
Genetic characterization of Oreochromis niloticus (♀) × Sarotherodon melanotheron (♂) hybrid F2 and F3 by microsatellite analysis
-
摘要:
利用罗非鱼第二代遗传连锁图谱中微卫星标记对尼罗罗非鱼(Oreochromis niloticus)(♀)×萨罗罗非鱼(Sarotherodon melanotheron)(♂)杂交后代F2、F3群体遗传特征进行了初步比较分析。结果显示,28对引物中有22对引物能有效扩增,未观察到杂交后代F2、F3中有多倍体个体现象。筛选出7个在尼萨罗非鱼杂交后代F2、F3群体中存在差异的位点。杂交F2群体平均等位基因(Na)为2.71,平均多态信息含量(PIC)为0.466,平均观测杂合度(Ho)为0.632;杂交F3群体平均Na为2.14,平均PIC为0.370,平均Ho为0.432,杂交F3遗传杂合性较杂交F2降低。F2自繁F3过程中,F3群体4个位点的等位基因与F2完全相同,3个位点的等位基因少于F2,且F3群体在2个位点出现完全纯合,杂交F3群体位点等位基因呈现纯合趋势。研究结果为尼萨罗非鱼杂交世代遗传变异与杂交利用积累基础资料。
Abstract:A preliminary comparative analysis of genetic characteristics between F2 and F3 hybrid populations of Oreochromis niloticus♀×Sarotherodon melanotheron♂ was conducted by using microsatellite markers from the second generation genetic linkage map of Nile tilapia. The results show that 22 pairs of primers were amplified and no polyploidy individual was detected in F2 and F3 populations, seven different amplified loci were screened out in hybrids F2 and F3 populations from 28 loci. In F2 hybrid population, average allele was 2.71;average polymorphic information content was 0.466;observed heterozygosity was 0.632. In F3 hybrids, the average allele was 2.14;average polymorphic information content was 0.370;observed heterozygosity was 0.432. The genetic heterozygosity of F3 population was lower than that of F2 population. In F3 population, the allele number at four loci was identical with F2 population, while the allele numbers at three loci were less than that of F2 population. Moreover, there were two homozygous loci in F3 population; the change of alleles in F3 population showed a homozygous trend. The results provide some fundamental data for inheritance of different generations and tilapia hybridization utilization.
-
