
Effect of high-carbohydrate diet on growth performance, feed utilization, glucose and lipid metabolism of GIFT Oreochromis niloticus
-
摘要:
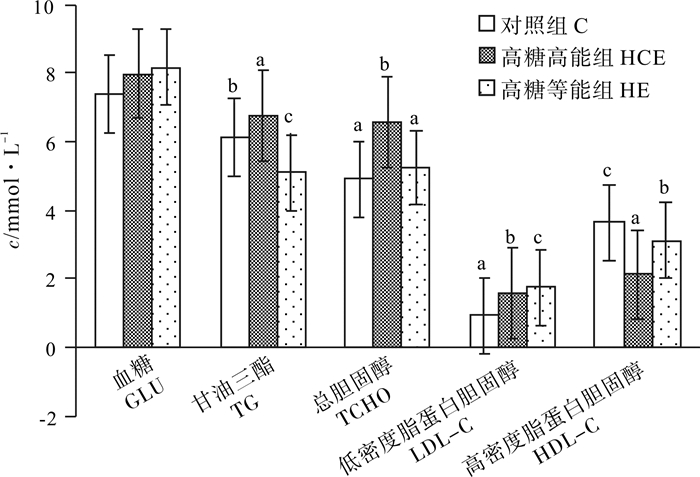
为探讨高糖饲料对吉富罗非鱼(GIFT Oreochromis niloticus)生长性能、饲料利用和糖脂代谢的影响, 设置糖水平为34%的对照饲料(C组, 能量水平13.68 kJ · g-1)、糖水平为48%的高糖高能饲料(HCE组, 能量水平16.03 kJ · g-1)和高糖等能饲料(HE组, 能量水平13.81 kJ · g-1), 在室内循环水系统中饲养初始体质量(20.34±0.42)g的吉富罗非鱼幼鱼60 d。结果表明, HCE组和HE组的增重率、特定生长率、肝脏粗蛋白均显著低于C组(P < 0.05), 饲料系数、肝脏粗脂肪、肝糖原、总糖表观消化率、血清低密度脂蛋白胆固醇和高密度脂蛋白胆固醇均显著高于C组(P < 0.05);HE组增重率、全鱼粗脂肪、血清甘油三酯和总胆固醇均显著低于HCE组(P < 0.05);高糖组肝脏葡萄糖激酶显著高于C组, HE组葡萄糖-6-磷酸脱氢酶、苹果酸酶、脂肪酸合成酶活性最高, C组最低(P < 0.05);由肝脏组织切片看出, 高糖饲料引起吉富罗非鱼肝脏细胞肿大变形, 出现空泡, HCE组对肝脏损伤更加显著。研究结果表明, 吉富罗非鱼能够耐受48%糖水平的饲料, 但长期摄食导致鱼体糖脂代谢紊乱, 尤其影响脂肪代谢, 造成鱼体脂肪蓄积, 肝功能损伤, 不利于生长与健康。
Abstract:To investigate the effects of high-carbohydrate diet on the growth performance, feed utilization, glucose and lipid metabolism of GIFT Oreochromis niloticus, we had fed the GIFT juveniles [initial body mass (20.34±0.42) g] with normal diet (C diet, 34% cholesterol level, dietary energy of 13.68 kJ · g-1), high-carbohydrate and high-energy diet (HCE diet, 48% cholesterol level, dietary energy of 16.03 kJ · g-1) and high-carbohydrate isoenergy diet (HE diet, dietary energy of 13.81 kJ · g-1) in indoor circulating water system for 60 d.The results show that the weight gain rate, specific growth rate, liver crude protein contents of HCE and HE groups were lower than those of C group significantly (P < 0.05);while HCE and HE groups had significantly higher feed conversion ratio, crude fat content and glycogen content of liver, apparent digestibility of carbohydrate, low density lipoprotein cholesterol (LDL-C) and high density lipoprotein cholesterol (HDL-C).The weight gain rate, whole body fat content, serum triglyceride (TG), total cholesterol (T-CHO) of HE group were lower than those of HCE group significantly (P < 0.05).The activities of glucokinase (GK) of HCE and HE group were higher than those of C group, and glucose-6-phosphate dehydrogenase (G-6-PDH), malic enzyme (ME), fatty acid synthase (FAS) of HCE were the highest in the three groups, and those of the C group were the lowest (P < 0.05).High carbohydrate groups induced swelling of liver cells and appearance of vacuoles.Hepatic injury was more serious in HCE group.In conclusion, GIFT can tolerate the feed with 48% carbohydrate content, but that is not conducive to the growth and health of GIFT, leading to fat accumulation and liver injury, disorder of glucose and lipid metabolism especially fat metabolism.
