
A preliminary study on biological characteristics of Auxis thazard in the South China Sea
-
摘要:
根据2014年春、秋两季灯光罩网船在南海海域生产调查期间采集的扁舵鲣(Auxis thazard)样本,对其生物学进行初步分析。结果显示,春季扁舵鲣优势叉长为240~300 mm,占总数的60.3%,秋季为220~260 mm,占77.8%;春、秋季样本叉长与体质量关系式差异不大(P>0.05);春、秋季样本总体性比无显著差异,但各叉长组性比有显著差异(P < 0.001);春季样本性腺成熟度各期均有分布,以Ⅰ期和Ⅱ期为主,且随着叉长增大,性成熟的雌鱼比例减小,雄鱼比例上升;不同性腺发育阶段的平均性腺指数(GSI)随着性腺的发育而上升,且雌性大于雄性;初次性成熟叉长为304.5 mm;摄食等级都以0级和1级为主,分别占总数的43.1%和43.4%。
Abstract:We investigated the biological characteristics of Auxis thazard collected from light-falling net fishery in the South China Sea in spring and autumn of 2014. The results show that the dominant fork length ranged from 240 to 300 mm in spring while from 220 to 260 mm in autumn, accounting for 60.3% and 77.8% of the total number, respectively. The relationship between fork length and body weight had little difference in spring and autumn. Significant difference was not found in total sex ratio in spring and autumn (P>0.05), but was found in fork length groups in spring and autumn (P < 0.001). StageⅡ had the maximum maturity in spring. The proportion of mature females decreased and mature males increased with increasing fork length. With the development of gonad, the average GSI values increased at different gonadal stages and females had higher GSI values than males. The fork length of the first gonadal maturity was 304.5 mm. Stage 0 and Stage 1 had the highest feeding intensity, accounting for 43.1% and 43.4% of the total number, respectively.
-
Keywords:
- Auxis thazard /
- the South China Sea /
- biological characteristics
-
