
Spatial pattern characteristics of albacore tuna resources at different spatial scales in South Pacific
-
摘要: 长鳍金枪鱼 (Thunnus alalunga) 是延绳钓渔业主要捕捞目标物种,占金枪鱼年渔获量的1/3。南太平洋的长鳍金枪鱼资源丰富,探究其渔业资源的空间格局特征,分析是否存在空间尺度差异,对其渔业资源的合理利用和保护具有重要意义。根据2015—2019年中国大陆延绳钓渔捞日志数据,按季度以3种空间尺度进行聚合 (1°、2°和5°),以及中西太平洋渔业委员会提供的5°网格数据,分别计算其空间格局特征,以比较不同空间尺度下资源分布格局的差异。结果表明:1) 南太平洋长鳍金枪鱼资源空间格局方向特征的椭圆扁率大,所有尺度的空间格局均呈现显著的东西向分布,且资源的空间格局特征 (资源的分布范围、扁率、分布方位角等) 呈明显的季节性波动特征;2) 同源的3种不同尺度数据反映的资源空间格局差异性较小,而不同源数据反映的则差异性较大;3) 在同尺度5°网格数据中,相较于中西太平洋委员会数据,渔捞日志数据中空间格局的资源中心更偏东南,偏东约10°经度;4) 采用较大尺度聚合的数据反映出的资源空间聚集特征更强,且不同尺度数据得到的资源空间热点区域有一定差异。Abstract: Albacore tuna (Thunnus alalunga) is the main target species of longline fishery, accounting for 1/3 of the annual catch of tuna. Since the fishery resources of T. alalunga are rich in South Pacific, to explore the spatial pattern characteristics of its fishery resources in that sea area, and to analyze whether there are differences in the spatial scale are important for the rational utilization and protection of its fishery resources in South Pacific. We aggregated the longline fishing logs of the Chinese mainland from 2015 to 2019 at three spatial scales (1°, 2° and 5°) by season, and calculated their spatial pattern characteristics based on the 5° grid data from Western and Central Pacific Fisheries Commission (WCPFC). The results show that: 1) The spatial pattern of T. alalunga resources in the South Pacific was characterized by great ellipticity, and there was a significant eastwest distribution in the direction of the spatial pattern at all scales. The spatial pattern characteristics of resources (Distribution range, flatness, azimuth, etc.) had similar and obvious seasonal fluctuation characteristics. 2) The spatial pattern of resources obtained from three different scales of the same origin had a small difference, while that from different sources had a great difference. 3) For the same scale of 5° grid data, the resource center of the spatial pattern in China was more southeast and about 10° east in longitude than that of the data from the WCPFC. 4) Data aggregated at larger scales reflect stronger spatial aggregation characteristics of resources. Besides, the hot spots of spatial pattern obtained from different scale data of same origin were different to some extent.
-
Keywords:
- Thunnus alalunga /
- Spatial pattern /
- Standard deviation ellipse /
- Hot spot analysis /
- South Pacific
-
花斑蛇鲻(Saurida undosquamis),属灯笼鱼目、狗母鱼科、蛇鲻属,是一种分布于东印度洋和西北太平洋近岸[1]暖水性底层鱼类,在中国主要分布于南海和台湾海峡,水深30~120 m的沙质海区,常常与多齿蛇鲻(S. tumbil)混栖而一同被捕获[2]。蛇鲻类鱼肉鲜嫩,多做鱼丸食用,是南海区重要的底层经济鱼类[3],1997—1999年南海北部底拖网调查数据显示多齿蛇鲻和花斑蛇鲻渔获量居底层经济鱼类的第1位和第3位[4-6]。在2009年南海鱼类的捕捞产量调查中,蛇鲻类的产量达到5.6×104 t,位居第9[7]。但近年来由于近海过度捕捞和水域污染等原因,蛇鲻类资源严重衰退[8],导致目前其渔获物主要以当龄鱼和幼鱼为主[4,9]。
