
Effects of linolenic acid level on growth performance, digestive enzymes and antioxidant capacity of juvenile golden pompano (Trachinotus ovatus)
-
摘要:
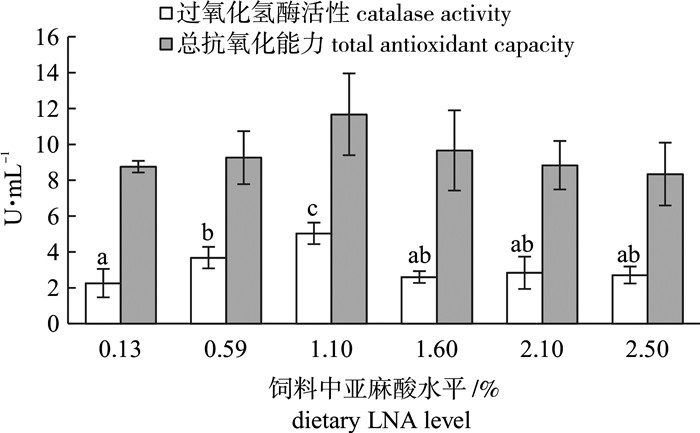
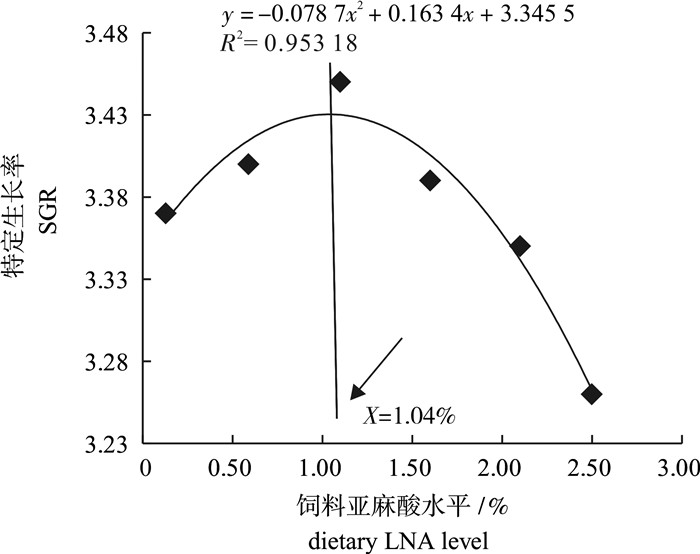
探讨了饲料中不同水平的亚麻酸对卵形鲳鲹(Trachinotus ovatus)幼鱼生长性能、消化酶活性及抗氧化能力的影响。选取均体质量为(10.38±0.10)g的卵形鲳鲹幼鱼随机分配6组,每组3个平行,每个平行20尾鱼。配制6种亚麻酸质量分数分别为0.13%、0.59%、1.10%、1.60%、2.10%和2.50%的实验饲料饲喂卵形鲳鲹,每天饲喂2次,周期为56 d。结果表明,饲喂含0.59%和1.10%亚麻酸饲料的实验鱼增重率、特定生长率显著高于2.1%组和2.5%组。亚麻酸水平显著影响了实验鱼的肝体比、脏体比、腹脂率、肥满度及其成活率。0.59%组实验鱼的胃蛋白酶显著高于0.13%组和2.50%组。1.10%组的淀粉酶活性显著低于其他各组。血清总抗氧化能力和过氧化氢酶活性随着饲料中亚麻酸含量的升高呈现出先升高后降低的趋势。以特定生长率为评价指标,二次曲线回归分析得出卵形鲳鲹幼鱼对亚麻酸的最适需求量为饲料干质量的1.04%。
Abstract:To determine the effects of dietary linolenic acid (LNA) level on growth performance, digestive enzymes and antioxidant capacity of juvenile golden pompano (Trachinotus ovatus), we assigned the fish [(10.38±0.10) g] randomly into six groups with three replicates per group and 20 fish per replicate. Six diets were formulated with six levels of LNA (dry matter, 0.13%, 0.59%, 1.10%, 1.60%, 2.10% and 2.50%). The fish were fed twice daily for 56 d. The results show that the weight gain and specific growth rate (SGR) of fish fed diet with 0.59% and 1.10% LNA were significantly higher than those in group 2.1% and group 2.5%.Viscerosomatic index (VSI), hepatosomatic index (HSI), condition factor, abdominal fat and survival rate of fish were affected by dietary LNA levels significantly. The activity of pepsin of fish fed diet with 0.59% LNA was significantly higher than those fed with 0.13% and 2.50% LNA. The activity of amylase of fish fed diet with 1.10% LNA was significantly lower than those in the other groups. The total antioxidant capacity and catalase activity of serum showed an increasing trend and then decreased with increasing dietary LNA level. Quadratic regression analysis on SGR indicates that the optimum dietary LNA level for optimal growth of juvenile golden pompano was 1.04% of dry matter.
