
Effects of poly-β-hydroxybutyrate (PHB) on immune and digestive indicators in hepatopancreas of Litopenaeus vannamei
-
摘要:
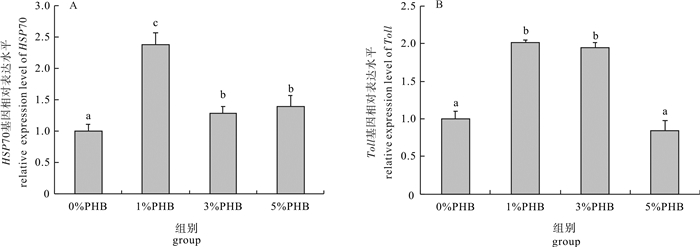
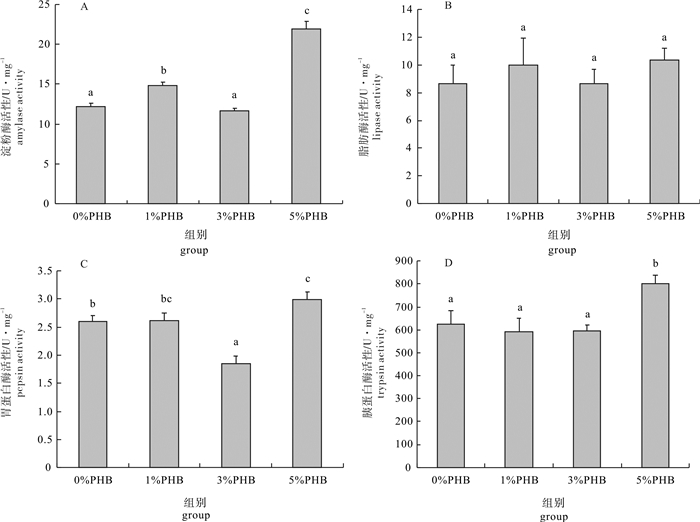
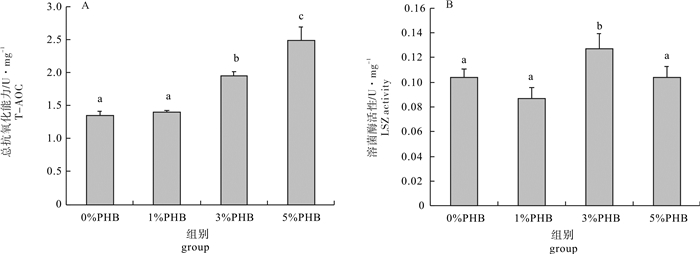
以平均体质量为(5.66±0.14)g的凡纳滨对虾(Litopenaeus vannamei)为研究对象, 分别投喂含聚β-羟基丁酸酯(PHB)质量分数为0%(对照组)、1%、3%和5%的对虾配合饲料, 饲养35 d后检测并比较不同水平PHB对凡纳滨对虾肝胰腺免疫和消化指标的影响。结果显示, 3%PHB添加组凡纳滨对虾的总抗氧化能力和溶菌酶活性显著高于对照组(P < 0.05)。Toll基因表达量随PHB含量的升高呈先上升后下降趋势, 其中1%和3%PHB添加组的表达量显著高于对照组(P < 0.05);对照组HSP70基因表达量显著低于其他处理组(P < 0.05)。5%PHB添加组的淀粉酶、胃蛋白酶和胰蛋白酶活性显著升高(P < 0.05), 而脂肪酶活性无显著变化(P>0.05)。由此可见, 饲料中添加适量的PHB有利于增强凡纳滨对虾肝胰腺的消化和非特异性免疫能力。
-
关键词:
- 凡纳滨对虾 /
- 聚β-羟基丁酸酯(PHB) /
- 肝胰腺 /
- 免疫 /
- 消化
Abstract:Litopenaeus vannamei of average weight of (5.66±0.14) g had been fed for 35 d with four different diets:0%, 1%, 3% and 5% poly-β-hydroxybutyrate (PHB) supplemented feed, and the immune and digestive indicators in hepatopancreas of shrimp were evaluated in this study.The results show that compared with the control group, the shrimps in 3% PHB group had higher total antioxidant capacity (T-AOC) and lysozyme (LSZ) activity (P < 0.05).The relative expression level of Toll gene increased but then decreased with increasing PHB content, and the expression levels in 1% and 3% PHB groups were higher than that in the control group significantly (P < 0.05).The relative expression levels of heat shock protein 70 (HSP70) gene in all three PHB treatment groups were higher than that in the control group significantly (P < 0.05).The amylase, pepsin and trypsin activities of 5% PHB group were significantly higher than those of the control group (P < 0.05), but the lipase activity showed no significant difference (P > 0.05).It is indicated that appropriate PHB content in diet can improve the immune and digestive abilities in hepatopancreas of shrimps significantly.