在渔业管理中,为防止过度捕捞导致种群适应力的衰退,科学的做法是根据不同种群的特征来分配不同的捕捞强度[10-12]。种群遗传学通过研究种群中基因和基因型频率的维持和变化,判定种群的遗传结构、评估种群的遗传多样性等特征,在渔业管理上具有重要参考价值[13]。目前对花斑蛇鲻的研究基本上集中在年龄与生长[2,4-5,9]、繁殖[14]、种群动力学[15-16]及资源量[17-18]等方面,未见针对种群遗传特征的相关报道。本研究利用种群遗传分析中广泛使用的线粒体细胞色素b (cytochrome b,cytb)基因序列作为分子标记,分析了花斑蛇鲻的种群遗传结构特征,并对其遗传多样性进行评价,以期为该渔业资源的保护和利用提供参考依据。
1. 材料与方法
1.1 实验材料
花斑蛇鲻样本通过底层单拖网渔船采集,采集时间为2013年7月至2016年7月。经鉴定和形态学测量后,置于−20 ℃冰冻带回实验室。共选取了8个采样点,每个采样点随机挑选30余尾样本进行实验。样本采集点位置见图1,位点的经纬度等信息见表1。
![]() 表 1 花斑蛇鲻样本信息及cytb基因序列遗传多样性参数Table 1 Specimen information of S. undosquamis and genetic diversity parameters based on cytb gene sequences
表 1 花斑蛇鲻样本信息及cytb基因序列遗传多样性参数Table 1 Specimen information of S. undosquamis and genetic diversity parameters based on cytb gene sequences采样点
sampling
site经度/纬度
longitude/
latitude样本量 (N)
number of
samples单倍型数量 (H)
number of
haplotypes多态性位点数 (S)
number of polym-
orphic sites单倍型多样性 (h±SD)
haplotype
diversity核苷酸多样性 (π±SD)
nucleotide
diversity防城港 FCG 108°30'E/21°00'N 32 21 25 0.943 5±0.028 7 0.003 145±0.001 823 北部湾 BBW 107°15'E/19°15'N 34 22 26 0.953 7±0.021 9 0.003 205±0.001 850 西沙 XS 109°24'E/16°40'N 34 31 37 0.992 9±0.009 9 0.003 852±0.002 168 三亚 SY 109°46'E/17°58'N 35 27 41 0.968 1±0.020 3 0.003 724±0.002 103 海口 HK 111°18'E/20°18'N 34 21 32 0.925 1±0.035 8 0.003 154±0.001 824 珠江口 ZJK 114°05'E/21°41'N 34 27 38 0.985 7±0.010 8 0.003 692±0.002 089 汕头 ST 116°55'E/23°00'N 32 23 34 0.949 6±0.029 0 0.003 497±0.001 997 泉州 QZ 119°02'E/24°36'N 31 22 29 0.961 3±0.021 6 0.003 393±0.001 948 总计 total − 266 142 144 0.965 0±0.006 1 0.003 455±0.001 921 1.2 基因组DNA提取
剪取花斑蛇鲻背部肌肉约30 mg置于1.5 mL离心管内,加入海洋动物基因组DNA抽提试剂盒(天根,北京)中的裂解液,使用快速组织细胞破碎仪(BulletBlender STORM,美国)研磨成糊状,再依据试剂盒说明提取基因组总DNA。用EonTM微孔板分光光度计(BioTek,美国)检测DNA浓度后,将DNA稀释至100 ng·μL−1,−20 ℃保存。
1.3 目标片段扩增与序列测定
参照GenBank中蛇鲻属的线粒体全基因组序列,设计并筛选出1对用于cytb基因序列的扩增引物:SunCytb-F (5'-CAGGATTTGAAGCCACCCCTACTAT-3')和Sun-Cytb-R (5'-GCTTTGGGAGTCAGTGGTAGAGGTT-3')。依照ExTaq酶(TaKaRa,大连)的使用说明构建序列扩增反应体系(按每个反应50 μL计算)。PCR反应程序为:94 ℃预变性2 min;94 ℃变性20 s,60 ℃退火20 s,72 ℃延伸90 s,30个循环;最后72 ℃延伸3 min。用1%的琼脂糖凝胶电泳检测和分离扩增产物,切取单一条带凝胶回收纯化,将回收产物双向测序。
1.4 数据分析
利用DNAStar软件包中的SeqMan拼接正反序列,用BioEdit 7.2.5[19]整合所有样品的序列并手工校正,用MEGA 7.0[20]中ClustalW进行多重排列比对(alignment)。通过软件ModelTest[21]选择核苷酸替代最适模型,基于AIC (Akaike information criterion)标准筛选出最适模型及相关参数用于进化分析。
