
Characteristics of macrozoobenthic community structure in typical wetlands of Macao
-
摘要:
于2012年12月~2013年9月对澳门4个典型湿地的大型底栖动物群落结构进行了研究。调查共鉴定出底栖动物66种,其中环节动物9种、软体动物45种、甲壳动物8种、鱼类3种。底栖动物种类组成具有河口低盐种、半咸水种和淡水种共存的特点;从物种生活型组成来看,底内型占据优势;从功能群组成来看,浮游生物食性和植食性底栖动物占据优势。最多的物种数出现在莲花大桥滩涂(46种),物种数于采样点间差异显著。沼蛤(Limnoperna fortunei)、多棱角螺(Angulyagra polyzonata)、梨形环棱螺(Bellamya purificat)、羽须鳃沙蚕(Dendronereis pinnaticirris)、谭氏泥蟹(Ilyoplax deschampsi)和纹斑棱蛤(Trapezium liratum)是主要的优势种。底栖动物密度呈现出秋季>夏季>冬季>春季的季节变化,生物量呈现出秋季>春季>冬季>夏季的季节变化。多样性分析结果表明,莲花大桥滩涂的物种多样性最高,多样性指数于采样点和季节均无显著差异。
Abstract:We conducted a seasonal investigation on the community structure of macrobenthos in four typical wetlands in Macao from December 2012 to September 2013. A total of 66 species were identified, including 45 species of molluscs, 9 polychaetes, 8 crustaceans and 3 fishes. The macrobenthic species composition in wetlands was characteristized by brackish water species, freshwater species and estuarial low salinity species. Bottom-type species dominated in the life form composition, herbivorous and plankton feeders dominated in functional feeding composition. The species occurred in Lotus Flower Bridge Flat (46 species) most. Two-Way analysis of variance show that there was significant difference in species number among different seasons. Limnoperna fortunei, Angulyagra polyzonata, Bellamya purificat, Dendronereis pinnaticirris, Ilyoplax deschampsi and Trapezium liratum were the main dominant species. The descending order of macrobenthos density was autumn > summer>winter>spring. The descending order of biomass was autumn>spring>winter>summer. Shannon-Wiener species diversity and Pielou evenness indices were the highest in Lotus Flower Bridge Flat, but no significant difference was detected in the two indices among different seasons and sampling sites.
-
Keywords:
- macrobenthos /
- community characteristics /
- biodiversity /
- wetland /