-
Keywords:
- Litopenaeus vannamei /
- poly-β-hydroxybutyrate (PHB) /
- hepatopancreas /
- immune /
- digestion
-
马面鲀(Navodon spp.)是东海区重要的海产经济鱼类,曾经是东海机轮拖网的主要捕捞对象之一[1]。东海马面鲀的产量主要包括绿鳍马面鲀(N.septentrionalis)和黄鳍马面鲀(N.xanthopterus)的渔获量[2-3]。20世纪50~60年代,东海绿鳍马面鲀的产量很少,1974年初在温台渔场发现大量的绿鳍马面鲀,吸引了众多生产渔船前来生产,绿鳍马面鲀迅速成为东海区新的捕捞对象[4]。东海区马面鲀有渔业统计记录的年份为1976年,当时马面鲀的产量主要以绿鳍马面鲀为主,其渔获量在东海区仅次于带鱼,逐渐位居经济捕捞品种的第二位,直至20世纪80年代末[5-6]。从1990年起黄鳍马面鲀开始形成渔汛,同期绿鳍马面鲀渔获量急速下降,东海区马面鲀渔汛转为以捕捞黄鳍马面鲀为主的局面[7]。黄鳍马面鲀的渔获量开始上升,1996年达最高峰,此后黄鳍马面鲀渔获量开始下降,在黄鳍马面鲀资源量下降的同时,东海的绿鳍马面鲀的资源则有所回升[1-2, 7]。
从1974年开始开发东海绿鳍马面鲀资源以来,马面鲀年间产量波动十分剧烈,这与不断增强的捕捞压力有关,也可能与马面鲀种群之间的饵料和栖息地竞争有关,此外,气候、水温等环境因素的变化也是马面鲀产量波动的主要影响因素,因此有必要结合气候环境的变化探讨东海马面鲀渔获量的变动原因。对此诸多学者已作了一些探讨,方瑞生等[8]研究钓鱼岛近海的涡旋及其与马面鲀渔场的关系时发现,马面鲀的产量与涡旋底部外洋水的上升强度呈负相关,当涡旋底部外洋水弱的年份产量偏高,反之,当涡旋底部外洋水强的年份产量偏低。许学龙等[9]在对东、黄海绿鳍马面鲀的资源评估时指出,由于传统的动态理论都把种群的初始期看作为最大,然后随着渔业的发展而逐步减少,这很难描述和解释马面鲀种群由小变大的动态过程,因此必须从东、黄海的生态系角度进行考察。王跃中等[10]研究东海带鱼渔获量变动时发现,渔获量变动不仅与捕捞压力的增长有关,还与海表水温、季风、陆地降雨和径流等气候环境的变化相联系。此研究主要利用东海马面鲀历年渔业统计资料和相应的气候环境资料来探讨捕捞效应和气候变化对其产量波动的影响,并在此基础上进一步预测东海马面鲀渔业的未来变化趋势。
1. 材料与方法
1.1 渔业统计资料
渔业统计资料含1976年~2006年的浙江省、福建省、江苏省和上海市的马面鲀渔获量和捕捞努力量统计数据。研究资料主要来自农业部水产司和渔业局等,包括1979年~2006年的《中国渔业统计年鉴》[2]、《中国渔业统计四十年(1949~1988)》[3]和《中国渔业统计汇编(1994~1998)》[11]等。渔船捕捞努力量统计数据含机动渔船和非机动渔船,单位用千瓦(kW)表示,其中非机动渔船捕捞努力量是按其单位捕捞努力量渔获量(CPUE)的年间变动趋势与该海区相同功率的机动渔船比例标准化而来。
1.2 气候变化资料
气候变化资料主要来自荷兰气候研究中心KNMI(Royal Netherlands Meteorological Institute)Climate Explorer(http://climexp.knmi.nl)和日本气象厅区域专业气象中心RSMC(Regional Specialized Meteorological Center)东京台风中心数据文档(http://www.jma.go.jp/)。主要的气候时间序列资料包含陆地降雨、季风和热带气旋影响指数。
因为缺乏东海沿岸主要河流的径流量数据,故用长江流域和东海沿岸地区的降雨量资料做替代。陆地降雨基于全球降水气候中心GPCC(Global Preciptiation Climatology Centre)1°×1°网格化的数据集,从KNMI Climate Explorer网站选择长江流域和东海沿岸地区下载。下载的陆地降雨量时间序列资料为长江流域和东海沿岸地区年平均降水量。
中国近海气候最明显的特征是冬、夏季风交替,因此取冬季和夏季2个季节的季风,夏季季风为每年的6月~8月,期间盛行西南风,冬季季风为每年的10月至翌年3月,期间主要盛行偏北风[12]。季风数据利用KNMI Climate Explorer从每月的2°×2℃OADS(Comprehensive Ocean-Atmosphere Data Set)获取。主要获取的季风为黄海海域(119°E~127°E,33°N~39°N)和东海海域(117°E~131°E,23°N~33°N)的海面风速数据。从获取的季风风速数据来看,明显存在上升趋势,RAMAGE[13]和CARDONE等[14]发现季风变化趋势主要是由于测量技术不断改进所引起,因此所有季风时间序列数据中的上升趋势都将被移除。
热带气旋时间序列数据基于RSMC东京台风中心下载的每6 h热带气旋分析数据。下载的数据范围位于东海海域(120°E~130°E,22°N~37°N)。热带气旋分析数据可用热带气旋影响指数(TCI)[15],TCI=∑T(1010-Pmin)2,其中T为热带气旋的持续时长,Pmin为最低气压。TCI运算结果是基于热带气旋发生时的最低气压以及该最低气压在东海海域的持续时长,TCI为一年中所有热带气旋数的累加值,可综合反映海域中热带气旋的发生频率、影响时长和热带气旋的强度等,其优于热带气旋的发生频率、持续时间和强度等单一指标。
1.3 表层水温资料