光裸方格星虫(Sipunculus nudus Linnaeus)又称方格星虫,为暖水性、世界广布种类,在北部湾沿海滩涂和国内浅海均有分布[1-3]。方格星虫营养价值较高,含有丰富的氨基酸、多糖、脂肪酸以及多种有益微量元素[4-5]。研究发现方格星虫具有重要的经济和药用价值[6],民间称其为“海洋中的冬虫夏草”,从其体内提取的活性物质在提高免疫力、抗疲劳等方面具有功效[7-10]。随着方格星虫繁育技术的突破,其滩涂增养殖规模逐年增加,也开展了一定规模的试验性池塘养殖。以往研究显示方格星虫、刺参(Stichopus japonicus)等底栖动物的矿物元素组成具有明显的地区或环境差异,如温岭地区的方格星虫体壁钙(Ca)、铁(Fe)、锌(Zn)含量均显著高于北海和海口(P<0.05)[11];深海和池塘养殖的刺参体壁矿物元素如铬(Cr)、锰(Mn)、铜(Cu)含量较为接近,而Zn、硼(B)、铝(Al)之间具有差异(P<0.05)[12]。方格星虫通过吻部摄取适宜粒径的表层沉积物[13],主要由底栖硅藻、浮游生物沉降、大型藻类碎片、陆源性有机物质以及沙质颗粒等组成,表层沉积物的元素含量会一定程度影响方格星虫体壁矿物元素组成。由于不同有机物质的脂肪酸组成不同,同种生物摄取不同有机质时,体内会呈现出不同的食物脂肪酸特征[14],因此不同环境中的方格星虫会表现出不同的脂肪酸组成。方格星虫的增养殖空间亟需拓展,而在近海生态环境保护下,开展池塘增养殖是重要措施。以往开展了方格星虫体内的矿物元素含量、脂肪酸和氨基酸组成的研究[4,15-16],而有关不同养殖方式对方格星虫体内矿物元素、脂肪酸和氨基酸组成变化的影响鲜有报道。本研究以滩涂和池塘2种养殖方式的方格星虫作为研究对象,测定方格星虫矿物元素及体壁营养组成,分析不同养殖方式对方格星虫营养物质的影响,以期为方格星虫的养殖模式及开发利用提供理论依据。
1. 材料与方法
1.1 研究地点
2016年11月对湛江遂溪滩涂(9.99 hm2)和3个池塘(每个3.33 hm2)养殖的方格星虫及相应沉积物进行采样与测定,分析不同养殖方式对方格星虫营养组成的影响。滩涂区域的采样点位于中低潮区,距离海岸线200 m,沉积物表层饵料丰富,底质含沙量大于95%。池塘养殖区的底质含沙量也在95%以上,藻类组成稳定。方格星虫样品(每条9~10 g)为2016年4月同一批次放苗,滩涂和池塘增养殖密度均为40 条·m–2,滩涂区域不投饵,而池塘增养殖过程中需要投入氮磷肥以调节藻相。
1.2 样品采集与测定
在滩涂和池塘分别设置3个采样区域,并在每个采样区域采集3个重复样品并混合。在每个采样区域共采集30条方格星虫和500 g表层沉积物(0~3 cm)样品,共检测3个生物样品和3个沉积物样品。将方格星虫的消化道及内容物清除,并剪碎混合冷冻保存。生物样品及沉积物样品均置于 – 20 ℃冷冻保存。
采用凯氏定氮法测定方格星虫体壁蛋白质含量(凯氏定氮仪,岛津UDK132,日本),索氏抽提法测定脂肪含量,先炭化再放入马弗炉中550 ℃灼烧测定灰分,含水率通过烘干前后的质量差值进行计算。根据国际通用方法[17]采用气相色谱仪(Agilent 6890,美国)测定生物样品中的脂肪酸组成。参照GB 5009.124—2016采用氨基酸分析仪(日立L-8900,日本)测定生物样品中的氨基酸组成。
称取0.5 g方格星虫体壁进行湿法消解,先后在75 ℃和135 ℃进行消解,再定容到25 mL进行矿物元素测定。采用质谱仪测定砷(As)、钴(Co) (安捷伦7700X,美国),全自动双道氢化物发生原子荧光光度计测定硒(Se)和锡(Sn) (北京海光AFS9750,中国),其余的矿物元素Ca、Fe、Zn、Cu、Mn、Cr、钾(K)、钠(Na)、镁(Mg)、铅(Pb)、镍(Ni)、镉(Cd)参照GB 5009.268—2014采用原子吸收分光光度仪测定(日立Z-2000,日本)。
采用100目筛取沉积物样品,四分法取过筛样品。称取0.2 g样品进行除As以外的元素测定。采用四酸法消解样品,定容至50 mL待测,参照GB/T 6041—2002采用质谱仪(安捷伦7700 x,美国)和光谱仪测定(安捷伦Duo AA 55B/240Z,美国)。称取0.2 g沉积物样品,加入10 mL王水,沸水浴加热2 h,定容至25 mL,参照GB 5009.268—2014采用全自动双道氢化物发生原子荧光光度计(北京海光AFS3100,中国)测定总As含量。以往研究采用C16:1 (n-7)/C16:0比值和C20:5 (n-3)的含量作为硅藻脂肪酸标志,这2个数值越大表明硅藻脂肪酸特征越明显,硅藻对动物体内的饵料贡献越大[18]。
1.3 数据分析
采用SPSS 17.0进行数据统计和分析,所得数据以“平均值±标准差(
$\overline X \pm {\rm SD} $ )”表示,百分数值进行反正弦转换后再方差分析,不同处理组间的数据差异采用One-Way ANOVA分析,P<0.05为差异显著。2. 结果
2.1 方格星虫基本营养成分
滩涂和池塘中的方格星虫营养组成无显著差异(P>0.05)。2种养殖方式的方格星虫体壁组织的含水率、蛋白质、脂肪和灰分组成均无显著差异(P>0.05,表1)。