-
浮游植物是海洋生态系统中最基础的一个环节,作为该系统最重要的初级生产者,在海洋生态系统的物质循环和能量流动中起着重要的作用。浮游植物的种类组成和数量变动都将直接或者间接地制约海洋生产力的发展,从而影响海洋生物资源的开发和利用[1-5]。珠江口南沙海域位于广州市最南端,濒珠江出海口伶仃洋,属咸淡水的交汇区域,同时受到地表径流和海洋潮汐的影响,生态环境复杂,形成独特的海洋生物群落。近年来,随着周边地区经济社会的快速发展,以及滩涂围垦、航道疏浚和养殖等人类活动规模不断扩大,使得海水的富营养化加剧,极大地影响了该海区生态群落结构的稳定性,增大了发生赤潮的危险性,珠江口海域已成为中国赤潮的多发区之一[6-7]。
目前,对珠江河口及近海浮游生物的群落特征的研究已有报道[8-12],但是对珠江口内海域研究的比较少,特别是人为扰动最为频繁的珠江口南沙区周围的海域。文章通过对2015年4月(丰水期)和11月(枯水期)珠江口南沙海域生态综合调查采集的浮游植物样品进行鉴定分析,初步研究了该海域海洋浮游植物的种类组成、优势种、细胞丰度的分布及多样性水平,旨在为研究该海域生产力水平、海洋生态系统的群落结构和环境质量状况提供基础数据。
1. 材料与方法
1.1 采样和分析
2015年4月(丰水期)和11月(枯水期)在珠江口南沙区周围的海域(113°29′52″E~113°47′19″E,22°28′10″N~22°51′23″N)进行浮游植物样品采集,丰水期和枯水期都布设了20个调查站位(S1~S20,图 1)。调查站点均位于珠江口南沙海域人类活动扰动频繁海域,覆盖了从近口淡水、咸淡水段到口外海水段的河口典型生境特征。浮游植物样品用浅水Ⅲ型浮游生物网自底至表垂直拖网采集,并当场加入甲醛溶液保存。带回实验室后浓缩至50 mL,于显微镜下进行鉴定与计数,测定分析其种类组成、数量、分布、优势度、多样性指数和均匀度等。
![]() 图 1 珠江口南沙海域浮游植物调查示意图Figure 1. Sampling sites of phytoplankton in the Nansha sea area of Pearl River Estuary
图 1 珠江口南沙海域浮游植物调查示意图Figure 1. Sampling sites of phytoplankton in the Nansha sea area of Pearl River Estuary使用YSI便携式温盐深仪现场采集各站位盐度、温度、溶解氧(DO)、pH和水深等物理环境数据;同时现场采集海水样品,带回实验室后分析水体中的溶解性无机氮(TIN)、活性磷酸盐(PO4-P)和叶绿素a(Chl-a)的含量。浮游植物样品的采集和分析均按《海洋监测规范》(GB17378—2007)和《海洋调查规范-海洋生物调查》(GB12763.6—2007)[13-14]中规定的方法进行。
1.2 评价方法
浮游植物优势度(Y)应用以下公式计算:
$ Y = \frac{{{n_i}}}{N} \times {f_i} $
式中ni为第i种的个体数;fi是该种在各站中出现的频率;N为所有站每个种出现的总个体数,以Y>0.02作为优势种。
采用Shannon-Wiener[15]、Pielou[16]测定浮游植物的多样性指数(H′)、均匀度指数(J′),其计算公式分别为:
$ \begin{array}{l} H' =-\sum\limits_{i = 1}^S {{P_i}{\rm{lo}}{{\rm{g}}_2}{P_i}} \\ J' = \frac{{H'}}{{{\rm{lo}}{{\rm{g}}_2}S}} \end{array} $
式中S为样品中的种类总数;Pi为第i种的个体数与总个体数的比值。
浮游植物与环境因子的相关性分析采用SPSS 19.0统计软件分析;典范对应分析(canonical correspondence analysis,CCA)应用CANOCO 4.0软件进行,排序结果用物种-环境因子关系的双序图表示。另外,采用孙军等[17]的浮游植物多样性的评判阈值(表 1)对浮游植物多样性进行等级评价。
表 1 浮游植物多样性指标的评判阈值Table 1. Threshold value evaluation for diversity indices of phytoplankton多样性指标
diversity index很低
very low
(Ⅰ)低
low
(Ⅱ)一般
normal
(Ⅲ)高
high
(Ⅳ)很高
very high
(Ⅴ)种类数species number <15 15~24 25~34 35~44 >45 多样性指数(H′)Shannon-Wiener diversity index <1.5 1.5~2.49 2.5~3.49 3.5~4.5 >4.5 均匀度指数(J′)Pielous evenness index <0.68 0.68~0.73 0.74~0.79 0.80~0.88 >0.88 2. 结果与分析
2.1 浮游植物物种组成
共鉴定出浮游植物44个属76种,隶属于硅藻门、甲藻门、绿藻门、蓝藻门和裸藻门5大门类。其中丰水期共鉴定出浮游植物5门37属68种(包括变种和变形),硅藻门种类最多(24属39种),占总种数的57.35%;其次是绿藻门(7属15种),占总种数的22.06%;甲藻门有2属7种,占总种数的10.29%;裸藻门有3属5种,占总种数的7.35%;蓝藻门有1属2种,占总种数的2.94%。S1站鉴定出的浮游植物种类最多(38种),而S17站鉴定出的浮游植物种类最少(6种),但都是属于硅藻类。各站位中新月筒柱藻(Cylindrotheca closterium)、中肋骨条藻(Skeletonema costatum)、虹彩圆筛藻(Coscinodiscus oculusiridis)在每个站位中都有分布。此外,奇异棍形藻(Bacillaria paradoxa)也分布比较广泛(表 2)。
表 2 珠江口南沙海域浮游植物物种组成Table 2. Species composition of phytoplankton in the Nansha sea area of Pearl River Estuary门类
phylum总种类数
total species number丰水期(5月)
wet season (May)枯水期(11月)
dry season (November)种类
species比例/%
percentage种类
species比例/%
percentage种类
species比例/%