东海海表水温数据从KNMI Climate Explorer 2°×2℃OADS网格化的数据集中下载(海域范围为117°E~131°E,23°N~33°N)。下载的海表水温时间序列数据存在明显增强趋势,其主要是因为测量技术的改进引起[16],因此,在与东海马面鲀渔获量变动分析中海表水温的增强趋势将被移除。陆地降雨、季风、热带气旋影响指数和海表水温时间序列数据均经过Z-score标准化处理,K-S检验(Kolmogorov-Smirnov test)气候时间序列资料的正态性,其结果显著性水平均大于0.05。因此认为气候时间序列数据均符合正态性,可直接与渔获量数据进行相关性计算和回归性分析。
1.4 数据分析
东海马面鲀渔业产量时间序列,1990年前主要包括东海绿鳍马面鲀渔获量,1990年后既含绿鳍马面鲀渔获量又含黄鳍马面鲀渔获量[2, 17-18]。尽管东海马面鲀渔业产量包含绿鳍马面鲀和黄鳍马面鲀的渔获量,但由于它们都属同一属,其生态习性与生活习性及其对环境的要求又具有相似性,固可将它们作为单一的群系来做分析[15]。根据传统渔业理论,捕捞压力的单调增长会引起渔获量的变动呈曲线型变化趋势,渔获量的曲线型变化趋势可用SPSS统计分析软件对其进行Fox模型拟合[19]。Fox模型通常可表示为:
$$\;\;\;\; Y_e=\mathrm{C} f_e \mathrm{e}^{-\mathrm{d} f_e} $$ 式中Ye为平衡渔获量,fe为捕捞努力量,C、d为参数。
在消除捕捞压力引起的渔获量变化趋势后,其变动与陆地降雨、季风、海表水温和热带气旋影响指数等气候变量相关。根据气候变化时间序列数据建立渔获量变动与各气候变量的多元线性回归模型,回归模型中各气候变量的偏相关系数可反映其对渔获量变动的影响程度。由于气候变化不仅影响到现有的鱼类种群和饵料生物数量,而且还会影响鱼类的成活、补充、生长和繁育等,甚至通过亲体补充关系影响下一世代鱼类的发生数量,因此将对气候变量后5年内的马面鲀渔获量变动进行回归分析和检验。
渔获量的变动不仅与捕捞压力的单调增长有关,还与气候环境的变化有关。在捕捞压力和气候变化的双重影响和作用下,渔获量的评估模型可以用优化的Fox模型[10, 20]来表示:
$$ Y = {\rm{C}}f{{\rm{e}}^{ - {\rm{d}}f}} + \left( {\sum\limits_{i = 1}^n {{{\rm{a}}_i}} {X_i} + {{\rm{a}}_0}} \right) $$ 式中(${\sum\limits_{i = 1}^n {{{\rm{a}}_i}} {X_i} + {{\rm{a}}_0}} $)为气候因素对渔获量的影响部分,Y为根据捕捞努力量和各气候变量估算出的渔获量,f为捕捞努力量,Xi为第i个气候影响因子,ai为第i个气候影响因子的系数,C、d和a0为常数项。
2. 结果
2.1 渔获量与捕捞努力量关系
1976年~2006年的东海马面鲀渔获量和东海捕捞渔船功率变化时间序列变动十分显著(图 1)。1976年~1978年渔获量呈小幅波动,但到1979年渔获量急剧下降,仅5.4×104 t,之后的3年不断上升;1983年再次大幅减产,仅4.3×104 t,1984年回升,1985年基本持平,1986年继续增加,到1987年创历史最高水平(22.7×104 t),之后再次经历下降和上升的过程,1993年降到最低(仅3.2×104 t),1993年后年间波动幅度减小,除1997年和2005年渔获量分别达13.2×104 t和11.7×104 t外,基本维持在约4×104~7×104 t。
![]() 图 1 东海马面鲀渔获量和东海捕捞努力量时间序列Figure 1. Time series of catches and fishing efforts of filefish in East China Sea
图 1 东海马面鲀渔获量和东海捕捞努力量时间序列Figure 1. Time series of catches and fishing efforts of filefish in East China Sea根据传统渔业理论,东海马面鲀渔获量与捕捞努力量的关系可以用Fox模型来拟合,运用SPSS统计分析软件对其进行迭代计算,拟合出的Fox模型为:
$$ Y_i=0.188 f_i \mathrm{e}^{-4.636 \times 10^{-3} f_i} $$ 式中fi为第i年的东海区捕捞努力量,Yi为根据Fox模型估算的第i年东海马面鲀渔获量。Fox模型估算的东海马面鲀渔获量与实际渔获量显著相关(R=0.6,P < 0.01),说明东海马面鲀渔获量的变化与捕捞努力量的增长有关(图 1)。
2.2 渔获量变动与气候变化关系
捕捞压力增长引起东海马面鲀渔获量年际间的变化趋势可根据Fox模型来移除,移除变化趋势后的马面鲀渔获量变动见图 2。移除趋势后的东海马面鲀渔获量变动与降雨、季风和热带气旋等气候变化时间序列相关,可以用相应的气候变量组合来对其进行拟合。气候变量含东海热带气旋影响指数、东海海表水温、黄海冬季季风和夏季季风、东海冬季季风和夏季季风、长江流域和东海沿岸降雨(图 3)。
![]() 图 2 东海马面鲀渔获量的年间变动(残差)Figure 2. Inter-annual fluctuation of filefish catches (residual) in East China Sea
