
Spatial analysis of vertical active layer of albacore tuna (Thunnus alalunga) in the South Pacific Ocean
-
摘要:
为了解南太平洋长鳍金枪鱼(Thunnus alalunga)的垂直活动水层分布特征及其适宜的垂直活动水层深度,采用Argo数据重构了研究海域次表层20 ℃和25 ℃等温线深度场,并结合2010年~2012年中水集团南太平洋长鳍金枪鱼延绳钓渔船实际生产统计数据,绘制了20 ℃和25 ℃等温线深度与长鳍金枪鱼单位捕捞努力量渔获量(CPUE)叠加图,分析南太平洋长鳍金枪鱼的垂直活动水层分布特征。结果表明,研究海域20 ℃和25 ℃等温线深度存在明显的季节性变化,且长鳍金枪鱼渔场时空分布随着20 ℃等温线深度的220 m等深线和25 ℃等温线深度的140 m等深线时空变动而季节性南北移动。长鳍金枪鱼中心渔场主要分布于10°S以南、160°E~175°E之间,中心渔场所处海域,其20 ℃等温线深度多在220 m以深,超过250 m的海域CPUE均偏低;25 ℃等温线深度多在140 m以浅,浅于80 m的海域则难以形成中心渔场。采用频次分析与经验累积分布函数(ECDF)相结合的方法,计算出南太平洋长鳍金枪鱼适宜的垂直活动水层深度为88~238 m。文章初步得出了南太平洋长鳍金枪鱼的垂直分布特征及其适宜的垂直活动水层深度,可用于指导延绳钓投钩深度,为中国南太平洋长鳍金枪鱼延绳钓生产作业提供理论参考。
Abstract:In order to understand the vertical distribution characteristics and suitable vertical active layer depth range of albacore tuna in the South Pacific Ocean, we plotted the subsurface isothermal depths at 20 ℃ and 25 ℃ on a spatial overlay map, using Argo data and monthly CPUE (catch per unit effort) from logbook of Chinese albacore tuna long-lines vessels in this area to analyze the vertical distribution characteristics of albacore tuna in the South Pacific Ocean. The analysis shows that the subsurface isothermal depths at 20 ℃ and 25 ℃ had significant seasonal changes; the distribution of central fishing grounds of albacore tuna had significant correlation with the seasonal variation of the isobaths at 220 m of isothermal depths at 20 ℃ and the isobaths at 140 m of isothermal depths at 25 ℃. The central fishing grounds of albacore tuna were distributed to the south of 10°S, between 160°E and 175°E, where the isothermal depths at 20 ℃ was mostly deeper than 220 m, or the isothermal depths at 25 ℃ was mostly shallower than 140 m; if the depth at 20 ℃ was more than 250 m or the depth at 25 ℃ was less than 80 m, the CPUE tended to be low. We used the methods of frequency analysis and the empirical cumulative distribution function (ECDF) to estimate the suitable range of vertical active layer depth of albacore tuna(88~238 m). Thus, the preliminary results on vertical distribution characteristics and the suitable vertical active layer depth range of albacore tuna have been obtained in this study, which can be used as guides for settlement depth of longline hook and provide references for longline production operation.
