
Community structure of Thaliacea in the Zhongsha and Xisha Islands, South China Sea
-
摘要:
根据2014年~2015年中沙西沙海域12°30'N~17°32'N、110°00'E ~110°11'E 4个航次调查资料, 对该海域海樽的种类组成及季节变化、总栖息密度和生物多样性分布以及群落格局进行了研究。结果表明, 共鉴定出海樽23种, 夏季种类最多为16种, 其次为冬季15种, 春季14种, 秋季10种; 冬季与其他3个季节在种类组成上存在比较明显的更替现象, 其他季节更替现象不明显; 中沙西沙海域海樽类平均丰度为0.99个·m-3, 夏季最高为1.93个·m-3, 其次为冬季1.12个·m-3, 春季为0.59个·m-3, 秋季最小为0.32个·m-3; 周年优势种小齿海樽(Doliolum denticulatum)和双尾纽鳃樽(Thalia democratic)以及春、夏季的优势种大西洋火体虫(Pyrosoma atlanticum)是影响中沙西沙海域海樽类数量分布格局的主要种类; 优势种的集聚分布导致中沙西沙海域海樽类多样性水平一般; 多维尺度分析表明中沙西沙海域海樽类群落的季节变化较小。海樽类栖息密度的平面分布及群落格局与海流的变化密切相关。
Abstract:Based on the four seasonal investigation data collected from the Zhongsha and Xisha Islands, South China Sea (12°30'N~17°32'N, 110°00'N~110°11'N) during 2014~2015, we investigated the characteristics of thaliacean community. Twenty-three species were catched (16 in summer, 15 in winter, 14 in spring and 10 in autumn). According to the result of alternation fraction (R), the seasonal alternation of the species between winter and the other seasons was more obvious. The average abundance was 0.99 ind·m-3 (1.93 ind·m-3 in summer, 1.12 ind·m-3 in winter and 0.32 ind·m-3 in autumn). Doliolum denticulatum and Thalia democratica in four seasons and Pyrosoma atlanticum in spring and summer were dominant in abundance distribution. The dominant species' clustering led to normal diversity. The results of MDS show no obvious change in thaliacean community structures among seasons. The distribution of thaliacean is highly related with surface currents.
-
海樽类属尾索动物亚门,海樽纲,半肌目,全海产,是一类低等脊索动物,是海洋浮游动物的一个重要类群,尤其是在热带海域的浮游生物中占有较大的比重。这类动物由于个体大,终生浮游,对水文环境敏感等,常作为水团或海流的指示种[1-2]。有报道海樽类是乌鲳(Formio niger)、南海北部鲐鱼(Pneumatophorus japonicus)和深水金线鱼(Nemipterus bathybius)等的饵料,其密集区常是渔获量的高产区[3]。此外,海樽类是唯一能以超微和微型浮游植物为食的大型浮游动物,是微型食物环能量流动的重要环节[4]。而且,海樽类有较快的沉降速率、排泄物中含有较高的碳,在海洋碳汇中也具有重要作用[5]。