图 2 东海马面鲀渔获量的年间变动(残差)Figure 2. Inter-annual fluctuation of filefish catches (residual) in East China Sea用多元线性回归方法把东海马面鲀渔获量变动和气候变量时间序列联系起来,对相应气候变量后5年内的马面鲀渔获量变动进行了分析。东海马面鲀渔获量变动和各气候变量的多元线性回归模型为:
ΔYi=5.501Mi+1.348Ni-1-1.523Ni-2-3.439Ni-4-2.514Qi-1+3.98Pi-8.774Ri-5+14.125Si-2+0.397
式中△Yi为第i年的东海马面鲀渔获量变动估算值,Mi为第i年的东海海表温度,Ni为第i年的东海热带气旋影响指数,Qi为第i年的长江流域和东海沿岸降雨,Pi为第i年的黄海夏季季风,Ri为第i年的黄海冬季季风,Si为第i年的东海冬季季风。
东海马面鲀渔获量变动与气候变量的多元线性回归模型回归系数R=0.87(P < 0.01),且回归模型中所有的气候变量都具有显著性偏相关(P < 0.05),各气候变量间的共线性分析容差均大于0.5,说明该回归方程有效。回归结果表明,渔获量变动不仅与东海海表温度、热带气旋影响指数、东海冬季季风和黄海夏季季风呈显著正偏相关外,还与热带气旋影响指数、黄海冬季季风和长江流域和东海沿岸降雨呈显著负偏相关(表 1)。
表 1 渔获量变动与气候变量多元线性回归方程中的偏相关分析Table 1. Partial correlations between variations of filefish catches and climatic variables in the multivariate linear regression equation气候变量
climatic variable偏相关系数(时滞/年)
partial correlation coefficient (time lag/a)P 热带气旋影响指数 tropical cyclone impact index 0.45(1);-0.48(2);-0.68(4) 0.02 东海海表温度 sea surface temperature in East China Sea 0.47(5) 0.02 东海冬季季风 winter monsoon in East China Sea 0.79(2) 0.00 黄海夏季季风 summer monsoon in Yellow Sea 0.49(0) 0.01 黄海冬季季风 winter monsoon in Yellow Sea -0.78(5) 0 长江流域和东海沿岸降雨 annual precipitation in Yangtze River Valley and coastal region of East China Sea -0.63(1) 0 2.3 渔获量拟合结果
根据东海马面鲀渔获量与捕捞努力量以及气候变量之间的关系,对东海马面鲀渔获量进行了拟合。东海马面鲀渔获量变动拟合值Y′i=Yi+ΔYi,其中Yi为捕捞压力的增长引起的渔获量趋势变动,ΔYi为由气候变化影响下的渔获量年间变动。该方程拟合的渔获量与实际渔获量之间的回归系数R=0.91,且P < 0.01。可以看出,不仅拟合的马面鲀渔获量的年间波动与实际马面鲀渔获量的年间波动相吻合,而且其年际间的趋势变化也一致(图 4)。
![]() 图 4 东海马面鲀渔获量和根据捕捞努力量及气候变量拟合的渔获量Figure 4. Actual catches and catches fitted by fishing effort and climatic variables for filefish in East China Sea
图 4 东海马面鲀渔获量和根据捕捞努力量及气候变量拟合的渔获量Figure 4. Actual catches and catches fitted by fishing effort and climatic variables for filefish in East China Sea3. 分析与讨论
3.1 捕捞压力对渔获量的影响
东海马面鲀渔业的兴起到衰退仅20年时间,这主要与捕捞力量的过快增长有关,过度捕捞导致种群资源量急速衰退[4, 7]。1976年~1986年东海区渔船功率从114×104 kW增至205×104 kW,期间马面鲀渔获量呈剧烈波动状态。在高强度的捕捞下,1976年~1978年鱼山和温台渔场的绿鳍马面鲀渔获量迅速下降[21]。1986年后随着渔船功率的不断增长及高捕捞强度渔具渔法的广泛应用,东海马面鲀渔获量分别在1987年和1989年超20×104 t之后开始急剧下降。江外和舟外渔场的产卵洄游群体1986年~1987年的渔获量高达约17×104 t,1992年则降至1.6×104 t[22]。捕捞压力的增强还导致渔获物中高龄鱼数量减少,低龄鱼和补充群体数量显著增加[19]。1991年~1992年大批中层拖网渔船在沙外渔场大量捕捞绿鳍马面鲀幼鱼,导致1992年渔获量降至6.7×104 t,此后连年下跌,使东海绿鳍马面鲀资源补充机制受到了严重的破坏[23]。
在绿鳍马面鲀资源衰退的同时,东海黄鳍马面鲀资源开始兴起,东海区马面鲀渔汛转为以捕捞黄鳍马面鲀为主[7]。然而,由于捕捞压力的持续增长,黄鳍马面鲀的资源量也逐渐下降,1996年黄鳍马面鲀的资源尾数和资源质量分别为41.2×108 尾和10.1×104 t, 1997年后开始下降,至2000年其资源尾数和资源质量分别为15.4×108 尾和4.5×104 t[22]。从黄鳍马面鲀群体组成变化来看,小型化和低龄化明显,20世纪90年代初渔获群体以2~3龄为主,90年代后期则以1龄鱼为主体[24]。从东海黄鳍马面鲀开发利用以来,其渔获量从最高年份的(6~7)×104 t降至2006年的3×104 t,说明黄鳍马面鲀的资源衰退明显,究其原因可能与过度捕捞有关。黄鳍马面鲀资源基础远比绿鳍马面鲀差,但投入的捕捞强度更大,因此造成黄鳍马面鲀资源出现衰退[22]。
