
Cloning and expression analysis of Chitinase-2 from Penaeus monodon during molting cycle and different larval developmental stages
-
摘要:
研究以斑节对虾(Penaeus monodon)转录组获得的几丁质酶基因(PmChi)片段,运用RACE技术克隆了PmChi基因cDNA全长,命名为PmChi-2。PmChi-2基因cDNA全长2 050 bp,其中5′-非编码区(5′-UTR)144 bp、3′-UTR 319 bp和开放阅读框(ORF)1 587 bp,编码528个氨基酸。生物信息学分析显示,PmChi-2与其他甲壳动物Chi-2的相似性为78%~97%。实时荧光定量PCR结果显示,PmChi-2在蜕皮前期(D期)表皮中表达水平最高,在胃、鳃、腹神经节和眼柄中表达水平依次降低,在其他组织(肝胰腺、肠、肌肉、心脏)中几乎不表达。不同蜕皮阶段,PmChi-2在鳃、胃和表皮3种组织中的表达变化模式基本一致,均在蜕皮期(E期)表达量最低,而最高表达量在鳃中出现在蜕皮后期(A期),在胃和表皮中为D期。幼体不同发育阶段分析揭示PmChi-2 mRNA在幼体发育阶段的无节幼体到糠虾幼体第二期表达量维持在一个低水平,在糠虾幼体第三期PmChi-2 mRNA表达水平显著升高,在仔虾期又显著下降,推测PmChi-2 mRNA可能与斑节对虾幼体发育密切相关。研究结果表明PmChi-2基因可能在斑节对虾蜕皮以及幼体的变态发育中发挥重要作用,为深入研究斑节对虾几丁质酶发育调控提供了重要信息。
Abstract:We obtained the full-length cDNA sequence of PmChi (GenBank accession number:KX610684, namely PmChi-2) from black tiger shrimps (Penaeus monodon) by transcriptome sequencing and rapid amplification of cDNA ends (RACE). The full length of PmChi-2 cDNA was 2 050 bp, including a 1 587 bp ORF (open reading frame) which encoded 528 amino acid residues, a 144 bp 5′-UTR and a 319 bp 3′-UTR. Phylogenetic analyses suggest that the predicted amino acid sequence of PmChi-2 shared 78%~97% identity with the type of Chi-2 of other crustaceans. Relative gene expression levels of PmChi-2 gene in various tissues were then detected by qRT-PCR (quantitative real-time PCR) during the molting cycle and different larval developmental stages. The results show that PmChi-2 had the highest gene expression level in cuticle in the premolt, then decreased in a descending order (stomach > gill > thoracic ganglia > eyestalk), while weak expression was detected in the other tissues. The gene expression patterns of PmChi-2 were different among tissues during the molting cycle. PmChi-2 mRNA had the highest expression level at Stage A and the lowest expression level at Stage E in gill. PmChi-2 mRNA had the highest expression level at Stage D and the lowest expression level at Stage E in stomach. PmChi-2 mRNA had the highest expression level at Stage D and the lowest expression level at Stage E in cuticle, which suggests that PmChi-2 may participate in the molting. At different larval developmental stages, PmChi-2 mRNA had the highest expression level at Mysis 3, indicating that PmChi-2 was associated with larval metamorphosis. The results lay a foundation for research on the biological function and regulation of chitinase in P.monodon.
-
Keywords:
- Penaeus monodon /
- chitinase /
- gene cloning /
- expression analysis /
- molting /
- metamorphosis
