295. doi: 10.3969/j.issn.1007-5739.2014.17.175 [29] 赵明森. 淡水小龙虾的生物学特性及其养殖技术[J]. 当代水产, 2006, 31(18): 37-40. [30] 金彩霞, 潘鲁青. 盐度变化对克氏原螯虾渗透调节影响机制的初步研究[J]. 水生生物学报, 2008, 32(6): 894-899. [31] 李庭古. 盐度对克氏原螯虾幼虾生长的影响[J]. 水产科学, 2009, 28(8): 465-467. doi: 10.3969/j.issn.1003-1111.2009.08.011 [32] 张曹进, 姚国兴, 吴国钧, 等. 克氏原 (螫) 虾对盐度适应性试验[J]. 水产养殖, 2011, 32(7): 35-37. [33] 季鹏飞. 克氏原螯虾在盐胁迫条件下差异化的microRNA表达谱的研究[D]. 南京: 南京大学, 2014: 1-15. [34] 朱春华. 盐度对南美白对虾生长性能的影响[J]. 水产科技情报, 2002, 29(4): 166-168. doi: 10.3969/j.issn.1001-1994.2002.04.004 -
期刊类型引用(7)
1. 陈浩,吉宏武,张迪,刘书成,宋文奎,郝记明. 克氏原螯虾虾青蛋白A2基因克隆、组织分布及原核表达. 食品与生物技术学报. 2024(02): 63-72 .  百度学术
百度学术
2. 苏禹,王力玄,孟泳岐,马源潮,鲁耀鹏,张泽龙,郑佩华,李军涛,冼健安,刘存歧,王冬梅. 环境因素对克氏原螯虾生长与繁育的影响. 中国饲料. 2024(21): 74-82 . 百度学术
3. 赵明光,冯广朋,陈建华,王海华,张燕萍,徐维康. 主要环境因子对克氏原螯虾生长发育的影响研究进展. 江西水产科技. 2023(06): 28-35 . 百度学术
4. 欧琳,张余,陈晓芳,周刚,黄宇宏,陈蕾,肖调义,刘巧林. 基于线粒体Cytb和COⅠ基因的洞庭湖区养殖克氏原螯虾遗传多样性分析. 水产科技情报. 2022(03): 137-142 . 百度学术
5. 黎兰诗,戴习林. 盐度对不同蜕皮时期罗氏沼虾生理生化及蜕皮相关基因表达的影响. 南方农业学报. 2022(08): 2302-2311 . 百度学术
6. 韩财安,李安东,周美玉,廖怀生. 小龙虾幼苗培育关键技术. 江西水产科技. 2022(05): 30-31+34 . 百度学术
7. 韦永春,程顺,贾永义,迟美丽,刘士力,郑建波,李飞,刘一诺,顾志敏. 不同pH对红螯螯虾胚胎离体孵化的影响. 渔业现代化. 2021(06): 64-71 . 百度学术
其他类型引用(6)

















计量
- 文章访问数: 5090
- HTML全文浏览量: 2425
- PDF下载量: 88
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