-
Keywords:
- golden pompano /
- linolenic acid /
- growth performance /
- digestive enzymes /
- antioxidant capacity
-
种群遗传结构是指种群内部等位基因频率的异质性[1]。种群遗传结构研究的目的在于探明物种在特定栖息范围内的种群数量,以及各个种群的遗传特征及他们之间的差异和联系。在渔业管理中,需要根据种群的划分来分配不同的捕捞压力[1-2],防止过度捕捞的发生和种群适应力的下降或衰退[3]。
圆舵鲣(Auxis rochei)属鲈形目、鲭科,广泛分布于大西洋、印度洋和西太平洋的浅海海域[4-5],是一种中上层洄游的小型金枪鱼类,具有重要的经济价值。FAO数据显示,以舵鲣类为代表的小型金枪鱼类资源的全球捕捞量逐年攀升[6]。近年来的灯光罩网探捕[7-8]也发现南海蕴藏有丰富的圆舵鲣资源,其中2012年9月~10月在南海中南部海域的探捕调查显示圆舵鲣为重要渔获[9]。而目前关于南海圆舵鲣的研究较少,仅见于生长发育[10]、资源生物学[11-12]和营养分析[13]等方面,尚未见种群遗传相关的报道。线粒体控制区(D-loop)为非编码区,进化速率快,多态性高,是探讨种内遗传分化较为理想的分子标记,应用广泛。该研究利用D-loop序列作为遗传标记,分析了南海圆舵鲣的种群遗传结构和遗传多样性,旨在为资源的合理开发和可持续利用提供参考。
1. 材料与方法
1.1 样本采集
圆舵鲣个体样本通过灯光罩网渔船采集,经形态学鉴定和生物学测量后冰冻带回实验室,剪取背部肌肉置于-20 ℃保存。采样点位置示意图见图 1,样本采集时间、位点经纬度及样本量见表 1。
![]() 图 1 圆舵鲣采样点示意图南海九段线参见彩色宣传页的调查区域图Fig. 1 Map of sampling sites for A.rochei in the South China SeaFor information about the nine-dash line of the South China Sea, see the survey area map on the colored leaflet.表 1 南海圆舵鲣样本信息及D-loop区遗传多样性参数Table 1 Specimen information of A.rochei and genetic diversity parameters based on D-loop sequences
图 1 圆舵鲣采样点示意图南海九段线参见彩色宣传页的调查区域图Fig. 1 Map of sampling sites for A.rochei in the South China SeaFor information about the nine-dash line of the South China Sea, see the survey area map on the colored leaflet.表 1 南海圆舵鲣样本信息及D-loop区遗传多样性参数Table 1 Specimen information of A.rochei and genetic diversity parameters based on D-loop sequences采样点
sampling site采样时间
sampling date经度/纬度
longitude/latitude样本量(N)
number of samples单倍型数量(H)
number of haplotypes多态性位点数(S)
number of polymorphic sites单倍型多样性
(h±SD) haplotype diversity核苷酸多样性
(π±SD) nucleotide diversityA 2014-11-22 108°03′E /18°12′N 30 30 103 1.000 0±0.008 6 0.034 966±0.017 627 B 2015-03-17 112°58′E /18°29′N 42 34 130 0.958 2±0.025 3 0.037 692±0.018 765 C 2015-04-22 116°53′E /20°28′N 36 36 125 1.000 0±0.006 5 0.035 815±0.017 933 D 2013-03-27 115°32′E /11°01′N 38 35 116 0.994 3±0.008 2 0.037 030±0.018 494 E 2013-03-20 110°31′E / 5°34′N 32 32 103 1.000 0±0.007 8 0.034 327±0.017 276 F 2014-11-28 112°03′E /20°14′N 10 9 31 0.977 8±0.054 0 0.034 347±0.018 732 G 2013-04-05 117°28′E /15°04′N 13 13 83 1.000 0±0.030 2 0.041 235±0.021 749 总计 total - - 201 185 202 0.997 2±0.001 5 0.036 807±0.017 981 1.2 实验方法
1.2.1 基因组DNA提取与检测
从每个样品中剪取30 mg左右肌肉置于1.5 mL离心管内,使用快速组织细胞破碎仪(BulletBlender STORM,美国)研磨后,用海洋动物基因组DNA抽提试剂盒(北京天根)提取基因组DNA。取5 μL DNA样品用1%TBE琼脂糖凝胶进行电泳,检测其基因组DNA完整性和纯度。剩余样品置于-20 ℃冰箱内保存备用。
1.2.2 PCR扩增与测序
PCR反应体积为50 μL,其中Taq PCR Master Mix(上海生工)25 μL,包括Taq DNA聚合酶、dNTP、PCR Buffer、PCR stabilizers、gel loading和核酸染液;模板DNA 1 μL,正反向引物(Ath-Dloop-F与Ath-Dloop-R[14])各1 μL(浓度为10 μmol · L-1),加灭菌超纯水至总体积50 μL。PCR反应在热循环仪(Eppendorf,德国产)上进行,反应程序为94 ℃预变性3 min,之后进行30个循环,每个循环包括94 ℃变性30 s、56 ℃退火30 s、72 ℃延伸1 min,最后72 ℃延伸5 min。扩增产物经电泳检验后,选取单一条带产物进行序列测定,测序引物与扩增引物相同。
1.3 数据处理
1.3.1 序列编辑及进化模型确定
将测序获得的序列经BioEdit 3.3[15]编排后,用MEGA 6.0[16]中ClustalW进行比对(alignment),多重排列的参数设置为程序默认值。核苷酸替代的最适模型通过软件Modeltest 3.7[17]来选择。基于AIC(Akaike Information Criterion) 标准从56种核苷酸替代模型中选出最适模型及相关参数,用于后续系统发生关系重建和种群遗传结构分析。
1.3.2 遗传多样性
利用Dnasp 5.0[18]和Arlequin 3.5[19]软件计算各个采样点(地理群体)样本的遗传多样性指数,包括单倍型数目(H)、多态位点数目(S)、单倍型多样性(h)和核苷酸多样性(π)等。
1.3.3 种群遗传结构