- Macao
-
凡纳滨对虾(Litopenaeus vannamei)又称南美白对虾,主要分布于加利福尼亚湾至秘鲁北部的太平洋沿岸水域,是当今世界上产量最高的养殖对虾品种之一[1]。良好的养殖水环境是凡纳滨对虾养殖成功的保障,氨氮(NH3-N)和溶解氧(DO)是对虾养殖水体环境中的重要理化因子,特别是工厂化高密度养殖模式下最值得关注的2个水质理化因子[2]。中国就凡纳滨对虾仔虾、幼虾、成虾阶段对氨氮的耐受性已有报道[3-5];对低溶氧胁迫的研究主要集中在行为、生理、免疫等方面[6-7]。有关凡纳滨对虾高氨氮、低溶氧抗逆性状遗传育种的研究少见报道。氨氮、溶解氧等胁迫因子对养殖凡纳滨对虾的遗传基础、抗逆性以及病原微生物的致病力均有较显著的影响。深入了解凡纳滨对虾高氨氮、低溶氧抗逆性的遗传性能,可为凡纳滨对虾抗逆遗传改良奠定基础。
配合力是筛选和评估人工选育亲本的重要遗传参数,最早应用于玉米杂交育种[9]研究中。配合力是指亲本(纯系、自交系或品种)材料在由其所产生的杂种后代的产量或其他性状表现中所起作用相对大小的度量[8],包括一般配合力和特殊配合力。一般配合力是由于基因加性效应所致,而特殊配合力主要是双亲杂交后通过互作表现出的非加性效应,即基因的显性、超显性和上位效应[10]。在水产动物育种研究中,吴杨平等[11]研究了红壳色文蛤(Meretrix meretrix)3个地理群体在早期发育阶段的壳长性状配合力,发现了红壳色文蛤最具优势的杂交组合。MALLET等[12]分析了野生牡蛎3个群体的生长和存活性状配合力,结果显示牡蛎早期发育阶段的生长和成活率受母体效应的影响较大。季士志等[13]通过双列杂交设计,研究了2个大菱鲆(Scophthalmus maximus)养殖群体的杂交效果,结果表明不同养殖群体间的杂交有可能是大菱鲆遗传改良的有效途径。林壮柄等[14]报道了杂色鲍(Haliotis diversicolor)双列杂交的杂种优势。国内外均有关于凡纳滨对虾不同群体配合力分析及杂种优势利用的研究报道[15-16],但关于群体间抗逆性状配合力分析的研究尚未见报道。该研究利用3个地理群体的凡纳滨对虾,采用Griffing双列杂交育种法Ⅰ,建立9个自繁和杂交组合F1代,比较其对高氨氮、低溶解氧的耐受性,分析交配组合杂种优势,进一步采用线性模型均数加权二乘分析法估算凡纳滨对虾各理化因子耐受性状的配合力,评估亲本和子代对各理化因子的耐受性能,为进一步家系选育提供候选材料和抗逆凡纳滨对虾新品系选育提供理论依据。
1. 材料与方法
1.1 实验材料
3个引进的凡纳滨对虾群体于2013年4月~6月分批相继运抵海威水产养殖有限公司东海对虾良种场(广东海洋大学产学研基地)。3个凡纳滨对虾引进群体包括美国迈阿密SIS群体、美国夏威夷SIS群体和泰国正大群体,分别命名为UM、UH和TZ群体。亲虾暂养稳定后,雌虾进行单侧眼柄切除,投喂冰冻鱿鱼、牡蛎和鲜活沙蚕进行促熟。选取性腺成熟的亲虾,通过完全双列杂交设计,成功建立了9个自繁和杂交群体,具体交配信息见表 1。每个交配组合分别有9~21尾雌虾成功孵出幼体,合计128个全同胞家系。采用标准化的方式对每个自交和杂交群体进行幼体培育直至仔虾。尽量保持各阶段的培育条件一致,以减少环境条件(主要包括各个阶段水的盐度、温度、幼体密度、饵料及充气等条件)差异对生长发育的影响。仔虾体长生长到3~4 cm时对9个群体进行凡纳滨对虾高氨氮和低溶氧的耐受性比较。
表 1 凡纳滨对虾3个引进群体的完全双列杂交Table 1. Complete diallel cross of three introduced populations of L.vannamei引进群体(♀)
introduced population引进群体(♂)
introduced populationUM UH TZ 美国迈阿密SIS群体UM UM×UM UM×UH UM×TZ 美国夏威夷SIS群体UH UH×UM UH×UH UH×TZ 泰国正大群体TZ TZ×UM TZ×UH TZ×TZ 1.2 耐受性试验设计
根据相关文献[17-18]设定氨氮和溶解氧梯度,进行胁迫因子的48 h耐受性预试验,得出48 h半致死温度、盐度值,即正式试验中高氨氮和低溶氧胁迫浓度值,分别为98 mg · L-1和0.7~0.9 mg ·L-1。采用单因素试验设计,每个群体挑选大小相近的凡纳滨对虾个体30尾,置于14 cm×12 cm×12 cm的小网箱中,每组的9个网箱放在同一个1.6 m×1.2 m×0.4 m的泡沫箱中。实验设3个平行组和1个对照组(对照组48 h内均全部成活),共计72个小网箱和8个泡沫箱。每隔12 h观察凡纳滨对虾的活力,当对虾停止任何运动,用玻棒触及无反应,则视为死亡[12],及时取出死亡个体。胁迫因子的调控在泡沫箱中完成,每12 h换水一次,清理粪便、蜕皮及死亡个体,实验持续48 h,实验结束时统计成活率。