利用Dnasp 6[22]生成cytb序列所有样本的单倍型,在Network 5 (www.fluxus-engineering.com)中基于最大似然法(maximum parsimony, MP)[23]构建中间连接网络图(median joining network,MJN)[24]。遗传多样性指数(表1)通过Dnasp 6和Arlequin 3.5[25]计算获得。利用Arlequin 3.5软件中的分子方差分析(analysis of molecular variances, AMOVA),通过5 000次重复抽样来检验不同遗传结构水平上的协方差的显著性,计算两两群体间遗传分化系数FST,并采用种群分化测试检测单倍型在两两群体间分布频率的差异,以上分析的显著性均通过10 000次重复抽样来检验。采用Tajima's D检验[26]和Fu's FS检验[27]两种中性检验(neutrality tests)来检测种群进化是否严格遵循中性理论,采用核苷酸错配分布分析(mismatch distribution)[28-29],分别检验在突然扩张模型(sudden expansion model)和空间扩散模型(spatial expansion model)下观测值和期望值的分布是否一致,以判断花斑蛇鲻种群历史上是否存在扩张事件,用粗糙指数[30] (Harpending's raggedness index, HRI)评估模型的有效性(10 000次重复抽样)。
根据公式t=τ/2u[31]计算种群扩张后的繁殖代数。其中τ为扩张时间参数(在Arlequin中根据核苷酸错配分布计算获得),u为序列的突变速率(u=2μk,其中k为cytb序列长度,μ为每个核苷酸位点的突变速率),突变速率取每百万年1%~2.5%[32]。以花斑蛇鲻平均代时(性成熟年龄) 1年[4]来换算种群扩张开始的时间。
2. 结果
2.1 序列特征及遗传多样性
花斑蛇鲻cytb基因全序列长度为1 141 bp,以不完全终止密码子(incomplete stop codon) T结尾。8个地理群体266尾样本共检测到257个变异位点,其中144个为多态性信息位点。样本总计包含142个单倍型,大多数单倍型(116个)属于个体特异单倍型,另外26个单倍型为共享单倍型,其中共享比例最高的为4个单倍型(图2):H8 (占所有个体的13.9%)、H11 (8.3%)、H1 (7.9%)和H30 (5%)。H8、H11和H1被全部8个群体共有,H30被6个群体共有。花斑蛇鲻各个群体间cytb序列的遗传多样性指数(表1)并无显著差异,与总体处于同一水平。样本总体表现出很高的单倍型多样性(0.965 0±0.006 1)和较低的核苷酸多样性(0.003 5±0.001 9)。
![]() 图 2 花斑蛇鲻cytb基因序列单倍型的中间连接网络图圆的大小代表单倍型频率,颜色代表所属群体,空心圆代表未检测到的单倍型,省略了单倍型之间的突变数Fig. 2 Median-joining network for cytb gene sequence haplotypes of S. undosquamisThe size of the circles is proportional to haplotype frequency and the colors represent the corresponding population. Small hollow circles represent missing haplotypes. The mutation steps between two connected haplotypes are omitted.
图 2 花斑蛇鲻cytb基因序列单倍型的中间连接网络图圆的大小代表单倍型频率,颜色代表所属群体,空心圆代表未检测到的单倍型,省略了单倍型之间的突变数Fig. 2 Median-joining network for cytb gene sequence haplotypes of S. undosquamisThe size of the circles is proportional to haplotype frequency and the colors represent the corresponding population. Small hollow circles represent missing haplotypes. The mutation steps between two connected haplotypes are omitted.2.2 单倍型网络关系
基于中间连接法构建的花斑蛇鲻cytb序列单倍型网络图见图2,各单倍型以4个主要的共享单倍型H8、H11、H1和H30为中心相连接形成星状发散(star-shape)结构,并没有区分出明显的支系。来自各个地理群体的样品交织在一起,也没有呈现特殊的单倍型偏好。
2.3 种群遗传结构
花斑蛇鲻cytb序列的分子方差分析表明绝大部分(99.79%)的遗传变异来自于地理群体内部,只有极少部分的遗传变异来自于群体间(表2)。总体的遗传分化系数仅为0.002 1 (P=0.572 6),表明各个群体间基因交流很频繁,不存在显著的遗传分化。花斑蛇鲻8个地理群体间遗传分化水平很低(表3),除了两个配对群体之间(XS-SY和XS-HK),其余群体之间的FST值均很小甚至是负值,且统计检验不显著,表明群体间存在高度的遗传同质性。另外,绝大部分群体间的随机交配假设检验水平并不显著(表4),符合单倍型在群体间是随机分布的假设。整体样本的检测结果(P=1.000 0)也显示花斑蛇鲻群体间符合随机交配群的假设,与分子方差分析结果一致。
表 2 花斑蛇鲻8个地理群体cytb基因序列遗传变异的分子方差分析Table 2 Analysis of molecular variance for eight populations of S. undosquamis based on cytb gene sequences变异来源
source of variation自由度
degree of freedom变异百分比
percentage of variation分化系数