华贵栉孔扇贝(Chlamys nobilis)属于热带海水贝类,具有肉质鲜美、营养价值高等优点,自20世纪70年代起,有关其人工育苗和规模化养殖迅速在南方各沿海省份展开,并取得了较好的经济效益,但由于长期养殖,华贵栉孔扇贝也面临种质质量退化的问题,严重影响了该产业的健康发展。目前相关机构已经对其开展了选育研究,并取得了初步成效。数量遗传学作为一种新型研究遗传育种的学科,在水产动物遗传育种中展现了很大的优越性,但将此方法运用于华贵栉孔扇贝良种选育的相关研究尚不多见。该研究在此方向作了初步探索,解决华贵栉孔扇贝养殖产业健康发展的关键所在是建立有效的良种选育方法[3-4]。
三亚崖州湾海区是海南省华贵栉孔扇贝主养区之一,已有5年养殖经历。笔者通过对该海区养殖的华贵栉孔扇贝数量性状的测量和分析,采用多元分析和回归方法,建立形态性状与质量性状间的多元线性回归方程[5],以期为华贵栉孔扇贝通过数量性状辅助选择育种提供相关理论依据。
1. 材料与方法
1.1 材料
受测的华贵栉孔扇贝来自于三亚崖州湾养殖海区,由三亚意源养殖有限公司提供。
1.2 取样和测量方法
2010年3月30日一次性随机选取各年龄阶段526个扇贝标本测量其数量性状。测量前清除其表面附着物。5个形态性状(壳高、壳长、绞合线长、壳厚和贝柱直径)用数显游标卡尺测量(精度为0.02 mm),6个质量性状(总质量、壳质量、贝柱鲜质量、鲜肉质量、干肉质量和贝柱干质量)用电子天平测量(精度为0.01 g)。鲜肉质量指活体去壳后含水分的质量,干肉质量指将鲜肉放入烘箱去除水分后的质量。
1.3 数据分析方法
测量的数据使用SPSS 13软件进行数据分析,对数据采用t检验,用多元回归分析法,分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝3个经济性状[鲜肉质量(y1)、肉柱鲜质量(y2)和肉柱干质量(y3)]的影响,同时作通径分析。
2. 结果与分析
2.1 各性状的参数分析
华贵栉孔扇贝壳高、壳长、绞合线长、壳厚、贝柱直径、总质量、壳质量、鲜肉质量、贝柱鲜质量、干肉质量和贝柱干质量等11个数量性状的各种参数见表 1。其中不同性状的参数单位有所不同,不能进行性状间比较。变异系数的单位相同,因而可以进行性状间比较。性状的变异系数越大,表明品种相应指标可供选择的范围越大,选择的潜力就越大。11个性状的变异系数由大到小依次为贝柱干质量>干肉质量>贝柱鲜质量>鲜肉质量>总质量>壳质量>贝柱直径>壳厚>绞合线长>壳长>壳高。因此,在华贵栉孔扇贝高产品种培育中对这些性状进行选择时,应首先考虑贝柱干质量,其次是干肉质量、贝柱鲜质量、鲜肉质量、总质量、壳质量、贝柱直径、壳厚、绞合线长和壳长,最后是壳宽。
表 1 各性状的表型统计量(n=526)Table 1. Phenotypic statistics of various quantitative traits参数parameter 壳高/mm SH 壳长/mm SL 绞合线长/mm HL 壳厚/mm ST 贝柱直径/mm CP 总质量/g TW 壳质量/g WWAM 鲜肉质量/g FW 贝柱鲜质量/g CFW 干肉质量/g DFW 贝柱干质量/g DWAM max 76.00 72.30 42.00 26.30 23.10 54.01 29.93 24.74 5.32 3.66 1.30 min 44.10 41.00 21.90 13.10 8.90 10.23 7.28 1.33 0.55 0.40 0.07 mean 60.52 55.57 32.58 18.97 15.83 27.86 17.07 10.79 2.40 1.46 0.53 R 31.90 31.30 20.10 13.20 14.20 43.78 22.65 23.41 4.77 3.26 1.23 SD 5.52 5.44 3.32 2.04 2.46 7.08 4.17 3.29 0.87 0.57 0.22 SE 0.24 0.24 0.14 0.09 0.11 0.31 0.18 0.14 0.04 0.02 0.01 CV 9.12% 9.80% 10.19% 10.77% 15.56% 25.43% 24.42% 30.51% 36.31% 38.99% 42.19% 2.2 5个数量性状的回归分析及通径分析