在Arlequin 3.5中采用分子方差分析(analysis of molecular variances,AMOVA)[20]来评价种群遗传变异水平与地理格局的相关性;进化模型选择1.3.1中确定的具有非变异位点(invariable sites)和Gamma参数(Gamma shape parameter)的TrN模型[21](TrN+I+G模型),显著性用10 000次重复抽样来检验。通过计算两两群体间的分化固定指数FST来检验群体间遗传距离的大小,利用Dnasp 5.0计算群体间的基因流。若群体间遗传分化不显著,则进行种群分化测试(exact test of population differentiation)。此测试基于随机交配群的单倍型分布频率的观测值低于期望值这个预定假设。若检测结果不显著,则接受群体间为随机交配群的假说。种群分化测试在Arlequin 3.5中计算,显著性通过10 000次重复抽样来检验。鉴于采样点F和G的样本量比较小,为避免样本量不对等带来的误差,这2个群体不参与种群遗传结构和历史动态的分析。
1.3.4 系统发生关系
利用Dnasp 5.0生成所有样本的D-loop区序列单倍型,将扁舵鲣(A.thazard)D-loop区同源序列作为系统分析的外类群,基于邻接法(neighbor-joining,NJ)在MEGA 6.0中构建样本单倍型的系统发育树。进化模型同样选择1.3.1中确定的TrN+I+G模型,系统树各分支的可靠性采用1 000次重复抽样评估。
1.3.5 种群历史动态
在Arlequin 3.5中使用核苷酸不配对分布分析(mismatch distribution)[22-23]和中性检验(neutrality tests)来检测南海圆舵鲣的种群历史动态,以检验种群历史上是否存在扩张事件。核苷酸不配对分布分析中,选取突然扩张模型(sudden expansion model)基于最小方差法来检验观测值和种群扩张模型下的期望分布之间是否一致。模型的有效性由粗糙指数[24](Harpending′s raggedness index,HRI)进行评估,HRI的统计检验由10 000次重复抽样获得。另外由于核苷酸不配对分布可能具有保守性[25],采用两种广泛使用的中性检验:Tajima′s D检验[26]和Fu′s FS检验[27]来检测种群是否严格遵循中性理论。
如检测到种群扩张事件,则依据公式t=τ/2 u[28]计算扩张开始到现在经历的代数。其中τ为根据核苷酸不配对分布计算获得的扩张时间参数,t指自扩张以来所经历的代数,u为序列的突变速率,可通过公式u=2μk计算得到,其中k代表序列长度,μ代表每个核苷酸位点的突变速率,采用相关研究中普遍采用的每百万年3% ~10%的数值[29]。实际扩张时间T=t×代时,圆舵鲣代时以2.6年[5]为计。
2. 结果
2.1 序列特征
共获得南海圆舵鲣7个地理群体共201尾样本的线粒体D-loop区序列,截取5′端高变区644 bp序列进行分析,未发现插入与缺失碱基。碱基平均组成比例为A=31.9%、T=31.5%、C=20.8%和G=15.8%,A+T含量(63.4 %)明显高于C+G含量(36.6 %)。
2.2 单倍型分布及遗传多样性
样本整体共包含202个多态性位点,核苷酸多样性指数为0.036 807 (SD=0.017 981)。201条序列一共检测到185个单倍型,单倍型多样性指数为0.997 2(SD=0.001 5)。绝大多数单倍型(180个)只属于一个个体,仅有5个单倍型被不同的个体共享(2~10个)。除位点E的个体外,其余各地理群体均互有共享单倍型。南海圆舵鲣遗传多样性参数如表 1所示,各个地理群体均呈现出非常高的单倍型多样性(0.958 2~1.000 0)和较高的核苷酸多样性(0.034 327~0.041 235)。
2.3 种群遗传结构
南海圆舵鲣地理群体D-loop序列的AMOVA结果(表 2)显示,群体间的遗传变异仅占所有遗传变异的极少部分,绝大部分的遗传变异(98.33%)来源于群体内部。样本总体分化系数为0.016 74,基因流为14.68,表明群体间基因交流频繁,不存在明显的遗传分化。两两地理群体间遗传分化系数FST及显著性见表 3,FST除了群体B与其他群体之间为显著(但数值较小)之外,其余两两群体间的FST均为很小的值甚至是负值,且统计检验不显著。随机交配假设检验的显著性(表 4)显示大部分群体间符合单倍型随机分布的假设,虽然B群体偏离假设,但整体样本的检测结果(P=1.00)表明南海圆舵鲣符合随机交配群的假设,与AMOVA结果一致。
表 2 圆舵鲣5个地理群体D-loop区遗传变异的分子方差分析Table 2 Analysis of molecular variance for five populations of A.rochei based on D-loop sequences变异来源
source of variation自由度
degree of freedom变异百分比
percentage of variation分化系数
F statisticsP值
P value群体间 among populations 4 1.67 FST = 0.016 74 0.000 50 群体内 within populations 173 98.33 所有样本 total 177 表 3 圆舵鲣两两地理群体间D-loop区的遗传分化系数(对角线下方)及显著性水平(对角线上方)Table 3 Pairwise FST (below diagonal) and P values (above diagonal) among geographic populations of A.rochei based on D-loop sequencesA B C D E A 0.004 36 0.056 23 0.327 59 0.429 86 B 0.036 45 0.001 09 0.002 48 0.005 35 C 0.015 65 0.033 77 0.847 24 0.336 11 D 0.002 12 0.034 59 -0.007 26 0.645 38 E -0.000 13 0.033 57 0.001 82 -0.003 69 表 4 圆舵鲣两两地理群体间随机交配假设检验的显著性水平Table 4 P values of exact test of sample differentiation of A.rochei based on D-loop haplotype frequenciesA B C D B 0.000 01 C 1.000 00 0.000 01 D 0.128 85 0.000 01 0.128 70 E 1.000 00 0.001 60 1.000 00 0.112 95 2.4 系统发生关系
利用D-loop序列单倍型基于邻接法(NJ)构建的南海圆舵鲣的系统发育树见图 2。每个采样点的单倍型均广泛分散于系统树的各个分支,系统树的拓扑结构很浅并且大部分分支的支持率(自展值)均不高,未形成分化显著的支系结构。
![]() 图 2 圆舵鲣D-loop区单倍型邻接系统发育树每一枝的末端代表单倍型来源群体;各分支标记大于70%的自展值Fig. 2 Neighbor-joining tree for D-loop haplotypes of A.thazardThe tails of branches represent the populations for each haplotypes. Bootstrap supports of >70% are shown at nodes.