-
Keywords:
- albacore tuna /
- Argo /
- vertical active layer /
- the South Pacific Ocean
-
长鳍金枪鱼(Thunnus alalunga)是南太平洋延绳钓金枪鱼渔业的重要目标种类,也是中国金枪鱼延绳钓渔船的主捕对象之一[1]。联合国粮农组织(FAO)2010年统计数据显示,南太平洋海域长鳍金枪鱼产量为15.29×104 t,占当年全球长鳍金枪鱼总产量的63%,中国渔船于1998年开始进入该海域进行长鳍金枪鱼延绳钓作业,近年来其产量和作业规模总体呈现出波动增长的趋势[2-3]。相关资源评估报告表明,目前南太平洋长鳍金枪鱼资源状态总体较好,仍具有一定的开发潜力[4]。远洋性鱼类,尤其是金枪鱼,为捕食深海散射层(deep scattering layer,DSL)生物,都具有“昼潜夜浮”的特性[5]。ZAGAGLIA等[6]认为这种垂直方向上的昼夜移动,会使得金枪鱼垂直活动水层与次表层水温的关系更为密切。DOMOKOS等[7]通过在美属萨摩亚群岛附近海域进行成年长鳍金枪鱼标志放流研究发现,长鳍金枪鱼在18~30 ℃水层均有分布,其中65%~70%的时间在20~25 ℃水层活动。延绳钓调查也得出相似结论,林显鹏等[8]在南太平洋所罗门群岛以及杨嘉樑等[9]在东太平洋库克群岛附近海域进行延绳钓调查得出,长鳍金枪鱼适宜栖息水温分别为20~26 ℃以及20~25 ℃。上述研究结果表明,水温垂直结构对长鳍金枪鱼垂直活动水层的分布有直接影响,同时也是长鳍金枪鱼渔场形成过程中极为重要的因素之一。因此,分析南太平洋长鳍金枪鱼的垂直活动水层分布特征,了解其适宜的垂直活动水层深度范围十分有必要。以往对长鳍金枪鱼垂直活动水层的研究主要依靠标志放流和延绳钓调查,但标志放流和延绳钓调查的区域跨度小且在时间上不连续,无法大面积、长时间序列的监测海洋次表层环境变化对长鳍金枪鱼资源分布的影响。该研究根据前人标志放流和延绳钓调查所得结论,采用Argo数据重构了南太平洋次表层20 ℃和25 ℃等温线深度场,结合2010年~2012年中水集团长鳍金枪鱼延绳钓生产统计数据,分析了南太平洋长鳍金枪鱼的垂直分布特征,并采用数值方法计算其适宜的垂直活动水层深度,以期为南太平洋延绳钓长鳍金枪鱼捕捞作业、资源保护和管理提供参考。
1. 材料与方法
1.1 数据
1.1.1 渔业数据
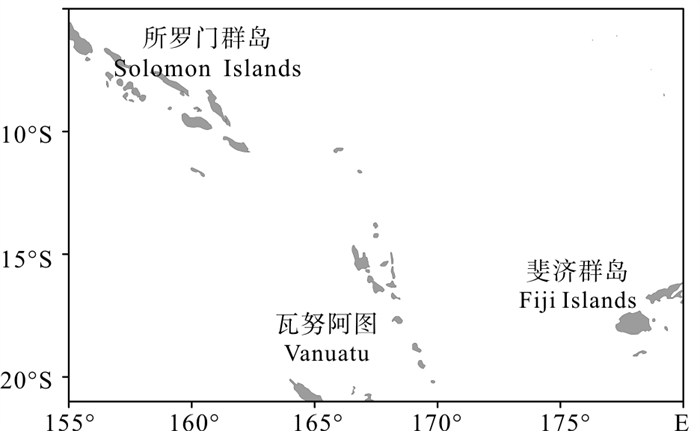
渔业数据来源于中水集团南太平洋长鳍金枪鱼延绳钓船队的21艘延绳钓渔船渔捞日志,数据包括作业时间、作业位置、渔获量、渔获尾数、下钩数等信息,时间为2010年~2012年,空间范围为5°S~21°S、155°E~180°E,具体研究区域见图 1。在实际分析统计中,以1°×1°为空间统计单元,并按月计算各1°×1°渔区网格内的单位捕捞努力量渔获量(catch per unit effort,CPUE)。
![]() 图 1 南太平洋长鳍金枪鱼渔场范围示意图Figure 1. Map of fishing ground of albacore tuna in the South Pacific Ocean
图 1 南太平洋长鳍金枪鱼渔场范围示意图Figure 1. Map of fishing ground of albacore tuna in the South Pacific OceanCPUE计算公式如下[10]:
$$ \operatorname{CPUE}_{(i, j)}=\frac{N_{f is h(i, j)} \times 1\;000}{N_{h o o k(i, j)}} $$ (1) 式中CPUE(i, j)、Nfish(i, j)、Nhook(i, j)分别是第i个经度、第j个纬度所处渔区网格内的CPUE(尾·千钩-1)、渔获尾数和实际下钩数。随后计算2010年~2012年各月各渔区网格内所有CPUE值的四分位数(Q1~Q3),并将所有大于Q3(第3个四分位点)的CPUE称为高值CPUE,其所属渔区定义为南太平洋长鳍金枪鱼的中心渔场[11]。2010年~2012年按月统计的1°×1°渔区网格内CPUE样本数共1 939个,其中高值CPUE样本485个,CPUE的四分位数划分如下:Q1=7.27尾·千钩-1,Q2=11.23尾·千钩-1,Q3=15.78尾·千钩-1。
1.1.2 Argo数据
综合前人标志放流和延绳钓调查所得结论,选取20 ℃和25 ℃分别作为南太平洋长鳍金枪鱼适宜栖息水温的下界和上界,用于分析其垂直活动水层分布特征及适宜的垂直活动水层深度范围。2010年~2012年南太平洋次表层20 ℃和25 ℃等温线深度数据来源于中国Argo实时数据中心(http://www.argo.org.cn/)。文章采用杨胜龙等[12]的方法将离散分布的Argo剖面深度数据,等距插值到规则等温线层上,垂直等距间隔为1 m。将2010年~2012年所有Argo数据按月进行分组,分别提取20 ℃和25 ℃等温面上离散分布的深度值,并采用Kriging插值方法将其插值到1°×1°网格节点上。