表 1 不同养殖方式的方格星虫主要营养成分Table 1. Essential nutrients of S.nudus in different aquaculture modes% 组别
group水分
moisture蛋白质
crude protein脂肪
crude lipid灰分
ash滩涂 tidal flat 80.12±0.13 13.16±0.25 0.61±0.03 6.08±0.18 池塘 pond 80.15±0.18 13.12±0.15 0.60±0.03 6.05±0.13 2.2 方格星虫体壁脂肪酸组成
滩涂和池塘养殖的方格星虫体壁脂肪酸总质量分数分别为3.47 mg·g–1和3.42 mg·g–1(P>0.05,表2),饱和脂肪酸质量分数分别为2.44 mg·g–1和2.49 mg·g–1,占相应样品脂肪酸总量的70%和73%,而不饱和脂肪酸(unsaturated fatty acid,UFA)分别占相应样品的30%和27%。滩涂和池塘增养殖的方格星虫脂肪酸组成均呈现出UFA小于SFA的趋势;多不饱和脂肪酸质量分数分别为0.907 mg·g–1和0.912 mg·g–1,占各样品脂肪酸总质量分数的26.11%和26.64% (P>0.05)。滩涂增养殖的方格星虫未检出C22:0和C20:3,而池塘中的方格星虫中未检出C16:1(表2)。滩涂养殖方格星虫C16:1 (n-7)/C16:0比值和C20:5 (n-3)质量分数均高于池塘(P<0.05),而C18:2 (n-6)和C23:0质量分数表现出相反趋势(表2和表3)。
表 2 不同养殖方式的方格星虫脂肪酸质量分数Table 2. Fatty acids of S.nudus in different aquaculture modesmg·g–1 脂肪酸种类
fatty acid滩涂养殖
tidal flat mode池塘养殖
pond model十四碳酸/C14∶0 myristic acid 0.391±0.010 0.360±0.009 十五碳酸/C15∶0 pentadecanoic acid 0.043±0.004 0.049±0.001 十六碳酸/C16∶0 hexadecanoic acid 0.578±0.016 0.466±0.007 顺-7-十六碳一烯酸/C16∶1 palmitoleic acid 0.054±0.006 – 十七碳酸/C17∶1 heptadecenoic acid 0.149±0.005 0.168±0.007 十八碳酸/C18∶0 stearic acid 0.416±0.008 0.442±0.002 顺-9-十八碳一烯酸/C18∶1 (n-9) oleic acid 0.069±0.003 0.076±0.005 顺,顺-9,12-十八碳二烯酸/C18∶2 (n-6) linoleic acid 0.273±0.013a 0.357±0.023b 二十碳酸/C20:0 arachidic acid 0.066±0.005 0.089±0.002 顺,顺-11,14-二十碳二烯酸/C20∶2 cis-11,14-eicosadienoic acid 0.075±0.006 0.074±0.002 二十二碳酸/C22∶0 behenic acid – 0.072±0.004 顺,顺,顺-8,11,14-二十碳三烯酸/C20∶3 (n-3) cis-8,14,17-eicosatrienoic acid _ 0.052±0.001 二十三碳酸/C23∶0tricosanoic acid 0.803±0.019a 0.914±0.025b 顺-5,8,11,14,17-二十碳五烯酸/C20∶5 (n-3) cis-5,8,11,14,17-eicosapentaenoic acid 0.478±0.023b 0.339±0.008a 顺-4,7,10,13,16,19-二十二碳六烯酸/C22∶6ns cis-4,7,10,13,16,19-docosahexaenoic acid 0.082±0.003 0.089±0.002 注:–. 未检测出;同行不同小写字母表示差异显著(P<0.05),下表同此 Note: –. not detected; different lowercase superscripts in the same row indicate significant difference (P<0.05). The same case in the following table. 表 3 不同养殖方式的方格星虫脂肪酸特征Table 3. Fatty acid characteristics of S.nudus in different aquaculture modes脂肪酸