国外对海樽类有不少报道,GODEAUX[6]系统地介绍了印度洋海樽类的研究结果;AMARA等[7]研究了巴西南部大陆架水域的海樽类分布、丰度、垂直移动以及与水团关系等;还有大量关于优势种的相关研究,如对纽鳃樽科种类的研究[8-11]。中国对海樽类的研究主要集中在东海和台湾海峡海域,主要有海樽类种类组成、多样性变化、数量分布以及与环境的关系等[2, 12-17]。另外,在黄海、大亚湾和北部湾也有少量相关研究[3, 18-20]。
中沙西沙海域位于中国南海中北部,海洋生物资源种类繁多,且具有独特的热带珊瑚礁和深海大洋性的生态特点,具有巨大的生产潜力,既是中国传统的优良渔场,又是海上的交通要冲,地理位置十分重要[21]。因此,对该海域海樽类的专题研究,对于海洋生态学研究的深入开展和渔业资源的开发,具有重要的理论和实践意义。但目前除1984年对其进行过相关研究[5]外,尚未见其他相关报道。文章根据2014年~2015年在中沙西沙海域所获样品,对春、夏、秋和冬4个季节海樽类的种类组成及季节变化、数量分布和群落格局进行研究,以弥补中沙西沙海域此方面研究的不足。
1. 材料与方法
1.1 样品采集与分析
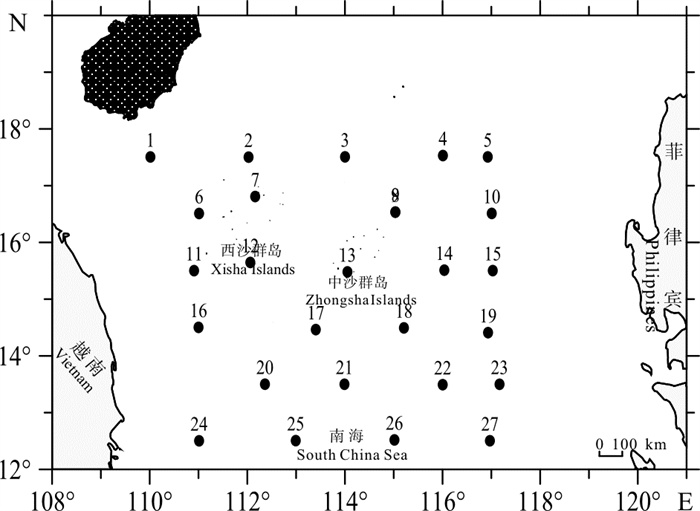
材料来源于2014年3月、7月~8月、11月和2015年1月中沙西沙海域(12°30′N~17°32′N、110°00′E~110°11′E)按季度开展的4个航次调查,采样站位设置见图 1。使用大型浮游生物网(网长280 cm,网口内径80 cm,网口面积0.5 m2,网孔直径0.505 mm)从水深200 m处至表层垂直采集样品。样品的处理、保存、计数等均按《海洋调查规范》进行[22]。
![]()
1.2 数据分析
采用种类更替率(R)进行优势种类组成的季节变化研究,其计算公式为R=[(a+b-2c)/(a+b-c)]×100%,a与b分别为相邻2个季节的种数,c为相邻2个季节共同的种数[23]。
优势种确定采用优势度(Y),计算公式为Y=(Ni/N)×fi,式中Ni表示第i种的个体数;N表示海樽类总个体数;fi表示某种生物的出现频率;Y≥0.02的物种为优势种[24]。
采用Shannon-Wiener物种多样性指数(H′)、Pielou物种均匀度指数(J′)及多样性阈值(Dv)综合分析海樽类的多样性,其计算公式分别为:
$$ H^{\prime}=-\sum P_i \log _2 P_i $$ (1) $$ J^{\prime}=H^{\prime} / \log (S) $$ (2) $$ D v=H^{\prime} \times J^{\prime} $$ (3) 式中S为群落中的总种类数,Pi为第i种的个体丰度占同一样品海樽类总丰度的比值[24]。
海樽类群落结构的多变量分析采用非参数多变量分析方法。首先将海樽类丰度进行4次方根转换,然后采用PRIMER 6.0进行非度量多维标度(MDS)分析来研究其群落结构组成[25]。
调查站位图、海樽类各季节丰度平面分布图以及多样性指数的平面分布采用Golden Software Surfer 11.0软件绘制。
2. 结果
2.1 种类组成与季节变化
2014年~2015年四季中沙西沙海域共鉴定海樽类23种,其中冬季15种,春季14种,夏季16种,秋季10种(表 1)。冬季与其他3个季节在种类组成上存在比较明显的更替现象,其中冬-春季种类更替率最高为55%,其次为冬-秋季为53%,冬-夏季相对较低为52.4%;秋季与春季和夏季也存在一定的更替,种类更替率分别为40%和38%;春-夏季种类更替率最低,为33.3%。
表 1 中沙西沙海域海樽类种类组成Table 1. Species during four seasonal surveys in the Zhongsha and Xisha Islands物种
species季节season 冬winter 春spring 夏autumn 秋summer 长吻纽鳃樽Brooksia rostrata + + + 羽环纽鳃樽Cyclosalpa pinnata + + + 软拟海樽Dolioletta gegenbauri + + 殖离海樽Doliolina separata + + + 小齿海樽Doliolum denticulatum + + + + 宽肌纽鳃樽Iasis zonaria + Pyrosoma aherniosum + 大西洋火体虫Pyrosoma atlanticum + + + + 梭形纽鳃樽Salpa fusiformis + + + Thalia cicar + + + + 双尾纽鳃樽Thalia