2.2.1 鲜肉质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝鲜肉质量(y)的影响(表 2)。总质量与鲜肉质量为极显著的正相关(相关系数为0.839 4,P < 0.01),壳厚与贝柱鲜质量为显著的负相关(相关系数为-0.110 7,0.01 < P < 0.05)。y=1.295-0.033 15x1-0.091 6x4+0.474 9x5,决定系数R2=0.878 563,剩余标准差SSE=1.150 2。
表 2 5个数量性状对鲜肉质量的逐步回归分析Table 2. Stepwise regression analysis between 5 quantitative traits and shell-free weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x1 -0.033 1 -0.055 5 -0.092 8 2.126 9 0.033 9 x4 -0.091 6 -0.056 8 -0.107 4 2.464 6 0.014 0 x5 0.474 9 1.022 5 0.839 4 35.246 5 0.000 0 通径系数反映自变量对依变量的直接影响,总质量对鲜肉质量的影响最为明显,直接系数为1.024 9(表 3)。x5与鲜肉质量的相关系数最大,而x1和x4与鲜肉质量呈负相关,x5与鲜肉质量呈正相关。
表 3 5个数量性状对鲜肉质量的通径分析Table 3. Path analysis between 5 quantitative traits and shell-free weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x1 -0.055 5 - -0.037 8 0.822 6 0.729 3 x4 -0.056 8 -0.036 9 - 0.757 1 0.663 4 x5 1.024 9 -0.044 7 -0.042 1 - 0.935 7 2.2.2 贝柱鲜质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝贝柱鲜质量(y)的影响(表 4),总质量与贝柱鲜质量呈极显著的正相关(相关系数为0.622 9,P < 0.01),绞合线长与贝柱鲜质量呈极显著的负相关(相关系数为-0.196 6,P < 0.01)。y=-0.168 8+0.027 34x2-0.046 04x3+0.091 4x5,决定系数R2=0.617 847,剩余标准差SSE=0.539 5。
表 4 5个数量性状对贝柱鲜质量的逐步回归分析Table 4. Stepwise regression analysis between 5 quantitative traits and adductor muscle weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x2 0.027 3 0.171 0 0.154 5 3.569 8 0.000 4 x3 -0.046 0 -0.175 7 -0.196 6 4.576 6 0.000 0 x5 0.091 4 0.744 1 0.629 9 18.513 1 0.000 0 从表 5中可以看出,通径系数反映自变量对依变量的直接影响。结果显示,总质量对贝柱鲜质量的影响最为明显,直接系数为0.744 1;x2和x5与贝柱鲜质量呈正相关,x3与贝柱鲜质量呈负相关; x5的总体影响最大(0.775 4),x2其次(0.596 0)。