fatty acid滩涂养殖
tidal flat mode池塘养殖
pond modew(总脂肪酸)/mg·g–1 TFA 3.47±0.12 3.42±0.09 w(不饱和脂肪酸)/mg·g–1 UFA 2.44±0.05 2.49±0.04 w(多不饱和脂肪酸)/mg·g–1 poly-unsaturated fatty acids
C16:1(n-7)/C16:0/%0.907±0.025
9.290.912±0.030
02.3 方格星虫体壁氨基酸组成
滩涂和池塘养殖的方格星虫体壁均检测出16种氨基酸(表4),且氨基酸组成均表现为谷氨酸(Glu)含量最高,其次为甘氨酸(Gly)、丙氨酸(Ala)、精氨酸(Arg)、天冬氨酸(Asp)、赖氨酸(Lys)及亮氨酸(Leu),而组氨酸(His)和蛋氨酸(Met)含量较低。池塘中的方格星虫氨基酸总量高于滩涂中的方格星虫(P<0.05),且池塘中方格星虫的其他氨基酸含量均略高于滩涂养殖(P>0.05,表4)。2种养殖方式下方格星虫体壁各氨基酸占总氨基酸的比例无显著差异(P>0.05),表现为滩涂养殖方格星虫的Ala比例略高于池塘养殖,而Gly和Arg比例略低于池塘养殖(表4)。
表 4 不同养殖方式的方格星虫氨基酸组成Table 4. Amino acids of S.nudus in different aquaculture modes氨基酸种类
amino acid滩涂养殖 tidal flat mode 池塘养殖 pond mode 质量分数/mg·g–1
mass fraction组成比例/%
proportion质量分数/mg·g–1
mass fraction组成比例/%
proportion谷氨酸 Glu 19.20±1.87a 16.55±1.53 24.30±2.39b 16.42±1.62 丙氨酸 Ala 12.80±1.22 11.03±1.06 14.10±1.92 9.53±1.46 甘氨酸 Gly 11.40±1.23 9.83±1.02 15.90±1.83 10.74±1.30 天冬氨酸 Asp 11.40±1.36 9.83±1.03 14.60±1.68 9.86±1.17 精氨酸 Arg 10.10±0.89 8.71±0.76 14.20±1.37 9.59±0.93 赖氨酸 Lys 9.00±0.82 7.76±0.70 10.90±1.25 7.36±0.92 亮氨酸 Leu 8.60±1.07 7.41±0.83 11.20±1.38 7.57±1.01 苏氨酸 Thr 5.20±0.85 4.48±0.75 6.80±0.89 4.59±0.65 丝氨酸 Ser 4.80±0.63 4.14±0.55 6.30±0.85 4.26±0.63 脯氨酸 Pro 4.00±0.52 3.45±0.46 4.90±0.55 3.31±0.38 苯丙氨酸 Phe 4.00±0.48 3.45±0.40 5.00±0.53 3.38±0.35 缬氨酸 Val 3.70±0.32 3.19±0.29 4.60±0.41 3.11±0.27 异亮氨酸 Ile 3.70±0.40 3.19±0.34 4.60±0.33 3.11±0.20 酪氨酸 Tyr 3.50±0.43 3.02±0.38 4.60±0.36 3.11±0.23 蛋氨酸 Met 2.50±0.22 2.16±0.19 3.10±0.34 2.09±0.25 组氨酸 His 2.20±0.27 1.90±0.25 2.80±0.39 1.89±0.31 总氨基酸 total amimo acid 116±13a 148±16b 滩涂和池塘中的方格星虫体壁总氨基酸(total amino acid,TAA)质量分数分别为116 mg·g–1和148 mg·g–1 (P<0.05),而必需氨基酸(essential amino acid,EAA)占TAA的比例分别为31.61%和31.23% (P>0.05),质量分数较高的是Leu和Lys (表4)。方格星虫体内的呈味氨基酸 (delicious amino acid,DAA)主要包括Asp、Gly、Ala、Glu、丝氨酸(Ser)和脯氨酸(Pro),滩涂和池塘中的方格星虫所含DAA的质量分数分别为54.83%和54.12%,两组之间无显著差异(P>0.05,表5)。