democratica + + + + 双尾纽鳃樽东方亚种Thalia orientalis + + + + 多手纽鳃樽Traustedtia multitentaculata + + + 韦氏纽鳃樽Weelia cylindrica + + + 环纽鳃樽属Cyclosalpa sp. + 海樽类幼虫Doliolum larvae + 黄纽鳃樽Thalia rhomboides + + + Dolionaria sp. + 佛环纽鳃樽Cyclosalpa floridana + 贝环纽鳃樽Cyclosalpa bakeri + 安纽鳃樽Ritteriella amboinensis + 双尾纽鳃樽多刺亚种Thalia democratica echinata + 火体虫Pyrosoma sp. + 2.2 海樽类丰度的平面分布
中沙西沙海域海樽类平均丰度为0.99个·m-3,夏季最高为1.93个·m-3,其次为冬季1.12个·m-3,春季和秋季数量明显减少,分别为0.59个·m-3和0.32个·m-3。
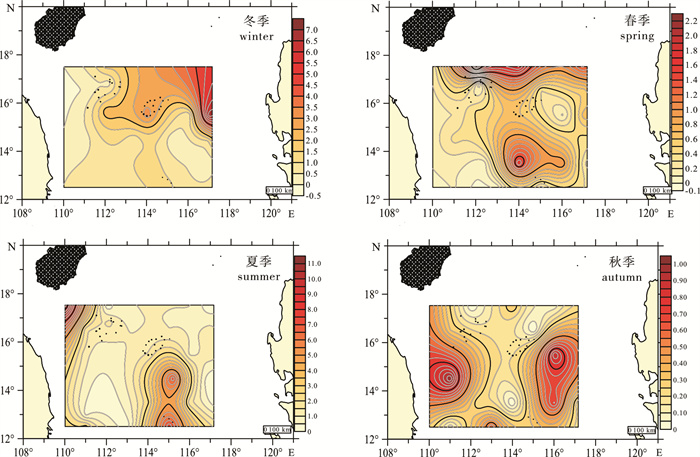
各季节海樽类的平面分布存在一定差异(图 2)。冬季海樽类在该海域的东北部形成密度高峰区,主要以小齿海樽(Doliolum denticulatum)、双尾纽鳃樽(Thalia democratica)构成;春季丰度则呈北部及南中部区域高于其他海域,小齿海樽、双尾纽鳃樽和大西洋火体虫(Pyrosoma atlanticum)为该季节优势种;夏季西北部及东南部形成高密度区,优势种类最多,双尾纽鳃樽、小齿海樽和大西洋火体虫同春季一致,长吻海樽(Brooksia rostrata)、殖离海樽(Doliolina separata)为该季节独有优势物种;秋季高密度区则移至东、西及南部的中部区域,以小齿海樽和双尾纽鳃樽为优势种。
![]() 图 2 中沙西沙海域海樽类栖息密度(个·m-3)的平面分布Figure 2. Spatial distribution of the thaliacean density (ind·m-3) in the Zhongsha and Xisha Islands
图 2 中沙西沙海域海樽类栖息密度(个·m-3)的平面分布Figure 2. Spatial distribution of the thaliacean density (ind·m-3) in the Zhongsha and Xisha Islands2.3 物种多样性分布
中沙西沙海域海樽类种类多样性水平较低,H′、J′及Dv分别为1.09、0.69和0.83(表 2)。其中H′夏季最高,秋季最低;J′则以秋季最高,冬季最低。根据多样性阈值标准(>3.5为非常丰富;2.6~3.5为丰富;1.6~2.5为较好;0.6~1.5为一般; < 0.6为差)对该海域海樽类多样性水平进行评价,各季海樽类多样性水平均为一般。多样性阈值既反映了总数量又反映了每个物种分配的均匀程度[15, 24]。海樽类具有一定的集聚行为,其栖息密度高峰区仅仅是优势物种的高密度分布,因此该海域海樽类多样性阈值较低。
表 2 中沙西沙海域海樽类多样性指数Table 2. The diversity indices of the tunieate in the Zhongsha and Xisha Islands季节
season多样性指数(H′)
diversity index均匀度指数(J′)
evenness index多样性阈值(Dv)
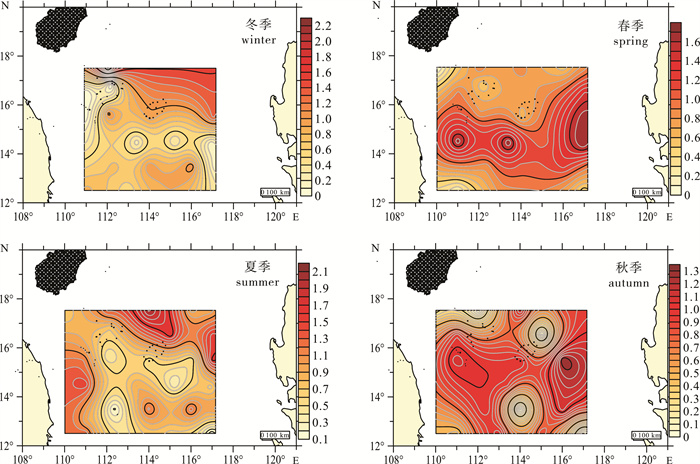
threshold value冬季winter 1.00±0.70 0.54±0.17 0.64±0.56 春季spring 1.15±0.56 0.74±0.25 0.96±0.50 夏季summer 1.33±0.46 0.67±0.18 0.95±0.48 秋季autumn 0.91±0.49 0.82±0.16 0.75±0.41 均值average 1.09±0.18 0.69±0.12 0.83±0.16 冬季中沙西沙海域北部多样性水平高于其他海域,春季多样性较高区域移至海域中部,夏季则是中北部、中东部及中西部高于中部和南部区域;秋季整体多样性较其他季节低,但分布较为均匀,东部和西部高于南部和北部(图 3)。