1.2 方法
1.2.1 空间分析
将2010年~2012年南太平洋次表层20 ℃和25 ℃等温线深度数据与长鳍金枪鱼CPUE数据按月进行空间匹配,采用空间分析软件Arcgis绘制20 ℃和25 ℃等温线深度与CPUE空间叠加分布图。根据南半球的季节划分(1月~3月为夏季、4月~6月为秋季、7月~9月为冬季、10月~12月为春季),分析不同季节下长鳍金枪鱼CPUE与20 ℃和25 ℃等温线深度的时空分布特征及其动态变化规律。
1.2.2 垂直活动水层深度计算
采用频次分析和经验累积分布函数(empirical cumulative distribution function,ECDF)相结合的方法,计算南太平洋长鳍金枪鱼适宜的垂直活动水层深度范围[13]。1)计算南太平洋长鳍金枪鱼中心渔场20 ℃和25 ℃等温线深度的平均值、均方差及适宜的垂直活动水层深度区间(平均值±均方差);2)计算高值CPUE与20 ℃和25 ℃等温线深度的经验累积分布函数及适宜的垂直活动水层深度区间[最大D(t)处深度值±均方差];取两者交集,即为长鳍金枪鱼适宜的垂直活动水层深度。
ECDF计算方法如下[14]:
$$ f(t)=\frac{1}{n} \sum\limits_{i=1}^n l\left(x_i\right) \quad l\left(x_i\right)= \begin{cases}1, & x_i \leq t \\ 0, & x_i>t\end{cases} $$ (2) $$ g(t)=\frac{1}{n} \sum\limits_{i=1}^n \frac{y_i}{\bar{y}} $$ (3) $$ D(t)=|f(t)-g(t)| $$ (4) 式中f(t)为深度值的经验累积分布函数,g(t)为高值CPUE与深度的加权经验累积分布函数,l(t)为分段函数,D(t)为t时刻处f(t)与g(t)差的绝对值。n为高值CPUE样本个数;t是以1 m为间距从低到高排列的深度范围序列;xi为第i个样本对应的深度值;yi为第i个样本对应的CPUE;y为所有高值CPUE样本的平均值;根据给定的显著性水平α=0.05,采用双样本K-S(Kolmogorov-Smirnov)检验方法分别检验高值CPUE与20 ℃和25 ℃等温线深度间的关系。
2. 结果与分析
2.1 长鳍金枪鱼垂直活动水层空间分析
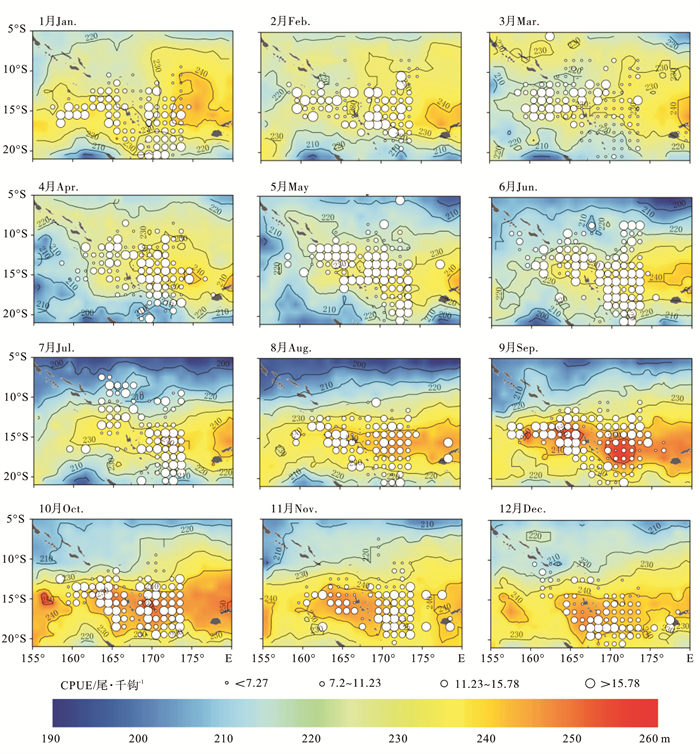
2010年~2012年南太平洋长鳍金枪鱼CPUE与20 ℃等温线深度的月平均空间分布见图 2。20 ℃等温线深度大致以15°S为中心,呈南北对称分布,且存在明显的季节性变化,长鳍金枪鱼CPUE的时空分布与20 ℃等温线深度的220 m等深线时空变动特征具有明显的相关性。1月,研究海域内20 ℃等温线深度基本在220 m以上,深度为220~230 m区域占整个研究海域面积的比例较大,高值CPUE主要分布于12°S~18°S,深度为220~240 m的海域,瓦努阿图东北部,20 ℃等温线深度小于220 m的海域,也有少量高值CPUE分布;2月~3月,研究海域南部的20 ℃等温线深度略有下降,中心渔场开始北移,渔场分布的纬度范围变窄,瓦努阿图东北部渔场消失,而深度为220~230 m的区域面积则进一步扩张,约占整个研究海域面积的一半;秋季(4月~6月),随着作业渔船数量的增加,高值CPUE的个数及分布范围均有所拓展,220 m等深线较1月~3月有明显的向南移动趋势,20 ℃等温线深度为220~230 m的海域面积开始逐步缩小,中心渔场所在区域20 ℃等温线深度则逐步加深,20 ℃等温线深度超过230 m的中心渔场数量显著增加,随着220 m等深线的南移,研究区域北部20 ℃水层的深度开始逐步变浅,而研究海域南部20 ℃等温线深度则开始逐步加深;从冬季(7月~9月)开始,10°S以北海域20 ℃等温线深度已基本全部处于220 m以下,此处区域全年均少有渔场出现,中心渔场位置开始逐步向南移动,深度为220~240 m的区域面积进一步缩小,渔场分布的纬度范围开始重新收窄;春季(10~12月),中心渔场继续向南移动,中心渔场所处海域的20 ℃等温线深度则继续加深,到12月,南太平洋长鳍金枪鱼中心渔场已完全分布于15°S以南,中心渔场所在海域的20 ℃等温线深度下界也随之拓展到250 m。
![]() 图 2 各月平均20 ℃等温线深度与平均CPUE空间叠加图Figure 2. Overlay map of average isothermal depths at 20 ℃ and average CPUE in each month
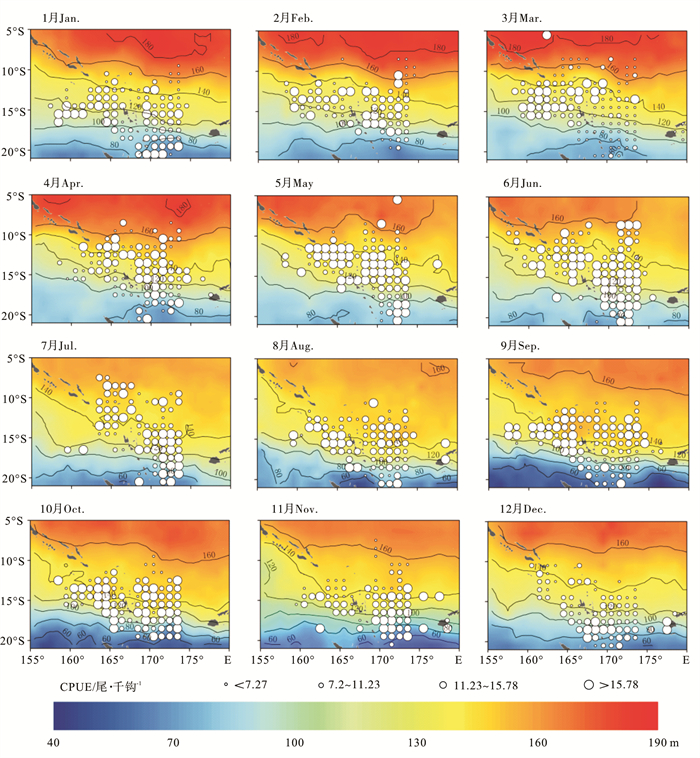