-
斑节对虾(Penaeus monodon)是对虾主要养殖品种之一[1]。目前主要通过传统育种的手段选育生长性状优良的品种,选育的时间长、效率低。因此,了解斑节对虾生长发育方面的分子调控机理,对于开展生长性状方面的分子辅助育种有非常重要的意义。斑节对虾的生长发育需要经历多次蜕皮,而其外壳主要成分是几丁质,因此开展几丁质酶的功能研究,对于阐明其生长发育过程中的蜕皮机理十分有必要。
几丁质是由N-乙酰-β-D-氨基葡萄糖以β-1, 4-糖苷键连接形成的长而无支链的高分子多聚物[2]。它是自然界中许多生物的结构性组分,如是真菌细胞壁的重要组成成分,也是节肢动物特别是甲壳动物外壳和围食膜的重要组成成分[3-5]。几丁质酶包含4个典型的结构域:信号肽、几丁质结合结构域(CBD)、第18家族几丁质酶催化结构域(GH18催化结构域)和富含Ser/Thr的连接区(或富含Thr/Pro的连接区)。甲壳动物几丁质酶不仅参与蜕皮调控和围食膜降解,还参与机体内多个生物学过程,如消化食物中的几丁质和抵抗外来病原体入侵等[6-9]。甲壳动物外壳中的几丁质需要经过几丁质酶和壳二糖(N-乙酰-β-D-氨基葡萄糖甘酶)的共同作用才能完全分解:几丁质先被几丁质酶降解为寡聚糖,接着寡聚糖被壳二糖酶继续水解为N-乙酰-β-D-氨基葡萄糖单体,这些单体可被机体再次吸收,用于甲壳动物新壳的形成[2]。与昆虫相比,甲壳动物几丁质酶研究开展较晚,但由于其对甲壳动物生长发育很重要,也逐渐成为研究热点。目前,在中华绒螯蟹(Eriocheir sinensis)[10-11]、斑节对虾[2, 12]、日本对虾(P.japonicus)[13-15]、凡纳滨对虾(Litopenaeus vannamei)[16-17]、中国对虾(Fenneropenaeus chinensis)[18]、日本仿长额虾(Pandalopsis japonica)[19]、脊尾白虾(Exopalaemon carinicauda)[20]、日本沼虾(Macrobrachium nipponense)[21]和三疣梭子蟹(Portunus trituberculatus)[22-23]中均已开展了几丁质酶基因的克隆及功能研究。甲壳动物中已发现了Chi-1~Chi-6 6种几丁质酶基因,其划分主要依据是CBD和催化结构域的组成和顺序,其中报道Chi-2与甲壳动物的蜕皮密切相关。目前对虾中仅日本对虾[13]克隆了Chi-2的全长cDNA序列,凡纳滨对虾[16]和斑节对虾[2]仅克隆了部分序列。因此,该研究利用RACE技术以斑节对虾转录组中筛选的斑节对虾几丁质酶基因(Chitinase-2)片段获得了PmChi cDNA全长,并分析了该基因在斑节对虾蜕皮周期以及幼体不同发育阶段中的表达模式,以为斑节对虾生长以及幼体变态发育过程中的蜕皮分子调控机制提供基础数据。
1. 材料与方法
1.1 实验材料
实验材料取自中国水产科学研究院南海水产研究所深圳试验基地的健康斑节对虾,体质量为19~22 g,体长为11~13 cm。斑节对虾先暂养在盐度32,水体溶氧充足,水温控制在(25±1)℃的虾池中(虾池长、宽、高分别为2.5 m、2.5 m、1.5 m,养殖的水体体积为4.3 m3),每池80尾,共3池,每天换水量为1/3,早上7: 00和下午6: 00投喂饲料,中午11: 30投喂鱿鱼,次日早上清除残饵和粪便。暂养3 d后开始实验,管理同暂养期间。用解剖剪剪取斑节对虾尾扇内肢置于载玻片上,Motic BA400光学显微镜观察(同一视野分别×40和×100)并拍照。根据尾扇内肢刚毛发生将其蜕皮周期分为蜕皮前期(D期)、蜕皮期(E期)、蜕皮后期(A期)和蜕皮间期(C期)[24] (图 1),取蜕皮各期的眼柄、鳃、肝胰腺、肠、胃、肌肉、心脏、胸神经节、表皮等组织样品,保存在RNAlater(Invitrogen, 美国)中备用。每个蜕皮阶段采集5~8尾虾。
![]() 图 1 斑节对虾蜕皮过程的A期、C期和D期A.A期尾扇内肢;C. C期尾扇内肢;D. D期尾扇内肢(×40);A′. A期尾扇内肢;C′. C期尾扇内肢;D′. D期尾扇内肢(×100);S.刚毛干;Sn.刚毛瘤;Sc.刚毛锥体;Nc.新表皮;Ss.新刚毛干;Pp.只看到一点光线的区域Figure 1. P.monodon molting process at Stage A, C and DA.postmoult(A) inner uropod; C. intermoult (C) inner uropod; D.premoult (D) inner uropod (×40);A′.postmoult (A) inner uropod; C′.intermoult (C) inner uropod; D′.premoult (D) inner uropod (×100);S. setal shaft; Sn. setal node; Sc. setal cone; Nc. new cuticle; Ss. new setal shaft; Pp. pinpoints of light where new setal node will develop