![]() 图 3 中沙西沙海域海樽类多样性阈值的平面分布Figure 3. Spatial distribution of the Dv index for thaliacean in the Zhongsha and Xisha Islands
图 3 中沙西沙海域海樽类多样性阈值的平面分布Figure 3. Spatial distribution of the Dv index for thaliacean in the Zhongsha and Xisha Islands2.4 群落格局
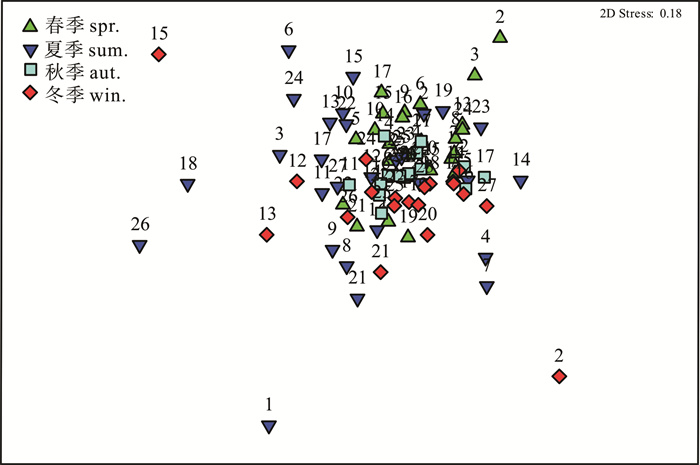
多维尺度分析显示(图 4),中沙西沙海域4个季节海樽类除冬季位于西北部的S2站和东北部的S15站以及夏季西北部的S1、东南部的S18和S26站外基本聚成一类。表明中沙西沙海域海樽类群落的季节变化较小,基本为一个群落。
![]() 图 4 中沙西沙海域海樽类序列分析Figure 4. Multi-Dimensional Scaling (MDS) ordination of thaliacean in the Zhongsha and Xisha Islands
图 4 中沙西沙海域海樽类序列分析Figure 4. Multi-Dimensional Scaling (MDS) ordination of thaliacean in the Zhongsha and Xisha Islands3. 讨论
3.1 海樽类种类数比较
2014年~2015年在中沙西沙海域共采获海樽类23种,种类数高于台湾海峡(13种)[12]、台湾南湾区(16种)[13]、东海(20种)[15]、南黄海(7种)[18]、北黄海(4种)[20]和北部湾(12种)[3]。海樽类是一类以暖水性种类为主的浮游动物[13],南海地处热带,终年温度较高,因此,南海海樽类的种类数高于其他海域。与1984年相比,虽然该海域海樽类出现种类数相等,但种类组成存在一定差异[5]。近年来随着全球变暖加剧,各大海域的海面温度随之不同程度的升高,海流分布也发生一定的改变,从而导致对水文环境比较敏感的浮游动物结构和分布的改变。环境的变化可能是导致中沙西沙海域海樽类种类组成变化的主要原因。
3.2 海樽类栖息密度平面分布与海流的关系
ACHA等[26]研究发现,浮游动物经常在水面辐合区域发生集聚。海樽类作为浮游动物的一个重要类群,由于个体大,终生浮游,对水文环境尤其敏感,其分布情况与海流有密切联系,常作为水团或海流的指示种,同时也是上升流区浮游动物的重要类群之一[12]。
冬季,中沙西沙海域盛行东北风,受一个较强的双涡旋气旋式环流控制,其中一个涡旋中心位于该海域的东北部(117.4°E, 17.0°N),且一支黑潮分支从吕宋海峡侵入南海[27]。因此,海樽类在中沙西沙海域东北部的涡旋中心处形成高密度区。而且,能准确反映黑潮水入侵陆架水的指示种双尾纽鳃樽东方亚种(Thalia orientalis)[15-17],在此处出现了数量最高值。
春季,是季风转换期,随着东北季风的减弱,海流特征也发生了变化。原来的气旋型双圈结构解体,从海南岛外海沿广东外海陆坡存在一支自西向东的海流,即南海暖流穿过中沙西沙海域的北部,在吕宋岛西南侧外海岸存在一个较强的气旋型环流[27]。该季节海樽类数量高值区正好与南海暖流流经处及气旋型环流的位置,在时间上和空间上吻合,呈北部及南中部区域高于其他区域的趋势。
夏季,中沙西沙海域盛行西南季风,整个海域受强大的东北向沿岸流影响,且海流在西北部海域即海南岛东南外海部区域与其附近的上升流交汇[27-28]。另有研究发现在15°N左右,南北两侧各有一个气旋式涡旋和反气旋式涡旋,其中心分别位于114.3°E, 15.6°N和115.3°E, 14.0°N附近[27, 29]。故夏季,在中沙西沙海域西北部及东南部的上升流区和涡旋区形成海樽类的高密度区,且优势种组成最为复杂。此外夏季由于季风的旋度影响,在越南中部存在上升流,但上升流附近的S16、S20和S24站优势种小齿海樽、双尾纽鳃樽、大西洋火体虫和长吻海樽数量较低,且无其他种类分布。海樽类为滤食性,过多的浮游植物会堵塞其进食部位,进而造成其密度的降低[5]。上升流处于越南沿岸流作用强区域,随之携带的丰富营养物质能促进浮游植物的大量繁殖,可能造成海樽类进食部位的堵塞。因此,该区域海樽类数量低、种类少。
秋季,处于夏季向冬季的过渡时期,沿岸流继续作用,整个海域存在多个中小尺度的环流。例如临近中沙西沙海域西中部的气旋型环流(中心位置(110.5°E,12.8°N附近)和中沙西沙海域中东部区域的环流区(即吕宋海峡附近119.4°E,18.3°N)。此时,海樽类栖息密度的高峰区则对应分布在环流的附近海域。