表 5 5个数量性状对贝柱鲜质量的通径分析Table 5. Path analysis between 5 quantitative traits and adductor muscle weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x2 0.171 0 - -0.124 4 0.549 5 0.596 0 x3 -0.175 7 0.121 1 - 0.402 1 0.347 5 x5 0.744 1 0.126 3 -0.094 9 - 0.775 4 2.2.3 贝柱干质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝贝柱干质量(y)的影响(表 6)。总质量与贝柱干质量为极显著的正相关(相关系数为0.743 0,P < 0.01),壳长与贝柱干质量为显著的负相关(相关系数为-0.092 2,P < 0.05),绞合线长与贝柱干质量为极显著的负相关(相关系数为-0.141 1,P < 0.01)。y=0.228 7-0.005 0x2-0.007 8x3+0.029 9x5,决定系数R2=0.664 705,剩余标准差SSE=0.129 1。
表 6 5个数量性状对贝柱干质量的逐步回归分析Table 6. Stepwise regression analysis between 5 quantitative traits and adductor muscle dry weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x2 -0.005 0 -0.123 7 -0.119 9 2.756 7 0.006 0 x3 -0.007 8 -0.117 0 -0.141 1 3.253 9 0.001 2 x5 0.029 9 0.954 0 0.743 0 25.341 1 0.000 0 从表 7可以看出,通径系数反映自变量对依变量的直接影响。结果显示,总质量对贝柱鲜质量的影响最为明显,直接系数为0.954 0。
表 7 5个数量性状对贝柱干质量的通径分析Table 7. Path analysis between 5 quantitative traits and adductor muscle dry weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x2 -0.123 7 - -0.082 9 0.704 5 0.497 9 x3 -0.117 0 -0.087 6 - 0.515 5 0.310 9 x5 0.954 0 -0.091 3 -0.063 2 - 0.799 4 x5与贝柱干质量呈正相关,x2和x3与贝柱干质量呈负相关,x5对贝柱干质量产生的影响最为明显。
3. 讨论
3.1 通径分析的运用
性状间的相关系数是进行多元分析的基础,但相关系数是两变量间关系的综合体现,其既包括变量间的直接关系,又包括通过其他变量影响的间接关系。单纯采用性状间的相关系数不能正确表述两性状间的真实关系。通径分析可用于分析多个自变量与变量之间的线性关系,是回归分析的拓展,不仅可以处理较为复杂的变量关系,而且能把性状间相关性分成直接影响和间接影响[6-8]。该研究中华贵栉孔扇贝壳高、壳长、绞合线长、壳厚和总质量与扇贝的鲜肉质量、贝柱鲜质量及贝柱干质量的相关系数均极显著,建立回归方程时将其余几个对经济性状相关系数影响不显著的予以剔除以建立理想的回归方程。
3.2 数量性状的选育
衡量华贵栉孔扇贝生长状况的数量性状一般包括长度(高度)和质量,该研究选取长度和质量2个性状来探讨华贵栉孔扇贝数量性状间的相关性。以相关形态性状和总质量为自变量,对3个主要经济性状的回归相关性极显著。把简单易测的性状作为自变量是符合期望的[9-11]。因此,在华贵栉孔扇贝的遗传育种中把壳长和总质量作为重要生长指标即可。通径分析在水产动物的遗传育种中运用非常广泛[12-16],如王爱民等[17]对马氏珠母贝(Pinctada martensii)不同地理种群内自繁和种群间杂交子一代形态性状参数进行了相关分析,郑怀平等[18]对近江牡蛎(Ostrea rivularis gould)养殖群体数量性状间的相关性进行了通径分析,喻子牛等[19]对香港巨牡蛎(Crassostrea hongkongensis)雌雄群体数量性状进行了通径分析。