表 5 不同养殖方式下方格星虫氨基酸组成特征Table 5. Amino acid characteristics of S.nudus in different aquacultre modes指标
indicator滩涂养殖
tidal flat mode池塘养殖
pond mode必需氨基酸 EAA/mg·g–1 36.7 46.2 呈味氨基酸 DAA/mg·g–1 63.6 80.1 必需氨基酸/总氨基酸 EAA/TAA/% 31.61 31.23 呈味氨基酸/总氨基酸 DAA/TAA/% 54.83 54.12 2.4 养殖环境沉积物和方格星虫体壁矿物元素组成
池塘沉积物矿物元素含量均高于滩涂沉积物,尤其是K、Na、Mg、Ca、Fe、Zn、Cu (P<0.05),除Mn、Cd以外的其他矿物元素也显著高于滩涂(P<0.05,表6)。2种方式下方格星虫体壁矿物元素组成均表现为K>Na>Mg>Ca> Fe>Zn>Sn>Se>As>Ni>Cd>Cr>Co。滩涂和池塘养殖方格星虫体壁的K质量分数最高,分别为3 010 mg·kg–1和3 390 mg·kg–1 (P<0.05),高于相应沉积物中的380 mg·kg–1与1 400 mg·kg–1;Se质量分数分别为0.54 mg·kg–1和0.81 mg·kg–1 (P<0.05),也高于相应沉积物中的0.22 mg·kg–1与0.46 mg·kg–1 (P<0.05)。池塘养殖方格星虫体壁的K、Na、Mg、Ca、Se含量高于滩涂养殖(P<0.05),并且随着沉积物中矿物元素含量的增加,方格星虫体壁中的以上5种矿物元素亦有所增加。池塘沉积物中的Fe、Zn、Mn、Ni、Pb、Cr、Co含量显著高于滩涂沉积物,然而2种养殖方式下方格星虫体壁中的以上几种矿物元素含量相近(P>0.05),沉积物中Fe、Zn、Mn、Ni、Pb、Cr、Co增加并未造成星虫体内的相应矿物元素含量明显增加,表明星虫对Fe、Zn、Mn等矿物元素无明显富集作用。
表 6 不同养殖方式下方格星虫的矿物元素组成Table 6. Mineral elements of S.nududs in different aquaculture modelsmg·g–1 微量元素
trace element滩涂养殖 tidal flat mode 池塘养殖 pond mode 沉积物 sediment 方格星虫 S.nududs 沉积物 sediment 方格星虫 S.nududs 钾 K 380.00±10.68A 3 010.00±23.83a 1 400.00±28.99B 3 390.00±4.74b 钠 Na 1 600.00±45.96A 1 760.00±1.63a 10 000.00±84.85B 2 460.00±20.51b 镁 Mg 520.00±5.37A 261.70±4.01a 1 900.00±19.80B 390.90±3.96b 钙 Ca 77.00±0.88A 113.70±0.82a 1 300.00±43.80B 163.60±6.54b 铁 Fe 1 600.00±13.43A 26.00±0.88 9 200.00±202.23B 28.90±0.18 锌 Zn 2.79±0.13A 5.60±0.16 11.95±0.37B 6.60±0.05 铜 Cu 1.21±0.01A 2.10±0.08b 5.73±0.22B 1.23±0.02a 锰 Mn 28.00±0.99 0.06±0.00 32.00±0.69 0.04±0.00 硒 Se 0.22±0.00A 0.54±0.05a 0.46±0.00B 0.81±0.07b 锡 Sn 0.001±0.000A 2.57±0.08 0.01±0.00B 2.50±0.09 砷 As 0.62±0.00A 0.61±0.02 4.60±0.03B 0.89±0.04 镍 Ni 0.79±0.01A 0.09±0.01 5.06±0.02B 0.09±0.00 铅 Pb 4.09±0.01A 0.09±0.00 7.89±0.09B 0.09±0.00 镉 Cd 0.003±0.000 0.076±0.000 0.005±0.000 0.076±0.001 铬 Cr 2.38±0.03A 0.060±0.00 15.42±0.02B 0.07±0.01 钴 Co 0.25±0.01A 0.03±0.00 1.20±0.04B 0.03±0.00 注:A、B表示不同沉积物矿物元素含量之间具有显著差异(P<0.05);a、b表示方格星虫体壁矿物元素含量之间具有显著差异(P<0.05) Note: A and B indicate significant difference among the element contents in sediments between the two groups (P>0.05); while a and b indicate significant difference of the element contents in S.nudus between the two groups (P<0.05). 3. 讨论