3.3 海樽类群落结构与环境的关系
从多维尺度分析可以看出,除冬季西北部的S2站和东北部S15站以及夏季西北部的S1站、东南部的S18和S26站明显离散分布外,4个季节海樽类基本聚在一起。该结果与LIN和LIN[5]研究较为一致,造成这种现象的主要原因可能是该海域温度、盐度整体上平面分布差异相对较小[24]。然而,虽然海樽类大多数适温、适盐性基本一致,但有些物种则只能在一定的温、盐条件下生存,不能在大范围内广泛分布[16-17]。冬季S2站与其他站位明显在种类组成方面有所差异,其出现独有的热带大洋种安纽鳃樽(Ritteriella amboinensis)[17],且研究发现安纽鳃樽的分布与高温水团有关,随着高温水团消退,该物种随之消失[30-31]。冬季位于北部的站位受来自于太平洋高温水团黑潮分支作用,由于气候条件限制,冬季只在北部的S2站进行了样品采集,因而安纽鳃樽有可能随黑潮分支携带由大洋进入该区域。而冬季S15站含有1个独有物种双尾纽鳃樽多刺亚种Thalia democratica echinata,且丰度达2个·m-3,属亚热带外海种的高盐适应亚群,具相对较低的最适温度和较高的最适盐度(>34)[17]。徐兆礼等[17]通过对东海海樽类生态类群研究同样发现,双尾纽鳃樽多刺亚种主要分布在盐度较高的外海区域。冬季,S15站附近海域受盐度较高的外海水团黑潮分支及吕宋半岛低温高盐冷涡的综合作用[27-28],从而使其成为该种最优的生存区域。同步所测温度和盐度数据同样显示冬季S15站具相对较低的温度和较高的盐度,该站表层温度(25.48 ℃)除高于S13站(25.09 ℃)外,均低于其他站位,而表层盐度(33.58)则均大于其他站位。夏季西北部的S1站仅有3种优势种,分别为长吻纽鳃樽、双尾纽鳃樽和小齿海樽,其丰度分别为8个·m-3、2个·m-3和0.8个·m-3。其中除小齿海樽属广布种外,长吻纽鳃樽和双尾纽鳃樽则属于亚热带外海种的高温适应亚群,最适温度均超过25 ℃[17]。夏季,表层平均温度(29.17±0.83) ℃为全年最高。因此,长吻纽鳃樽为夏季的独有优势种,且S1站处于越南沿岸流和海南上升流的交汇处,具有丰富的营养物质,故其具最高栖息密度。夏季S18和S26站附近的气旋式涡旋和反气旋式涡,可能是造成这2个站位于整体群落外的主要原因。因此,总体来说,海樽类群落结构是对温度、盐度以及海流等多种环境因素适应的结果。
致谢: 样品鉴定得到了厦门大学黄加祺教授、许振祖教授和国家海洋局第三海洋研究所的陈瑞祥研究员和林茂研究员的帮助,“南锋”号调查船全体人员在样品采集过程中提供帮助,谨此致谢! -
![]()
![]()
图 2 中沙西沙海域海樽类栖息密度(个·m-3)的平面分布
Figure 2. Spatial distribution of the thaliacean density (ind·m-3) in the Zhongsha and Xisha Islands
![]()
图 3 中沙西沙海域海樽类多样性阈值的平面分布
Figure 3. Spatial distribution of the Dv index for thaliacean in the Zhongsha and Xisha Islands
![]()
图 4 中沙西沙海域海樽类序列分析
Figure 4. Multi-Dimensional Scaling (MDS) ordination of thaliacean in the Zhongsha and Xisha Islands
表 1 中沙西沙海域海樽类种类组成
Table 1 Species during four seasonal surveys in the Zhongsha and Xisha Islands
物种