percentage硅藻Bacillariophyta 39 51.32 39 57.35 33 55.00 甲藻Pyrrophyta 15 19.74 7 10.29 14 23.33 绿藻Chlorophyta 15 19.74 15 22.06 8 13.33 蓝藻Cyanophyta 2 2.63 2 2.94 2 3.33 裸藻Euglenophyta 5 6.58 5 7.35 3 5.00 总计total 76 - 68 - 60 - 枯水期共鉴定出浮游植物5门44个属60种(包括变种和变形),其中还是硅藻门种类最多(22属33种),占总种数的55.00%;其次是甲藻门(11属14种),占总种数的23.33%;绿藻门有6属8种,占总种数的13.33%;裸藻门有3属3种,占总种数的5.00%;蓝藻门有2属2种,占总种数的3.33%。中肋骨条藻、虹彩圆筛藻、有棘圆筛藻(C.spinosus)、布氏双尾藻(Ditylum brightwelli)在大部分调查站位中均有分布(表 2)。可见珠江口南沙海域丰水期和枯水期的浮游植物物种组成相似,都是硅藻类占主体。
2.2 浮游植物种类的分布
浮游植物的区域分布具有季节差异,以丰水期较明显。丰水期S1站和S2站分别有38和30种,而S17站只有6种,表现为由河口内向外海递减的特征。枯水期该水域各站位浮游植物种类数比较接近(12~16种),也表现为由河口内向外海递减的特征(图 2)。
![]() 图 2 珠江口南沙海域各站位浮游植物种类数Figure 2. Number of phytoplankton species of each station in the Nansha sea area of Pearl River Estuary
图 2 珠江口南沙海域各站位浮游植物种类数Figure 2. Number of phytoplankton species of each station in the Nansha sea area of Pearl River Estuary2.3 浮游植物细胞丰度的变化及分布
丰水期调查结果表明(图 3),浮游植物细胞丰度为(361.52~2 000.73)×104个· L-1,平均为651.91×104个· L-1,主要集中在万顷沙旁的水道,以S17站最高,其次是S19站(912.59×104个·L-1),S14站最低。细胞丰度组成大部分以硅藻为主,硅藻类平均细胞丰度为636.46×104个· L-1,占该海域浮游植物平均细胞丰度的97.63%,不同站位硅藻细胞丰度占比范围为92.95%~100.00%,其中S14站和S17站均只检出硅藻(图 3)。
![]() 图 3 珠江口南沙海域各站位浮游植物细胞丰度Figure 3. Abundance of phytoplankton cells of each station in the Nansha sea area of Pearl River Estuary
图 3 珠江口南沙海域各站位浮游植物细胞丰度Figure 3. Abundance of phytoplankton cells of each station in the Nansha sea area of Pearl River Estuary枯水期调查结果表明,浮游植物细胞丰度为(4.00~698.40)×104个· L-1,平均为129.21×104个· L-1,也主要集中于万顷沙周围的水道,以S19站最高,其次是S6站(430.13×104个· L-1),S13站最低。细胞丰度组成大部分以硅藻为主,硅藻平均细胞丰度为117.25×104个· L-1,占海域浮游植物平均细胞丰度的90.74%,不同站位硅藻细胞丰度占比为4.88%~100.00%;此外,绿藻在S11站、S13站和S14站都占有较高的比例,裸藻在S17站也同样有较高的比例。
从浮游植物细胞丰度的平均值来看,丰水期的细胞丰度明显高于枯水期。两个时期从各个站位的细胞丰度来看,都是离岸的站位(S6~S9,S18~S20)高于近岸的站位(S1~S5,S10~S13),呈现出近岸向离岸递增的趋势。
2.4 优势种的组成及变化
调查海域浮游植物优势种结果显示,丰水期优势种2种,均为硅藻,新月筒柱藻的优势度最高(0.85);其次是中肋骨条藻,优势度为0.09。枯水期优势种3种,也均为硅藻,其中萎软海链藻(Thalassiosira mala)的优势度最高(0.25);其次是中肋骨条藻(0.20);有棘圆筛藻最低(0.02)。通过对丰水期和枯水期的对比,发现该海域的优势种均为硅藻类,其中中肋骨条藻是两时期的共有优势种,但也表现出季节性的演替现象,丰水期的优势种新月筒柱藻在枯水期更替为萎软海链藻。
2.5 群落多样性
珠江口南沙海域丰水期的H′为0.18~3.32,平均值为1.64;J′为0.07~0.72,平均值为0.40;而该海域枯水期的H′为0.59~2.56,平均值为1.58;J′为0.16~0.64,平均值为0.41(图 4)。丰水期时S4站的H′和J′最高;其次是S5站,分别是2.93和0.68;最低是S17站。枯水期时S1站、S3站、S4站和S10站的H′和J′都较其他站位高;而S9站和S17站则相对较低。
表 3为珠江口南沙海域浮游植物多样性评价结果,表中百分数表示各多样性指标隶属评判阈值的站位数占总站位数的百分比。可见丰水期时种类数、H′和J′都基本处于很低的水平,丰水期的多样性评判结果处于Ⅰ级水平。枯水期时由于H′略高,因此枯水期的多样性评判结果处于Ⅱ级水平。
表 3 珠江口南沙海域浮游植物多样性评价Table 3. Diversity evaluation for phytoplankton in the Nansha sea area of Pearl River Estuary季节
season很低
very low
(Ⅰ)低
low
(Ⅱ)一般
normal
(Ⅲ)高
high
(Ⅳ)很高
very high
(Ⅴ)总评价
general evaluation丰水期wet season 种类数(S)species number 87.50% 12.50% Ⅰ 多样性指数(H′)Shannon-Wiener diversity index 62.50% 37.50% Ⅰ 均匀度指数(J′)Pielous evenness index 100% Ⅰ 枯水期dry season 种类数(S) species number 40% 60% Ⅱ 多样性指数(H′)Shannon-Wiener diversity index 45% 45% 10% Ⅱ 均匀度指数(J′)Pielous evenness index 100% Ⅰ 2.6 浮游植物丰度与环境因子的关系
将浮游植物细胞丰度与水温、盐度进行Pearson相关回归系数分析(表 4),可以看出,丰水期浮游植物细胞丰度与水温呈显著负相关性(P < 0.05);枯水期浮游植物细胞丰度与盐度呈显著正相关性(P < 0.01)。可见温度和盐度对该海域浮游植物的群落结构有较大影响,而其他环境因子与细胞丰度相关性不明显。