3.3 选育对加工产业的意义
选育优质的良种对华贵栉孔扇贝加工产业具有相当重要的意义。加工企业需用扇贝的贝柱来加工产品,因而对贝柱的质量格外关注。该研究中对扇贝贝柱鲜质量和贝柱干质量均作了相关分析[20]与通径分析,研究结果显示,总质量对于3个经济性状的正面影响最大。因此,在高产选育时应选择总质量大的品种来进行选育,使获得的优良品种能够产生大的贝柱。
-
![]()
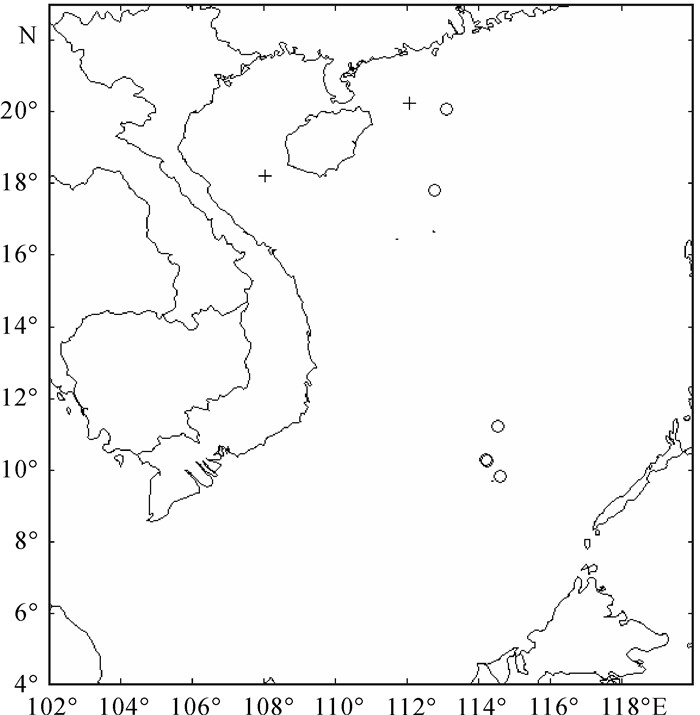
图 1 调查站位图
○表示春季调查站位,+表示秋季调查站位
Figure 1. Investigation stations
○. stations in spring; +. stations in autumn
![]()
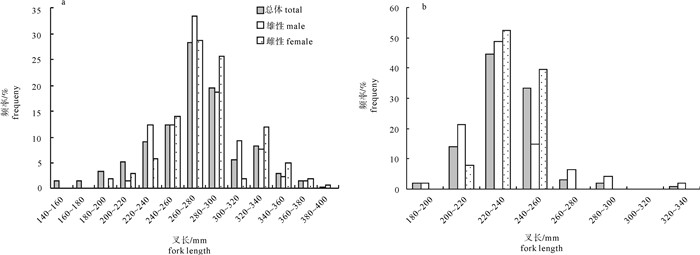
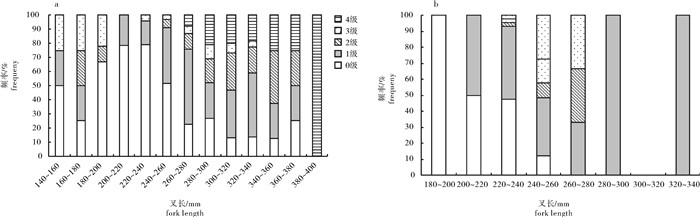
图 2 春季(a)和秋季(b)扁舵鲣叉长分布
Figure 2. Distribution of fork length of A.thazard in spring (a) and autumn (b)
![]()
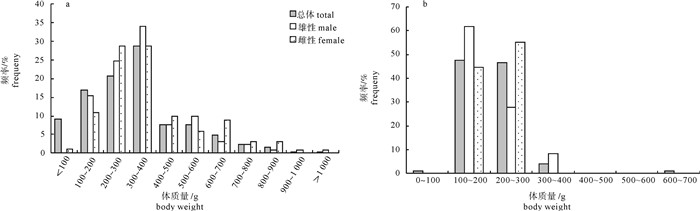
图 3 春季(a)和秋季(b)扁舵鲣体质量分布
Figure 3. Distribution of body weight of A.thazard in spring (a) and autumn (b)
![]()
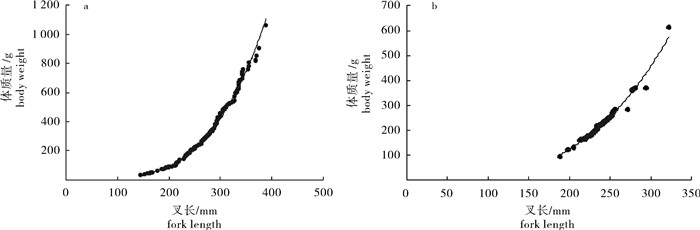
图 4 春季(a)和秋季(b)季扁舵鲣体质量与叉长关系
Figure 4. Relationship between body weight and fork length of A.thazard in spring (a) and autumn (b)
![]()
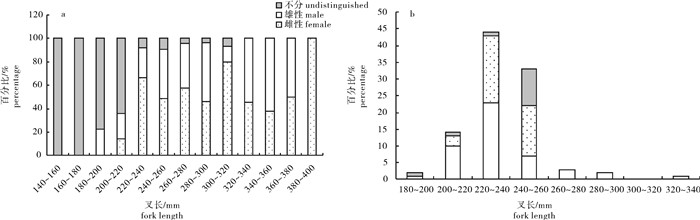
图 5 春季(a)和秋季(b)扁舵鲣性比与叉长关系
Figure 5. Relationship between sex ratio and fork length of of A.thazard in spring (a) and autumn (b)
![]()
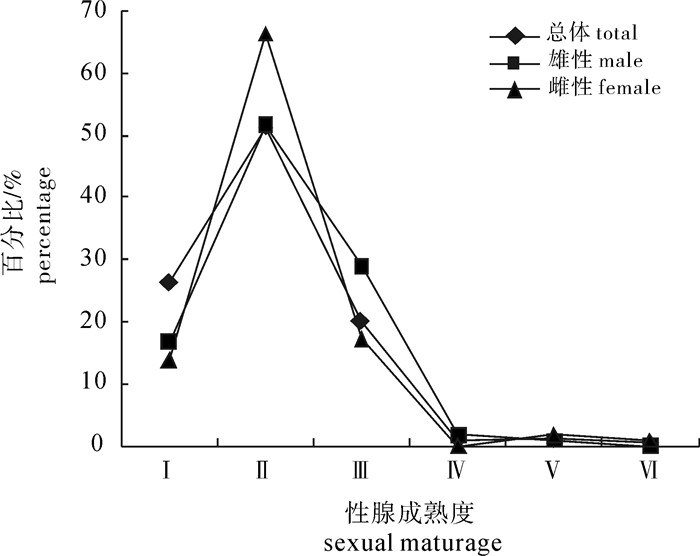
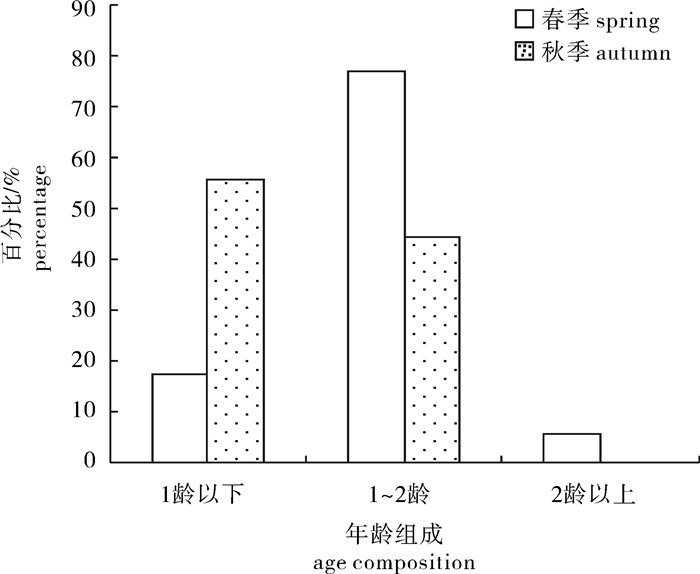
图 6 春季扁舵鲣性腺成熟度百分比组成
Figure 6. Percentage of gonad developmental stages of A.thazard in spring
![]()
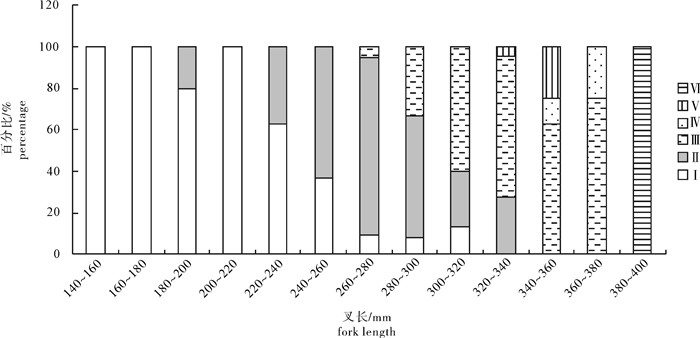
图 7 春季扁舵鲣不同叉长组性腺成熟度百分比组成
Figure 7. Percentage of gonad developmental stages in different fork-length groups of A.thazard in spring
![]()
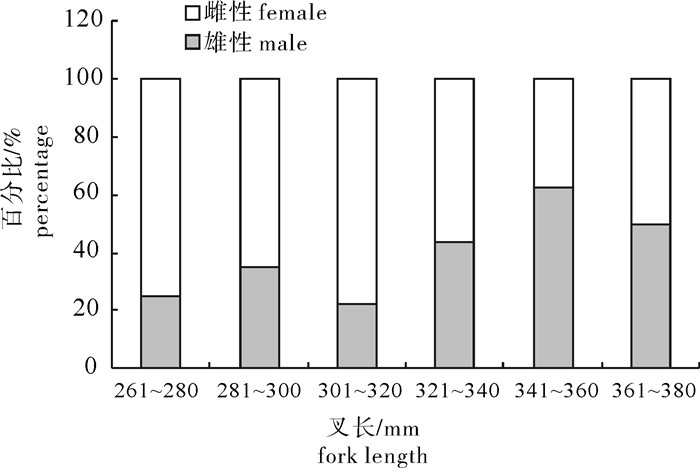
图 8 春季扁舵鲣成熟个体性比与叉长关系
Figure 8. Relationship between sex ratio and fork length of A.thazard in spring
![]()
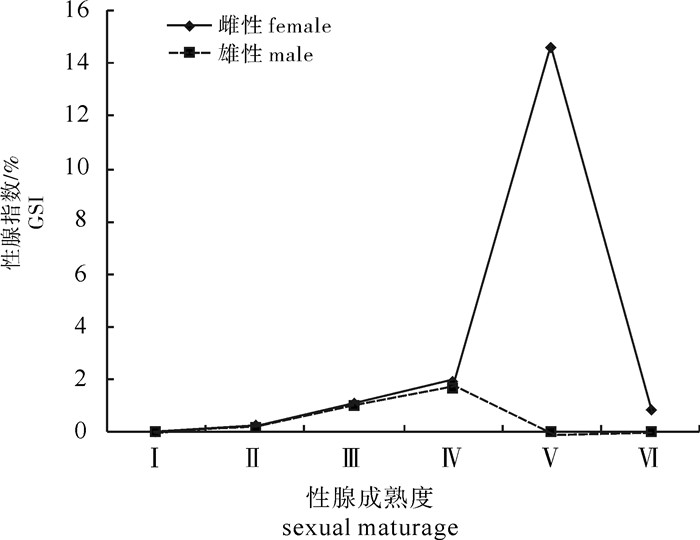
图 9 春季不同性腺发育阶段扁舵鲣的平均性腺指数