图 1 斑节对虾蜕皮过程的A期、C期和D期A.A期尾扇内肢;C. C期尾扇内肢;D. D期尾扇内肢(×40);A′. A期尾扇内肢;C′. C期尾扇内肢;D′. D期尾扇内肢(×100);S.刚毛干;Sn.刚毛瘤;Sc.刚毛锥体;Nc.新表皮;Ss.新刚毛干;Pp.只看到一点光线的区域Figure 1. P.monodon molting process at Stage A, C and DA.postmoult(A) inner uropod; C. intermoult (C) inner uropod; D.premoult (D) inner uropod (×40);A′.postmoult (A) inner uropod; C′.intermoult (C) inner uropod; D′.premoult (D) inner uropod (×100);S. setal shaft; Sn. setal node; Sc. setal cone; Nc. new cuticle; Ss. new setal shaft; Pp. pinpoints of light where new setal node will develop幼体不同发育期样品的采集:自受精卵孵化后,每天每隔0.5 h不间断的观察并记录发育时期,对达到一定发育时期的幼体进行样品的采集,采集的幼体发育期包括无节幼体(N),溞状幼体第一期(Z1)、第二期(Z2)、第三期(Z3), 糠虾幼体第一期(M1)、第二期(M2)、第三期(M3), 仔虾第一期(P1)、第二期(P2)、第六期(P6)、第十二期(P12)。样品用RNAlater(Invitrogen, 美国)保存。
1.2 实验方法
1.2.1 总RNA提取和cDNA合成
根据Trizol(Invitrogen,美国)说明书提取蜕皮各期组织以及幼体发育各阶段样品的总RNA,经琼脂糖凝胶电泳检测RNA完整性,NANODROP 2000检测RNA纯度[25]。使用SMARTer ® RACE 5′/3′ Kit (Clontech,日本)进行cDNA第一链的合成。
1.2.2 PmChi-2的克隆与测序
根据斑节对虾转录组获得的PmChi基因片段运用Primer premier 5.0设计基因特异性引物,利用RACE技术扩增目的基因的5′-非编码区(UTR)和3′-UTR。5′RACE和3′RACE中用到的引物为PmChi-2-5′-GSP1(5′-AATCGGCTTGGGGGAAGTGGTCGGT-3′)、PmChi-2-5′-GSP2(5′-CGTGTTAGGCATCGGGGTGCGGGA-3′)、PmChi-2-3′-GSP1(5′-GCTGCTTTTGCCGAATGGAT -3′)、PmChi-2-3′-GSP2(5′-CCGACAACGGACTCA-ACCAGA-3′)、UPM(long 5′-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3′,0.4 μmol·L-1;short 5′-CTAATACGACTCACTATAGGGC-3′,2 μmol·L-1)、NUP (5′-AAGCAGTGGTATCAACGCAGAGT-3′),RACE反应程序参考王艳等[26],琼脂糖凝胶电泳、目的条带的回收、连接、转化、挑阳性克隆、测序等参考赵超等[27]。
1.2.3 序列分析
使用DNAman软件拼接测序片段;使用BlastP (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)分析相似性;使用ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/)查找开放阅读框;使用Bioedit软件进行多序列比对;使用SMART 4.0(http://smart.embl-heidelberg.de/)分析蛋白结构域;用SingalP 4.1 (http://www.cbs.dtu.dk/services/SignalP/)程序预测信号肽;构建系统进化树采用Clustal X和MEGA 6.06软件。
1.2.4 实时荧光定量PCR(real-time quantitative PCR)
采用Trizol(Invitrogen, 美国)法提取斑节对虾蜕皮各期组织样品及幼体不同发育阶段的总RNA,按照SYBR Premix Ex TaqTM Kit (TaKaRa,日本)说明书逆转录为cDNA。根据Beacon Designer 7设计荧光定量引物PmChi-2-qPCR-F/R。内参基因的选择及引物的设计参考王艳等[26]。取斑节对虾D期的眼柄、鳃、肝胰腺、肠、胃、肌肉、心脏、胸神经节、表皮,按照荧光定量PCR试剂盒(TaKaRa,Cat.DRR420A)说明书进行定量PCR,研究PmChi-2基因在不同组织中的相对表达,每个样品设3个重复。根据D期组织表达结果,继续研究PmChi-2在蜕皮各个阶段鳃、胃、表皮中的表达变化。斑节对虾幼体不同发育期PmChi-2相对表达情况也采取同样的方法进行实验。
1.2.5 数据分析
通过2-ΔΔCt法计算PmChi-2在蜕皮各期组织以及幼体发育各阶段相对表达量。使用统计分析软件SPSS 23.0对实验结果进行单因素方差(One-Way ANOVA)分析,分析结果以“平均值±标准误(X±SE)”表示,P < 0.05表示差异显著。
2. 结果
2.1 PmChi-2生物信息学分析
斑节对虾几丁质酶基因(PmChi-2)cDNA全长2 050 bp,NCBI登录号为KX610684。包括144 bp 5′-UTR、319 bp 3′-UTR和1 587 bp开放阅读框,编码528个氨基酸,分子量为59.04 kD,等电点为7.86。PmChi-2包含甲壳动物几丁质酶4个典型结构域中的2个:CBD(41~93 aa)和GH18催化结构域(151~495 aa) (图 2)。
![]() 图 2 PmChi-2氨基酸序列及cDNA序列展示双下划线表示几丁质结合结构域;灰色表示GH18催化结构域;粗体表示起始密码子(ATG)和终止密码子(TAA);斜体表示PolyA结构。Figure 2. Predicted amino acid and nucleotide sequence of PmChi-2 cDNADouble-underlines indicate chitin binding domain(CBD); GH18 domain is shown in gray; the initiation codon (ATG) and termination codon (TAA) are highlighted in bold; the poly A signal sequence is italicized.