F statisticsP 群体间 among populations 7 0.21 0.002 1 0.572 6 群体内 within populations 258 99.79 所有样本 total samples 265 表 3 花斑蛇鲻两两地理群体间cytb基因序列的遗传分化系数 (对角线下方) 及显著性水平 (对角线上方)Table 3 Pairwise FST (below diagonal) and P values (above diagonal) among geographic populations of S. undosquamis based on cytb gene sequences群体 population FCG BBW XS SY HK ZJK ST QZ 防城港 FCG 0.773 8 0.372 0 0.342 6 0.288 2 0.540 9 0.779 1 0.807 2 北部湾 BBW −0.013 1 0.394 1 0.460 9 0.304 5 0.650 3 0.846 7 0.659 4 西沙 XS −0.000 9 −0.001 2 0.077 4 0.041 5 0.225 2 0.419 6 0.622 3 三亚 SY −0.000 2 −0.003 9 0.021 1 0.896 7 0.729 2 0.762 6 0.178 8 海口 HK 0.002 4 0.001 9 0.031 8 −0.012 9 0.747 6 0.588 1 0.118 5 珠江口 ZJK −0.006 7 −0.008 1 0.005 7 −0.008 9 −0.009 9 0.896 3 0.247 1 汕头 ST −0.012 5 −0.013 1 −0.002 4 −0.010 5 −0.007 9 −0.012 9 0.695 9 泉州 QZ −0.014 1 −0.009 9 −0.007 4 0.010 5 0.018 9 0.004 5 −0.010 3 表 4 花斑蛇鲻两两地理群体间随机交配假设检验的显著性水平Table 4 P values of exact test of sample differentiation of S. undosquamis based on cytb gene haplotype frequencies群体 population FCG BBW XS SY HK ZJK ST 北部湾 BBW 0.014 6 西沙 XS 0.067 7 0.109 4 三亚 SY 0.019 9 0.362 7 0.187 7 海口 HK 0.035 1 0.197 3 0.027 3 0.736 8 珠江口 ZJK 0.285 5 0.254 7 0.692 4 0.292 1 0.181 8 汕头 ST 0.390 6 0.236 1 0.269 5 0.704 2 0.873 0 0.674 6 泉州 QZ 0.527 0 0.310 6 0.128 8 0.303 3 0.486 4 0.392 6 0.945 3 2.4 种群历史动态
花斑蛇鲻cytb序列的核苷酸错配分布的参数估算值(表5)显示,所有群体和样本总体的吻合度检验的HRI值在2种扩张模型的假设下均不显著(P>0.05),表明核苷酸错配分布符合这2种扩张模型的假设,即种群历史上既有数量上的快速扩张也有空间分布范围的扩散。核苷酸错配分布与发生频率的关系见图3,两两序列差异的分布呈明显的单峰形,且观测值与期望值非常吻合,同样印证了上述观点。种群历史的中性检验结果(表5)显示,所有群体的Tajima's D值和Fu's FS值均呈现显著(P<0.05)和极显著的负值(P=0),显著偏离中性理论下的Wright-Fisher模型,是种群历史上快速扩张事件的典型表现。根据Arlequin计算的扩张时间参数(τ)的观测值4.43,估算出花斑蛇鲻种群的扩张时间约发生在4万~10万年前。
表 5 花斑蛇鲻cytb基因序列核苷酸错配分布分析的参数估计值和中性检验统计值Table 5 Mismatch distribution parameter estimates and neutrality tests statistics for S. undosquamis based on cytb gene sequences群体
population错配分布 mismatch distribution 中性检验 neutrality test 突然扩张模型
sudden expansion model空间扩散模型
spatial expansion modelTajima' D Fu's FS 粗糙指数
HRIP 粗糙指数
HRIP D P FS P 防城港 FCG 0.782 9 0.936 2 0.590 0 0.950 1 −1.493 5 0.048 6 −14.339 4 0 北部湾 BBW 0.066 7 0.067 2 0.028 5 0.061 4 −1.492 5 0.048 5 −15.139 0 0 西沙 XS 0.075 9 0.313 3 0.214 5 0.173 7 −1.854 0 0.013 3 −25.721 9 0 三亚 SY 0.049 8 0.188 5 0.151 9 0.513 6 −2.068 2 0.004 2 −23.293 0 0 海口 HK 0.749 0 0.897 7 0.629 0 0.911 5 −1.926 2 0.010 6 −13.437 8 0 珠江口 ZJK 0.050 7 0.047 7 0.045 6 0.077 7 −1.973 3 0.008 2 −24.127 8 0 汕头 ST 0.890 7 0.834 6 0.909 0 0.934 4 −1.909 1 0.011 5 −17.081 6 0 泉州 QZ 0.136 3 0.217 8 0.156 5 0.352 5 −1.680 7 0.026 9 −15.934 9 0 合计 total 0.177 0 0.439 0 0.076 1 0.601 0 −2.555 7 0 −25.510 6 0 ![]() 图 3 花斑蛇鲻cytb序列单倍型核苷酸错配分布曲线柱状图表示观测值, 虚线为突然扩张模型下的期望值,实线为空间扩散模型下的期望值Fig. 3 Mismatch distribution of cytb haplotypes for S. undosquamisThe observed pairwise differences are shown in bars and the expected values under the sudden expansion model and spatial expansion model are in dash line and solid line, respectively.