图 2 各月平均20 ℃等温线深度与平均CPUE空间叠加图Figure 2. Overlay map of average isothermal depths at 20 ℃ and average CPUE in each month2010年~2012年南太平洋长鳍金枪鱼CPUE与25 ℃等温线深度的月平均空间分布见图 3。25 ℃等温线深度大致呈由北向南逐步递减的纬向带状分布。25 ℃等温线深度也存在明显的季节性变化,长鳍金枪鱼中心渔场的时空分布与25 ℃等温线深度的140 m等深线季节性变化趋势具有明显的相关性。1月,长鳍金枪鱼高值CPUE主要分布于25 ℃等温线深度为100~140 m的海域,瓦努阿图东北部,25 ℃等温线水深在80 m以下的海域,也有少量高值CPUE分布;2月~3月,中心渔场逐步北移,中心渔场对应的25 ℃等温线深度范围变窄,瓦努阿图东北部渔场消失;秋季(4月~6月),随着研究海域内捕捞努力量的增加,中心渔场的数量和空间分布范围均有所拓展,140 m等深线开始逐步南移,水深为140~160 m的海域面积不断扩张,中心渔场对应的25 ℃等温线深度范围拓宽至80~160 m;冬季(7月~9月),研究海域北部25 ℃等温线深度开始加深,140~160 m水深海域面积达到全年最大值,140 m等深线已完全分布于15°S以南,中心渔场对应的25 ℃等温线深度也随之变浅至80~150 m,研究海域南部25 ℃等温线深度开始变浅,中心渔场也随着140 m等深线的变动而向南转移,此处海域等深线分布密集,水层结构较为复杂,同时也是长鳍金枪鱼高值CPUE密集分布的海域;从春季(10月~12月)开始,140 m等深线开始北移,研究海域北部25 ℃等温线深度逐渐变浅,140~160 m水深的海域面积逐渐缩小,而在研究海域南部,25 ℃等温线深度则开始逐步加深,随着研究海域南部25 ℃等温线深度的增加,长鳍金枪鱼中心渔场的分布也随之不断南移,到12月,长鳍金枪鱼中心渔场已完全分布于15°S以南,中心渔场对应的25 ℃等温线深度范围缩小至80~120 m。
![]() 图 3 各月平均25℃等温线深度与平均CPUE空间叠加图Figure 3. Overlay map of average isothermal depths at 25 ℃ and average CPUE in each month
图 3 各月平均25℃等温线深度与平均CPUE空间叠加图Figure 3. Overlay map of average isothermal depths at 25 ℃ and average CPUE in each month2.2 长鳍金枪鱼适宜垂直活动水层深度计算
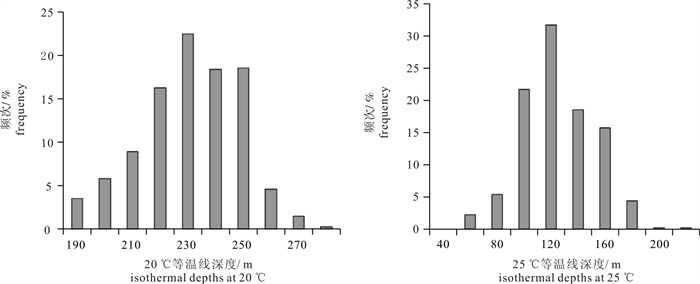
2010年~2012年南太平洋长鳍金枪鱼中心渔场所在区域,20 ℃等温线深度在181~272 m,25 ℃等温线深度在41~201 m均有分布。长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度的频次分布直方图(图 4)呈现出明显的偏正态分布趋势,20 ℃等温线深度平均值为227 m、均方差为18,25 ℃等温线深度平均值为115 m,均方差为27。采用频次分析方法计算得出,71%的长鳍金枪鱼中心渔场趋向于集中分布在20 ℃等温线深度为209~245 m(227±18)的海域,70%的长鳍金枪鱼中心渔场趋向于集中分布在25 ℃等温线深度为88~142 m(115±27)的海域。
![]() 图 4 长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度频次分布直方图Figure 4. Frequency distribution histogram of high CPUE and isothermal depths at 20 ℃ and 25 ℃
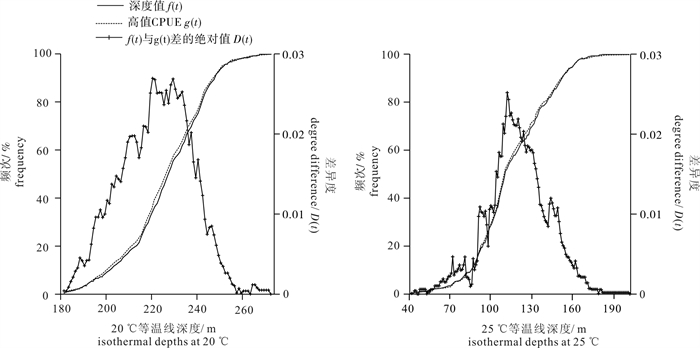
图 4 长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度频次分布直方图Figure 4. Frequency distribution histogram of high CPUE and isothermal depths at 20 ℃ and 25 ℃采用K-S检验方法对长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度之间的关系进行检验,样本数为485,在给定显著性水平α=0.05下,得出D0.05=0.061 7。ECDF分析结果见图 5。20 ℃和25 ℃等温线深度对应的最大D(t)值分别为0.026 9和0.025 1,均远小于D0.05,表明样本间服从同一分布,长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度间有密切关系。20 ℃和25 ℃等温线深度对应的最大D(t)值分别出现在220 m和111 m,与中心渔场20 ℃和25 ℃等温线深度的均值稍有差异(分别为227 m和115 m)。采用ECDF方法得出长鳍金枪鱼中心渔场20 ℃和25 ℃等温线深度分别为202~238 m(220±18)、84~138 m(111±27)。综合频次分析和ECDF分析的结果,取两者交集,得出南太平洋长鳍金枪鱼适宜的垂直活动水层深度为88~238 m。