图 2 PmChi-2氨基酸序列及cDNA序列展示双下划线表示几丁质结合结构域;灰色表示GH18催化结构域;粗体表示起始密码子(ATG)和终止密码子(TAA);斜体表示PolyA结构。Figure 2. Predicted amino acid and nucleotide sequence of PmChi-2 cDNADouble-underlines indicate chitin binding domain(CBD); GH18 domain is shown in gray; the initiation codon (ATG) and termination codon (TAA) are highlighted in bold; the poly A signal sequence is italicized.多重序列比对发现,PmChi-2与日本对虾PjChi-2、凡纳滨对虾LvChi-2氨基酸序列相似度最高。与日本对虾PjChi-2一样,PmChi-2 CBD中也含有6个半胱氨酸和芳香族氨基酸残基。甲壳动物同种几丁质酶氨基酸序列相似度也较高,相似度在78%~97%,Chi-1、Chi-2、Chi-3的GH18结构域中的催化活性位点(FDGLDLDWEYP)高度保守(图 3)。
![]() 图 3 PmChi-2与其他甲壳动物的Chi的氨基酸多重序列比对方框内为结构域和催化活性位点;“●”和“★”分别表示半胱氨酸和芳香族氨基酸;PmChi为斑节对虾几丁质酶,PjChi为日本对虾几丁质酶,LvChi为凡纳滨对虾几丁质酶,FcChi代表中国明对虾几丁质酶。Figure 3. Multiple alignment of predicted amino acid sequence between PmChi-2 and Chi of other crustacean speciesThe domains and catalytic active site are marked by boxes; "● "and"★" stand for the cysteine and aromatic amino acid residues, respectively; PmChi(P.monodon chitinase), PjChi (P.japonicus chitinase), LvChi (L.vannamei chitinase), FcChi (F.chinensiss chitinase)
图 3 PmChi-2与其他甲壳动物的Chi的氨基酸多重序列比对方框内为结构域和催化活性位点;“●”和“★”分别表示半胱氨酸和芳香族氨基酸;PmChi为斑节对虾几丁质酶,PjChi为日本对虾几丁质酶,LvChi为凡纳滨对虾几丁质酶,FcChi代表中国明对虾几丁质酶。Figure 3. Multiple alignment of predicted amino acid sequence between PmChi-2 and Chi of other crustacean speciesThe domains and catalytic active site are marked by boxes; "● "and"★" stand for the cysteine and aromatic amino acid residues, respectively; PmChi(P.monodon chitinase), PjChi (P.japonicus chitinase), LvChi (L.vannamei chitinase), FcChi (F.chinensiss chitinase)多重序列比对显示,PmChi-2与凡纳滨对虾LvChi-2、日本对虾PjChi-2序列的一致性分别为97%、86%。运用MEGA 6.06软件,采用NJ法构建了PmChi-2与其他甲壳动物几丁质酶基因的系统进化树(图 4)。从进化树可以看出Chi-1、Chi-2、Chi-3、Chi-4分别聚为一大支,而Chi-5和Chi-6分别单独聚为一支。PmChi-2在进化树中先与凡纳滨对虾LvChi-2聚为一支,再与日本对虾PjChi-2聚在一起。因此,该研究中克隆的斑节对虾Chi基因进一步确认为PmChi-2。
![]() 图 4 运用Clustal X 2.1和MEGA 6.06软件基于NJ法构建的PmChi-2系统进化树Figure 4. NJ phylogenetic tree of PmChi-2 by Clustal X 2.1 and MEGA 6.06
图 4 运用Clustal X 2.1和MEGA 6.06软件基于NJ法构建的PmChi-2系统进化树Figure 4. NJ phylogenetic tree of PmChi-2 by Clustal X 2.1 and MEGA 6.062.2 PmChi-2在D期不同组织中的表达
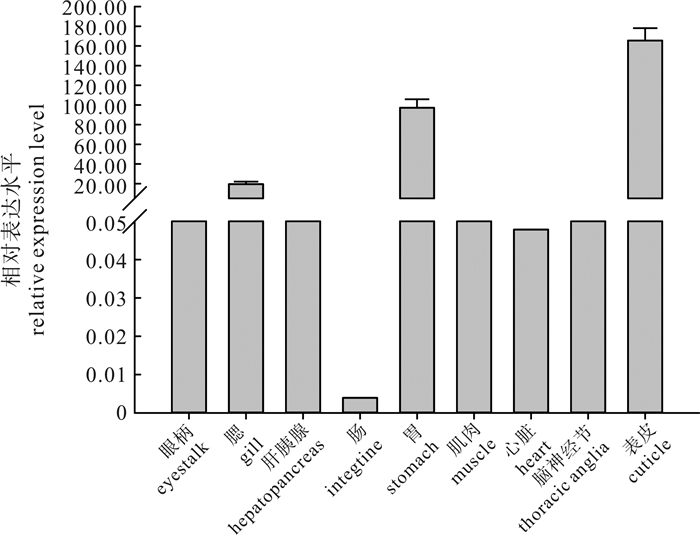
实时荧光定量PCR结果显示,PmChi-2在D期的表皮中表达最高,在胃、鳃、眼柄和腹神经节中也有一定的表达,表达水平高低顺序依次为胃>鳃>胸神经节>眼柄(图 5),在其他组织无表达。
![]() 图 5 PmChi-2基因在D期不同组织中mRNA相对表达水平Figure 5. mRNA relative expression level of PmChi-2 in different tissues of P.monodon at premolt
图 5 PmChi-2基因在D期不同组织中mRNA相对表达水平Figure 5. mRNA relative expression level of PmChi-2 in different tissues of P.monodon at premolt2.3 PmChi-2在蜕皮各期的表达
基于PmChi-2在D期不同组织的相对表达量,选取蜕皮各期的鳃、胃、表皮,研究PmChi-2在各个蜕皮期这3个组织相对表达量的变化。结果显示鳃中PmChi-2在不同蜕皮阶段的相对表达量变化不显著,而胃和表皮中PmChi-2表达量变化呈现显著性差异,只是变化趋势有所不同。鳃中PmChi-2 mRNA的相对表达水平在A期最高(是E期的2.93倍),E期最低,总体来说变化不是很大;胃中PmChi-2 mRNA表达水平在D期最高(是E期的3.10倍),到E期显著下降,A期又显著升高,但仍低于D期,C期又开始下降,但高于E期;表皮中PmChi-2 mRNA表达水平在D期最高(是E期的184.66倍),到E期显著下降,A期又显著升高,但低于D期,C期继续升高,但仍低于D期(图 6)。
![]() 图 6 PmChi-2 mRNA在斑节对虾蜕皮周期中3种组织的相对表达水平Figure 6. Expression of PmChi-2 transcripts in three tissues of P.monodon during the molting cycle
图 6 PmChi-2 mRNA在斑节对虾蜕皮周期中3种组织的相对表达水平Figure 6. Expression of PmChi-2 transcripts in three tissues of P.monodon during the molting cycle2.4 PmChi-2在幼体不同发育阶段的表达
PmChi-2基因在幼体不同发育阶段的表达情况见图 7。从无节幼体一直到糠虾幼体第二期,PmChi-2的表达量没有显著变化,到糠虾幼体第三期,表达量显著升高,且达到最高的表达水平(是无节幼体期的19.65倍),紧接着仔虾第一期又显著下降,仔虾第二期、第六期、第十二期与仔虾第一期表达量没有显著变化。
![]() 图 7 斑节对虾幼体不同发育阶段PmChi-2 mRNA相对表达水平N.无节幼体;Z1.溞状幼体第一期;Z2.溞状幼体第二期;Z3.溞状幼体第三期;M1.糠虾幼体第一期;M2.糠虾幼体第二期;M3.糠虾幼体第三期;P1.仔虾第一期;P2.仔虾第二期;P6.仔虾第六期;P12.仔虾第十二期;仔虾第一期:变态后仔虾第一天,依此类推。Figure 7. Expression of PmChi-2 transcripts at different larval developmental stagesN. nauplius; Z1. zoea1;Z2. zoea2;Z3. zoea3;M1. mysis1; M2. mysis2;M3. mysis3;P1. post larvae1;P2. post larvae2; P6.post larvae6;P12. post larvae12;post larvae1:the first day post-larvae after metamorphosis, and so on.