图 3 花斑蛇鲻cytb序列单倍型核苷酸错配分布曲线柱状图表示观测值, 虚线为突然扩张模型下的期望值,实线为空间扩散模型下的期望值Fig. 3 Mismatch distribution of cytb haplotypes for S. undosquamisThe observed pairwise differences are shown in bars and the expected values under the sudden expansion model and spatial expansion model are in dash line and solid line, respectively.3. 讨论
遗传多样性是生物多样性形成的基础,物种遗传多样性的高低与其对环境的适应能力成正比[33]。在渔业资源开发利用中,正确认识和评价鱼类的遗传多样性水平,是可持续开发利用该渔业资源的基础。花斑蛇鲻作为重要的经济鱼类,资源开发力度很大。就本研究的结果来看,目前中国海域的花斑蛇鲻具有较高的遗传多样性(h=0.925 1~0.992 9),与同样基于cytb序列标记分析的其他海水鱼类如绿鳍马面鲀(Thamnaconus septentrionalis,h=0.927 0)[34]、短尾大眼鲷(Priacanthus macracanthus,h=0.874 1)[35]、鮸鱼(Miichthys miiuy,h=0.960 0)[36]、小黄鱼(Larimichthys polyactis,h=0.984 0)[37]和大泷六线鱼(Hexagrammos otakii,h=0.739 0)[38]等处于同一水平。已有研究证实鱼类的遗传多样性会受捕捞压力的影响,尤其是对于有效群体大小(Ne)较低且群体遗传结构显著的种类来说,过度捕捞会显著降低其遗传多样性[39]。目前在持续较高强度的捕捞压力下,花斑蛇鲻的h仍处于较高水平,可能由于原有的资源量基数大,且分布范围内基因交流频繁,这在一定程度上抵消或延迟了人类活动对其种质衰退效应的显现。另外也存在遗传多样性已经降低的可能,但由于缺乏历史研究数据而无从对比。因此本研究可作为一个基线资料,可供后续的遗传种质监测作本底参照。
花斑蛇鲻总体的单倍型多样性高(h=0.965 0)而核苷酸多样性较低(π=0.003 455)的模式,也与上述几种海洋鱼类类似。根据Grant和Bowen[40]对海水鱼类遗传多样性(h和π值)的划分模式,具有较高的h (>0.5)和较低的π (<0.005)可能是种群历史上经历过瓶颈效应后快速扩张的表现。在扩张过程中群体数量增加,使得单倍型数量显著提高,但尚未有足够的时间来积累核苷酸差异。这也是大多数海洋鱼类具备的特征。
花斑蛇鲻历史上种群扩张的推测也被中性检测所佐证,Tajima's D和Fu's FS检验结果均表明其显著偏离稳定种群模型。另外,单倍型网络的星状发散模式和核苷酸错配分布曲线的单峰形状同样表明花斑蛇鲻历史上经历过种群的迅速扩张,扩张时间约在更新世晚期。更新世所在的第四纪晚期,由于冰期和间冰期交替气候[41]的影响,西北太平洋边缘海的海平面反复升降[42]。推测在末次冰期结束、海平面上升后,花斑蛇鲻从深海避难所重新向外殖化,栖息地发生大面积扩张,种群得以迅速扩张。该海域其他鱼类的种群历史分析也显示,不少种类如短尾大眼鲷[35]、黄斑篮子鱼(Siganus oramin)[43]、扁舵鲣(Auxis thazard)[44]和圆舵鲣(A. rochei)[45]等均经历了更新世的群体扩张事件。
种群是渔业资源利用和保护的基本单位,鱼类种群遗传特征的研究有助于合理划分渔业管理单元。本研究中基于cytb序列单倍型网络分析显示花斑蛇鲻不存在显著遗传分化的支系,分子方差分析显示遗传变异几乎(99.79%)都来自群体内部,群体间也普遍不具备明显的遗传分化(FST为−0.014 14~0.031 79),表明花斑蛇鲻地理群体间的基因交流很强烈,可以视为随机交配的单一种群(unit population)。
海洋环境由于缺乏物理屏障,许多具有较强扩散力的生物体在很大的空间范围内表现出很低的遗传分化[46-47]。原因主要在于他们的浮游性卵和幼体可以借助洋流扩散,使不同地理群体间产生遗传同质化[40]。南海的季风漂流、中国沿岸流[48]和黑潮支流等可以促使花斑蛇鲻的卵和仔稚鱼扩散到较远的海域,从而产生基因交流,与同属的多齿蛇鲻[49]以及南海其他鱼类(如上述短尾大眼鲷等种类)遗传结构模式类似。依据Laikre等[50]的定义,这种没有分化的种群(no differentiation)在渔业资源的开发利用上可作为一个管理单元(management units,MUs)来看待。然而,利用线粒体序列标记进行种群遗传分析毕竟仍有分辨率不够精细的局限性,后续研究中可以结合更多的手段如RAD、GBS等技术来进行研究,以期获得更为确切的结果,为渔业管理措施的制定提供支撑。