图 2 圆舵鲣D-loop区单倍型邻接系统发育树每一枝的末端代表单倍型来源群体;各分支标记大于70%的自展值Fig. 2 Neighbor-joining tree for D-loop haplotypes of A.thazardThe tails of branches represent the populations for each haplotypes. Bootstrap supports of >70% are shown at nodes.2.5 种群历史动态
从核苷酸不配对分布的参数估算(表 5)可见,圆舵鲣样本总体的吻合度检验的HRI值不显著(P>0.05),表明核苷酸不配对分布符合假设(Rogers and Harpending的种群扩张模型)。总体样本两两序列差异与发生频率的关系图(图 3)显示,核苷酸不配对分布呈明显的单峰分布,且观测值与突然扩张模型下的期望值非常吻合,表明种群在历史上经历过快速扩张。中性检验的Tajima′s D值为不显著的微小负值,但Fu′s FS值在所有群体中均呈现极显著或显著的负值(表 6),显著偏离中性理论下的Wright-Fisher模型,同样表明南海圆舵鲣经历过种群的扩张事件。根据扩张时间参数(τ)的观测值21.8,估算出种群的扩张时间约发生在22万~73万年前。
表 5 圆舵鲣D-loop区序列核苷酸不配对分布分析的参数估计值Table 5 Mismatch distribution parameter estimates for A.rochei based on D-loop sequence核苷酸不配对分布 mismatch distribution 吻合度检验 goodness-of-fit test 扩张时间
τ初始值
θ0最终值
θ1平方和
SSD显著性
P粗糙指数
HRI显著性
PA 21.7 0.003 52 248.325 00 0.001 82 0.817 20 0.003 80 0.926 20 B 19.8 3.636 91 152.992 97 0.004 68 0.202 30 0.011 64 0.016 80 C 21.3 0.047 46 387.304 30 0.001 53 0.713 70 0.004 09 0.710 80 D 22.7 0.007 03 189.716 41 0.001 71 0.740 30 0.004 31 0.653 60 E 19.9 0.451 76 703.750 00 0.001 40 0.818 30 0.004 54 0.735 10 总计 total 21.8 0.029 88 219.521 48 0.000 37 0.680 50 0.001 35 0.831 20 ![]() 图 3 圆舵鲣D-loop区序列单倍型核苷酸不配对分布曲线Fig. 3 Mismatch distribution of D-loop haplotypes for A.rochei表 6 圆舵鲣D-loop区序列的Tajima′s D和Fu′s FS统计值及显著性水平Table 6 Tajima′s D, Fu′s FS statistics, corresponding P values for A.rochei based on D-loop sequences
图 3 圆舵鲣D-loop区序列单倍型核苷酸不配对分布曲线Fig. 3 Mismatch distribution of D-loop haplotypes for A.rochei表 6 圆舵鲣D-loop区序列的Tajima′s D和Fu′s FS统计值及显著性水平Table 6 Tajima′s D, Fu′s FS statistics, corresponding P values for A.rochei based on D-loop sequencesTajima′s D Fu′s FS D P FS P A -0.858 93 0.208 60 -15.264 13 0.000 40 B -1.044 75 0.145 60 -7.948 97 0.022 00 C -1.170 23 0.113 00 -20.597 32 0.000 01 D -0.851 71 0.203 60 -13.510 01 0.000 80 E -0.840 11 0.209 60 -17.268 99 0.000 01 总计 total -1.215 72 0.089 00 -23.701 30 0.003 60 3. 讨论
物种的遗传多样性是其生存适应和进化的前提,也是评价物种资源状况和进行合理利用的重要依据。在渔业资源开发和管理中,对其遗传多样性水平的监测是不可忽视的部分。遗传多样性的下降会降低鱼类对不良环境的适应力,威胁到种群的可持续性发展[3]。该研究检测到南海海域圆舵鲣具有很高的单倍型多样性(h=0.997 2)和较高的核苷酸多样性(π=0.036 8),与同样基于D-loop序列分析的南海其他鱼类相比较,遗传多样性水平与扁舵鲣(h=0.999 5;π=0.019 1)[14]、短尾大眼鲷(Priacanthus macracanthus)(h=0.990 3;π=0.094 2)[30]和红鳍笛鲷(Lutjanus erythropterus)(h=0.997 0;π=0.030 0)[31]相当,明显高于黄斑胡椒鲷(Plecto-rhinchus flavomaculatus)(h=0.665 5;π=0.005 4)[32]。南海海域圆舵鲣丰富的遗传多样性可能与其繁殖较快、资源量和有效群体较大并且受到的捕捞压力较小有关。其种质资源良好,体现出较强的环境适应能力和进化潜力[33]。
南海圆舵鲣中性检验的Tajima′s D值虽不显著,但对种群扩张非常敏感的Fu′s FS检验产生了绝对值较大的负的FS值,且所有群体和整体的FS值在统计意义上均为显著或极显著水平,表明其显著偏离稳定种群模型。核苷酸不配对分布的单峰形曲线同样表明南海圆舵鲣历史上曾经历过种群的快速扩张,时间约发生在22万~73万年前(更新世晚期)。第四纪晚期,西太平洋的边缘海受冰期—间冰期交替气候的影响[34],海平面反复升降[35],在更新世冰期,南海海平面下降变成一个封闭的内陆海[36],圆舵鲣栖息地缩小。随后的间冰期海平面上升,圆舵鲣可能在较大的范围发生重新殖化,种群得以迅速扩张。
种群是渔业资源开发和管理的基本单位。鱼类种群遗传结构的研究有助于种群的判定和合理划分渔业管理单元,指导可持续发展的渔业资源开发。南海圆舵鲣样本单倍型的系统发生关系分析显示不存在显著遗传分化的支系,地理群体的AMOVA也未发现明显的种群遗传结构。整体样本FST仅为0.016 74,依据FREELAND[37]和WRIGHT[38-39],遗传分化系数FST < 0.05表明群体间无遗传分化;基因流>4,群体就是一个随机的单位。该研究中南海圆舵鲣FST为0.016 74,个体间基因流高达14.68,表明群体间基因流动相当强烈,是一个随机交配的单一种群(unit population)。
在缺乏阻碍扩散因素的海洋环境中,许多生物的浮游性卵、幼体和成体可以借助洋流扩散,使群体间产生频繁的基因交流,从而导致海洋生物在较大的地理尺度内呈现很低的遗传分化[40-42]。圆舵鲣是一种洄游性很强的中上层鱼类,成体的移动范围很大。产卵场遍布南海北部[12, 43],中沙、西沙和南沙等岛礁浅海水域[44-45],卵和仔稚鱼可随季风主导的海洋环流、水平环流、沿岸流[46-47]及部分区域的上升流等海流而扩散,从而形成基因交流和遗传均质化。依据LAIKRE等[1],这种非分化种群(no differentiation),在渔业开发和管理上可视为一个管理单元(management units,MUs)。然而,南海海域仅仅是圆舵鲣的部分分布区,其很可能与相邻海域分布的个体同属一个种群。今后需要通过扩大研究范围,结合更多的标记如SSR、SNP等分析技术全面评估圆舵鲣的种群遗传结构,为渔业管理措施的制定提供支撑。
-
![]()
图 1 饲料中亚麻酸水平与卵形鲳鲹幼鱼特定生长率的关系
Figure 1. Relation between LNA level and SGR of juvenile T.ovatus
![]()
图 2 饲料中亚麻酸水平对卵形鲳鲹幼鱼抗氧化能力的影响
Figure 2. Effects of LNA level on antioxidant capacity of juvenile T.ovatus
表 1 实验饲料配方和化学组成(饲料干质量)