图 7 斑节对虾幼体不同发育阶段PmChi-2 mRNA相对表达水平N.无节幼体;Z1.溞状幼体第一期;Z2.溞状幼体第二期;Z3.溞状幼体第三期;M1.糠虾幼体第一期;M2.糠虾幼体第二期;M3.糠虾幼体第三期;P1.仔虾第一期;P2.仔虾第二期;P6.仔虾第六期;P12.仔虾第十二期;仔虾第一期:变态后仔虾第一天,依此类推。Figure 7. Expression of PmChi-2 transcripts at different larval developmental stagesN. nauplius; Z1. zoea1;Z2. zoea2;Z3. zoea3;M1. mysis1; M2. mysis2;M3. mysis3;P1. post larvae1;P2. post larvae2; P6.post larvae6;P12. post larvae12;post larvae1:the first day post-larvae after metamorphosis, and so on.3. 讨论
3.1 PmChi-2结构分析
生物信息学分析显示,PmChi-2与其他甲壳动物的Chi的一致性达到78%~97%,说明甲壳动物的Chi整体上趋于保守,并且在昆虫中也比较保守。PmChi-2推导的氨基酸序列包含CBD、GH18催化结构域。CBD主要功能是通过其内部的芳香族氨基酸残基与不溶性几丁质结合,进而提高对几丁质的降解效率[11]。CBD内部保守的芳香族氨基酸残基及带电残基参与糖类的结合,是CBD必需的,直接影响到CBD与糖类的结合活性[4, 11, 28],而内部的6~8个半胱氨酸能形成3~4个二硫键,其中只要有一个半胱氨酸发生突变,就会完全失去几丁质结合活性,而且该结构对几丁质酶基因的稳定性可能起到很大的作用[11, 16, 19, 29]。与已报道甲壳类几丁质酶基因一样,PmChi-2 CBD中也含有6个半胱氨酸残基,能够形成3个内部二硫键,推测对PmChi-2基因的稳定性起到一定的作用。FDGLDLDWEYP是甲壳动物几丁质酶GH18催化结构域中的高度保守Motif,也是几丁质酶的催化活性位点,其中的谷氨酸(E)可以为催化活动提供质子[11, 30-32],若谷氨酸发生突变就会使几丁质酶催化活性丧失[9],因此该位点对几丁质酶催化作用有重要意义。尽管甲壳动物的几丁质酶基因整体上较为保守,但还是存在差异,目前几丁质酶类别的划分主要依据是CBD和催化结构域的组成和顺序[33]。凡纳滨对虾已成功克隆了6种几丁质酶,基本上涵盖了甲壳动物已发现的几丁质酶种类,因此以凡纳滨对虾几丁质酶为例分析其分类。LvChi-1、LvChi-3以及LvChi-5的顺序和组成为1个催化结构域和1个CBD;LvChi-2的顺序和组成为1个CBD和1个催化结构域;LvChi-4的顺序和组成为1个催化结构域和2个CBD;而LvChi-6中目前只发现1个催化结构域[16]。在日本对虾PjChi-2中顺序和组成为1个CBD和1个催化结构域;此研究中斑节对虾Chi-2中顺序和组成也为1个CBD和1个催化结构域,与日本对虾PjChi-2和凡纳滨对虾LvChi-2顺序和组成一致。因此,进一步确认PmChi-2属于Chi-2。
3.2 PmChi-2在蜕皮阶段、幼体不同发育阶段的功能研究
Chi-2相继被报道在甲壳动物的表皮、尾扇、第二触角鳞片中高表达,胃和眼柄等组织也发现有少量表达,但却发现在肝胰腺中微量表达甚至不表达,现有的研究表明其基本功能是参与蜕皮调控[2, 13, 16]。该研究发现PmChi-2最高表达量出现在蜕皮前期的表皮中,此外在胃和鳃中也有一定的表达量,这与以往其他甲壳动物中的Chi-2在组织中的分布基本吻合。进一步确认PmChi-2应该属于Chi-2。
不同蜕皮阶段,PmChi-2基因在鳃组织中表达差异不显著,而在胃、表皮中的表达存在显著性差异。表皮中PmChi-2 mRNA表达水平在D期最高,可能与旧壳中几丁质的降解有关,因为这个时期外壳中的几丁质被大量降解,为即将来临的蜕皮期做最后的准备,这与日本沼虾的研究结果一致[21]。胃中PmChi-2 mRNA表达水平在D期最高,推测可能与旧壳中几丁质的降解与重吸收有关。
斑节对虾幼体要经历无节幼体、溞状幼体、糠虾幼体才能变态为与幼虾以及成虾形态相似的仔虾,糠虾幼体期浮游生活,头向下倒置;仔虾1期开始变为水平游泳以及底栖习性。PmChi-2 mRNA在斑节对虾幼体不同发育阶段中,从无节幼体一直到糠虾幼体第二期,PmChi-2的表达量没有显著变化,到糠虾幼体第三期,表达量显著升高,紧接着到仔虾期又显著下降,表明PmChi-2可能调控着斑节对虾幼体的变态发育,这与中华绒鳌蟹Chi-2的研究结果很类似[11]。不过还需进一步的深入研究才能确定。
该研究克隆了PmChi-2基因的cDNA全长,分析PmChi-2基因在蜕皮周期和幼体发育各期的表达变化,发现PmChi-2基因在斑节对虾蜕皮发育和幼体变态发育中都发挥了一定的作用。但由于缺乏相关的功能验证,故PmChi-2对斑节对虾蜕壳以及幼体变态发育的更为精确的生理功能以及调控机制仍有待进一步深入研究。
-
![]()
图 1 斑节对虾蜕皮过程的A期、C期和D期