1.3 理化因子的控制
高氨氮和低溶解氧等水环境全部通过人工48 h不间断调节控制,主要调控方法:1)氨氮控制系统。实验溶液以NH4ClA.R.溶于过滤海水中制得,使用5B-2N轻巧便携型氨氮测定仪随时监控实验组水体中氨氮含量;2)低溶解氧控制系统。通过向泡沫箱水体中充入不同的空气和氮气的比例来调节。供气系统由氮气罐、氧气罐、气阀、输气管和气石等组成。溶解氧用Mi605便携式溶解氧测试仪直接快速测量,随时监控试验组水体溶解氧。
1.4 数据处理
利用Excel 2007软件对成活率数据进行初步整理,用SPSS 19.0软件对成活率数据进行平方根反正弦转换后作单因素方差分析,多重比较法分析不同群体间的差异显著性。杂种优势(Heterosis)参照CRUZ等[19]的方法计算:
$$ \begin{aligned} & M P(\%)=\frac{F_1-P_0}{P_0} \times 100 \\ & B P(\%)=\frac{F_1-P_B}{P_B} \times 100 \end{aligned} $$ 其中MP是中亲优势;F1为杂交组均值;P0是相应双亲均值;BP为超亲优势;PB是杂交组较好的亲本均值。
配合力分析统计模型[20]为:
$$ Y_{i j k}=u+S_i+D_j+S D_{i j}+E_{i j k} $$ 式中Yijk代表第i个父本群体与第j个母本群体的第k个杂种个体的表型观测值,u表示总体均数,Si、Dj分别为第i个父本和第j个母本一般配合力效应,SDij为第i个父本与第j个母本的互作效应即正交特殊配合力效应,Eijk表示随机误差效应,并假设其独立地服从N(0,σe2)分布。整个群体的变异可以剖分为一般配合力(GCA)、特殊配合力(SCA)、随机误差等组成部分。模型采用二因素有互作的均数加权最小二乘分析法进行配合力分析。其正规方程为(X′X)b=X′Y
其约束条件为$ \sum_i \hat{S}=\sum_j \hat{D}=\sum_i \hat{S} D=\sum_j \hat{S} D=\sum \sum_{i j} \hat{S} D=0$
2. 结果与分析
2.1 急性胁迫耐受性比较
F1代9个群体幼虾各理化因子48 h急性胁迫耐受成活率见表 2。9个群体低溶氧48 h急性胁迫耐受成活率为15.56%~84.45%,其中TZ×UM群体成活率最低,UM×UH群体成活率最高。UH×TZ群体低盐48 h急性胁迫耐受成活率最低[(16.67±1.93)%],UM×UH群体高氨氮48 h急性胁迫耐受成活率最高[(64.44±2.39)%]。各交配组合耐低溶氧和高氨氮成活率单因素方差分析多重比较,差异均达到显著水平(P < 0.05),表明可以进一步进行交配组合的杂种优势和配合力分析。
表 2 各理化因子48 h胁迫下不同凡纳滨对虾群体的耐受成活率Table 2. Effect of physicochemical factors on survival rate of different populations of L.vannamei杂交组合
cross combination耐受成活率/% survival rate 低溶氧lower DO 高氨氮higher ammonia nitrogen UM×UH 84.45±6.19a 64.44±2.94a UM×UM 53.33±3.33de 21.11±1.06de UM×TZ 80.00±7.70ab 55.56±6.37ab UH×UH 66.67±3.47b 54.44±8.29ab UH×TZ 50.00±3.85e 16.67±1.93f UH×UM 55.55±7.78cd 56.67±5.09bc TZ×UM 15.56±2.44f 21.11±5.56ef TZ×UH 65.55±2.22c 62.22±2.82ab TZ×TZ 52.22±4.84e 47.78±7.78cd 均值mean 62.59±4.63 44.44±4.61 注:不同字母表示组间差异达到显著水平(P < 0.05)