表 4 珠江口南沙海域浮游植物细胞丰度与环境因子的相关性Table 4. Correlation between cell density of phytoplankton and environmental factors in the Nansha sea area of Pearl River Estuary季节
season水温
temperature盐度
salinity溶解氧
DO酸碱度
pH丰水期wet season -0.670* -0.209 0.208 0.378 枯水期dry season 0.491 0.597** -0.233 0.133 注:*.P < 0.05;* *.P < 0.01 2.7 典范对应分析
在CCA排序图中(图 5),丰水期与第1排序轴呈正相关的环境因子为盐度(SAL)、温度(TEM)、总氮(DIN)、透明度(SD)和磷酸盐(PO4),相关系数分别为0.978、0.813、0.786、0.567和0.490;与第1排序轴呈负相关的环境因子为溶解氧和pH,相关系数分别为-0.528和-0.018;与第2排序轴呈最大正相关和负相关的环境因子分别为磷酸盐和pH,相关系数分别为0.436和-0.623。枯水期与第1排序轴呈正相关关系的环境因子依次为温度、磷酸盐、盐度、总氮和透明度,相关系数分别为0.846、0.740、0.711、0.702和0.553;与第1排序轴呈负相关的环境因子为pH和溶解氧,相关系数分别为-0.754和-0.439;与第2排序轴呈最大正相关的环境因子为pH,相关系数0.430;透明度与第2排序轴呈最大负相关,相关系数为-0.804。结果表明丰水期和枯水期环境因子对浮游植物群落结构的影响作用存在差异,丰水期环境因子的重要性依次为盐度、温度、总氮、pH、透明度、溶解氧和磷酸盐,枯水期则为温度、透明度、磷酸盐、盐度、总氮、pH和溶解氧。
![]() 图 5 丰水期(a)、枯水期(b)珠江口南沙海域浮游植物物种-环境因子典范对应分析排序图Figure 5. Ordination diagram of CCA based on phytoplankton species and environmental factors in the Nansha sea area of Pearl River Estuary
图 5 丰水期(a)、枯水期(b)珠江口南沙海域浮游植物物种-环境因子典范对应分析排序图Figure 5. Ordination diagram of CCA based on phytoplankton species and environmental factors in the Nansha sea area of Pearl River Estuary丰水期浮游植物优势种新月筒柱藻和中肋骨条藻分别位于第三象限和第四象限,新月筒柱藻与温度、总氮和磷酸盐呈负相关,与pH呈正相关;而中肋骨条藻与盐度、温度、总氮、磷酸盐、透明度和pH均呈正相关。枯水期浮游植物优势种萎软海链藻位于第三象限,与pH和透明度呈正相关,与温度和磷酸盐呈负相关;中肋骨条藻位于第四象限,与温度、透明度、磷酸盐、盐度和总氮呈正相关,与pH呈负相关;有棘圆筛藻则位于第一象限,与温度、透明度、pH、磷酸盐、盐度和总氮均呈正相关。
3. 讨论
3.1 浮游植物种类组成
根据珠江口海域浮游植物的特点,可划分为河口类、近岸类、外海类等3大生态类群[8],由于受到珠江水系径流汇入的影响,调查海域的盐度比较低,沿岸带来的营养盐较丰富,因此该海域的浮游植物大量生长和繁殖,种类的组成跨度较大,包括沿岸性种和淡水性种。浮游植物种类组成季节变化明显,丰水期共获得浮游植物37属68种(包括变种和变形),而枯水期共获得浮游植物44属60种(包括变种和变形)。与2008年~2010年在该海域的调查结果相比[10],当前该海域中硅藻类在总种类数中所占比例呈下降趋势,绿藻类所占比例呈现明显上升趋势。
3.2 浮游植物细胞丰度与优势种分布特征
调查发现,该海区浮游植物细胞丰度平面分布在丰水期和枯水期都呈现出由近岸区向离岸区递增的趋势(图 3)。位于龙穴岛东面的S6站、S7站、S8站和S9站以及离岸区的S19站和S20站都是浮游植物平均细胞丰度的高值区,较其他站位平均细胞丰度高,分析发现丰水期离岸高值区站位主要是由硅藻类新月筒柱藻大量繁殖造成的;枯水期离岸高值区站位主要是由硅藻类的萎软海链藻和中肋骨条藻繁殖引起。
优势种在群落中起着主导作用,决定着群落结构中的能量传递和物质循环的途径和方式,支配着群落结构的演替方向[18]。丰水期时的优势种为新月筒柱藻和中肋骨条藻,这2种硅藻都属于广温广盐性藻类,其中新月筒柱藻是一种分布广、高繁殖力的底栖硅藻,也是引发赤潮的一种重要藻类[19],在丰水期其细胞丰度占总细胞丰度的85.15%,特别是S17站,该藻细胞丰度百分比为97.70%;枯水期时的优势种是萎软海链藻、中肋骨条藻和有棘圆筛藻,种类相对较多且优势种发生了更替,由以新月筒柱藻向萎软海链藻和中肋骨条藻优势度并重演替。丰水期时优势种以淡水种和广温广盐种为主,但枯水期时主要是以广温广盐种为主,其中有棘圆筛藻是低温高盐种,这与各个时期由于地表径流引起的水体温度和盐度变化相适应。
丰水期的新月筒柱藻和枯水期的萎软海链藻及中肋骨条藻的细胞丰度分布格局呈现近岸向离岸递增的趋势,这同该海区浮游植物总细胞丰度的分布格局相一致。同时该海域的优势种新月筒柱藻、中肋骨条藻和萎软海链藻都是属于赤潮藻类,说明该海域富营养化严重,有较大的赤潮爆发风险[19]。
3.3 浮游植物群落结构多样性
珠江口南沙海域丰水期和枯水期浮游植物种类数的分布都表现为由河口处向外递减的特征(图 2、图 3),这是由于该海域生长繁殖的主要是以淡水种和广温广盐种为主的藻类[8],盐度的升高反而成为抑制淡水种生长繁殖的因子。藻类平均种类数丰水期略多于枯水期,这是由于丰水期地表径流的增加,导致低盐度的水域扩大,从而使淡水藻类和广温广盐藻类生长繁殖区间扩大,其中丰水期离河口最近的S1站的种类数明显高于同时期的各站位种类数,离河口较远的S19站和S20站的种类数都比较低;而枯水期各站位的种类数较接近(12~16种),这是由于枯水期地表径流的减小,高盐度的海水入侵,导致淡水藻类生长繁殖区间减小,因此以广温广盐类藻类为主。多样性评价结果表明枯水期高于丰水期,但总体而言,两时期的多样性和均匀度都处于较低水平,表明珠江口南沙段海域水质状况一般,水环境受到了一定程度的污染,水域生态系统较为脆弱。
3.4 浮游植物群落结构与水温、盐度的关系