Table 1 Formulation and chemical proximate composition of experimental diets (dry matter basis)
% 原料
ingredient组别group 0.13% 0.59% 1.10% 1.60% 2.10% 2.50% 鱼粉1 fish meal 23 23 23 23 23 23 大豆浓缩蛋白2 SPC 20 20 20 20 20 20 豆粕soybean meal 14 14 14 14 14 14 花生粕peanut meal 11 11 11 11 11 11 啤酒酵母brewer′s yeast 3 3 3 3 3 3 面粉wheat flour 16 16 16 16 16 16 鱼油3 fish oil 1.4 1.4 1.4 1.4 1.4 1.4 防腐剂ethoxyquin 0.1 0.1 0.1 0.1 0.1 0.1 维生素4 vitamin premix 1 1 1 1 1 1 矿物质5 mineral premix 1 1 1 1 1 1 氯化胆碱choline chloride(50%) 0.5 0.5 0.5 0.5 0.5 0.5 猪油lard oil 9 7.9 6.8 5.7 4.6 3.5 亚麻籽油6 linseed oil 0 1.1 2.2 3.3 4.4 5.5 营养成分分析proximate analysis 干物质dry matter 92.02 92.14 92.29 92.14 92.29 92.43 粗蛋白crude protein 48.60 48.69 48.94 48.79 48.70 48.76 粗脂肪crude lipid 12.54 12.56 12.63 12.64 12.61 12.61 灰分ash 9.60 9.76 9.73 9.77 9.82 9.72 注:1. 鱼粉购自CFGINVESTMENTS.A.C.,粗蛋白和粗脂肪的实测值分别为66.71%和9.01%;2. 大豆浓缩蛋白购自山东长润生物有限公司,粗蛋白和粗脂肪的实测值分别为65.86%和0.66%;3. 鱼油购自广州永兴浓缩饲料有限公司,脂肪酸组成为C14:0 2.8%,C15:0 0.26%,C16:0 13.5%,C16:1 n-7 3.5%,C17:0 0.38%,C18:0 3.8%,C18:1 n-9 29.2%,C18:2 n-6 22.6%,C18:3 n-3 4.1%,C20:0 0.71%,C20:1 n-9 1.7%,C20:2 n-7 0.61%,C20:3 n-6 0.17%,C20:4 n-6 1.5%,C20:5 n-3 4.1%,C22:6 n-3 5.2%;4. 维生素预混料为每千克饲料添加VB1 25 mg,VB2 45 mg,VB6 20 mg,VB12 0.1 mg,VC 2.0 g,VD3 5 mg,VE 120 mg,VK3 10 mg,烟酸200 mg,泛酸60 mg,叶酸1.2 mg,生物素32 mg,氯化胆碱2.0 g,肌醇800 mg,乙氧基喹啉150 mg和麸皮14.52 g[16];5. 矿物质预混料为每千克饲料添加以下矿物质:一水合磷酸二氢钙(Ca(H2PO4)2 · H2O)6.0 g,七水硫酸镁(MgSO4 · 7H2O)2.4 g,五水硫酸铜(CuSO4 ·5H2O)20 mg,一水合硫酸亚铁(FeSO4 · H2O)160 mg,一水合硫酸锌(ZnSO4 · H2O)100 mg,一水合硫酸锰(MnSO4 · H2O)120 mg,碘化钾(KI)1.6 mg,氯化钠(NaCl)200 mg,氟化钠(NaF)4 mg,六水氯化钴(CoCl2 · 6H2O)(1%) 100 mg,沸石粉30.90 g[16];6. 亚麻籽油购自广州市承跃贸易有限公司,脂肪酸组成为C16:0 5.6%,C16:1 n-7 0.08%,C17:0 0.06%,C18:0 4.6%,C18:1 n-9 23.3%,C18:2 n-6 16.5%,C18:3 n-3 48.8%,C20:0 0.40%,C20:1 n-9 0.16%,C20:3 n-6 0.15%
Note:1. fish meal:CFGINVESTMENTS.A.C.,Plant Tambo de Mora. The measured values of crude protein and crude lipid were 66.71% and 9.01%,respectively;2. soy protein concentrate,SPC:from Shandong Long-Run Biological Ltd.. The measured values of crude protein and crude lipid were 65.86% and 0.66%,respectively;3. fish oil:from Guangzhou Yongxing Concentrated Feed Ltd.,fatty acid composition:C14:0 2.8%,C15:0 0.26%,C16:0 13.5%,C16:1 n-7 3.5%,C17:0 0.38%,C18:0 3.8%,C18:1 n-9 29.2%,C18:2 n-6 22.6%,C18:3 n-3 4.1%,C20:0 0.71%,C20:1 n-9 1.7%,C20:2 n-7 0.61%,C20:3 n-6 0.17%,C20:4 n-6 1.5%,C20:5 n-3 4.1%,C22:6 n-3 5.2%;4. vitamin premix provided the following per kg of diet:VB1 25 mg,VB2 45 mg,VB6 20 mg,VB12 0.1 mg,VC 2.0 g,VD3 5 mg,VE 120 mg,VK3 10 mg,nicotinic acid 200 mg,pantothenic acid 60 mg,folic acid 1.2 mg,biotin 32 mg,choline chloride 2.0 g,inositol 800 mg,ethoxyquin 150 mg and manna-croup 14.52 g[16];5. mineral premix provided the following per kg of diet:Ca(H2PO4)2 · H2O 6.0 g,MgSO4 · 7H2O 2.4 g,CuSO4 · 5H2O 20 mg,FeSO4 · H2O 160 mg,ZnSO4 · H2O 100 mg,MnSO4 · H2O 120 mg,KI 1.6 mg,NaCl 200 mg,NaF 4 mg,CoCl2 · 6H2O (1%) 100 mg,zeolite powder 30.90 g[16];6. Linseed oil:from Guangzhou Cheng Yue Trade Co.,Ltd.,fatty acid composition:C16:0 5.6%,C16:1 n-7 0.08%,C17:0 0.06%,C18:0 4.6%,C18:1 n-9 23.3%,C18:2 n-6 16.5%,C18:3 n-3 48.8%,C20:0 0.40%,C20:1 n-9 0.16%,C20:3 n-6 0.15% 下载: 导出CSV
下载: 导出CSV
表 2 实验饲料的脂肪酸组成(饲料干物质)
Table 2 Fatty acid composition of experimental diets for juvenile T.ovatus (dry matter basis)
% 脂肪酸