![]() 图 5 长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度值的经验累积分布函数Figure 5. ECDF for high CPUE of albacore tuna and isothermal depths at 20 ℃ and 25 ℃
图 5 长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度值的经验累积分布函数Figure 5. ECDF for high CPUE of albacore tuna and isothermal depths at 20 ℃ and 25 ℃3. 讨论
3.1 长鳍金枪鱼渔场分布与20 ℃、25 ℃等温线深度的关系
以往对南太平洋长鳍金枪鱼渔场环境的研究,采用的多是遥感获取的海表面环境因子[15-19]。但标志放流和延绳钓调查结果表明,长鳍金枪鱼多分布于80 m以深的水层,10 m以浅的近表层很少有长鳍金枪鱼活动[20-21]。因此,相较于表层环境因子,采用次表层环境变量分析长鳍金枪鱼的栖息环境更为适合。长鳍金枪鱼耳石研究发现,长鳍金枪鱼经常出没于海洋环境变化较大的水层,如温跃层和混合层[22]。笔者在研究过程中也发现,15°S以南,长鳍金枪鱼高值CPUE密集分布的海域,其25 ℃等温线深度的等深线分布也十分密集,推测此处可能有混合层存在(图 3)。以往的研究中,常用20 ℃等温线深度来表示太平洋海域温跃层的深度,表明在太平洋海域次表层20 ℃等温线分布与温跃层的分布十分相似[23-24]。DOMOKOS等[6]指出,长鳍金枪鱼白天下潜至温跃层以下觅食DSL生物,夜晚则游回混合层上方,混合层及温跃层的深度直接影响到长鳍金枪鱼垂直活动水层的深度。对太平洋海域DSL生物的调查研究发现,DSL生物白天主要分布于温跃层以下,夜晚则分布于温跃层以上[25]。据此推断,20 ℃等温线可能通过影响DSL生物的垂直分布,进而影响长鳍金枪鱼垂直活动水层的空间分布。
K-S检验结果表明,长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度之间有密切关系。长鳍金枪鱼中心渔场所处海域,其20 ℃等温线深度多在220 m以深,超过250 m则少有中心渔场出现;25 ℃等温线深度多在140 m以浅,浅于80 m则少有中心渔场出现。长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度的时空分布均呈现出明显的季节性变化特征,20 ℃和25 ℃等温线深度的这种季节性变化进而导致长鳍金枪鱼垂直活动水层深度的季节性变化。长鳍金枪鱼中心渔场的时空分布与20 ℃等温线深度的220 m等深线和25 ℃等温线深度的140 m等深线的时空变动具有显著的相关性,进一步表明水温垂直结构对长鳍金枪鱼的垂直活动水层深度及渔场分布有直接影响。
3.2 长鳍金枪鱼适宜垂直活动水层深度分析
综合频次分析和ECDF方法计算得出,南太平洋长鳍金枪鱼适宜的垂直活动水层深度为88~238 m。所得结果在中西太平洋长鳍金枪鱼垂直分布水层(80~380 m)之内[19],与临近的所罗门群岛附近海域长鳍金枪鱼活动较为频繁的水层(130~190 m)相符[7],与相似纬度的东太平洋海域成年长鳍金枪鱼钓获深度(102.3~236.1 m)基本相同[26],与东南太平洋长鳍金枪鱼延绳钓钓获深度(74.33~296.50 m)相似[27]。文章初步得出了南太平洋长鳍金枪鱼适宜的垂直活动水层深度范围,在实际的长鳍金枪鱼延绳钓生产作业中,应尽可能使延绳钓钓钩沉降至88~238 m,从而有效节省饵料投放、提高长鳍金枪鱼的生产效率。
4. 展望
文章从次表层水温垂直结构的角度研究了南太平洋长鳍金枪鱼的垂直活动水层分布特征及其适宜的垂直活动水层深度范围,事实上,长鳍金枪鱼的垂直分布还受到海流、溶解氧、海水盐度、饵料生物分布等其他诸多环境因素的共同影响[28-30]。Argo数据可以实时获取次表层水温、水深、盐度以及溶解氧等信息,极大地拓展了远洋渔业资源和渔场学研究中海洋环境因子的选择范围,在远洋渔业研究中有着良好的应用前景。综合多种环境因子,恰当地选择神经网络、支持向量机、栖息地适应性指数等预报模型,准确、全面地把握渔场时空分布,是今后中国长鳍金枪鱼渔业研究的一个重要方向。
-
![]()
图 1 南太平洋长鳍金枪鱼渔场范围示意图
Figure 1. Map of fishing ground of albacore tuna in the South Pacific Ocean
![]()
图 2 各月平均20 ℃等温线深度与平均CPUE空间叠加图
Figure 2. Overlay map of average isothermal depths at 20 ℃ and average CPUE in each month
![]()
图 3 各月平均25℃等温线深度与平均CPUE空间叠加图
Figure 3. Overlay map of average isothermal depths at 25 ℃ and average CPUE in each month
![]()