-
![]()
图 2 2015—2019年不同空间尺度的长鳍金枪鱼资源第一级标准差椭圆
Figure 2. First order standard deviational ellipse spatial distribution of T. alalunga at different scales from 2015 to 2019
![]()
图 3 2015—2019年不同尺度的长鳍金枪鱼资源空间格局差异指数
Figure 3. Spatial pattern difference index of T. alalunga resources at different scales from 2015 to 2019
![]()
图 4 2015—2019年不同尺度的长鳍金枪鱼资源空间格局的分布范围
Figure 4. Distribution range of spatial pattern of T. alalunga resources at different scales from 2015 to 2019
![]()
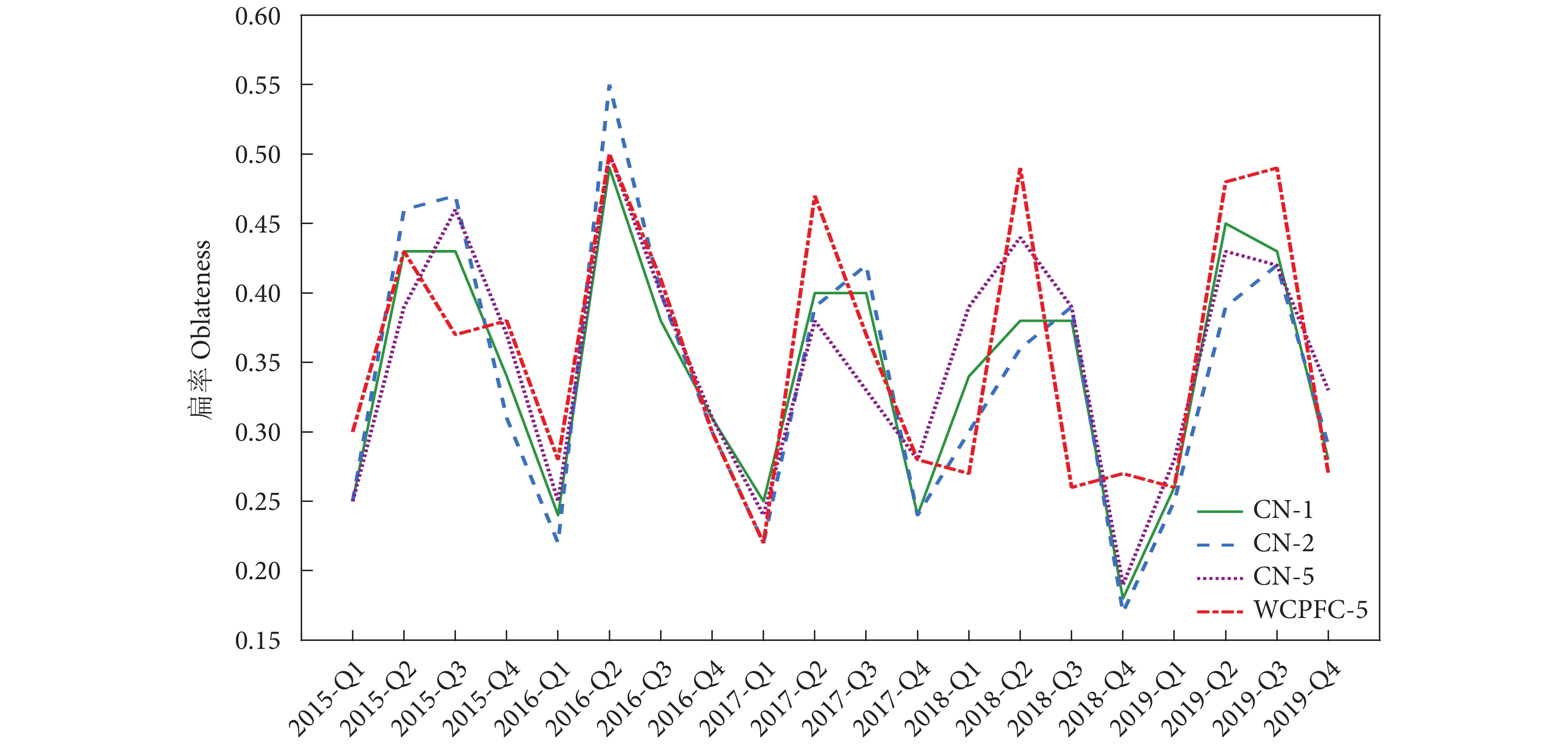
图 5 2015—2019年不同尺度的长鳍金枪鱼资源标准差椭圆扁率分布
Figure 5. Oblateness of standard deviation ellipticity of T. alalunga resources at different scales from 2015 to 2019
![]()
图 6 2015—2019年不同尺度的长鳍金枪鱼资源标准差椭圆方位角分布
Figure 6. Azimuth of standard deviation ellipticity of T. alalunga resources at different scales from 2015 to 2019
![]()
图 7 南太平洋长鳍金枪鱼渔业资源第三季度不同尺度的空间格局的局部聚集特征
Figure 7. Aggregation characteristics of spatial pattern of T. alalunga fishery resources at different scales in South Pacific in third quarter
表 1 南太平洋长鳍金枪鱼延绳钓渔业船次
Table 1 Number of vessels of longline fishery of T. alalunga in South Pacific
年份Year 船次 (CN)Number of vessels 船次 (WCPFC)Number of vessels 2015 189 1 752 2016 138 1 695 2017 159 1 764 2018 150 1 764 2019 156 1 817  下载: 导出CSV
下载: 导出CSV
表 2 南太平洋长鳍金枪鱼资源空间格局的全局分布聚集性显著性检验
Table 2 Significance test of global distribution and aggregation of spatial pattern of T. alalunga in South Pacific
统计时间
Statistical time中国大陆1°
CN-1中国大陆2°
CN-2中国大陆5°
CN-5中西太平洋渔业委员会5°
WCPFC-5I Z_score I Z_score I Z_score I Z_score 第一季度
Quarter 10.032 3 3.975 1 0.236 6 15.015 6 0.337 4 18.301 4 0.628 7 43.204 7 第二季度
Quarter 20.032 1 2.184 4 0.047 3 10.867 5 0.371 1 17.594 5 0.563 7 47.969 7 第三季度
Quarter 30.042 2 6.137 2 0.064 5 20.338 6 0.487 0 30.940 6 0.614 7 50.730 2 第四季度
Quarter 40.038 1 6.675 1 0.046 4 20.380 1 0.532 6 29.146 5 0.637 4 49.767 8 注:Z得分的临界值为1.65、1.96和2.58分别对应的置信度为90%、95%和99%。Note: The critical values of Z score are 1.65, 1.96 and 2.58, respectively, with confidence levels of 90%, 95% and 99%.