Note:Different letters indicate significant difference among treatments at 0.05 level.2.2 各理化因子耐受性的杂种优势
成活率数据经平方根反正弦转换后的杂种优势分析结果见表 3。6个杂交组合耐低溶氧和高氨氮性状的中亲优势和超亲优势均存在较大差异。耐低溶氧性状MP和BP的变化为-52.48%~31.39%和-54.34%~24.66%,其中UM和UH群体的杂交中亲优势均值最高(13.10%);耐高氨氮性状MP和BP的变化为14.09%~42.57%和1.860%~26.18%,UM和UH群体的杂交中亲优势均值最高(36.48%)。
表 3 各理化因子48 h胁迫下各杂交组合耐受成活率的杂种优势Table 3. Survival rate of heterosis of different populations of L.vannamei under stress in 48 h理化因子
physicochemical factor杂交组合cross combination UM×UH UH×UM 均值mean UM×TZ TZ×UM 均值mean UH×TZ TZ×UH 均值mean 中亲优势MP/% 溶氧 31.39 -5.19 13.10 29.73 -52.48 -11.38 2.36 -14.79 -6.22 氨氮 42.57 30.39 36.48 35.60 25.33 30.47 14.09 31.47 22.78 超亲优势BP/% 溶氧 21.99 -11.97 24.66 -54.34 -1.25 -17.79 氨氮 12.29 2.70 10.21 1.86 9.51 26.18 2.3 配合力分析
低溶氧和高氨氮胁迫下成活率性状一般配合力与特殊配合力方差分析见表 4。结果表明低溶氧和高氨氮的一般配合力和特殊配合力方差检验均达到显著(P < 0.05)或极显著水平(P < 0.01),说明这些耐受性性状遗传除受加性遗传效应控制,还受亲本间互作效应的影响。表 5显示了不同群体不同性状的配合力效应值。可以看出,UH群体耐高氨氮和低溶氧性状的一般配合力较高,分别为0.215 7和0.124 2;UM群体和UH群体组合的各抗逆性状的特殊配合力效应值较高,体现了两亲本群体较高的非加性效应。
表 4 各理化因子耐受成活率性状配合力的方差分析Table 4. Variance components of combining ability of survival rate of L.vannamei under stress性状
trait来源
source一般配合力general combining ability 特殊配合力
specific combining ability残差
residual父本♂ 母本♀ 低溶氧lower DO MS 0.344 1 0.079 3 0.240 0 0.023 4 F 14.715 0** 3.390 9 10.262 0** 高氨氮higher ammonia nitrogen MS 0.130 0 0.011 1 0.147 6 0.035 9 F 3.621 7* 0.309 4* 4.110 0* 注:*表示差异显著(P < 0.05);* *表示差异极显著(P < 0.01).
Note:* and * * represent P < 0.05 and P < 0.01,respectively.表 5 凡纳滨对虾群体各耐受性状配合力效应值分析Table 5. Combining ability of survival rate of L.vannamei under stress杂交群体
cross combination理化因子physicochemical factors 低溶氧lower DO 高氨氮higher ammonia nitrogen 一般配合力general combining ability UM父本 0.151 5 -0.042 0 UH父本 0.034 4 0.071 5 TZ父本 -0.185 9 -0.029 5 UM母本 -0.190 5 -0.189 8 UH母本 0.089 9 0.144 2 TZ母本 0.100 7 0.045 6 UM群体 -0.039 0 -0.231 8 UH群体 0.124 2 0.215 7 TZ群体 -0.085 3 0.016 0 母本效应maternal effect UM群体 -0.342 0 -0.147 8 UH群体 0.055 5 0.072 7 TZ群体 0.286 6 0.075 1 特殊配合力specific combining ability UM×UH 0.081 7 0.108 0 UM×TZ -0.060 5 -0.025 0 UH×TZ -0.034 0 0.070 0 3. 讨论