图 4 长鳍金枪鱼高值CPUE与20 ℃和25 ℃等温线深度频次分布直方图
Figure 4. Frequency distribution histogram of high CPUE and isothermal depths at 20 ℃ and 25 ℃
-
[1] 苗振清, 黄锡昌. 远洋金枪鱼渔业[M]. 北京: 科学技术文献出版社, 2003: 28-34. [2] FAO. Global tuna catches by stock 1950-2010[DB/OL]. http://www.fao.org/figis/servlet/TabSelector.
[3] 唐衍力, 徐湛. 中西太平洋长鳍金枪鱼(Thunnus alalunga)延绳钓渔场分布的时空变化[J]. 渔业科学进展, 2015, 36(3): 22-29. doi: 10.11758/yykxjz.20150304 [4] 吕泽华. 南太平洋长鳍金枪鱼可持续发展的初步研究[D]. 上海: 上海海洋大学, 2015, : 6-9. [5] SCHAEFER K M, FULLER D W, GABRIEL A. Movements, behavior, and habitat utilization of yellowfin tuna (Thunnus albacares) in waters surrounding the Revillagigedo Islands Archipelago Biosphere Reserve, Mexico[J]. Fish Oceanogr, 2014, 23(1): 65-82. doi: 10.1111/fog.12047
[6] ZAGAGLIA C R, LORENZZETTI J A, STECH J L. Remote sensing data and longline catches of yellow-fin tuna (Thunnus albacares) in the equatorial Atlantic[J]. Remot Sens Environ, 2004, 93(1/2): 267-281. https://www.sciencedirect.com/science/article/pii/S0034425704002354
[7] DOMOKOS R, SEKI M P, POLOVINA J J, et al. Oceanographic investigation of the American Samoa albacore (Thunnus alalunga) habitat and longline fishing grounds[J]. Fish Oceanogr, 2007, 16(6): 555-572. doi: 10.1111/j.1365-2419.2007.00451.x
[8] 林显鹏, 郭爱, 张洪亮, 等. 所罗门群岛海域长鳍金枪鱼的垂直分布与环境因子的关系[J]. 浙江海洋学院学报(自然科学版), 2011, 30(4): 303-306. doi: 10.3969/j.issn.1008-830X.2011.04.005 [9] 杨嘉樑, 黄洪亮, 宋利明, 等. 基于分位数回归的库克群岛海域长鳍金枪鱼栖息环境综合指数[J]. 中国水产科学, 2014, 21(4): 832-851. doi: 10.3724/SP.J.1118.2014.00832 [10] LEE P F, CHEN I C, TZENG W N. Spatial and temporal distribution patterns of bigeye tuna (Thunnus obesus) in the Indian Ocean[J]. Zool Stud, 2005, 44(2): 260-270. https://www.researchgate.net/publication/267198305_Zoological_Studies_442_Spatial_and_Temporal_Distribution_Patterns_of_Bigeye_Tuna_Thunnus_obesus_in_the_Indian_Ocean
[11] 陈雪忠, 杨胜龙, 张禹, 等. 热带印度洋大眼金枪鱼垂直分布空间分析[J]. 中国水产科学, 2013, 20(3): 660-671. doi: 10.3724/SP.J.1118.2013.00660 [12] 杨胜龙, 化成君, 靳少非, 等. 热带印度洋黄鳍金枪鱼水平-垂直分布空间分析[J]. 海洋与湖沼, 2014, 45(2): 281-290. [13] 杨胜龙, 张忭忭, 靳少非, 等. 中西太平洋延绳钓黄鳍金枪鱼渔场时空分布与温跃层关系[J]. 海洋学报, 2015, 37(6): 78-87. doi: 10.3969/j.issn.0253-4193.2015.06.008 [14] ZAINUDDIN M, SAITOH K, SAITOH S I. Albacore (Thunnus alalunga) fishing ground in relation to oceanographic conditions in the western North Pacific Ocean using remotely sensed satellite data[J]. Fish Oceanogr, 2008, 17(2): 61-73. doi: 10.1111/j.1365-2419.2008.00461.x