3.2 渔获量变动对气候变化的响应
东海马面鲀渔获量的年间变动十分显著,这种变动不仅与捕捞压力的增长有关,还与相应的气候环境变动相关。气候变化对渔业的影响主要是通过影响海洋物理和生物环境条件来实现。
3.2.1 长江流域和东海沿岸降雨
尽管沿海陆地降雨和径流能携带大量氮(N)、磷(P)等营养盐及其他无机物质进入海洋,而丰富的营养盐是河口和邻近海域的浮游植物营养补充的主要来源[25],但长江流域和东海沿岸降雨与东海马面鲀渔获量变动呈显著负偏相关(P < 0.01),说明该地域降雨对东海马面鲀渔业产量有着显著负面影响。这可能是因为降雨和径流所形成的冲淡水引起水体层化现象,限制了营养盐的垂直流动和初级生产力的提高[26]。另外冲淡水所带来的海水盐度的变化也会影响到鱼类的集群和分布,马面鲀喜欢栖息在高盐度的水中,其栖息的海域盐度一般为33.5~35,产卵期其适盐范围为34~35[27],因此沿岸冲淡水带来的盐度降低不利于马面鲀的繁育和生长。
3.2.2 东海和黄海季风
东、黄海沿岸具有典型的季风特性,冬季盛行偏北风,夏季盛行偏南风。在黄海,长年存在一支黄海沿岸流,其为低盐、低温的水流,水色混浊[28]。冬季,在强的东北季风作用下,南下的黄海沿岸流势力增强[29],因黄海沿岸流具有低盐、低温的特点,其势力增强将影响马面鲀种群的活动范围及生存空间[27],因此,黄海冬季季风的增强不利于马面鲀渔业产量的增加,这与黄海冬季季风与马面鲀渔获量呈负偏相关相一致(P < 0.01);夏季,在西南季风的作用下,南下的黄海沿岸流流速减弱[29],而减弱的黄海沿岸流对马面鲀种群的索饵及其发展较为有利[27],因此,黄海夏季季风的增强有利于马面鲀渔业产量的增加,这与黄海夏季季风与东海马面鲀渔获量呈显著正偏相关相吻合(P<0.02)。东海冬季季风与东海马面鲀渔获量变动呈显著正偏相关(P < 0.01),说明东海冬季季风对东海马面鲀渔获量有显著影响,这种影响可能与冬季季风带来的降温引起马面鲀的集群有关。在强的冬季季风作用下,沿岸冷水团发展强势,流隔也明显加强,导致马面鲀出现贴底集群,形成渔汛[30]。
3.2.3 东海海表水温
海水温度的变化成为影响海洋生物变化的重要水文因素,水温的变化对海洋鱼类的分布、洄游、生长以及其繁育等有重要影响[31]。东海海表水温与东海马面鲀渔获量变动呈显著正偏相关(P < 0.03),说明水温的变化对马面鲀渔业有显著影响。从1971年中国钓鱼岛北部渔场调查的水文来看,表层水温从21.6 ℃升到24.5 ℃、底层水温从16.7 ℃升至19.0 ℃时绿鳍马面鲀的性腺发育[32]。因此,渔场水温的升高会促进马面鲀产卵群体的性腺发育,增加马面鲀产卵群体的数量,而马面鲀亲体数量的增多和产卵时间的提前有利于马面鲀补充群体数量的增加。另外,渔场水温的提高能促进浮游植物的光合作用和繁殖生长,对提高海域的生产力、增加马面鲀饵料生物的数量有帮助[31]。因此,海水温度的变化对东海马面鲀的繁育和生长以及补充群体数量的增加均有重要的影响。
3.2.4 热带气旋
热带气旋是发生在热带或副热带洋面上的低压涡旋,东海热带气旋均来自于西太平洋和南海海域[12]。热带气旋带来的强风和降雨等改变了海洋生物的生存环境,不可避免地对其产生影响。从热带气旋影响指数与东海马面鲀渔获量变动的相关分析中可以看出,其与马面鲀渔获量变动在不同的时滞段分别呈显著正偏相关和负偏相关(P < 0.03),说明热带气旋对东海马面鲀渔获量的影响具有双重效应。热带气旋对东海马面鲀渔获量变动影响的正面效应可能与热带气旋的发生增加了海洋生态系统营养盐的供应有关。BABIN等[33]通过海洋遥感技术对1998年~2001年北大西洋地区的13个热带气旋研究时发现,热带气旋经过后海洋表面叶绿素浓度增高,增高的叶绿素水平能维持约2~3周,认为热带气旋之所以能增加海洋表面叶绿素浓度,主要是因为其所带来的水层强烈混合作用以及其所诱导的涌升流等因素增加了海洋上层营养盐的供应,提高了海域的初级生产力。SHIAH等[34]在研究热带气旋对台湾西北大陆架海域影响时发现,热带气旋经过后所引起的风生混合、再悬浮以及陆地径流等明显增加了海域的硝酸盐浓度,初级生产力也得到很大提高,使该海域变为一个高生产力的区域。热带气旋的负面效应可能是其经过后改变了水域中马面鲀的生存环境,从而影响了马面鲀的集群和洄游等。热带气旋所带来的强风和降雨对其他海洋生物带来的负面影响已有报道[35-38]。马面鲀种群的群居特性明显,具有特定的栖息场所,并且一年中有规则地进行索饵、产卵和越冬洄游等[27]。热带气旋带来的强降雨和海水混合等可能对马面鲀种群的分布、繁育和摄食等活动带来一定的负面影响。
3.3 渔获量的变化趋势分析
自20世纪80年代开发以来,东海马面鲀渔获量一直处于强烈的波动之中。根据1976年~2006年的捕捞努力量以及相应的气候环境变动时间序列数据对东海马面鲀渔获量进行了拟合,拟合结果显示,拟合的渔获量与实际渔获量显著相关(R=0.91,P < 0.01),并且对马面鲀渔获量的年间波动也能做出较好的拟合,说明东海马面鲀渔获量变动不仅与捕捞压力的增长有关,还与气候环境的变化有关。气候环境的变化不仅影响到海洋中营养盐分布和初级生产力的变化,还影响东海马面鲀种群的分布、洄游、摄食以及繁育等。未来全球气候温暖化会造成气候环境的变动幅度加剧,联合国政府间气候变化专门委员会(IPCC)对全球过去50年中发生的极端天气事件展开了调查,结果显示极端天气事件有增多增强的趋势,并预计未来这种增多增强的趋势还会持续增加[39]。因此推测,受极端天气事件的影响,未来东海马面鲀的渔业产量还会出现更大范围的波动。