species季节season 冬winter 春spring 夏autumn 秋summer 长吻纽鳃樽Brooksia rostrata + + + 羽环纽鳃樽Cyclosalpa pinnata + + + 软拟海樽Dolioletta gegenbauri + + 殖离海樽Doliolina separata + + + 小齿海樽Doliolum denticulatum + + + + 宽肌纽鳃樽Iasis zonaria + Pyrosoma aherniosum + 大西洋火体虫Pyrosoma atlanticum + + + + 梭形纽鳃樽Salpa fusiformis + + + Thalia cicar + + + + 双尾纽鳃樽Thalia democratica + + + + 双尾纽鳃樽东方亚种Thalia orientalis + + + + 多手纽鳃樽Traustedtia multitentaculata + + + 韦氏纽鳃樽Weelia cylindrica + + + 环纽鳃樽属Cyclosalpa sp. + 海樽类幼虫Doliolum larvae + 黄纽鳃樽Thalia rhomboides + + + Dolionaria sp. + 佛环纽鳃樽Cyclosalpa floridana + 贝环纽鳃樽Cyclosalpa bakeri + 安纽鳃樽Ritteriella amboinensis + 双尾纽鳃樽多刺亚种Thalia democratica echinata + 火体虫Pyrosoma sp. +  下载: 导出CSV
下载: 导出CSV
表 2 中沙西沙海域海樽类多样性指数
Table 2 The diversity indices of the tunieate in the Zhongsha and Xisha Islands
季节
season多样性指数(H′)
diversity index均匀度指数(J′)
evenness index多样性阈值(Dv)
threshold value冬季winter 1.00±0.70 0.54±0.17 0.64±0.56 春季spring 1.15±0.56 0.74±0.25 0.96±0.50 夏季summer 1.33±0.46 0.67±0.18 0.95±0.48 秋季autumn 0.91±0.49 0.82±0.16 0.75±0.41 均值average 1.09±0.18 0.69±0.12 0.83±0.16
下载: 导出CSV
-
[1] 郑重, 李少菁, 许振祖. 海洋浮游生物学[M]. 北京: 海洋出版社, 1984: 532-546. https://xueshu.baidu.com/usercenter/paper/show?paperid=cb79007b4da85634aeb3099598bf4218&site=xueshu_se&hitarticle=1 [2] 张金标, 连光山, 王云龙, 等. 台湾海峡东部海樽类的分布[J]. 台湾海峡, 2003, 22(3): 279-285. doi: 10.3969/j.issn.1000-8160.2003.03.001 [3] 王学锋, 李纯厚, 贾晓平, 等. 北部湾海樽类的种类组成及分布[J]. 渔业现代化, 2010, 37(2): 59-63. doi: 10.3969/j.issn.1007-9580.2010.02.014 [4] NAKAMURA Y. Blooms of tunicates Oikopleura spp. and Dolioletta gegenbauri in the Seto Inland Sea, Japan, during summer[J]. Hydrobiologia, 1998, 385(1/2/3): 183-192. https://xueshu.baidu.com/usercenter/paper/show?paperid=84bd683f19de4b160409e1f520be6f18&site=xueshu_se&hitarticle=1
[5] LIN M, LIN R C. Seasonal abundance and distribution of pelagictunicates(Chordata: Thaliacea)in the central South China Sea[J]. Acta Oceanologica Sinica, 2006, 25(1): 100-107. https://xueshu.baidu.com/usercenter/paper/show?paperid=807cd18e06b71d363d257d0a6456049b&site=xueshu_se
[6] GODEAUX J. Pelagic tunicates of the Indian Ocean[J]. J Mar Biol Assoc Ind, 1972, 14(1): 263-292.