浮游植物群落结构与盐度、水温、营养盐、海流、光照等环境因子存在一定的相关性[20-22],其中水温、盐度是影响浮游植物分布的重要因素[23-25]。广州南沙海域浮游植物群落是以硅藻类为主体的浮游植物群落结构,丰水期和枯水期的细胞丰度都呈现出近岸向离岸递增的趋势。从浮游植物丰度和环境因子的关系,并结合CCA分析可以看出,温度、盐度仍是影响该海区浮游植物的主要环境因子。丰水期细胞丰度与水温、盐度呈负相关性;枯水期细胞丰度则与水温、盐度呈正相关性。由于丰水期浮游植物丰度主要受优势种新月筒柱藻丰度的影响,而新月筒柱藻与温度和盐度呈负相关(图 5-a),这可能是丰水期浮游植物丰度与温度和盐度呈负相关的原因。而枯水期优势种中肋骨条藻和有棘圆筛藻以及浮动弯角藻等多数常见种与温度、盐度呈正相关(图 5-b),而这些种类是枯水期浮游植物丰度的重要组成部分,这可能是浮游植物丰度与温度、盐度呈正相关的主要原因。此外,浮游植物种类不同,其适应的水温和盐度的范围也不同。丰水期时优势种以淡水种和广温广盐种为主,盐度的升高反而抑制了淡水种的生长繁殖,导致细胞丰度的分布格局跟温盐的分布呈现负相关;而枯水期时优势种主要以广温广盐种为主,水温的变化范围为24.7~26.6 ℃,此时细胞丰度与水温呈正相关,表明该水域水温适合大部分藻类的生长,并且随着水温的升高,浮游植物细胞丰度呈增长的趋势,盐度的变化范围为0.14~21.39,适合广温广盐类的藻类生长繁殖,因此枯水期细胞丰度的分布格局与温盐分布格局呈正相关。
4. 小结
1) 调查结果表明,珠江口南沙海域共鉴定浮游植物5门44属76种,种类的组成跨度较大,包括淡水种和广温广盐种,其中硅藻为最主要优势种群。丰水期优势种有2种,分别为新月筒柱藻(0.85)和中肋骨条藻(0.09);枯水期优势种有3种,分别为萎软海链藻(0.25)、中肋骨条藻(0.20)和有棘圆筛藻(0.02)。
2) 丰水期和枯水期浮游植物平均细胞丰度分别为651.91×104个· L-1和129.21×104个· L-1,分别以S17站和S19站最高,而细胞丰度最低的站位位置接近(分别是S14站和S13站)。丰水期和枯水期的细胞丰度分布都表现为近岸向离岸递增的趋势。
3) H′和J′的平均值在丰水期分别为1.64和0.40,而在枯水期分别为1.58和0.41。调查海区的浮游植物群落结构多样性总体处于较低水平,表明珠江口南沙海域水质状况一般,水环境受到了一定程度的污染。
4) 环境因子对珠江口南沙海域浮游植物群落结构有明显影响,丰水期主要影响因子为盐度、温度、总氮和pH;枯水期为温度、透明度、磷酸盐和盐度。
此研究采样频次偏少,今后应加强对其季节演替及动态变化的研究,以更好地理解人类活动干扰下珠江口生态系统的变化过程。
-
![]()
![]()
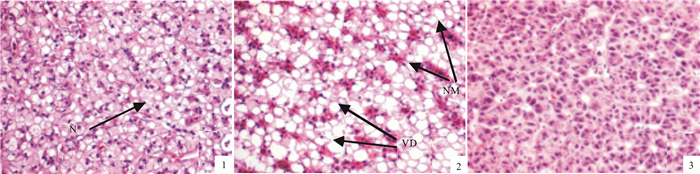
图 2 高糖饲料对吉富罗非鱼肝脏组织的影响
1.对照组(C组)肝脏切片(×200);2.高糖高能组(HCE组, ×200);3.高糖等能组(HE组, ×200); N.细胞核; NM.细胞核偏移; VD.空泡变性
Figure 2. Effect of high carbohydrate level in diets on hepatic tissue of GIFT O.niloticus
1.liver section from the control group (×200);2.liver section from the high-carbohydrate-high-fat group (×200); 3.liver section from high-carbohydrate-isoenergy group (×200);N.nucleus; NM.nucleus migration; VD.vacuolar degeneration
![]()
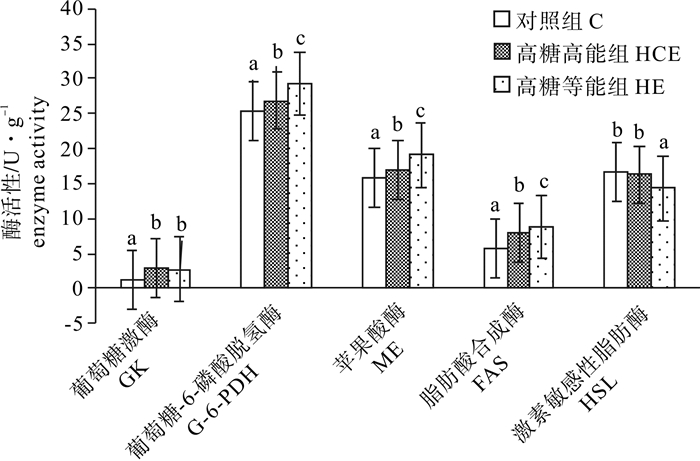
图 3 高糖饲料对吉富罗非鱼糖脂代谢相关酶活性及肝糖原的影响
Figure 3. Effect of high carbohydrate level in diets on liver activity of carbohydrate and lipid metabolism enzymes of GIFT O.niloticus
表 1 基础饲料组成及基本营养成分组成
Table 1 Formulation and proximate analysis of basal diets
成分
ingredient对照组
(C)高糖高能组
(HCE)高糖等能组
(HE)酪蛋白/% casein 29.00 29.00 29.00 明胶/% gelatin 7.00 7.00 7.00 糊精/% dextrine 34.00 48.00 48.00 玉米油/% corn oil 4.20 4.20 1.20 大豆油/% soybean oil 4.20 4.20 1.20 氯化胆碱/% choline chloride 0.25 0.25 0.25 维生素预混料①/% vitamin premix 1.00 1.00 1.00 无机盐预混料②/% mineral premix 2.00 2.00 2.00 磷酸二氢钙/% calcium phosphate 2.00 2.00 2.00 膨润土/% bentonite 1.60 1.60 1.60 微晶纤维素/% micro-cellulose 14.65 0.65 6.65 三氧化二钇/% Y2O3 0.1 0.1 0.1 营养成分 proximate composition 粗蛋白/% crude protein 28.93 28.63 28.73 粗脂肪/% crude lipid 8.23 8.23 2.35 总能/kJ·g-1 gross energy 13.68 16.03 13.81 总糖/% gross carbohydrate 34.11 49.01 48.78 注:营养水平中总能按粗蛋白23.64 kJ · g-1, 粗脂肪39.54 kJ · g-1, 总糖17.15 kJ · g-1计算, 其他均为实测值; ①维生素预混料可为每千克饲料提供, 维生素A 5 000 IU, 维生素D3 2 000 IU, 维生素E 60 mg, 维生素C 120 mg, 维生素K 5 mg, 维生素B2 20 mg, 维生素B1 5 mg, 维生素B6 10 mg, 烟酸120 mg, 泛酸钙10 mg, 叶酸1 mg, 生物素0.1 mg, 肌醇400 mg; ②矿物质预混料可为每千克饲料提供磷酸氢钙6 000 mg, L-乳酸钙6 540 mg, 硫酸铁42.5 mg, 硫酸镁1 340 mg, 磷酸氢钠1 744 mg, 氯化钠870 mg, 氯化铝3 mg, 碘化钾2.5 mg, 氯化钾1 500 mg, 氯化铜2 mg, 硫酸镁16 mg, 氯化钴20 mg, 硫酸锌60 mg