-
![]()
图 1 聚β-羟基丁酸酯对凡纳滨对虾肝胰腺总抗氧化能力和溶菌酶活性的影响
不同小写字母表示组间差异显著(P < 0.05), 相同小写字母表示组间差异不显著(P>0.05);后图同此
Figure 1. T-AOC and LSZ activity in hepatopancreas of L.vannamei after PHB treatment
Different letters indicate significant difference (P < 0.05), while the same letters indicate no significant difference (P>0.05).The same case in the following figures.
![]()
图 2 聚β-羟基丁酸酯对凡纳滨对虾肝胰腺HSP70基因表达量和Toll基因表达量的影响
Figure 2. Relative expression level of HSP70 and Toll gene in hepatopancreas of L.vannamei after PHB treatment
![]()
图 3 聚β-羟基丁酸酯对凡纳滨对虾肝胰腺淀粉酶、脂肪酶、胃蛋白酶和胰蛋白酶活性的影响
Figure 3. Amylase, lipase, pepsin and trypsin activities in hepatopancreas of L.vannamei after PHB treatment
表 1 该研究所用引物序列
Table 1 Sequence of primers in this study
引物名称
primer name引物序列(5′→3′)
primer sequence登录号
accession No.HSP70-F AGGAGACCGCTGAGGCTTAC AY645906 HSP70-R AGCACATTCAGACCCGAGAT Toll-F CCAGCTTAGAAGACCGGCAA DQ923424 Toll-R GTTGTCCGAGCAGAAGTCCA β-actin-F GCCCTGTTCCAGCCCTCATT AF300705 β-actin-R ACGGATGTCCACGTCGCACT  下载: 导出CSV
下载: 导出CSV
-
[1] 李玉虎, 宋芹芹, 张志怀, 等.凡纳滨对虾生长发育规律及生长曲线拟合研究[J].南方水产科学, 2015, 11(1):89-95. http://www.schinafish.cn/CN/abstract/abstract9184.shtml [2] 向赟, 吴宇学, 王刚, 等.不同温度下哈维氏弧菌和白斑综合症病毒对凡纳滨对虾的致病性[J].大连海洋大学学报, 2015, 30(1):56-61. http://edu.wanfangdata.com.cn/Periodical/Detail/hnrdnydxxb201601001 [3] 李晶晶, 李云, 刘红, 等.感染溶藻弧菌及白斑综合症病毒后凡纳滨对虾不同组织的Toll样受体基因表达变化研究[J].海洋与湖沼, 2016, 47(2):476-483. http://www.cqvip.com/QK/90072X/201602 [4] 罗词兴, 黄旭雄, 李桑, 等.溶藻弧菌感染后凡纳滨对虾鳃组织免疫相关基因的表达[J].中国水产科学, 2014, 21(1):189-196. http://www.cqvip.com/QK/90072X/201602/669133827.html [5] 李健, 梁俊平, 李小彦, 等.黄芩苷在中国对虾体内对诺氟沙星消除及细胞色素P450酶的影响[J].海洋科学, 2012, 36(3):81-88. http://www.oalib.com/references/17806877 [6] 文国樑, 林黑着, 李卓佳, 等.饲料中添加复方中草药对凡纳滨对虾生长、消化酶和免疫相关酶活性的影响[J].南方水产科学, 2012, 8(2):58-63. http://www.schinafish.cn/CN/abstract/abstract8940.shtml [7] 杨铿, 林黑着, 夏冬梅, 等.饲料中海洋红酵母对尼罗罗非鱼幼鱼生长性能、消化酶及免疫酶活性的影响[J].南方水产科学, 2016, 12(6):51-58. http://www.schinafish.cn/CN/abstract/abstract9401.shtml [8] BARUAH S K, NOROUZITALLAB P, DEBNATH D, et al.Organic acids as non-antibiotic nutraceuticals in fish and prawn feed[J].Aquacult Health Int, 2008(12):4-6. https://www.researchgate.net/publication/293336603_Organic_acids...