fatty acid组别group 0.13% 0.59% 1.10% 1.60% 2.10% 2.50% C14:0 0.22 0.20 0.19 0.18 0.16 0.15 C15:0 0.01 0.01 0.01 0.01 0.01 0.01 C16:0 2.90 2.60 2.40 2.20 2.00 1.80 C16:1 n-7 0.25 0.23 0.22 0.20 0.18 0.16 C17:0 0.04 0.04 0.40 0.03 0.03 0.30 C18:0 1.60 1.40 1.30 1.20 1.10 0.94 C18:1 n-9 3.90 3.80 3.70 3.60 3.60 3.20 C18:2 n-6 1.70 1.80 1.80 1.90 1.90 1.90 C18:3 n-3 0.13 0.59 1.10 1.60 2.10 2.50 C18:3 n-6 0.01 0.01 0.01 0.01 0.01 0.01 C20:0 0.02 0.04 0.04 0.04 0.04 0.40 C20:1 n-9 0.10 0.09 0.09 0.08 0.08 0.07 C20:2 n-7 0.07 0.07 0.07 0.06 0.06 0.05 C20:3 n-6 0.01 0.01 0.01 0.01 0.01 0.01 C20:4 n-6 0.04 0.03 0.03 0.03 0.03 0.03 C20:5 n-3 0.11 0.11 0.11 0.11 0.11 0.11 C22:6 n-3 0.15 0.15 0.15 0.15 0.15 0.15 ∑饱和脂肪酸saturates 4.79 4.29 4.34 3.66 3.34 3.60 ∑MUFA 4.25 4.12 4.01 3.88 3.86 3.43 ∑PUFA 2.21 2.76 3.27 3.86 4.36 4.75 ∑n-3 0.39 0.85 1.36 1.86 2.36 2.76 ∑n-6 1.75 1.84 1.84 1.94 1.94 1.94 DHA/EPA 1.36 1.36 1.36 1.36 1.36 1.36 ∑n-3/∑n-6 0.22 0.46 0.74 0.96 1.22 1.42 注:MUFA. 单不饱和脂肪酸;PUFA. 多不饱和脂肪酸
Note:MUFA. monounsaturated fatty acid;PUFA. polyunsaturated fatty acid
下载: 导出CSV
表 3 饲料中亚麻酸水平对卵形鲳鲹幼鱼生长性能的影响
Table 3 Effects of LNA level on growth performance of juvenile T.ovatus
组别group 0.13% 0.59% 1.10% 1.60% 2.10% 2.50% 初质量/g initial weight 10.41±0.21 10.34±0.09 10.4±0.10 10.34±0.12 10.35±0.08 10.42±0.05 末质量/g final weight 68.55±1.61ab 69.21±1.33b 71.91±1.84b 68.88±4.52ab 67.56±0.69ab 64.68±1.51a 采食量/g feed intake 73.34±0.45 73.85±4.54 78.69±6.58 73.89±6.93 72.93±1.67 79.3±8.34 增重率/g weight gain 558.07±6.57ab 569.69±11.11b 591.55±24.19b 566.82±50.63ab 552.58±1.84ab 520.71±14.58a 特定生长率/% SGR 3.37±0.02ab 3.40±0.03b 3.45±0.07b 3.39±0.14b 3.35±0.01ab 3.26±0.04a 饲料系数FCR 1.26±0.03a 1.26±0.07a 1.28±0.08a 1.26±0.09a 1.28±0.03a 1.47±0.18b 成活率/% survival rate 71.67±2.89a 81.67±2.89ab 85.00±5.00b 83.33±10.41ab 90.00±5.00b 80.00±8.67ab 脏体比/% VSI 6.31±0.35c 5.95±0.34b 5.81±0.34b 5.67±0.48b 5.65±0.30b 5.24±0.36a 肝体比/% HSI 0.82±0.1ab 0.84±0.09b 0.81±0.08ab 0.78±0.08ab 0.78±0.06ab 0.74±0.09a 腹脂率/% abdominal fat 1.05±0.22d 0.99±0.18cd 0.85±0.15bc 0.74±0.19ab 0.74±0.07ab 0.67±0.16a 肥满度CF 3.32±0.05b 3.35±0.09b 3.32±0.19b 3.20±0.15a 3.16±0.10a 3.17±0.06a 注:同一行数据右上角上标字母不同,表示有显著性差异(P<0.05),后表同此。
Note:Values with different small letter superscripts within the same row indicate significant difference (P<0.05). The same case in the following tables.
下载: 导出CSV
表 4 饲料中亚麻酸水平对卵形鲳鲹幼鱼全鱼体成分的影响
Table 4 Effects of dietary LNA level on whole-body approximate composition of juvenile T.ovatus
组别group 0.13% 0.59% 1.10% 1.60% 2.10% 2.50% w(水分)/% moisture 67.74±1.31 67.28±0.16 68.23±1.27 66.78±0.72 67.23±0.15 68.05±0.92 w(粗蛋白质)/% crude protein 57.05±0.33 56.10±0.79 57.93±1.07 57.03±2.09 57.84±2.79 56.32±0.58 w(粗脂肪)/% lipid 31.35±3.24 29.05±2.10 27.53±2.11 29.94±1.19 29.05±3.75 31.32±2.75 w(灰分)/% ash 12.09±0.86b 11.89±0.10ab 12.01±0.24b 11.10±0.37a 11.83±0.46ab 12.44±0.45b
下载: 导出CSV
表 5 饲料中亚麻酸水平对卵形鲳鲹幼鱼消化酶活性的影响
Table 5 Effects of dietary ARA levels on digestive enzymes of juvenile T.ovatus
组别group 0.13% 0.59% 1.10% 1.60% 2.10% 2.50% 胃蛋白酶/U·g-1 pepsin 12.35±0.42a 17.92±3.69c 16.62±0.69bc 14.84±1.91abc 15.34±1.15abc 13.19±1.93ab 淀粉酶/U·g-1 amylase 2.89±0.05d 1.69±0.31b 1.35±0.03a 2.35±0.05c 1.87±0.18b 1.84±0.13b
下载: 导出CSV
-
[1] NRC. Nutrient requirements of fish and shrimp[M]. Washington: National Academies Press, 2011: 102-105. https://nap.nationalacademies.org/catalog/13039/nutrient-requirements-of-fish-and-shrimp
[2] DAS U N. Essential fatty acids: a review[J]. Curr Pharm Biotechnol, 2006, 7(6): 467-482. doi: 10.2174/138920106779116856
[3] CHEN C Y, SUN B L, LI X X, et al. N-3 essential fatty acids in Nile tilapia, Oreochromis niloticus: quantification of optimum requirement of dietary linolenic acid in juvenile fish[J]. Aquaculture, 2013, 416/417 : 99-104. doi: 10.1016/j.aquaculture.2013.09.003
[4] LI M, CHEN L Q, LI E C, et al. Growth, immune response and resistance to Aeromonas hydrophilaof darkbarbel catfish, Pelteobagrus vachelli(Richardson), fed diets with different linolenic acid levels[J]. Aquac Res, 2013, 46(4): 789-800.