3.1 不同养殖方式对方格星虫脂肪酸组成的影响
方格星虫体壁脂肪酸组成具有地区差异[16],其他研究也表明同一物种摄食不同食物时,体内会呈现出不同的食物脂肪酸特征[14]。以往研究采用C16:1 (n-7)/C16:0比值和C20:5 (n-3)的含量作为硅藻脂肪酸标志,在动物体内硅藻脂肪酸特征越明显,表明硅藻的饵料贡献越大[18]。本研究发现2组方格星虫体壁脂肪酸组成存在一定差异,并且滩涂方格星虫的体壁脂肪酸C16:1 (n-7)/C16:0和C20:5 (n-3)含量均显著高于池塘中的方格星虫,主要原因可能是滩涂和池塘内的饵料组成不同,且滩涂沉积物的硅藻生物量高于池塘。以往研究发现刺参体壁中的C22:0和C20:3 (n-3)具有明显的季节变化特征[19],本实验也发现这2种脂肪酸在不同养殖方式下表现出明显差异,由于刺参和方格星虫均属沉积食性动物,推测主要原因是不同养殖方式、不同季节的微藻等有机质组成变化造成的动物脂肪酸差异。其他研究发现方格星虫体腔液中的UFA浓度高于体壁和内脏[16],且其比例高于SFA,而本研究发现方格星虫体壁的SFA占有较大比例,主要原因是不同组织的脂肪酸含量不同。需要对方格星虫体腔液与体壁、消化道中的脂肪酸组成开展进一步研究。在池塘养殖过程中,通过定向培养高营养价值的底栖硅藻可能更有利于方格星虫营养价值的提升。
3.2 不同养殖方式对方格星虫氨基酸组成的影响
水生动物氨基酸组成受到多种因素的影响,如种类、生长阶段、季节及营养状况等[15]。研究表明,刺参在不同生长阶段具有不同的氨基酸变化规律[20],而方格星虫与刺参的氨基酸总量变化相近[15],可能是两者均以底栖微藻为主的沉积物为食,体内氨基酸总量会受到微藻生物量的影响。本研究池塘中的方格星虫氨基酸总量和Glu含量显著高于滩涂,且每种氨基酸组成均略高于滩涂中的方格星虫,表明养殖方式会影响方格星虫的氨基酸组成。
本研究中滩涂和池塘中的方格星虫均表现出较高含量的Glu,这与其他研究结果相近[15,21-22],可能是方格星虫被称为“天然味精”的主要原因。滩涂和池塘中的方格星虫体壁DAA均表现为Glu>Ala>Gly>Asp>Ser>Pro,且DAA/TAA比值分别为54.83%和54.12%,其他研究也具有相似结果[23]。2种养殖方式下方格星虫的EAA组成无显著差异,表明相同海域的不同养殖方式并不会明显影响方格星虫体内EAA的组成。
3.3 不同养殖方式对方格星虫矿物元素组成的影响
方格星虫的营养价值与其矿物元素组成具有密切关系,可补充调节人体所缺乏的必需矿物元素[4]。本研究滩涂和池塘内的方格星虫矿物元素组成均表现为K>Na>Mg>Ca>Fe>Zn>Cu>Se,并且与其他研究中的矿物元素含量高低顺序相一致[4,24],只是不同研究中的元素含量有所区别,可能与生物规格和养殖环境有关。本研究滩涂和池塘增养殖的方格星虫体壁Sn、As、Ni、Pb、Cd、Cr、Co元素含量相近,并且以上几种元素含量符合食品安全值范围。
以往研究表明方格星虫、刺参等底栖动物体内的矿物元素具有环境差异。温岭地区方格星虫体壁中的Ca、Fe、Zn、Cu、K、P含量均显著高于北海和海口(P<0.05),而北海和海口的方格星虫Fe、Zn、Cu、P含量相近,两地之间的K、Mg、Ca组成具有显著差异[11]。深海和池塘收获的刺参体壁矿物元素如Cr、Mn、Cu含量较为接近,而Zn、B、Al表现出显著差异[12]。本研究滩涂和池塘中的方格星虫体壁Fe、Zn、Mn、Sn、As、Ni、Pb、Cd、Cr、Co等矿物元素之间无显著差异,然而K、Na、Mg、Ca、Cu、Se之间存在显著差异,并且体壁组织中的K、Na、Mg、Ca、Se表现出随着沉积物中相应元素含量增加而升高的趋势,表明方格星虫对以上矿物元素存在一定的富集利用作用。李珂娴等[11]研究发现北海和海口方格星虫体内K、Mg、Ca组成易受生长环境影响,并且饵料研究也发现方格星虫体壁或体腔液中Se、Zn含量随着饲料中的添加量增加而升高[24-25]。Se、Zn等作为重要的微量元素,其含量也是体现营养价值的重要指标,深入研究方格星虫对矿物质元素的富集作用具有重要意义。