下载: 导出CSV
-
[1] PILLING G M, HARLEY S J, WILLIAMS P, et al. Trends in the south Pacific albacore longline and troll fisheries (WCPFC-SC14-2018/SA-IP-08)[R]. 14th Regular Session of the Scientific Committee, Busan, Republic of Korea, 8-16 August, 2018.
[2] NIKOLIC N, MORANDEAU G, HOARAU L, et al. Review of albacore tuna, Thunnus alalunga, biology, fisheries and management[J]. Rev Fish Biol Fish, 2017(27): 775-810.
[3] 牛明香, 王俊, 黄海中, 等. 黄海中南部越冬鳀空间格局的年际变化[J]. 海洋环境科学, 2019, 38(2): 263-271. doi: 10.12111/j.cnki.mes20190215 [4] YANG X P, JIA Y T, WANG Q H, et al. Space-time evolution of the ecological security of regional urban tourism: the case of Hubei Province, China[J]. Environ Monit Assess, 2021, 193(9): 1-20.
[5] 侯娟, 周为峰, 樊伟, 等. 基于集成学习的南太平洋长鳍金枪鱼渔场预报模型研究[J]. 南方水产科学, 2020, 16(5): 42-50. doi: 10.12131/20200022 [6] 毛江美, 陈新军, 余景. 基于神经网络的南太平洋长鳍金枪鱼渔场预报[J]. 海洋学报, 2016, 38(10): 34-43. [7] TREMBLAY B L, HAMPTON J, MCKECHNIE S, et al. Stock assessment of South Pacific albacore tuna (WCPFC-SC14-2018/SA-WP-05) [R]. Busan, Republic of Korea: The Pacific Community (SPC), 2018: 8-16.
[8] 安树伟, 常瑞祥. 中国沿海地区生产性服务业与制造业空间关系演变研究——基于113个城市面板数据的分析[J]. 中国软科学, 2017(11): 101-110. doi: 10.3969/j.issn.1002-9753.2017.11.010 [9] PING J L, GREEN C J, ZARTMAN R E, et al. Exploring spatial dependence of cotton yield using global and local autocorrelation statistics[J]. Field Crops Res, 2004, 89(2): 219-236.
[10] ESRI帮助文档. 空间自相关 (Global Moran's I) 的工作原理[EB/OL]. [2022-07-04]. https://desktop.arcgis.com/zh-cn/arcmap/latest/tools/spatial-statistics-toolbox/h-how-spatial-autocorrelation-moran-s-i-spatial-st.htm. [11] MUHLING B A, LAMKIN J T, ALEMANY F, et al. Reproduction and larval biology in tunas, and the importance of restricted area spawning grounds[J]. Rev Fish Biol Fisher, 2017, 27(4): 697-732. doi: 10.1007/s11160-017-9471-4
[12] ESRI帮助文档. 热点分析 (Getis-Ord Gi*) 的工作原理. [EB/OL]. [2022-07-04]. https://desktop.arcgis.com/zh-cn/arcmap/10.3/tools/spatial-statistics-toolbox/h-how-hot-spot-analysis-getis-ord-gi-spatial-stati.htm. [13] BRIAND K, MOLONY B, LEHODEY P. A study on the variability of the albacore (Thunnus alalunga) longline catch rates in the Southwest Pacific Ocean[J]. Fish Oceanogr, 2011, 20(6): 517-529. doi: 10.1111/j.1365-2419.2011.00599.x