3.1 一般配合力
氨氮是对虾养殖水环境中最主要的污染物质之一,主要以离子氨和非离子氨的形式存在,对对虾的生长、蜕皮、耗氧量、氨氮排泄、渗透压调节及抗病力等多个方面有不利影响[21]。而溶解氧含量也直接影响水生动物的生长和成活。因此,提高凡纳滨对虾对养殖环境的应激适应力是对虾健康种质改良的重要方向。大量研究[22-24]表明,虾苗胁迫耐受性强,在后期养成过程中会具有更高的成活率和显著的生长优势。目前有关对虾耐高氨氮和低溶氧性状选育的研究报道较少,孙苗苗等[23]利用巢式设计构建了13个斑节对虾(Penaeus monodon)家系,评估了稚虾对氨氮的耐受性;何玉英等[24]比较了中国对虾(Fenneropenaeus chinensis)家系幼体的pH和氨氮的耐受性,为中国对虾的抗逆选育提供了基础。该研究中UH群体的耐低溶氧、高氨氮等性状的一般配合力效应值均为正向效应,分别为0.124 2和0.215 7,表明该群体内存在丰富的加性遗传变异。可见凡纳滨对虾不同交配群体对氨氮的耐受性存在较大的遗传变异,这为选育耐高氨氮性状的对虾品系提供了理论依据,也使得从对虾种质改良方面解决对虾养殖环境恶化与追求高产量间的矛盾成为可能。而养殖水质环境与遗传育种工作相结合进行选育的研究工作处于刚起步阶段。该研究以溶氧和氨氮48 h急性胁迫的耐受性成活率为指标,比较分析不同群体之间的体质差别;并将其作为抗逆性状进一步分析杂种优势和配合力效应,比较不同交配组合之间的遗传差别,可为凡纳滨对虾遗传选育工作提供参考。
3.2 特殊配合力和母本效应
凡纳滨对虾高氨氮和低溶氧抗逆性状的表现与其亲本的配合力密切相关,通过亲本配合力分析可以评估各亲本材料作为杂交亲本时的优劣,以及对每个杂交组合的性状好坏作出判断[25]。分析该研究中正反交组合耐高氨氮和低溶氧性状的特殊配合力效应值表明,群体UM和群体UH的杂交后代耐低溶氧、高氨氮等性状的特殊配合力效应值具有较明显的正向优势,杂交后代杂种优势明显,存在较高的抗逆非加性效应,而群体UM或群体UH与群体TZ组合均表现为负的特殊配合力效应值,因此在留种群体中可适当增加UH群体和UM群体数量。此外该研究使用材料为3~4 cm仔虾,有研究指出对虾早期生长性状受母本效应影响显著,黄永春等[26]研究发现,正、反交凡纳滨对虾的抗WSSV遗传上偏向母本,即母体效应占主导地位。该实验中耐高氨氮和低溶氧性状均存在较高的母本效应。因此推断凡纳滨对虾无论是生长性状、抗病性状还是抗逆性状,其早期发育阶段均受到母本效应显著影响,此方面有待深入研究。
3.3 性状相关性
刘海英等[27]研究表明,对虾在氧浓度高的水体中氨氮耐受性强于氧浓度低的水体。对其他水产动物,如大菱鲆[28]、半滑舌鳎(Cynoglossus semilaevis)[29]、黄颡鱼(Pelteobagrus fulvidraco)[30]等的研究也有相似结果。该研究中3个群体凡纳滨对虾双列杂交F1幼虾高氨氮耐受性与低溶氧耐受性间表型相关系数为0.730,相关性检验显著(P < 0.05),而一般配合力相关系数为0.637,相关性检验不显著(P>0.05),其具体原因还有待进一步探讨。但是,该实验发现UM×UH交配组合低溶氧与氨氮耐受性均较好,说明耐低溶氧与耐高氨氮的基因有可能在同一凡纳滨对虾个体上表达。因此,在制定对虾育种方案时,可以采用选择指数法同时考虑高氨氮耐受性和低溶氧耐受性,培育出既耐低溶氧又耐氨氮的凡纳滨对虾品种。NIELSEN等[31]采用选择指数法,提高了鲤(Cyprinus carpio)的生长性状和成活率性状。OFLYNN等[32]利用选择指数法同时提高了大西洋鲑(Salmo salar)幼鲑的成活率和抗BKD成活率。随着研究进一步的深入,笔者将优化育种方案,结合现代育种技术,构建半同胞、全同胞家系,利用最佳线性无偏估计法(BLUP法)估算高氨氮和低溶氧耐受性状遗传参数;确定家系或者个体的选择指数,加快选育速度,培育出具有中国自主知识产权的凡纳滨对虾抗逆新品种。
-
![]()
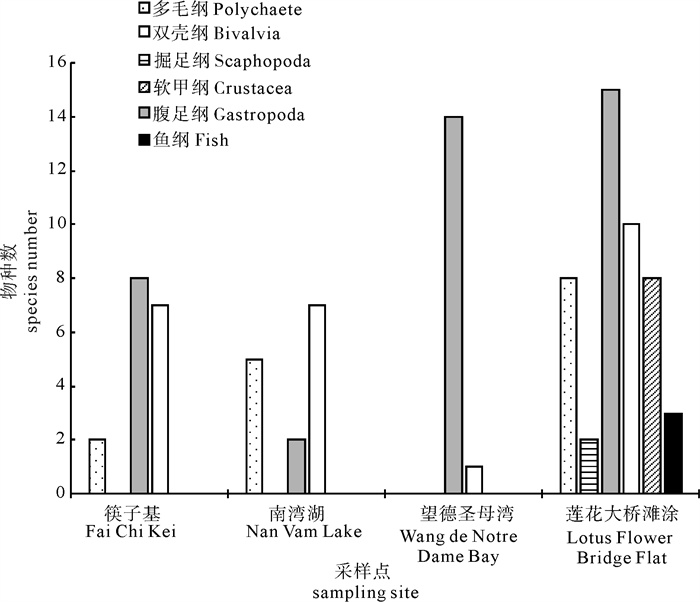