星虫类动物体内的矿物组成与生长环境、食物粒径、总有机碳以及代谢特征等有关[26-27]。研究发现方格星虫可以吸收并储存Fe,但是并不会因持续吸收而造成Fe富集[28]。本研究2种养殖环境沉积物中的Fe含量差异显著,但是方格星虫体壁组织中的Fe组成却相近,表明方格星虫不会过度吸收利用Fe元素。以往研究表明,方格星虫体内的Cu、Pb、Cr、Zn组成与周围环境沉积物之间不存在正相关关系[25,29],本研究池塘沉积物中的Fe、Zn、Mn、Ni、Pb、Cr、Co含量显著高于滩涂沉积物,然而2种养殖方式下方格星虫体壁中的以上几种矿物元素含量相近(P>0.05),池塘养殖方格星虫的Cu含量反而低于滩涂养殖,沉积物中矿物元素含量的增加并未造成方格星虫体内的相应矿物元素明显增加,表明星虫对Fe、Zn、Mn、Cu、Ni、Pb、Cr、Co等矿物元素无明显富集作用,也表明这几种矿物元素在星虫体内含量与周围环境沉积物中的元素组成不存在明显的正相关关系。
4. 结论
滩涂和池塘养殖方式对于方格星虫的粗蛋白、粗脂肪等基本营养成分无显著影响,然而对于脂肪酸组成和矿物元素表现出一定程度的影响,不同养殖方式下的有机物质沉积是影响方格星虫营养组成的主要因素。方格星虫对于K、Na、Mg、Ca和Se具有一定的富集作用,而对于Fe、Zn、Mn、Cu、Ni、Pb、Cr、Co无明显富集作用,且方格星虫体壁中的Cu、Zn、Pb、Ni、Cr、Co与周围养殖环境沉积物中的含量不存在明显的正相关关系。本研究表明增养殖环境对方格星虫体内矿物元素组成的影响大于对其脂肪酸和氨基酸组成的影响,优化底栖微藻种类和改进沉积物矿物元素组成可能是提高方格星虫营养价值的重要途径。
-
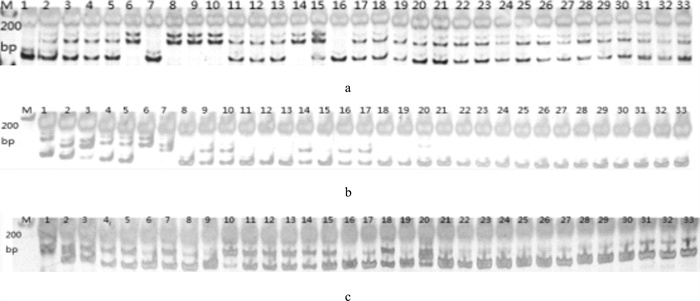
![]()
图 1 尼萨杂交F2、F3群体GM021(a)、UNH906(b)和GM222(c)位点扩增结果
M. DNA标记;1~20. F2;21~33. F3
Figure 1. Amplification results of GM021(a), UNH906(b) and GM222(c) in F2 and F3 hybrid populations of O.niloticus♀×S.melanotheron♂
M. molecular marker; 1~20. F2; 21~33. F3
表 1 7对微卫星引物信息
Table 1 Information of seven pairs of microsatellite primers
位点
locusGenBank登录号
GenBank No.引物序列(5′→3′)
primer sequence重复序列
repeat unit退火温度/℃
annealing temperatureGM258 BV005380 F:CCTTCACCTCCACCACTTTCT
R:AGATCGAACGTCGTCCTCTG(CA)n 64 GNH990 G69270 F:GCCACAGGTGACCATGTTAG
R:GGTGTCTGATTGCACTGACG(TG)n 62 GM222 BV005366 F:AACGGTGACATCTTCGCAACT
R:GATTTGGCTATCTGGCGTGTG(CA)n 62 UNH906 G68220 F:AACATGCTTTCAGCCTTCGT
R:TGAGCAAATCCCGTCCATA(AC)n 56 GM021 BV005272 F:CTGGCTGTGCACAACA
R:TTTGTAAGCAGTCAACACATT(CA)n 59 GM017 BV005269 F:CCCTCTGTTTCCATCTCA
R:GATACCTGTCCATACCTCCTC(CA)n 56 GM145 BV005330 F:AGCCATCCCCGTCTTTCT
R:TATTTTCTGTGAGCCCGTTTG(AC)n 58 注:F.正向引物;R.反向引物
Note:F. the forward primer;R.the reverse primer 下载: 导出CSV
下载: 导出CSV
表 2 尼萨杂交F2、F3微卫星位点的多态性
Table 2 Polymorphism of microsatellite loci of hybrid populations F2 and F3
位点
locus等位基因数(Na)
number of alleles有效等位基因数(Ne)
effective number of alleles观测杂合度(Ho)
observed heterozygosity期望杂合度(He)
expected heterozygosiy多态信息含量(PIC)