[15] 樊伟, 张晶, 周为峰. 南太平洋长鳍金枪鱼延绳钓渔场与海水表层温度的关系分析[J]. 大连水产学院学报, 2007, 22(5): 366-371. doi: 10.3969/j.issn.1000-9957.2007.05.010 [16] 范江涛, 陈新军, 钱卫国, 等. 瓦努阿图周边海域长鳍金枪鱼渔场分布及其与表温关系[J]. 海洋湖沼通报, 2011(1): 71-78. doi: 10.3969/j.issn.1003-6482.2011.01.010 [17] 刘洪生, 蒋汉凌, 戴小杰. 中西太平洋长鳍金枪鱼渔场与海温的关系[J]. 上海海洋大学学报, 2014, 23(4): 602-607. https://www.nstl.gov.cn/paper_detail.html?id=4b1537683b1099af91eacfbe84afee2c [18] 范永超, 陈新军, 汪金涛. 基于多因子栖息地指数模型的南太平洋长鳍金枪鱼渔场预报[J]. 海洋湖沼通报, 2015(2): 36-44. [19] 闫敏, 张衡, 伍玉梅, 等. 2011年南太平洋长鳍金枪鱼渔场时空分布及其与主要海洋环境变化特征[J]. 渔业信息与战略, 2015, 30(2): 119-127. doi: 10.13233/j.cnki.fishis.2015.02.007 [20] SAITO S. Studies on Fishing of Albacore, Thunnus Alalunga (Bonnaterre) by experimental deep-sea tuna long-line[J]. Mem Fac Fish Hokkaido Univ, 1973, 21(2): 107-184. http://agris.fao.org/agris-search/search.do?recordID=AV20120109257
[21] WILLIAMS A J, ALLAIN V, NICOL S J, et al. Vertical behavior and diet of albacore tuna (Thunnus alalunga) vary with latitude in the South Pacific Ocean[J]. Deep-Sea Res Ⅱ, 2015, 113: 154-169. https://www.sciencedirect.com/science/article/pii/S0967064514000757
[22] MACDONALD J I, FARLEY J H, CLEAR N P, et al. Insights into mixing and movement of South Pacific albacore Thunnus alalunga derived from trace elements in otoliths[J]. Fish Res, 2013, 148(6): 56-63. https://www.sciencedirect.com/science/article/pii/S0165783613001859
[23] 赵永平, 陈永利, 王凡, 等. 热带太平洋海洋混合层水体振荡与ENSO循环[J]. 中国科学D辑, 2007, 37(8): 1120-1133. doi: 10.3321/j.issn:1006-9267.2007.08.016 [24] 容新尧, 杨修群. 全球海洋大气耦合环流模式中的ENSO特征对气候背景态改变的敏感性[J]. 气象学报, 2003, 61(1): 52-65. https://www.cnki.com.cn/Article/CJFDTotal-QXXB200301005.htm [25] HAZEN E L, JOHNSTON D W. Meridional patterns in the deep scattering layers and top predator distribution in the central equatorial Pacific[J]. Fish Oceanogr, 2010, 19(6): 427-433. https://www.researchgate.net/publication/227723620_Meridional_patterns_in_the_deep_scattering_layers_and_top_predator_distribution_in_the_central_equatorial_Pacific