[7] AMARA W J A, MONTU M A, GLOEDEN I M, et al. Salpidae (Thaliacea) in the continental shelf of southern Brazil: composition, distribution and abundance(summer, 1990)[J]. Atlantica, 1997, 19: 51-66.
[8] DEIBEL D. Laboratory determined mortality, fecundity and growth rates of Thaliade mocratica for skaland Dolioletta gegenbauri Uljanin(Tunicata, Thaliacea)[J]. J Plankton Res, 1982, 4(1): 537-541.
[9] DAPONTE M C, CAPITANIO L F, MACHINANDIARENA L, et al. Planktonictunicates(Chordata, Tunicata)of the RTMA "Evrika" in the southwestern Atlantic Ocean(1988)[J]. Iheringia, Serie Zoológica, 1993, 74: 71-80.
[10] ESNAL G B, DAPONTE M C, SIMONE L C. Pelagictunicates(Appendicularia and Thaliacea)from the tropical western Pacific Ocean[J]. Physis Session, 1993, 48(114/115): 41-48. https://xueshu.baidu.com/usercenter/paper/show?paperid=2b8b16675bfeeaf5a5da39e398862f65&site=xueshu_se&hitarticle=1
[11] GIBSON D M, PAFFENHOFER G A. Feeding and growth rates of the doliolid, Dolioletta gegenbauri Uljanin (Tunicata, Thaliacea)[J]. J Plankton Res, 2000, 22(8): 1485-1500. doi: 10.1093/plankt/22.8.1485
[12] 林茂. 台湾海峡西部海域海樽类的初步分析[J]. 海洋通报, 1988, 7(4): 66-71. [13] 张金标, 黄将修, 连光山. 台湾南湾区秋末冬初海樽类的种类组成和数量分布[J]. 海洋通报, 2003, 22(6): 9-16. doi: 10.3969/j.issn.1001-6392.2003.06.002 [14] 徐兆礼, 林茂, 张金标. 东海海樽类(Thaliacea)优势种的数量变化和生态适应分析[J]. 动物学报, 2006, 52(1): 53-62. [15] 徐兆礼, 林茂. 东海海樽类种类组成和多样性[J]. 海洋水产研究, 2007, 28(1): 26-32. https://xueshu.baidu.com/usercenter/paper/show?paperid=1t720ea0e23w0p00yp5c0ck07j470855&site=xueshu_se&hitarticle=1 [16] 徐兆礼, 林茂, 张金标. 东海海樽类数量分布及与环境的关系[J]. 海洋与湖沼, 2007, 38(6): 549-554. doi: 10.3321/j.issn:0029-814x.2007.06.010 [17] 徐兆礼, 孙军, 林茂. 东海海樽类生态类群统计分析[J]. 生态学报, 2008, 28(11): 5698-5705. doi: 10.3321/j.issn:1000-0933.2008.11.056 [18] 高尚武. 东海水母类, 浮游贝类及被囊类的垂直分布[J]. 海洋科学集刊, 1990, 31: 83-92. [19] 林茂. 大亚湾海樽类生态的研究[M]. 北京: 海洋出版社, 1990: 390-396. [20] 陈洪举, 刘光兴, 姜强, 等. 北黄海海樽类的种类组成和分布特征[J]. 中国海洋大学学报(自然科学版), 2015, 45(6): 39-44. https://www.cnki.com.cn/Article/CJFDTOTAL-QDHY201506006.htm [21] 颜云榕, 冯波, 卢伙胜. 中、西沙海域2种灯光作业渔船的捕捞特性及其技术效率分析[J]. 南方水产, 2009, 5(6): 59-64. doi: 10.3969/j.issn.1673-2227.2009.06.011 [22] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763.6—2007. 海洋调查规范(第6部分海洋生物调查)[S]. 北京: 中国标准出版社, 2007: 34-38. [23] 杜飞雁, 王雪辉, 谷阳光, 等. 南沙群岛西南大陆斜坡海域浮游动物的垂直分布[J]. 