[9] 孙慧贤, 隋丽英.聚β-羟基丁酸酯对卤虫幼体的强化作用[J].上海海洋大学学报, 2011, 20(3):392-398. http://www.cnki.com.cn/Article/CJFDTOTAL-SCKX201408013.htm [10] 魏茂繁, 邵云海, 钟卫鸿.生物合成聚-β-羟基丁酸(PHB)的研究现状与应用前景[J].黑龙江科学, 2014, 5(6):24-25. http://www.cnki.com.cn/Article/CJFDTOTAL-GSKJ200603049.htm [11] 刘春, 张小凡.PHB在生产可降解塑料方面的应用及其微生物累积的研究进展[J].塑料工业, 2005, 33(8):1-3, 18. http://www.cnki.com.cn/Article/CJFDTotal-XJJB201432022.htm [12] 邓元告, 黄琼叶, 马灌楠, 等.PHB剂量和饲喂时间对中华绒螯蟹肝胰腺酶活力和肠道菌群多样性的影响[J].中国水产科学, 2016, 23(1):138-145. http://d.wanfangdata.com.cn/Periodical_zgsckx201601013.aspx [13] De SCHRYVER P, SINHA A, KUNWAR P, et al.Poly-β-hydroxybutyrate (PHB) increases growth performance and intestinal bacterial range-weighted richness in juvenile European sea bass, Dicentrarchus labrax[J].Appl Microbiol Biotechnol, 2010, 86(5):1535-1541. doi: 10.1007/s00253-009-2414-9
[14] 曹俊明, 严晶, 王国霞, 等.家蝇蛆粉替代鱼粉对凡纳滨对虾消化酶、转氨酶活性和肝胰腺组织结构的影响[J].南方水产科学, 2012, 8(5):72-79. http://www.schinafish.cn/CN/abstract/abstract8983.shtml [15] 邓康裕, 孔杰, 孟宪红, 等.聚β-羟基丁酸酯对中国明对虾抗WSSV能力的影响[J].水产学报, 2014, 38(8):1167-1174. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [16] SUGUNA P, BINURAMESH C, ABIRAMI P, et al.Immunostimulation by poly-β hydroxybutyrate-hydroxyvalerate (PHB-HV) from Bacillus thuringiensis in Oreochromis mossambicus[J].Fish Shellfish Immunol, 2014, 36(1):90-97. doi: 10.1016/j.fsi.2013.10.012
[17] NHAN D T, WILLE M, de SCHRYVER P, et al.The effect of poly-β-hydroxybuty rate on larviculture of the giant fresh water prawn Macrobrachium rosenbergii[J].Aquaculture, 2010, 302(1/2):76-81. https://www.sciencedirect.com/science/article/pii/S105046481630715X
[18] 刘玉, 隋丽英, 邓元告, 等.聚β-羟基丁酸酯(PHB)对中华绒螯蟹(Eriocheir sinensis)幼蟹生长和肝胰腺酶活力的影响[J].海洋与湖沼, 2013, 44(5):1333-1338. http://www.cnki.com.cn/Article/CJFDTOTAL-HYFZ201305031.htm [19] WU F, XUE Y, LIU X, et al.The protective effect of eicosapentaenoic acid-enriched phospholipids from sea cucumber Cucumaria frondosa on oxidative stress in PC12 cells and SAMP8 mice[J].Neurochem Int, 2014, 64:9-17. doi: 10.1016/j.neuint.2013.10.015
[20] 张克烽, 张子平, 陈芸, 等.动物抗氧化系统中主要抗氧化酶基因的研究进展[J].动物学杂志, 2007, 42(2):153-160. http://www.wenkuxiazai.com/doc/eec3817c1711cc7931b716df-4.html [21] 段亚飞, 董宏标, 王芸, 等.干露胁迫对日本囊对虾抗氧化酶活性的影响[J].南方水产科学, 2015, 11(4):102-108. http://www.schinafish.cn/CN/abstract/abstract9241.shtml [22] 段亚飞, 董宏标, 王芸, 等.鳗弧菌感染对斑节对虾免疫相关指标的影响[J].海洋科学, 2015, 39(9):44-50. doi: 10.11759//hykx20150409001 [23] 邓康裕, 孔杰, 孟宪红, 等.聚β-羟基丁酸酯对凡纳滨对虾抗WSSV能力及免疫基因表达量的影响[J].中国水产科学, 2015, 22(5):877-886. http://www.cqvip.com/QK/98270X/201505/666134560.html [24] SUI L Y, MA G N, LU W L, et al.Effect of poly-β-hydroxybutyrate on growth, enzyme activity and intestinal microbial community of Chinese mitten crab, Eriocheir sinensis (Milne-Edwards) juveniles[J].Aquacult Res, 2016, 47(11):3644-3652. doi: 10.1111/are.2016.47.issue-11