[14] 樊伟, 张晶, 周为峰. 南太平洋长鳍金枪鱼延绳钓渔场与海水表面温度的关系分析[J]. 大连水产学院学报, 2007, 22(5): 366-371. [15] 闫敏, 张衡, 伍玉梅, 等. 2011年南太平洋长鳍金枪鱼渔场时空分布及其与主要海洋环境变化特征[J]. 渔业信息与战略, 2015, 30(2): 119-127. doi: 10.13233/j.cnki.fishis.2015.02.007 [16] 周甦芳, 樊伟. 太平洋延绳钓长鳍金枪鱼及渔场水温分析[J]. 海洋湖沼通报, 2006(2): 38-43. doi: 10.3969/j.issn.1003-6482.2006.02.007 [17] 张嘉容, 杨晓明, 田思泉. 基于最大熵模型的南太平洋长鳍金枪鱼栖息地预测[J]. 中国水产科学, 2020, 27(10): 1222-1233. [18] 魏广恩, 陈新军. 不同环境模态下空间分辨率对北太平洋柔鱼CPUE标准化的影响[J]. 海洋科学, 2021, 45(4): 147-158. doi: 10.11759/hykx20190722003 [19] 闫敏, 张衡, 伍玉梅, 等. 基于GAM模型研究时空及环境因子对南太平洋长鳍金枪鱼渔场的影响[J]. 大连海洋大学学报, 2015, 30(6): 681-685. doi: 10.16535/j.cnki.dlhyxb.2015.06.018 [20] 张嘉容, 杨晓明, 戴小杰, 等. 南太平洋长鳍金枪鱼延绳钓渔获率与环境因子的关系研究[J]. 南方水产科学, 2020, 16(1): 69-77. doi: 10.12131/20190178 [21] 郭刚刚, 张胜茂, 樊伟, 等. 南太平洋长鳍金枪鱼垂直活动水层空间分析[J]. 南方水产科学, 2016, 12(5): 123-130. doi: 10.3969/j.issn.2095-0780.2016.05.016 [22] 官文江, 陈新军, 高峰, 等. GLM 模型和回归树模型在CPUE标准化中的比较分析[J]. 上海海洋大学学报, 2014, 23(1): 123-130. [23] HARLEY S J, MYERS R A, DUNN A. Is catch-per-unit-effort proportional to abundance?[J]. Can J Fish Aquat Sci, 2001, 58(9): 1760-1772. doi: 10.1139/f01-112
[24] YE Y, DENNIS D. How reliable are the abundance indices derived from commercial catch-effort standardization?[J]. Can J Fish Aquat Sci, 2009, 66(7): 1169-1178. doi: 10.1139/F09-070
[25] 张勋, 张禹, 周爱忠, 等. 我国远洋渔业渔具发展概况[J]. 中国农业科技导报, 2013, 15(6): 16-19. doi: 10.3969/j.issn.1008-0864.2013.06.03 [26] 刘世禄, 冯小花, 陈辉. 关于加快发展我国远洋渔业的战略思考[J]. 渔业现代化, 2014, 41(4): 63-67, 2. doi: 10.3969/j.issn.1007-9580.2014.04.014 [27] 张衡, 张瑛瑛, 叶锦玉. 中国远洋渔业发展的新思路及建议[J]. 渔业信息与战略, 2019, 34(1): 30-35. doi: 10.13233/j.cnki.fishis.2019.01.005 [28] ASHLEY J W, VALERIE A, SIMON J N, et al. Vertical behavior and diet of albacore tuna (Thunnus alalunga) vary with latitude in the South Pacific Ocean[J]. Deep-Sea Res II, 2015, 113: 154-169. doi: 10.1016/j.dsr2.2014.03.010
[29] 宋利明, 谢凯, 赵海龙, 等. 库克群岛海域海洋环境因子对长鳍金枪鱼渔获率的影响[J]. 海洋通报, 2017, 36(1): 96-106. doi: 10.11840/j.issn.1001-6392.2017.01.013 [30] 原作辉, 杨东海, 樊伟, 等. 基于卫星AIS的中西太平洋金枪鱼延绳钓渔场分布研究[J]. 海洋渔业, 2018, 40(6): 649-659. doi: 10.3969/j.issn.1004-2490.2018.06.002 [31] 刘禹希, 王学锋, 吕少梁, 等. 南海北部海域大眼鲷空间自相关性[J]. 水产学报, 2021, 45(8): 1361-1373. [32] 江承旭. 斐济专属经济区长鳍金枪鱼渔场分析[D]. 上海: 上海海洋大学, 2017: 38-39. [33] CHILDERS J, BETCHER A. Summary of the 2005 U. S. North and South Pacific albacore troll fisheries[R/OL]. [2022-1-25]. http://swfsc.noaa.gov/uploadedFiles/Divisions/FRD/Large_Pelagics/Albacore/SUMMARY2005.
[34] LU H J, LEE K T, CHENG H L. On the relationship between El Niño/Southern oscillation and South Pacific albacore[J]. Fish Res, 1998, 39(1): 1-7. doi: 10.1016/S0165-7836(98)00174-X
[35] 杨晓明, 戴小杰, 田思泉, 等. 中西太平洋鲣鱼围网渔业资源的热点分析和空间异质性[J]. 生态学报, 2014, 34(13): 3771-3778.












计量
- 文章访问数: 563
- HTML全文浏览量: 208
- PDF下载量: 44
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