[5] YANG X, TABACHEK J L, DICK T A. Effects of dietary n-3 polyunsaturated fatty acids on lipid and fatty acid composition and haematology of juvenile Arctic charr Salvelinus alpinus (L.)[J]. Fish Physiol Biochem, 1994, 12(5): 409-420. doi: 10.1007/BF00004305
[6] HALVER J E, HARDY R W. Fish nutrition[M]. New York: Academic Press, 2002: 220-222. https://www.sciencedirect.com/book/9780123196521/fish-nutrition
[7] RUYTER B, ROSJO C, EINEN O, et al. Essential fatty acids in Atlantic salmon: time course of changes in fatty acid composition of liver, blood and carcass induced by a diet deficient in n-3 and n-6 fatty acids[J]. Aquac Nutr, 2000, 6(2): 109-117. doi: 10.1046/j.1365-2095.2000.00136.x
[8] SMITH D M, HUNTER B J, ALLAN G L, et al. Essential fatty acids in the diet of silver perch (Bidyanus bidyanus): effect of linolenic and linoleic acid on growth and survival[J]. Aquaculture, 2004, 236(1/2/3/4): 377-390. doi: 10.1016/j.aquaculture.2003.10.021
[9] 张伟涛. 卵形鲳鲹(Trachinotus ovatus)对饲料脂肪利用的研究[D]. 苏州: 苏州大学, 2009: 15.https://www.zhangqiaokeyan.com/academic-degree-domestic_mphd_thesis/020313169519.html [10] 刘楚斌, 陈锤. 卵形鲳鲹的生物学与养殖技术[J]. 齐鲁渔业, 2009(6): 32-33. https://qikan.cqvip.com/Qikan/Article/Detail?id=30670140 [11] 刘兴旺, 许丹, 张海涛, 等. 卵形鲳鲹幼鱼蛋白质需要量的研究[J]. 南方水产科学, 2011, 7(1): 45-49. doi: 10.3969/j.issn.2095-0780.2011.01.007 [12] WANG L N, LIU W B, LU K L, et al. Effects of dietary carbohydrate/lipid ratios on non-specific immune responses, oxidative status and liver histology of juvenile yellow catfish Pelteobagrus fulvidraco[J]. Aquaculture, 2014, 426/427 : 41-48. doi: 10.1016/j.aquaculture.2014.01.022
[13] ZHOU C P, GE X P, NIU J, et al. Effect of dietary carbohydrate levels on growth performance, body composition, intestinal and hepatic enzyme activities, and growth hormone gene expression of juvenile golden pompano, Trachinotus ovatus[J]. Aquaculture, 2015, 437: 390-397. doi: 10.1016/j.aquaculture.2014.12.016
[14] LIN H Z, TAN X H, ZHOU C P, et al. Effect of dietary arginine levels on the growth performance, feed utilization, non-specific immune response and disease resistance of juvenile golden pompano Trachinotus ovatus[J]. Aquaculture, 2015, 437: 382-389. doi: 10.1016/j.aquaculture.2014.12.025
[15] NIU J, DU Q, LIN H Z, et al. Quantitative dietary methionine requirement of juvenile golden pompano Trachinotus ovatus at a constant dietary cystine level[J]. Aquac Nutr, 2013, 19(5): 677-686. doi: 10.1111/anu.12015
[16] 杜强, 林黑着, 牛津, 等. 卵形鲳鲹幼鱼的赖氨酸需求量[J]. 动物营养学报, 2011, 23(10): 1725-1732. doi: 10.3969/j.issn.1006-267x.2011.10.012 [17] 杜强. 卵形鲳鲹赖氨酸和蛋氨酸需求量及饲料中鱼粉替代的研究[D]. 上海: 上海海洋大学, 2012: 20.https://www.doc88.com/p-2824915922316.html [18] SATOH S, POE W E, WILSON R P. Studies on the essential fatty acid requirement of channel catfish, Ictalurus punctatus[J]. Aquaculture, 1989, 79(1/2/3/4): 121-128. https://www.sciencedirect.com/science/article/abs/pii/0044848689904523
[19] TOCHER D R. Fatty acid requirements in ontogeny of marine and freshwater fish[J]. Aquac Res, 2010, 41(5): 717-732. doi: 10.1111/j.1365-2109.2008.02150.x
[20] GLENCROSS B D, SMITH D M, THOMAS M R, et al. Optimising the essential fatty acids in the diet for weight gain of the prawn, Penaeus monodon[J]. Aquaculture, 2002, 204(1/2): 85-99. doi: 10.1016/s0044-8486(01)00644-5
[21] SKALLI A, ROBIN J H. Requirement of n-3 long chain polyunsaturated fatty acids for European sea bass (Dicentrarchus labrax) juveniles: growth and fatty acid composition[J]. Aquaculture, 2004, 240(1/2/3/4): 399-415. doi: 10.1016/j.aquaculture.2004.06.036