Note:Gross energy:crude protein 23.64 kJ · g-1, crude fat 39.54 kJ · g-1, total carbohydrate 17.15 kJ · g-1, and the others are measured values; ①The vitamin premix provids the following per kg of diets:VA 5 000 IU, VD3 2 000 IU, VE 60 mg, VC 120 mg, VK 5 mg, VB2 20 mg, VB1 5 mg, VB6 10 mg, nicotinic acid 120 mg, calcium pantothenate 10 mg, folic acid 1 mg, biotin 0.1 mg, inositol 400 mg; ②The mineral premix provids the following per kg of diets:Ca(HPO4)2 6 000 mg, Ca(CH3CHOHCOO)2 6 540 mg, FeSO4 42.5 mg, MgSO4 1 340 mg, NaHPO4 1 744 mg, NaCl 870 mg, AlCl3 3 mg, KIO3 2.5 mg, KCl 1 500 mg, CuCl2 2 mg, MnSO4 16 mg, CoCl2 20 mg, ZnSO4 60 mg. 下载: 导出CSV
下载: 导出CSV
表 2 高糖饲料对吉富罗非鱼生长性能和饲料利用的影响
Table 2 Effect of high carbohydrate level in diets on growth performance and feed utilization of GIFT O.niloticus
指标indicator 对照组
(C)高糖高能组
(HCE)高糖等能组
(HE)初始体质量/g IBM 20.20±0.28 19.80±0.06 20.62±0.51 终末体质量/g FBM 101.90±8.31c 90.21±1.39b 87.21±3.68a 增重率/% WGR 404.40±3.01c 340.47±2.11b 327.83±2.54a 特定生长率/%·d-1 SGR 2.97±0.13b 2.47±0.03a 2.46±0.09a 蛋白质效率 PER 2.15±0.11b 1.98±0.08a 2.01±0.10a 饲料系数 FCR 1.48±0.11a 1.88±0.16b 1.95±0.03b 肝体比/% HSI 2.93±0.07a 3.23±0.02b 3.01±0.16b 肥满度/% CF 3.54±0.17 3.98±0.18 3.41±0.29 脏体比/% VSI 9.77±0.12b 10.51±0.15c 9.31±0.11a 注:同行上标字母不同代表差异显著(P < 0.05);后表同此
Note:Values with different superscript letters have significant difference (P < 0.05).The same case in the following tables.
下载: 导出CSV
表 3 高糖饲料对吉富罗非鱼全鱼和肌肉常规成分的影响
Table 3 Effect of high carbohydrate level in diets on whole body and muscle composition of GIFT O.niloticus
% 成分
ingredient对照组
(C)高糖高能组
(HCE)高糖等能组
(HE)全鱼 whole fish 粗蛋白 crude protein 12.80±0.20 13.24±0.10 12.78±0.11 粗脂肪 crude lipid 6.42±0.11a 9.37±0.62b 4.97±0.54a 粗灰分 ash 4.23±0.31 3.88±0.33 4.18±0.16 水分 moisture 68.22±1.35 67.07±2.14 69.40±0.21 肌肉 muscle 粗蛋白 crude protein 18.91±0.43 18.32±0.65 18.14±0.58 粗脂肪 crude lipid 1.51±0.17 1.46±0.11 1.43±0.21 粗灰分 ash 1.18±0.25 1.25±0.02 1.23±0.07 水分 moisture 76.52±0.67 77.24±1.29 77.58±0.42 肝脏 liver 粗蛋白 crude protein 12.25±0.08b 11.21±0.09a 11.16±0.10a 粗脂肪 crude lipid 10.15±0.11a 12.21±0.09b 12.05±0.13b 糖原 glycogen 6.03±0.15a 8.74±0.11b 7.25±0.10c
下载: 导出CSV
表 4 高糖饲料对吉富罗非鱼营养物质表观消化率的影响
Table 4 Effect of high carbohydrate level in diets on apparent digestibility of nutrients of GIFT O.niloticus
% 成分
ingredient对照组
(C)高糖高能组
(HCE)高糖等能组
(HE)干物质 dry matter 65.32±0.12a 77.26±0.17b 76.15±0.13b 蛋白质 protein 95.65±0.19 96.06±0.21 95.87±0.11 脂肪 lipid 85.23±0.82b 84.59±0.74b 74.51±0.15a 总糖 carbohydrate 72.52±0.16b 65.71±0.83a 67.48±0.80a
下载: 导出CSV
-
[1] TENDENCIA E A, FERMIN A C, DELA PENA M R, et al.Effect of Epinephelus coioides, Chanos chanos, and GIFT tilapia in polyculture with Penaeus monodon on the growth of the luminous bacteria Vibrio harveyi[J].Aquaculture, 2006, 253(1/2/3/4):48-56. http://www.academia.edu/31952417/Integrated_mariculture_its_role_in...