polymorphim informationF2 F3 F2 F3 F2 F3 F2 F3 F2 F3 GM258 3 3 2.32 2.17 0.672 0.831 0.592 0.643 0.547 0.498 GNH990 3 3 2.41 2.17 0.439 0.389 0.563 0.334 0.553 0.271 GM017 3 3 2.22 2.06 0.673 0.608 0.557 0.519 0.451 0.501 GM222 2 1 1.97 1.46 0.669 0 0.509 0 0.370 0 UNH906 3 1 2.34 1.00 0.560 0 0.589 0 0.507 0 GM021 3 2 2.66 2.00 0.779 0.611 0.641 0.509 0.542 0.367 GM145 2 2 1.92 1.87 0.613 0.558 0.468 0.413 0.460 0.369 平均mean 2.71 2.14 2.26 1.82 0.632 0.432 0.561 0.340 0.466 0.370
下载: 导出CSV
表 3 尼萨杂交F2、F3中微卫星位点的基因型分布
Table 3 Genotype distribution of seven microsatellite loci of hybrid populations F2 and F3
位点
locusF2 基因型(数目)
F2 Genotype (number)F3 基因型(数目)
F3 Genotype (number)GM258 AA(2)、AB(9)、AC(5)、BC(6) AA(5)、AB(6)、AC(11)、 GNH990 AA(10)、AB(4)、BC(8) AA(10)、AB(7)、AC(5) GM017 AA(4)、AB(11)、BC(3)、BB(4) AA(6)、AB(8)、BC(6)、BB(2) GM222 AA(6)、AB(14)、BB(2) AA(22) UNH906 AA(6)、AB(8)、AC(4)、CC(4) AA(22) GM021 AA(6)、AB(10)、BC(6) AB(22) GM145 AA(11)、AB(11) AA(12)、AB(10)
下载: 导出CSV
表 4 尼萨杂交F2、F3中微卫星位点的等位基因频率
Table 4 Allele frequency in seven microsatellite loci of hybrid populations F2 and F3
位点
locusF2等位基因频率F2 allele frequency F3等位基因频率F3 allele frequency A B C A B C GM258 0.409 0.341 0.250 0.523 0.227 0.250 GNH990 0.545 0.273 0.182 0.727 0.159 0.114 GM017 0.432 0.500 0.068 0.455 0.409 0.136 GM222 0.591 0.409 - 1.000 - - UNH906 0.545 0.182 0.273 1.000 - - GM021 0.500 0.364 0.136 0.500 0.500 - GM145 0.750 0.250 - 0.773 0.227 -
下载: 导出CSV
-
-
期刊类型引用(6)
1. 李俊伟,胡瑞萍,郭永坚,朱长波,陈素文,齐占会,李婷. 方格星虫矿物元素组成及其生物富集特征. 中国海洋药物. 2021(02): 1-9 .  百度学术
百度学术
2. 徐艳,邹杰,谢一兴,蒋艳,孙雪萍,张琴. 广西北海方格星虫脂肪酸的提取及组成分析. 中国海洋药物. 2020(02): 19-30 . 百度学术
3. 陈振国,班庭辉,王明显,梁龙子,陈文清. 不同加工方式沙虫干品的脂肪酸组成比较. 河北渔业. 2020(10): 4-8+40 . 百度学术
4. 林海生,秦小明,章超桦,黄艳球,高加龙,刘琳琳,罗贝,杨发明. 中国沿海主要牡蛎养殖品种的营养品质和风味特征比较分析. 南方水产科学. 2019(02): 110-120 . 本站查看
5. 张顺琦,陈文,王湘君,郭琼琼,钱明,张聪聪. 水提法提取方格星虫粗多糖研究综述. 内江科技. 2019(05): 100-101 . 百度学术
6. 牛改改,游刚,王培,张自然,郭德军,陈文娟. 干燥方法对方格星虫干品质及特征风味的影响. 食品与发酵工业. 2019(23): 128-135 . 百度学术
其他类型引用(3)

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 9
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