Figure 9. Average GSI values of A.thazard at different gonadal stages in spring
![]()
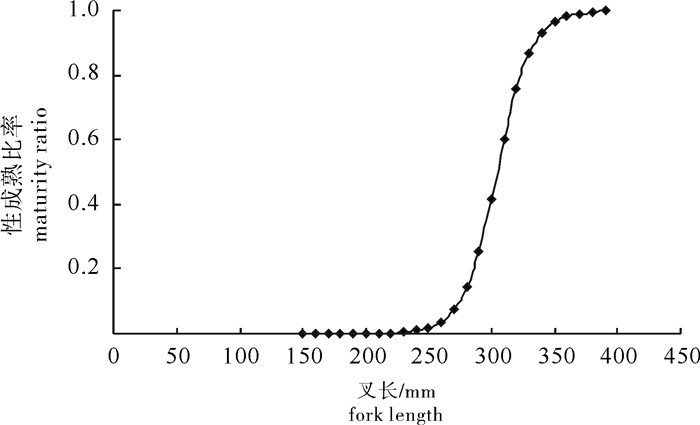
图 10 春季扁舵鲣性成熟与叉长关系
Figure 10. Ralationship between sexual maturity and fork length of A.thazard in spring
![]()
图 11 南海海域春季(a)和秋季(b)季扁舵鲣摄食等级与叉长关系
Figure 11. Relationship between feeding stage and fork length of A.thazard in the South China Sea in spring (a) and autumn (b)
表 1 不同研究所得的叉长-体质量关系
Table 1 Comparison of relationship between fork length and body weight by different studies
研究区域
region时间
time渔具
gear叉长-体质量关系
fork length-body weight relationship文献
Reference台湾海峡及其邻近海域Taiwan Strait and its adjacent sea 1981年5~8月,1982年4~7月,1983年4~9月,1984年4~6月 大围缯 W=1.658×10-6L3.405 5 卢振彬等[14],1991 台湾海峡Taiwan Strait 2005年4月~2006年3月 围网 W=0.1815×10-5L3.389 9 YU等[13],2012 南海the South China Sea 春季 灯光罩网 WS=3×10-7L3.691 3 此研究 南海the South China Sea 秋季 灯光罩网 WA=5×10-6L3.202 此研究  下载: 导出CSV
下载: 导出CSV
-
[1] 马超, 沈长春, 刘勇, 等. 钓鱼岛北部海域光诱敷网渔业资源调查[J]. 福建水产, 2014, 36(1): 55-61. doi: 10.3969/j.issn.1006-5601.2014.01.008 [2] 张鹏, 杨吝, 张旭峰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. doi: 10.3969/j.issn.1673-2227.2010.01.012 [3] 刘佩妤. 以台湾近海鲔延绳钓渔业大目鲔体长数据估计其年龄、成长与死亡率以及单位加入量分析[D]. 台北: 国立台湾大学海洋研究所, 2005. [4] 苏楠杰. 台湾近海金枪鱼延绳钓渔业黄鳍金枪鱼的年龄、成长、死亡率与单位加入生产量[D]. 台北: 国立台湾大学海洋研究所, 2003. [5] LANGLEY A, HAMPTON J, KLEIBER P. Stock assessment of yellowfin tuna in the western and central Pacific Ocean, including an analysis of management options [ C/OL]//Scientific Committee Third Regular Session, August 2007, Honolulu, USA. [ 2007-12-07]. http://www.wcpfc.int/system/files/documents/meeting-s/scientific-committee/3rd-regular-session/stock-assess-ment-swg-w-orking-papers/WCPFC-SC3%20SA-SWG%20WP-01.pdf.