A.A期尾扇内肢;C. C期尾扇内肢;D. D期尾扇内肢(×40);A′. A期尾扇内肢;C′. C期尾扇内肢;D′. D期尾扇内肢(×100);S.刚毛干;Sn.刚毛瘤;Sc.刚毛锥体;Nc.新表皮;Ss.新刚毛干;Pp.只看到一点光线的区域
Figure 1. P.monodon molting process at Stage A, C and D
A.postmoult(A) inner uropod; C. intermoult (C) inner uropod; D.premoult (D) inner uropod (×40);A′.postmoult (A) inner uropod; C′.intermoult (C) inner uropod; D′.premoult (D) inner uropod (×100);S. setal shaft; Sn. setal node; Sc. setal cone; Nc. new cuticle; Ss. new setal shaft; Pp. pinpoints of light where new setal node will develop
![]()
图 2 PmChi-2氨基酸序列及cDNA序列展示
双下划线表示几丁质结合结构域;灰色表示GH18催化结构域;粗体表示起始密码子(ATG)和终止密码子(TAA);斜体表示PolyA结构。
Figure 2. Predicted amino acid and nucleotide sequence of PmChi-2 cDNA
Double-underlines indicate chitin binding domain(CBD); GH18 domain is shown in gray; the initiation codon (ATG) and termination codon (TAA) are highlighted in bold; the poly A signal sequence is italicized.
![]()
图 3 PmChi-2与其他甲壳动物的Chi的氨基酸多重序列比对
方框内为结构域和催化活性位点;“●”和“★”分别表示半胱氨酸和芳香族氨基酸;PmChi为斑节对虾几丁质酶,PjChi为日本对虾几丁质酶,LvChi为凡纳滨对虾几丁质酶,FcChi代表中国明对虾几丁质酶。
Figure 3. Multiple alignment of predicted amino acid sequence between PmChi-2 and Chi of other crustacean species
The domains and catalytic active site are marked by boxes; "● "and"★" stand for the cysteine and aromatic amino acid residues, respectively; PmChi(P.monodon chitinase), PjChi (P.japonicus chitinase), LvChi (L.vannamei chitinase), FcChi (F.chinensiss chitinase)
![]()
图 4 运用Clustal X 2.1和MEGA 6.06软件基于NJ法构建的PmChi-2系统进化树
Figure 4. NJ phylogenetic tree of PmChi-2 by Clustal X 2.1 and MEGA 6.06
![]()
图 5 PmChi-2基因在D期不同组织中mRNA相对表达水平
Figure 5. mRNA relative expression level of PmChi-2 in different tissues of P.monodon at premolt
![]()
图 6 PmChi-2 mRNA在斑节对虾蜕皮周期中3种组织的相对表达水平
Figure 6. Expression of PmChi-2 transcripts in three tissues of P.monodon during the molting cycle
![]()
图 7 斑节对虾幼体不同发育阶段PmChi-2 mRNA相对表达水平
N.无节幼体;Z1.溞状幼体第一期;Z2.溞状幼体第二期;Z3.溞状幼体第三期;M1.糠虾幼体第一期;M2.糠虾幼体第二期;M3.糠虾幼体第三期;P1.仔虾第一期;P2.仔虾第二期;P6.仔虾第六期;P12.仔虾第十二期;仔虾第一期:变态后仔虾第一天,依此类推。
Figure 7. Expression of PmChi-2 transcripts at different larval developmental stages
N. nauplius; Z1. zoea1;Z2. zoea2;Z3. zoea3;M1. mysis1; M2. mysis2;M3. mysis3;P1. post larvae1;P2. post larvae2; P6.post larvae6;P12. post larvae12;post larvae1:the first day post-larvae after metamorphosis, and so on.