海洋学报(中文版), 2014, 36(6): 94-103. https://www.cnki.com.cn/Article/CJFDTOTAL-SEAC201406012.htm [24] 王亮根, 杜飞雁, 李亚芳, 等. 西南季风暴发前后南沙海域浮游桡足类群落特征比较研究[J]. 南方水产科学, 2015, 11(5): 47-55. doi: 10.3969/j.issn.2095-0780.2015.05.006 [25] CLARKE K R, WARWICK R M. Change in marine communities: an approach to statistical analysis and interpretation[M]. 2nd ed. UK: PRIMER-E Ltd, 2001: 42-54. https://xueshu.baidu.com/usercenter/paper/show?paperid=1f6y0eb0yf0f0r20we3q0mr01n514763&site=xueshu_se
[26] ACHA E M, MIANZAN H W, GUERRERO R A, et al. Marine fronts at the continental shelves of austral South America physical and ecological processes[J]. J Mar Sys, 2004, 44(1/2): 83-105. https://xueshu.baidu.com/usercenter/paper/show?paperid=8c3bdfba26a62c7a0a91b13f4448ecaa&site=xueshu_se&hitarticle=1
[27] 鲍李峰, 陆洋, 王勇, 等. 利用多年卫星测高资料研究南海上层环流季节特征[J]. 地球物理学报, 2005, 48(3): 543-550. [28] 万道静. 南海海表盐度的分布特征[D]. 青岛: 中国海洋大学, 2014: 22-25. https://xueshu.baidu.com/usercenter/paper/show?paperid=d3a3623fde87818d6d33d887bfe9aec7&site=xueshu_se&hitarticle=1 [29] 刘勇刚, 袁耀初, 苏纪兰, 等. 1998年夏季南海环流[J]. 科学通报, 2000, 45(12): 1252-1259. https://xueshu.baidu.com/usercenter/paper/show?paperid=8db583476a3220ce4c78fcfd64c75c6d&site=xueshu_se&hitarticle=1 [30] FRANCO P, CHEN H J, LIU G X. Distribution and abundance of pelagic tunicates in the North Yellow Sea[J]. J Ocean Univ China, 2014, 13(5): 782-790. https://xueshu.baidu.com/usercenter/paper/show?paperid=5786719662021b7cf0b4bb4505a79536&site=xueshu_se
[31] CLARA M H, BERTHA E L, GILBERTO G C, et al. Marine Ecology Progress Series. Composition and pot ential grazing impact of salp assemblages off Baja California during the 1997-1999 El Niño and La Niña[J]. Mar Ecol Prog Ser, 2006, 318: 123-140.
-
期刊类型引用(6)
1. 熊遇安,邬博文,陈万东,林立,蔡厚才,陈永久. 浙江南麂列岛潮间带多毛类环节动物多样性研究. 浙江海洋大学学报(自然科学版). 2024(05): 449-452+458 .  百度学术
百度学术
2. 刘晓红,石军,徐阁,王德鸿,娄全胜. 西沙宣德群岛中北部海域浮游动物群落结构. 广东海洋大学学报. 2024(06): 134-140 . 百度学术
3. 李开枝,柯志新,王军星,谭烨辉. 西沙群岛珊瑚礁海域浮游动物群落结构初步分析. 热带海洋学报. 2022(02): 121-131 . 百度学术
4. 黄彬彬,吴风霞,郑淑娴,田丰歌,何静. 雷州半岛以东近岸海域浮游动物群落结构与环境因子关系. 广东海洋大学学报. 2022(02): 53-61 . 百度学术
5. 陈俊锦,张经纬,刘时桥,陈万利,秦永鹏,吴时国. 南海中沙群岛海域表层沉积物粒度特征及其输运趋势. 海洋地质与第四纪地质. 2022(02): 15-27 . 百度学术
6. 龚玉艳,杨玉滔,范江涛,蔡研聪,徐姗楠,陈作志. 南海北部陆架斜坡海域夏季浮游动物群落的空间分布. 南方水产科学. 2017(05): 8-15 . 本站查看
其他类型引用(0)
计量
- 文章访问数: 3411
- HTML全文浏览量: 178
- PDF下载量: 1200
- 被引次数: 6
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