[6] LANGLEY A, HAMPTONL J, KLEIBER P, et al. Stock assessment of bigeye tuna in the western and central Pacific Ocean, including an analysis of management options[C/OL]//Scientific Committee Fourth Regular Session, August 2008, Papua New Guinea. [2008-07-31]. http://www.wcpfc.int/system/files/doc-uments/meetings/scientific-committee/4th-regular-session/stock-assessment-specialist-working-gro-/SC4-SA-WP1-rev.1%20%5B-BET%20Assessment%D.pdf.
[7] GHOSH, PILLAI N, DHOKIA H. Fishery population characteristics and yield estimates of coastal tunas at Veraval [J]. Ind J Fish, 2010, 57(2): 7-13. doi: 10.1093/icesjms/fsp251
[8] ABDUSSAMAD E, RAO G, KOYA K, et al. Indian tuna fishery-production trend during yesteryears and scope for the future[J]. Ind J Fish, 2012, 59(3): 1-13. doi: 10.1051/alr/2012034
[9] 张鹏, 曾晓光, 杨吝, 等. 南海区大型灯光罩网渔场渔期和渔获组成分析[J]. 南方水产科学, 2013, 9(3): 74-79. doi: 10.3969/j.issn.2095-0780.2013.03.012 [10] 颜云榕, 冯波, 卢伙胜. 中、西沙海域2种灯光作业渔船的捕捞特性及其技术效率分析[J]. 南方水产, 2009, 5(6) : 59 -64. doi: 10.3969/j.issn.1673-2227.2009.06.011 [11] 刘维, 张羽翔, 陈积明, 等. 南沙群岛春季灯光围网渔业资源调查初步分析[J]. 上海海洋大学学报, 2012, 21(1): 105-109. https://xueshu.baidu.com/usercenter/paper/show?paperid=a825015b67dd3d8e92ef97ac4e2c2f97&site=xueshu_se&hitarticle=1 [12] 卢振彬, 戴泉水, 颜尤明. 台湾海峡及其邻近海区扁舵鲣的生长和死亡[J]. 水产学报, 1991, 15(3): 228-234. [13] YU Tao, CHEN Mingru, DU Jianguo, et al. Age and growth changes and population dynamics of the black pomfret (Parastromateus niger) and the frigate tuna (Auxis thazard thazard), in the Taiwan Strait[J]. Lat Am J Aquat Res, 2012, 40(3): 649-656. doi: 10.3856/vol40-issue3-fulltext-13
[14] 卢振彬, 颜尤明, 戴泉水. 闽中、闽东渔场扁舵鲣的资源生物学[J]. 台湾海峡, 1992, 11(3): 251-256. https://xueshu.baidu.com/usercenter/paper/show?paperid=1138965dc1744661fc3c7d8972f741ed&site=xueshu_se&hitarticle=1 [15] JUDE D, NEETHISELVAN N, GOPALAKRISHNAN P, et al. Gill net selectivity studies for fishing frigate tuna, Auxis thazard Lacepede (Perciformes: Scombridae) in Thoothukkudi waters, southeast coast of India[J]. Ind J Mar Sci, 2002, 31(4): 329-333. https://www.researchgate.net/publication/237378359_Gill_net_selectivity_studies_for_fishing_frigate_tuna_Auxis_thazard_Lacepede_PerciformesScombridae_in_ThoothukkudiTuticorinwaters_southeast_coast_of_India
[16] 张仁斋. 三种金枪鱼类(鲣、黄鳍金枪鱼、扁舵鲣)的仔、稚鱼在南海的分布和产卵期[J]. 海洋学报: 中文版, 1983, 5(3): 368-375. [17] 童铃, 金毅, 徐坤华, 戴志远. 3种鲣鱼背部肌肉的营养成分分析及评价[J]. 南方水产科学, 2014, 10(5): 51-59. doi: 10.3969/j.issn.2095-0780.2014.05.008 [18] MANAT C, SAPPASITH K, SOOTTAWAT B. Characterisation of muscles from Frigate mackerel (Auxis thazard) and catfish (Clarias macrocephalus)[J]. Food Chem, 2013, 139(1/2/3/4): 414-419. doi: 10.1016/j.foodchem.2013.01.111