图 1 澳门4个典型湿地的底栖动物群落种类组成
Figure 1. Species composition of macrobenthic communities in each site
![]()
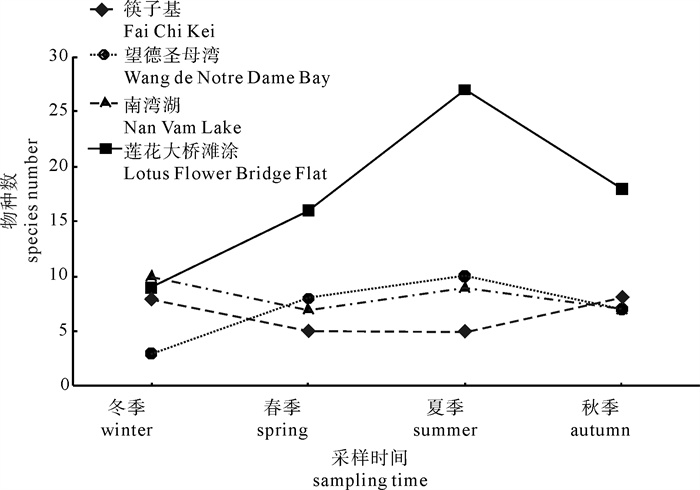
图 2 各采样点底栖动物种类数的季节变化
Figure 2. Seasonal change of species number of macrobenthos at each site
![]()
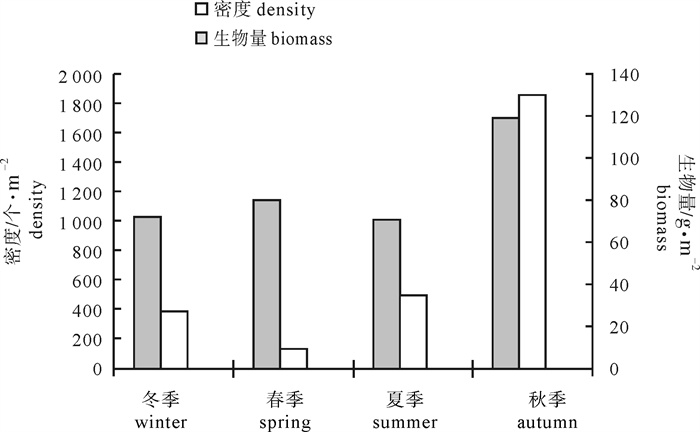
图 3 底栖生物年均栖息密度和生物量变化
Figure 3. Annual change of average density and biomass of macrobenthos
![]()
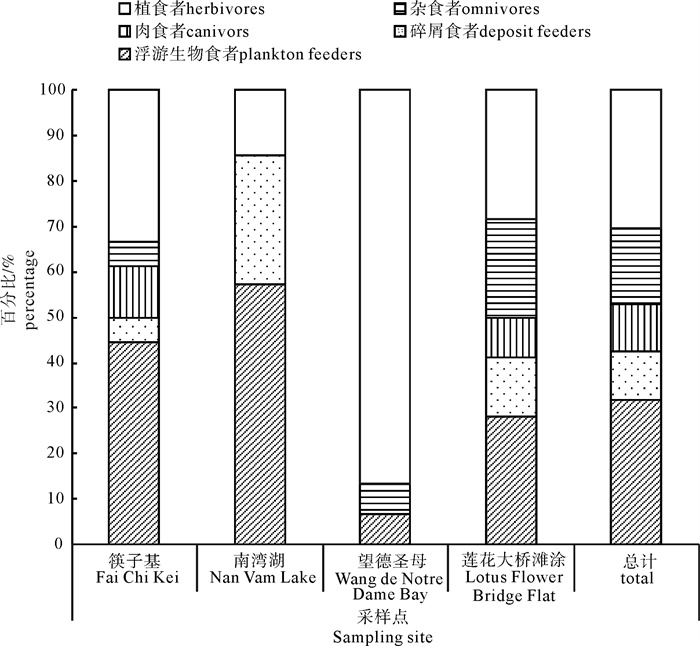
图 4 不同采样点大型底栖动物功能群物种组成百分比
Figure 4. Percentage of species composition in different functional feeding groups at different sampling sites
表 1 各采样点水温、盐度和透明度
Table 1 Water temperature, salinity and transparency at each sampling site
采样点 sampling site 季节 season 透明度/cm transparency 盐度 salinity 温度/℃ temperature 筷子基 Fai Chi Kei 冬季 60 5 24 春季 62 3 22 夏季 100 2 26 秋季 70 5 27 南湾湖 Nan Vam Lake 冬季 80 10 23 春季 75 11 23 夏季 130 5 26 秋季 90 8 27 望德圣母湾 Wang de Notre Dame Bay 冬季 25 0 24 春季 - 0 24 夏季 60 0 25 秋季 - 0 27 莲花大桥滩涂 Lotus Flower Bridge Flat 冬季 25 10 23 春季 15 5 23 夏季 63 4 26 秋季 20 4 27 注:-. 水太浅,不能测量
Note:-. The water was too shallow to determine the transparency. 下载: 导出CSV
下载: 导出CSV
表 2 澳门湿地大型底栖动物名录与分布
Table 2 Composition and distribution of macrobenthos in Macao wetlands
类群 species 采样点 sampling site 生活型 life form 功能群 FFG 筷子基 Fai Chi Kei 南湾湖 Nan Vam Lake 望德圣母湾 Wang de Notre Dame Bay 莲花大桥滩涂 Lotus Flower Bridge Flat 环节动物 Annelida 多毛纲 Polychaete 双腮内卷齿蚕 Aglaophamus dibranchis + + + IN D 羽须鳃沙蚕 Dendronereis pinnaticirris + + IN D 角沙蚕一种 Ceratonereis sp. + + IN D 尖刺樱虫 Potamilla acuminata + + IN Pl 单叶沙蚕一种 Namalycastis sp. + IN D 异足索沙蚕 Lumbriconeris heteropoda + IN D 裸沙蚕一种 Nicon sp. + IN D 全刺沙蚕属一种 Nectoneanthes sp. + + IN C 膜质伪才女虫 Pseudopolydora kempi + IN D 软体动物 Mollusca 掘足纲 Scaphopoda 肋变角贝 Dentalium octangyiatum + IN C 中国沟角贝 Striodentalium chinensis + IN C 腹足纲 Gastropoda 梨形环棱螺 Bellamya purificat + + AD Ph 铜锈环棱螺 B.aeruginosa + AD Ph 方形环棱螺 B.quadrata + AD Ph 尖膀胱螺 Physa acuta + AD Ph 