[2] WANG Y, LIU Y J, TIAN L X, et al.Effects of dietary carbohydrate level on growth and body composition of juvenile tilapia, Oreochromis niloticus×O.aureus[J].Aquacult Res, 2005, 36(14):1408-1413. doi: 10.1111/are.2005.36.issue-14
[3] WILSON R P.Utilization of dietary carbohydrate by fish[J].Aquaculture, 1994, 124(1/2/3/4):67-80. https://www.sciencedirect.com/science/article/pii/0044848694903638
[4] ASADUZZAMAN M, WAHAB M A, VERDEGEM M, et al.Effects of carbohydrate source for maintaining a high C:N ratio and fish driven re-suspension on pond ecology and production in periphyton-based freshwater prawn culture systems[J].Aquaculture, 2010, 301(1/2/3/4):37-46. https://www.sciencedirect.com/science/article/pii/S0044848610000591
[5] 王菲, 李向飞, 李贵锋, 等.不同糖脂比对建鲤幼鱼生长、体组成、消化及糖酵解能力的影响[J].水产学报, 2015, 39(9):1386-1394. http://www.oalib.com/paper/5257186 [6] 蔡春芳, 陈立侨.鱼类对糖的利用评述[J].水生生物学报, 2006, 30(5):608-613. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [7] 赵艳景, 胡虹, 王颖.缢蛏多糖的提取及抗氧化作用研究[J].水产科学, 2010, 29(2):91-94. http://www.cqvip.com/qk/93386X/201002/34073523.html [8] ANDERSEN D E, REID S D, MOON T W, et al.Metabolic effects associated with chronically elevated cortisol in rainbow trout (Oncorhynchus mykiss)[J].Can J Fish Aquat Sci, 2011, 48(9):1811-1817. https://core.ac.uk/display/62587210
[9] PANSERAT S, MÉDALE F, BLIN C, et al.Hepatic glucokinase is induced by dietary carbohydrates in rainbow trout, gilthead seabream, and common carp[J].Am J Physiol Regul Integr Comp Physiol, 2000, 278(5):R1164-R1170. https://www.ncbi.nlm.nih.gov/pubmed/10801283
[10] DIAS J, RUEDA-JASSO R, PANSERAT S, et al.Effect of dietary carbohydrate-to-lipid ratios on growth, lipid deposition and metabolic hepatic enzymes in juvenile Senegalese sole (Solea senegalensis, Kaup)[J].Aquacult Res, 2004, 35(12):1122-1130. doi: 10.1111/are.2004.35.issue-12
[11] BAUTISTA J M, GARRIDO-PERTIERRA A, SOLER G.Glucose-6-phosphate dehydrogenase from Dicentrarchus labrax liver:kinetic mechanism and kinetics of NADPH inhibition[J].Biochim Biophys Acta, 1988, 967(3):354-363. doi: 10.1016/0304-4165(88)90098-0
[12] 刘泓宇, 毛义波, 谭北平, 等.饲料糖水平对不同食性鱼类生长及葡萄糖耐受能力的影响[J].水产学报, 2015, 39(12):1852-1862. https://www.cnki.com.cn/qikan-SCKX201306015.html [13] 吴凡, 文华, 蒋明, 等.饲料碳水化合物水平对奥尼罗非鱼幼鱼生长、体成分和血清生化指标的影响[J].华南农业大学学报, 2011, 32(4):91-95. doi: 10.7671/j.issn.1001-411X.2011.04.020 [14] 吴凡, 文华, 蒋明, 等.饲料碳水化合物水平对吉富罗非鱼幼鱼生长性能和血液主要生化指标的影响[J].西北农林科技大学学报(自然科学版), 2012, 18(12):8-14. http://www.cqvip.com/QK/90760A/201212/44415193.html [15] OTCHOUMOU A K, BLE C M, NIAMKE S L, et al.Effect of dietary carbohydrate:lipid ratio on growth performance and body composition of African catfish (Heterobranchus longifilis) fingerlings[J].Israeli J Aquacult-Bamidgeh, 2011, 63(2):169-180. http://lprapp14.fao.org/XML_Output/2012/IR/IR1250.xml
[16] AMANAT AL, AL-ASGAH N A.Effect of feeding different carbohydrate to lipid ratios on the growth performance and body composition of Nile tilapia (Oreochromis niloticus) fingerlings[J].Animal, 2001, 50(1):91-100. https://www.sciencedirect.com/science/article/pii/S0044848613002834
[17] ALI M Z, JAUNCEY K.Optimal dietary carbohydrate to lipid ratio in African catfish Clarias gariepinus (Burchell 1822)[J].Aquacult Int, 2004, 12(2):169-180. doi: 10.1023/B:AQUI.0000032065.28059.5b
[18] HILTON J W, DIXON D G.Effect of increased liver glycogen and liver weight on liver function in rainbow trout, Salmo gairdneri richardson:recovery from anaesthesia and plasma 35 S-sulphobromophthalein clearance[J].J Fish Dis, 1982, 5(3):185-195. doi: 10.1111/jfd.1982.5.issue-3
[19] LOPÉZ L M, OLMOS SOTO J, TREJO ESCAMILLA I A, et al.Evaluation of carbohydrate-to-lipid ratio in diets supplemented with Bacillus subtilis probiotic strain on growth performance, body composition and digestibility in juvenile white seabass (Atractoscion nobilis, Ayres 1860)[J].Aquacult Res, 2016, 47(6):1864-1873. doi: 10.1111/are.2016.47.issue-6
[20] HAIDAR M N, PETIE M, HEINSBROEK L T, et al.The effect of type of carbohydrate (starch vs.nonstarch polysaccharides) on nutrients digestibility, energy retention and maintenance requirements in Nile tilapia[J].Aquaculture, 2016, 463:241-247. doi: 10.1016/j.aquaculture.2016.05.036