[19] IVANE R P, ALTAIR B A, MUDJEKEEWIS D S. Genetic diversity, population genetic structure, and demographic history of Auxis thazard (Perciformes), Selar crumenophthalmus (Perciformes), Rastrelliger kanagurta (Perciformes) and Sardinella lemuru (Clupeiformes) in Sulu-Celebes Sea inferred by mitochondrial DNA sequences[J]. Fish Res, 2015, 162: 64-74. doi: 10.1016/j.fishres.2014.10.006
[20] 国家海洋局. 海洋调查规范[S]. 北京: 中国标准出版社, 1999. https://xueshu.baidu.com/usercenter/paper/show?paperid=9f80a0159f9b1bcf96cac50a54537592&site=xueshu_se [21] 苏锦祥. 鱼类学与海水鱼类养殖[M]. 北京: 中国农业出版社, 1995: 300-306. https://xueshu.baidu.com/usercenter/paper/show?paperid=3b37cde2fd2853421f903f62b74dd6c8&site=xueshu_se [22] 李建生, 严利平, 胡芬, 等. 温台渔场日本鲭的繁殖生物学特征[J]. 中国水产科学, 2015, 22(1): 99-105. doi: 10.3724/SP.J.1118.2015.00112 [23] BAGENAL T B, TESCH E W. Age and growth[M]//BAGENAL T. Methods for assessment of fish in fresh-waters. Oxford: Black well Scientific Publications, 1978: 101-136. https://xueshu.baidu.com/usercenter/paper/show?paperid=9dc49e89c6b73e096cd63574d6339524&site=xueshu_se
[24] 黄真理, 常剑波. 鱼类体长与体重关系中的分形特征[J]. 水生生物学报, 1999, 23(4): 330-337. doi: 10.3321/j.issn:1000-3207.1999.04.006 [25] 刘岩, 张秀梅, 周游, 等. 2011年西南大西洋拖网渔获物阿根廷滑柔鱼生物学分析[J]. 南方水产科学, 2012, 8(3): 39-47. doi: 10.3969/j.issn.2095-0780.2012.03.006 [26] 龚彩霞, 陈新军, 刘必林, 等. 西南大西洋公海阿根廷滑柔鱼生物学特性的初步研究[J]. 大连海洋大学学报, 2010, 25(4): 353-358. doi: 10.3969/j.issn.1000-9957.2010.04.013 [27] 殷名称. 鱼类生态学[M]. 北京: 中国农业出版社, 2000: 34-38. https://xueshu.baidu.com/usercenter/paper/show?paperid=57ce72a57694feba4d0f887226ad1faf&site=xueshu_se [28] 江素菲, 王德明. 闽南-台湾浅滩渔场金枪鱼类仔鱼的分布和产卵期[J]. 厦门大学学报: 自然科学版, 1986, 25(4): 476-483. doi: 10.3321/j.issn:0438-0479.1986.04.015 [29] 赵传絪, 陈莲芳, 臧增嘉. 东海舵鲣的早期发育和生殖习性[J]. 水产学报, 1982, 6(3): 253-267. https://xueshu.baidu.com/usercenter/paper/show?paperid=1ee8fd39fad08ed2ed1556662a0de2bb&site=xueshu_se&hitarticle=1 -
期刊类型引用(3)
1. 周星星,范江涛,于杰,徐姗楠,蔡研聪,陈作志. 基于地统计学的南海扁舵鲣时空分布研究. 南方水产科学. 2022(05): 153-159 .  本站查看
本站查看
2. 粟丽,陈作志,张鹏,李杰,王欢欢,黄佳兴. 2017年南海中南部渔场灯光罩网渔获物组成及渔获率时空分布. 南方水产科学. 2018(05): 11-20 . 本站查看
3. 张俊,张鹏,陈作志,陈国宝,张魁,许友伟,孙铭帅. 南海外海鲹科鱼类资源量及其分布. 南方水产科学. 2016(04): 38-48 . 本站查看
其他类型引用(0)
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 3
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