微红萝卜螺 Radix rubiginosa + AD Ph 内切小钝螺 Obtusella cf. intersecta + AD Ph 福寿螺 Pomacea canaliculata + AD Ph 多棱角螺 Angulyagra polyzonata + + AD Ph 瘤拟黑螺 Melanoides tuberculata + + + + AD Ph 斜肋齿蜷 Sermyla riqueti + + + + AD Ph 塔蜷 Thiara scabra + + + AD Ph 斜粒粒蜷 T.granifera + + AD Ph 习见蛙螺 Bursa rana + IN C 中国耳螺 Ellobium chinense + AD Ph 光滑狭口螺 Stenothyra glabar + + + IN O 纵肋织纹螺 Nassarius variciferus + IN C 石蟥 Onchididum verruculatum + AD O 紫游螺 Neritina violacea + AD Ph 短拟沼螺 Assiminea brevicula + IN Ph 台湾山椒螺 A.taiwanensis + AD Ph 圆筒原盒螺 Eocylichna braunsi + IN Pl 扭唇拟捻螺 Acteocina gordonis + IN Pl 河口螺 Iravadia sp. + AD O 中华拟蟹手螺 Cerithidea sinensis + AD O 大脐圆扁螺 Hippentis umbilicalis + AD Ph 尖口圆扁螺 H.cantori + AD Ph 双壳纲 Bivalvia 沼蛤 Limnoperna fortunei + + IN Pl 凸壳肌蛤 Musculus senhousia + IN Pl 凹线仙女蚬 Crernobatissa subsulcata + + IN Pl 河蚬 Corbicula fluminea + + IN Pl 闪蚬 C.nitens + IN Pl 纹斑棱蛤 Trapezium liratum + + IN Pl 幼形亮樱蛤 Nitidotellina juvenilis + IN Pl 黑荞麦蛤 Vignadula atrata + + IN Pl 萨氏仿贻贝 Mytilopsis sallei + + + IN Pl 黑龙江河篮蛤 Potamocorbula amurensis + IN Pl 光滑河篮蛤 P.laevis + IN Pl 四角蛤蜊 Mactra veneriformis + IN Pl 台湾朽叶蛤 Coecella formaosae + IN Pl 波纹巴非蛤 Paphia undulata + IN Pl 菲律宾蛤仔 Ruditapes philippinarum + IN Pl 丽文蛤 Meretrix lusoria + IN Pl 连珠蚶 Mabellarca consociata + IN Pl 双纹须蚶 Barbatia bistrigata + IN Pl 甲壳纲 Crustacea 谭氏泥蟹 Ilyoplax deschampsi + CA Ph 褶痕拟相手蟹 Parasesarma plicatum + CA O 双齿相手蟹 Perisesarma bidens + CA O 无齿相手蟹 Sesarma dehaani + CA O 弧边招潮蟹 Uca arcuata + CA Ph 锯缘青蟹 Scylla serrata + CA C 脊尾白虾 Palaemon carincauda + SW O 日本沼虾 Macrobrachium nipponense + SW O 鱼类 fish 弹涂鱼 Periophthalmus cantonensis + SW O 中华乌塘鳢 Bostrychus sinensis + SW C 条纹三叉 
+ SW O 注:+. 表示在该采样点发现此生物;生活型IN. 底内型;CA. 穴居型;AD. 底上附着型;SW. 游泳底栖型;功能摄食类群;Pl. 浮游生物食者;Ph. 植食者;D. 碎屑食者;C. 肉食者;O. 杂食者
Note:+. species were found at this site. Life form composition:bottom type (IN);cave type (CA);benthic type (AD);swimming benthic type (SW). Functional feeding composition:planktophagous group (Pl);phytophagous group(Ph);detritivorous group(D);camivorous group(C);omnivorous group(O)