[25] 雷爱莹, 曾地刚.复方中草药对凡纳滨对虾热应激蛋白70基因表达的影响[J].广西农业科学, 2008, 39(6):830-833. http://www.cqvip.com/QK/83434X/200804/29012150.html [26] 曹煜成, 文国樑, 张华军, 等.地衣芽孢杆菌对凡纳滨对虾Toll和HSP70基因表达的影响[J].中国微生态学杂志, 2013, 25(8):882-886. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGWS201308003.htm [27] THAI T, WILLE M, GARCIA-GONZALEZ L, et al.Poly-β-hydroxybutyrate content and dose of the bacterial carrier for Artemia enrichment determine the performance of giant freshwater prawn larvae[J].Appl Microbiol Biotechnol, 2014, 98(11):5205-5215. doi: 10.1007/s00253-014-5536-7
[28] 邓康裕, 孟宪红, 孔杰, 等.聚β-羟基丁酸酯对凡纳滨对虾(Litopenaeus vannamei)非特异性免疫力及氨氮胁迫后免疫基因表达量的影响[J].渔业科学进展, 2015, 36(6):71-78. doi: 10.11758/yykxjz.20150611 [29] MORTENSEN F, NIELSEN H, MULVANY M, et al.Short chain fatty acids dilate isolated human colonic resistance arteries[J].Gut, 1990, 31(12):1391-1394. doi: 10.1136/gut.31.12.1391
[30] PARTANEN K, MROZ Z.Organic acids for performance enhancement in pig diets[J].Nutr Res Rev, 1999, 12(1):117-145. doi: 10.1079/095442299108728884
[31] NAJDEGERAMI E, TRAN T, DEFOIRDT T, et al.Effects of poly-β-hydroxybutyrate (PHB) on Siberian sturgeon (Acipenser baerii) fingerlings performance and its gastrointestinal tract microbial community[J].FEMS Microbiol Ecol, 2012, 79(1):25-33. doi: 10.1111/j.1574-6941.2011.01194.x
[32] NAJDEGERAMI E H, BARUAH K, SHIRI A A, et al.Siberian sturgeon (Acipenser baerii) larvae fed Artemia nauplii enriched with poly-β-hydroxybutyrate (PHB):effect on growth performance, body composition, digestive enzymes, gut microbial community, gut histology and stress tests[J].Aquacult Res, 2015, 46(4):801-812. doi: 10.1111/are.12231
[33] WELTZIEN F A, HEMRE G I, EVJEMO J O, et al.β-hydroxybutyrate in developing nauplii of brine shrimp (Artemia franciscana K.) under feeding and non-feeding conditions[J].Comp Biochem Physiol B, 2000, 125(1):63-69. doi: 10.1016/S0305-0491(99)00159-5
-
期刊类型引用(16)
1. 高天翔,张浩博,王晓艳,陈治. 基于eDNA技术的舟山近海中国团扇鳐定性与定量分析. 水产学报. 2024(03): 118-127 .  百度学术
百度学术
2. 夏明霞,张辉,线薇薇. 气候变化对中国近海中上层小型鱼类影响的研究进展. 海洋科学. 2024(06): 78-92 . 百度学术
3. 徐姗楠,郭建忠,陈作志,张魁,许友伟,李纯厚. 胶州湾鱼类生物量粒径谱特征. 水产学报. 2020(04): 596-605 . 百度学术
4. 徐姗楠,郭建忠,范江涛,许友伟,李纯厚. 夏季大亚湾鱼类群落结构与多样性. 生态学杂志. 2020(04): 1254-1264 . 百度学术
5. 徐姗楠,郭建忠,范江涛,许友伟,粟丽,李纯厚. 大亚湾夏季鱼类生物量粒径谱年际变化特征. 南方水产科学. 2020(04): 28-38 . 本站查看
6. 徐姗楠,郭建忠,陈作志,许友伟,周林滨,柯志新,李纯厚. 大亚湾鱼类生物量粒径谱特征. 中国水产科学. 2019(01): 34-43 . 百度学术
7. 徐姗楠,郭建忠,陈作志,张魁,许友伟,蔡研聪,李纯厚. 胶州湾鱼类资源量的时空分布特征. 水产学报. 2019(07): 1615-1625 . 百度学术
8. 祖凯伟,程家骅,刘阳,张弛,李建超,潘新冬,叶振江,田永军. 产卵期及越冬期蓝点马鲛渔场分布变化及其与海表温度的关系. 海洋湖沼通报. 2019(06): 48-57 . 百度学术
9. 郭建忠,陈作志,许友伟,徐姗楠,李纯厚. 大亚湾鱼类资源数量的时空分布特征. 中国海洋大学学报(自然科学版). 2018(12): 47-55 . 百度学术
10. 郭建忠,陈作志,徐姗楠. 鱼类粒径谱研究进展. 海洋渔业. 2017(05): 582-591 . 百度学术
11. 刘笑笑,王晶,徐宾铎,薛莹,任一平. 捕捞压力和气候变化对黄渤海小黄鱼渔获量的影响. 中国海洋大学学报(自然科学版). 2017(08): 58-64 . 百度学术
12. 郭禹,陈国宝,于杰,李斌,王东旭. 南海北部马面鲀渔获率初步研究. 水产科学. 2016(02): 123-129 . 百度学术
13. 张志敏,徐年军. 东海渔获量受捕捞努力与气候因子影响的年际变动分析. 宁波大学学报(理工版). 2016(04): 112-116 . 百度学术
14. 刘尊雷,袁兴伟,杨林林,严利平,田永军,程家骅. 气候变化对东海北部外海越冬场渔业群落格局的影响. 应用生态学报. 2015(03): 901-911 . 百度学术
15. 刘春胜,庄志猛,陈四清,刘长琳,赵鹏,陈昭廷. 绿鳍马面鲀(Navodon septentrionalis)对4种大型水母的捕食行为. 渔业科学进展. 2014(06): 30-38 . 百度学术
16. 李玉芳,李永振,陈国宝,孙典荣,于杰,崔科. 基于线粒体Cyt b基因的黄鳍马面鲀种群分析. 水产学报. 2014(01): 23-32 . 百度学术
其他类型引用(10)




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 26
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