[26] 翟天晨, 戴小杰, 朱江峰. 东太平洋长鳍金枪鱼个体大小与钓获深度的关系[J]. 海洋渔业, 2015, 37(1): 10-16. https://www.cnki.com.cn/Article/CJFDTotal-HTYY201501002.htm [27] 张艳波, 戴小杰, 朱江峰, 等. 东南太平洋金枪鱼延绳钓主要渔获种类垂直分布[J]. 应用生态学报, 2015, 26(3): 912-918. https://www.nstl.gov.cn/paper_detail.html?id=4ff1e36253580076e4115310c4ed3c83 [28] 陈雪忠, 樊伟, 崔雪森, 等. 基于随机森林的印度洋长鳍金枪鱼渔场预报[J]. 海洋学报, 2013, 35(1): 158-164. doi: 10.3969/j.issn.0253-4193.2013.01.018 [29] 任中华, 陈新军, 方学燕. 基于栖息地指数的东太平洋长鳍金枪鱼渔场分析[J]. 海洋渔业, 2014, 36(5): 385-395. doi: 10.3969/j.issn.1004-2490.2014.05.001 [30] 闫敏, 张衡, 樊伟, 等. 南太平洋长鳍金枪鱼渔场CPUE时空分布及其与关键海洋环境因子的关系[J]. 生态学杂志, 2015, 34(11): 3191-3197. https://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_stxzz201511029 -
期刊类型引用(15)
1. 刘雨青,吴世腾,宋利明,伏州,张子宸. 大滚筒智能钓机出绳速度控制及其对长鳍金枪鱼捕捞效果的影响. 上海海洋大学学报. 2025(02): 282-294 .  百度学术
百度学术
2. 郑淳文,刘力文,郭绍健,孟晓宇,周成. 海洋中尺度涡对南太平洋长鳍金枪鱼延绳钓渔获量的影响. 上海海洋大学学报. 2025(02): 413-421 . 百度学术
3. 王志华,杨晓明,田思泉. 南太平洋长鳍金枪鱼资源不同尺度的空间格局特征. 南方水产科学. 2023(02): 31-41 . 本站查看
4. 许回,宋利明,沈介然,李玉伟,张敏. 基于GAM的库克群岛海域长鳍金枪鱼CPUE时空分布与海洋环境的关系. 海洋通报. 2023(04): 444-455 . 百度学术
5. XU Hui,SONG Liming,ZHANG Tianjiao,LI Yuwei,SHEN Jieran,ZHANG Min,LI Kangdi. Effects of Different Spatial Resolutions on Prediction Accuracy of Thunnus alalunga Fishing Ground in Waters Near the Cook Islands Based on Long Short-Term Memory(LSTM) Neural Network Model. Journal of Ocean University of China. 2023(05): 1427-1438 . 必应学术
6. 宋利明,任士雨,洪依然,张天蛟,隋恒寿,李彬,张敏. 大西洋热带海域长鳍金枪鱼渔场预报模型的比较. 海洋与湖沼. 2022(02): 496-504 . 百度学术
7. 袁红春,高子玥,张天蛟. 基于改进的XGBoost模型预测南太平洋长鳍金枪鱼资源丰度. 海洋湖沼通报. 2022(02): 112-120 . 百度学术
8. 周胜杰,杨蕊,于刚,吴洽儿,马振华. 青干金枪鱼和小头鲔循环水养殖生长研究. 水产科学. 2021(03): 339-346 . 百度学术
9. 程懿麒,张俊波,汪金涛,雷林. 基于神经网络的印度洋长鳍金枪鱼(Thunnus alalunga)时空分布与海洋环境关系研究. 海洋与湖沼. 2021(04): 960-970+1-6 . 百度学术
10. 谢笑艳,汪金涛,陈新军,陈丕茂. 南印度洋长鳍金枪鱼渔获率与水深温度关系研究. 南方水产科学. 2021(05): 86-92 . 本站查看
11. 张嘉容,杨晓明,戴小杰,邹莉瑾. 南太平洋长鳍金枪鱼延绳钓渔获率与环境因子的关系研究. 南方水产科学. 2020(01): 69-77 . 本站查看
12. 张亚男,官文江,李阳东. 印度洋长鳍金枪鱼栖息地指数模型的构建与验证. 上海海洋大学学报. 2020(02): 268-279 . 百度学术
13. 张嘉容,杨晓明,田思泉. 基于最大熵模型的南太平洋长鳍金枪鱼栖息地预测. 中国水产科学. 2020(10): 1222-1233 . 百度学术
14. 侯娟,周为峰,樊伟,张衡. 基于集成学习的南太平洋长鳍金枪鱼渔场预报模型研究. 南方水产科学. 2020(05): 42-50 . 本站查看
15. 江建军,许柳雄,朱国平,郭志勋,朱伟俊. 利用鳍条研究北太平洋长鳍金枪鱼的年龄与生长. 水产学报. 2019(04): 917-927 . 百度学术
其他类型引用(14)
 下载:
下载:
计量
- 文章访问数: 3310
- HTML全文浏览量: 157
- PDF下载量: 1106
- 被引次数: 29
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