[21] MOHAPATRA M, SAHU N P, CHAUDHARI A.Utilization of gelatinized carbohydrate in diets of Labeo rohita fry[J].Aquacult Nutr, 2003, 9(3):189-196. doi: 10.1046/j.1365-2095.2003.00243.x
[22] 刘迎隆. 不同添加量的糖对军曹鱼生长代谢的影响[D]. 青岛: 中国海洋大学, 2014: 17-21. http://cdmd.cnki.com.cn/Article/CDMD-10423-1014328654.htm [23] 王朝明, 罗莉, 张桂众, 等.饲料脂肪水平对胭脂鱼生长性能、肠道消化酶活性和脂肪代谢的影响[J].动物营养学报, 2010, 22(4):969-976. http://www.wenkuxiazai.com/doc/6befd47a4693daef5ef73dcb.html [24] STONE D J.Dietary carbohydrate utilization by fish[J].Rev Fish Sci, 2003, 11(4):337-369. doi: 10.1080/10641260390260884
[25] 崔志峰, 郝彦周, 孙卫明, 等.纤维素在鱼类饲料中适宜含量的研究概况[J].华北农学报, 2006, 21(12):79-81. http://www.cqvip.com/QK/91494X/2006F12/26868130.html [26] 刘波, 谢骏, 苏永腾, 等.高碳水化合物日粮对翘嘴红鲌生长、GK及GK mRNA表达的影响[J].水生生物学报, 2008, 32(1):47-53. http://www.wenkuxiazai.com/doc/343772e9551810a6f5248639.html [27] 缪凌鸿, 刘波, 戈贤平, 等.高碳水化合物水平日粮对异育银鲫生长、生理、免疫和肝脏超微结构的影响[J].水产学报, 2011, 35(2):221-230. http://cqvip.com/QK/90183X/201102/37030208.html [28] 蔡春芳. 青鱼(Mylopharyngodon pieces Richardson)和鲫(Carassius auratus)对饲料糖的利用及其代谢机制的研究[D]. 上海: 华东师范大学, 2004: 40-53. http://cdmd.cnki.com.cn/Article/CDMD-10269-2004087208.htm [29] QIANG J, YANG H, MA X Y, et al.Comparative studies on endocrine status and gene expression of hepatic carbohydrate metabolic enzymes in juvenile GIFT tilapia (Oreochromis niloticus)fed high-carbohydrate diets[J].Aquacult Res, 2014, 47(3):758-768. doi: 10.3389/fendo.2017.00073/full
[30] LOVELL T.Nutrition and feeding of fish[J].Aquaculture, 1989, 267(s1/2/3/4):13-70. doi: 10.1007%2F978-1-4757-1174-5
[31] GAO W, LIU Y J, TIAN L X, et al.Effect of dietary carbohydrate-to-lipid ratios on growth performance, body composition, nutrient utilization and hepatic enzymes activities of herbivorous grass carp (Ctenopharyngodon idella)[J].Aquacult Nutr, 2010, 16(3):327-333. doi: 10.1080/10641260390260884
[32] TRIGATTI B L, KRIEGER M, RIGOTTI A.Influence of the HDL receptor SR-BI on lipoprotein metabolism and atherosclerosis[J].Arterioscler Thromb Vasc Biol, 2003, 23(10):1732-1738. doi: 10.1161/01.ATV.0000091363.28501.84
[33] 陈孟瑶. 饲料中不同淀粉、蛋白质和脂肪水平对罗非鱼生长及生理生化指标的影响[D]. 厦门: 集美大学, 2014: 28-40. http://cdmd.cnki.com.cn/Article/CDMD-10390-1014354717.htm [34] 蔡春芳, 王永玲, 陈立侨, 等.饲料糖种类和水平对青鱼、鲫生长和体成分的影响[J].中国水产科学, 2006, 13(3):452-459. http://d.wanfangdata.com.cn/Thesis_Y609956.aspx [35] AMEER F, SCANDIUZZI L, HASNAIN S, et al.De novo lipogenesis in health and disease[J].Metabolism, 2014, 63(7):895-902. doi: 10.1016/j.metabol.2014.04.003
[36] ROLLIN X, MÉDALE F, GUTIERES S, et al.Short-and long-term nutritional modulation of acetyl-CoA carboxylase activity in selected tissues of rainbow trout (Oncorhynchus mykiss)[J].Br J Nutr, 2003, 89(6):803-810. doi: 10.1079/BJN2003844
[37] LIKIMANI T A, WILSON R P.Effects of diet on lipogenic enzyme activities in channel catfish hepatic and adipose tissue[J].J Nutr, 1982, 112(1):112-117. doi: 10.1007/s11745-010-3521-1
[38] JOCKEN J W E, LANGIN D, SMIT E, et al.Adipose triglyceride lipase and hormone-sensitive lipase protein expression in subcutaneous adipose tissue is decreased after an isoenergetic low-fat high-complex carbohydrate diet in the metabolic syndrome[J].Metab Clin Experiment, 2012, 61(10):1404-1412. doi: 10.1016/j.metabol.2012.03.017
[39] 孙君君, 卢荣华, 常志光, 等.糖对草鱼肝脂代谢关键基因转录水平的调控研究[J].水产科学, 2015, 12(11):696-701. https://www.wenkuxiazai.com/doc/3929704d8762caaedc33d468... [40] 蒋利和, 吴宏玉, 黄凯, 等.饲料糖水平对吉富罗非鱼幼鱼生长和肝脏代谢功能的影响[J].水产科学, 2012, 37(2):245-255. http://www.cnki.com.cn/Article/CJFDTotal-SCKX201507011.htm -
期刊类型引用(9)
1. 张良奎,林雅君,向晨晖,黄亚东,郑传阳,宋星宇. 粤港澳大湾区近海浮游植物群落结构特征及其驱动因素. 生态科学. 2024(04): 1-10 .  百度学术
百度学术
2. 高广银,李学恒,唐灵刚,李颖颖. 秋冬季浮游植物群落结构特征及其环境因子关系——以珠江口南沙海域为例. 绿色科技. 2024(20): 74-78+84 . 百度学术
3. 陈亮东,詹建坡,王庆. 珠江典型河段浮游植物群落结构特征及其对水质的指示作用. 南方水产科学. 2023(06): 1-10 . 本站查看
4. 蒋佩文,李敏,张帅,陈作志,徐姗楠. 基于环境DNA宏条码和底拖网的珠江河口鱼类多样性. 水生生物学报. 2022(11): 1701-1711 . 百度学术
5. 李桂娇,冯洁娉,叶锦韶. 南沙港海区水质变化与浮游植物群落更替特征. 生态科学. 2021(05): 164-171 . 百度学术
6. 姚艳欣,陈楠生. 珠江口及其邻近海域赤潮物种的生物多样性研究进展. 海洋科学. 2021(09): 75-90 . 百度学术
7. 陈思,李艺彤,蔡文贵,陈海刚,田斐,张林宝,张喆,郭志勋. 虾蟹混养池塘浮游植物群落结构的变化特征. 南方水产科学. 2020(03): 79-85 . 本站查看
8. 李亚军,王先明,程贤松,魏盟智,李江月,朱为菊,邓晓东. 海南岛海尾湾浮游植物群落结构及水质的调查. 热带生物学报. 2020(03): 257-265 . 百度学术
9. 李希磊,于潇,卢钰博,杨俊丽,崔龙波. 2015—2017年莱州湾招远海域扇贝养殖区浮游藻类群落变化. 南方水产科学. 2019(04): 11-18 . 本站查看
其他类型引用(4)





计量
- 文章访问数: 3251
- HTML全文浏览量: 1013
- PDF下载量: 887
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