-
[1] 黄建华, 马之明, 周发林, 等.南海北部野生斑节对虾卵巢解剖结构及组织学的研究[J].南方水产, 2005, 1 (3): 49-53. http://www.cqvip.com/QK/96022A/200603/22020519.html [2] PROESPRAIWONG P, TASSANAKAJON A, RIMPHANITCHAYAKIT V. Chitinases from the black tiger shrimp Penaeus monodon: phylogenetics, expression and activities [J].Comp Biochem Phys B, 2010, 156 (2): 86-96. doi: 10.1016/j.cbpb.2010.02.007
[3] KURITA K. Chitin and chitosan: functional biopolymers from marine crustaceans [J].Mar Biotechnol, 2006, 8(3): 203-226. doi: 10.1007/s10126-005-0097-5
[4] KRAMER K J, KOGA D. Insect chitin: physical state, synthesis, degradation and metabolic regulation [J].Insect Biochem, 1986, 16 (6): 851-877. doi: 10.1016/0020-1790(86)90059-4
[5] FUKAMIZO T, KRAMER K J. Mechanism of chitin hydrolysis by the binary chitinase system in insect moulting fluid [J].Insect Biochem, 1985, 15 (2): 141-145. doi: 10.1016/0020-1790(85)90001-0
[6] LI D C. Review of fungal chitinases [J].Mycopathologia, 2006, 161 (6): 345-360. doi: 10.1007/s11046-006-0024-y
[7] MALI B, MöHRLEN F, FROHME M, et al. A putative double role of a chitinase in a cnidarian: pattern formation and immunity [J].Dev Comp Immunol, 2004, 28 (10): 973-981. doi: 10.1016/j.dci.2004.04.002
[8] DAHIYA N, TEWARI R, HOONDAL G S. Biotechnological aspects of chitinolytic enzymes: a review [J].Appl Microbiol Biot, 2006, 71 (6): 773-782. doi: 10.1007/s00253-005-0183-7
[9] ARAKANE Y, MUTHUKRISHNAN S. Insect chitinase and chitinase-like proteins [J].Cell Mol Life Sci, 2010, 67 (2): 201-216. doi: 10.1007/s00018-009-0161-9
[10] 姚琴琴, 杨志刚, 王瑶, 等.中华绒螯蟹几丁质酶基因HXchit全长cDNA克隆及其在蜕皮过程中的表达分析[J].中国水产科学, 2015, 22 (2): 185-195. http://www.doc88.com/p-0713138954129.html [11] LI X G, XU Z Q, ZHOU G, et al. Molecular characterization and expression analysis of five chitinases associated with molting in the Chinese mitten crab, Eriocheir sinensis [J].Comp Biochem Phys B, 2015, 187: 110-120. doi: 10.1016/j.cbpb.2015.05.007
[12] TAN S H, DEGNAN B M, LEHNERT S A. The Penaeus monodon chitinase 1 gene is differentially expressed in the hepatopancreas during the molt cycle [J].Mar Biotechnol, 2000, 2 (2): 126-135. https://www.sciencedirect.com/science/article/pii/S1050464817306599
[13] WATANABE T, KONO M. Isolation of a cDNA encoding a chitinase family protein from cuticular tissues of the kuruma prawn Penaeus japonicus [J].Zool Sci, 1997, 14 (1): 65-68. doi: 10.2108/zsj.14.65
[14] WATANABE T, KONO M, AIDA K, et al. Isolation of cDNA encoding a putative chitinase precursor in the kuruma prawn Penaeus japonicus [J].Mol Mar Biol Biotechnol, 1996, 5 (4): 299-303. https://www.sciencedirect.com/science/article/pii/S105046480300144X
[15] WATANABE T, KONO M, AIDA K, et al. Purification and molecular cloning of a chitinase expressed in the hepatopancreas of the penaeid prawn Penaeus japonicus [J].Bba-Proteins Proteom, 1998, 1382 (2): 181-185. https://www.sciencedirect.com/science/article/pii/S1050464808001861
[16] HUANG Q S, YAN J H, TANG J Y, et al. Cloning and tissue expressions of seven chitinase family genes in Litopenaeus vannamei [J].Fish Shellfish Immunol, 2010, 29 (1): 75-81. doi: 10.1016/j.fsi.2010.02.014
[17] ROCHA J, GARCIA-CARREÑO F L, MUHLIA-ALMAZÁN A, et al. Cuticular chitin synthase and chitinase mRNA of whiteleg shrimp Litopenaeus vannamei during the molting cycle [J].Aquaculture, 2012, 330: 111-115. https://www.sciencedirect.com/science/article/pii/S1096495916301567
[18] ZHANG J Q, SUN Y Y, LI F H, et al. Molecular characterization and expression analysis of chitinase (Fcchi-3) from Chinese shrimp, Fenneropenaeus chinensis [J].Mol Biol Rep, 2010, 37 (4): 1913-1921. doi: 10.1007/s11033-009-9633-0