[22] 王煜恒, 刘文斌, 王会聪, 等. 饲料中亚油酸和亚麻酸含量对团头鲂幼鱼生长、体成分和消化酶活性的影响[J]. 大连海洋大学学报, 2014, 29(4): 373-380. doi: 10.3969/J.ISSN.2095-1388.2014.04.010 [23] KJAER M A, TODORCEVI Ć M, TORSTENSEN B E, et al. Dietary n-3 HUFA affects mitochondrial fatty acid beta-oxidation capacity and susceptibility to oxidative stress in Atlantic salmon[J]. Lipids, 2008, 43(9): 813-827. doi: 10.1007/s11745-008-3208-z
[24] LEE S M, LEE J H, KIM K D. Effect of dietary essential fatty acids on growth, body composition and blood chemistry of juvenile starry flounder (Platichthys stellatus)[J]. Aquaculture, 2003, 225(1/2/3/4): 269-281. https://www.sciencedirect.com/science/article/abs/pii/S0044848603002953
[25] 关勇. 亚麻籽油对吉富罗非鱼生长、体组成、脂质代谢及抗氧化能力的影响[D]. 重庆: 西南大学, 2014: 20-22.https://d.wanfangdata.com.cn/thesis/Y2573704 [26] 彭祥和, 李法见, 林仕梅, 等. 亚麻籽油对罗非鱼生长性能及肉品质的影响[J]. 动物营养学报, 2014, 27(1): 147-153. https://lib.cqvip.com/Qikan/Article/Detail?id=663502819&from=Qikan_Article_Detail [27] SMITH G I, ATHERTON P, REEDS D N, et al. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: a randomized controlled trial[J]. Am J Clin Nutr, 2011, 93(2): 402-412. doi: 10.3945/ajcn.110.005611
[28] 陈乃松, 肖温温, 梁勤朗, 等. 饲料中脂肪与蛋白质比对大口黑鲈生长、体组成和非特异性免疫的影响[J]. 水产学报, 2012, 36(8): 1270-1280. https://qikan.cqvip.com/Qikan/Article/Detail?id=43076448 [29] 彭墨, 徐玮, 麦康森, 等. 亚麻籽油替代鱼油对大菱鲆幼鱼生长、脂肪酸组成及脂肪沉积的影响[J]. 水产学报, 2014, 38(8): 1131-1139. [30] 麦康森, 陈立侨. 水产动物营养与饲料学[M]. 北京: 中国农业出版社, 2011: 191. [31] FURNÉ M, HIDALGO M C, LÓPEZ A, et al. Digestive enzyme activities in Adriatic sturgeon Acipenser naccarii and rainbow trout Oncorhynchus mykiss. A comparative study[J]. Aquaculture, 2005, 250(1/2): 391-398. https://www.sciencedirect.com/science/article/abs/pii/S0044848605003534
[32] DENG J M, MAI K S, AI Q H, et al. Alternative protein sources in diets for Japanese flounder Paralichthys olivaceus (Temminck and Schlegel): Ⅱ. Effects on nutrient digestibility and digestive enzyme activity[J]. Aquac Res, 2010, 41(6): 861-870. doi: 10.1111/j.1365-2109.2009.02363.x
[33] ZAMBONINO INFANTE J L, CAHU C L. High dietary lipid levels enhance digestive tract maturation and improve Dicentrarchus labrax larval development[J]. J Nutr, 1999, 129(6): 1195-1200. doi: 10.1093/jn/129.6.1195
[34] 王朝明, 罗莉, 张桂众, 等. 饲料脂肪水平对胭脂鱼幼鱼生长、体组成和抗氧化能力的影响[J]. 淡水渔业, 2010, 40(5): 47-53. https://www.doc88.com/p-5754422060738.html [35] 邱小琮, 赵红雪, 王远吉, 等. 牛磺酸对鲤非特异性免疫及抗氧化能力的影响[J]. 上海水产大学学报, 2008, 17(4): 429-434. https://wenku.baidu.com/view/08f9294d3b3567ec102d8ac0?fr=xueshu_top [36] 魏玉婷, 王小洁, 麦康森, 等. 饲料中的维生素E对大菱鲆幼鱼生长、脂肪过氧化及抗氧化能力的影响[J]. 中国海洋大学学报(自然科学版), 2011, 41(6): 45-50. [37] 潘瑜. 亚麻油对鲤生长性能、脂质代谢及抗氧化能力的影响[D]. 重庆: 西南大学, 2013: 25-27.https://apps.wanfangdata.com.cn/thesis/article:Y2309993 [38] 左然涛. 饲料脂肪酸调控大黄鱼免疫力和脂肪酸代谢的初步研究[D]. 青岛: 中国海洋大学, 2013: 102.http://dx.doi.org/10.7666/d.D328923 [39] MOURENTE G, DÍAZ-SALVAGO E, TOCHER D R, et al. Effects of dietary polyunsaturated fatty acid/vitamin E (PUFA/tocopherol ratio on antioxidant defence mechanisms of juvenile gilthead sea bream (Sparus aurata L., Osteichthyes, Sparidae)[J]. Fish Physiol Biochem, 2000, 23(4): 337-351. doi: 10.1023/A:1011128510062
-
期刊类型引用(5)
1. 郭孝辉,王颉,连翠飞. 后熟过程对牡蛎风味及营养物质的影响. 中国调味品. 2023(06): 104-107 .  百度学术
百度学术
2. 林海生,王雯,秦小明,曹文红,章超桦,高加龙,郑惠娜,陈忆宾,伍彬. 不同方法提取马氏珠母贝外套膜胶原蛋白理化性质比较. 广东海洋大学学报. 2022(01): 106-112 . 百度学术
3. 廖津,林海生,伍彬,秦小明,曹文红,高加龙,郑惠娜,章超桦,谭绮晴. 马氏珠母贝闭壳肌活性肽制备及其辅助降血糖功效评价. 南方水产科学. 2022(05): 145-152 . 本站查看
4. 吕佳桐,林海生,秦小明,章超桦,曹文红,高加龙,郑慧娜. 牡蛎及其酶解产物抗皮肤光老化的初步研究. 南方水产科学. 2021(01): 91-100 . 本站查看
5. 王利霞,章超桦,高加龙,郑惠娜,曹文红,秦小明,陈建平. 马氏珠母贝肉提取物对小鼠急性酒精性肝损伤的影响. 广东海洋大学学报. 2021(06): 99-107 . 百度学术
其他类型引用(5)





计量
- 文章访问数: 3296
- HTML全文浏览量: 204
- PDF下载量: 934
- 被引次数: 10
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