[19] SALMA U, UDDOWLA M H, KIM M, et al. Five hepatopancreatic and one epidermal chitinases from a pandalid shrimp (Pandalopsis japonica): cloning and effects of eyestalk ablation on gene expression [J].Comp Biochem Phys B, 2012, 161 (3): 197-207. doi: 10.1016/j.cbpb.2011.11.005
[20] DUAN Y, LIU P, LI J, et al. The responsive expression of a chitinase gene in the ridge tail white prawn Exopalaemon carinicauda against Vibrio anguillarum and WSSV challenge [J].Cell Stress Chaperon, 2014, 19 (4): 549-558. doi: 10.1007/s12192-013-0488-x
[21] ZHANG S Y, JIANG S F, XIONG Y W, et al. Six chitinases from oriental river prawn Macrobrachium nipponense: cDNA characterization, classification and mRNA expression during post-embryonic development and moulting cycle [J].Comp Biochem Phys B, 2014, 167: 30-40. doi: 10.1016/j.cbpb.2013.09.009
[22] 张凤, 吕建建, 刘萍, 等.三疣梭子蟹几丁质酶基因克隆鉴定及在低盐胁迫和蜕皮周期中的表达分析[J].海洋与湖沼, 2015, 46 (4): 948-957. http://www.wenkuxiazai.com/doc/0cb35a8427284b73f3425096.html [23] 王伟, 吴旭干, 潘桂平, 等.三疣梭子蟹几丁质酶基因的克隆及其在蜕皮过程中的表达分析[J].水产学报, 2015, 39 (9): 1291-1301. http://www.cqvip.com/QK/90072X/201504/665890308.html [24] OWENS L. A simple technique to stage the moult of Penaeus monodon [J].Asian Fish Sci, 2011, 24: 1-11. http://www.asianfisheriessociety.org/publication/abstract.php?id=50
[25] 傅明骏, 赵超, 杨其彬, 等.斑节对虾过氧化氢酶基因全长cDNA克隆及表达分析[J].南方水产科学, 2015, 11 (6): 107-113. http://www.schinafish.cn/CN/abstract/abstract9278.shtml [26] 王艳, 傅明骏, 赵超, 等.斑节对虾GTF2H4基因的分子克隆及表达分析[J].南方水产科学, 2016, 12 (2): 36-43. http://www.schinafish.cn/CN/abstract/abstract9301.shtml [27] 赵超, 傅明骏, 江世贵, 等.斑节对虾细胞周期蛋白E基因的克隆与表达分析[J].中国水产科学, 2014, 21 (3): 464-473. http://d.wanfangdata.com.cn/Periodical_zgsckx201505006.aspx [28] TJOELKER L W, GOSTING L, FREY S, et al. Structural and functional definition of the human chitinase chitin-binding domain [J].J Biol Chem, 2000, 275 (1): 514-520. doi: 10.1074/jbc.275.1.514
[29] AKAGI K, WATANABE J, HARA M, et al. Identification of the substrate interaction region of the chitin-binding domain of Streptomyces griseus chitinase C [J].J Biochem, 2006, 139 (3): 483-493. doi: 10.1093/jb/mvj062
[30] TEWS I, van SCHELTINGA A C T, PERRAKIS A, et al. Substrate-assisted catalysis unifies two families of chitinolytic enzymes [J].J Am Chem Soc, 1997, 119 (34): 7954-7959. doi: 10.1021/ja970674i
[31] HUANG X, ZHANG H, ZEN K C, et al. Homology modeling of the insect chitinase catalytic domain-oligosaccharide complex and the role of a putative active site tryptophan in catalysis [J].Insect Biochem Mol Biol, 2000, 30 (2): 107-117. doi: 10.1016/S0965-1748(99)00101-0
[32] LU Y, ZEN K C, MUTHUKRISHNAN S, et al. Site-directed mutagenesis and functional analysis of active site acidic amino acid residues D142, D144 and E146 in Manduca sexta (tobacco hornworm) chitinase [J].Insect Biochem Mol Biol, 2002, 32 (11): 1369-1382. doi: 10.1016/S0965-1748(02)00057-7
[33] 吕黎, 宁黔冀.甲壳动物几丁质酶基因结构与功能的研究进展[J].生理科学进展, 2011, 42(6): 457-459. http://d.wanfangdata.com.cn/Periodical/slkxjz201106013 -
期刊类型引用(6)
1. 张娣,金鑫,程润强,耿昌盛,李晔. 甲壳动物几丁质酶基因的结构与功能. 生物学杂志. 2024(02): 91-96 .  百度学术
百度学术
2. 沙伟,秦瑞峰,孟灵军,信雨萌,马天意,张梅娟. 砂藓(Racomitrium canescens)抗旱相关基因RcAOS2的克隆及表达分析. 分子植物育种. 2020(04): 1131-1137 . 百度学术
3. 郝淑贤,杨肖杰,黄卉,李来好,潘创,杨贤庆,岑剑伟. 虾类剥壳加工现状及预处理便捷剥壳技术. 南方水产科学. 2020(04): 121-128 . 本站查看
4. 沙伟,蒋卓君,张梅娟,马天意. 砂藓抗旱相关基因RcSOD的克隆及表达分析. 基因组学与应用生物学. 2019(05): 2144-2151 . 百度学术
5. 周发林,杨其彬,黄建华,姜松,杨丽诗,张汤生,江世贵. 斑节对虾耐氨氮和淡水应激性状的遗传参数估计. 南方水产科学. 2019(05): 63-68 . 本站查看
6. 吴勉之,杨丽诗,周发林,黄建华,姜松,杨其彬,江世贵. 斑节对虾2种高血糖激素家族基因的基因组序列分析和表达研究. 南方水产科学. 2018(04): 27-36 . 本站查看
其他类型引用(7)
 下载:
下载:






计量
- 文章访问数: 3481
- HTML全文浏览量: 1094
- PDF下载量: 844
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