下载: 导出CSV
表 3 澳门湿地底栖动物群落不同季节的密度和生物量
Table 3 Seasonal change of density and biomass of macrobenthos communities in different habitats in Macao wetlands
采样季节 season 筷子基 Fai Chi Kei 南湾湖 Nan Vam Lake 望德圣母湾 Wang de Notre Dame Bay 莲花大桥滩涂 Lotus Flower Bridge Flat 密度/个·m-2 density 生物量/g·m-2 biomass 密度/个·m-2 density 生物量/g·m-2 biomass 密度/个·m-2 density 生物量/g·m-2 biomass 密度/个·m-2 density 生物量/g·m-2 biomass 冬季 winter 21.34 1.92 1 152.00 255.52 10.67 23.89 341.33 5.14 春季 spring 0 0 437.33 248.64 37.33 67.52 56.00 4.40 夏季 summer 0 0 1 866.67 248.59 10.67 30.45 106.67 3.12 秋季 autumn 0 0 7 345.67 449.01 32.00 24.48 42.67 2.00 周年平均值 annual mean 5.34 0.48 2 700.42 300.44 22.67 36.59 136.67 3.67
下载: 导出CSV
表 4 各采样点底栖动物优势种及优势度
Table 4 Dominant species and dominance of macrobenthos in each sampling site
采样点 sampling site 优势种及优势度 dominant species and dominance 筷子基 Fai Chi Kei 萨氏仿贻贝M.sallei 0.500;全刺沙蚕属一种Nectoneanthes sp. 0.500 南湾湖 Nan Vam Lake 沼蛤L.fortunei 0.786;凸壳肌蛤M.senhousia 0.083;纹斑棱蛤T.liratum 0.062;萨氏仿贻贝M.sallei 0.041 望德圣母湾 Wang de Notre Dame Bay 多棱角螺A.polyzonata 0.588;梨形环棱螺B.purificat 0.353 莲花大桥滩涂 Lotus Flower Bridge Flat 羽须鳃沙蚕 D.pinnaticirris 0.688;谭氏泥蟹I.deschampsi 0.098;角沙蚕Ceratonereis sp. 0.059
下载: 导出CSV
表 5 各个采样点底栖动物群落生物多样性指数
Table 5 Biodiversity indices of macrobenthos in each sampling site
采样点 sampling site 季节 season 丰富度指数 D 多样性指数 H′ 均匀度指数 J′ 南湾湖 Nan Vam Lake 冬季 1.116 0.848 1.650 春季 1.542 0.921 1.277 夏季 1.443 1.000 0.693 秋季 0.553 0.356 0.573 平均值 1.163 0.781 1.048 望德圣母湾 Wang de Notre Dame Bay 冬季 0.910 0.918 0.637 春季 0.908 0.727 1.171 夏季 1.024 0.309 0.602 秋季 0.976 0.859 0.480 平均值 0.955 0.703 0.722 莲花大桥滩涂 Lotus Flower Bridge Flat 冬季 0.558 0.650 0.451 春季 1.642 0.838 1.502 夏季 1.669 0.969 1.735 秋季 0.962 0.887 0.974 平均值 1.208 0.836 1.166
下载: 导出CSV
表 6 不同采样点大型底栖动物生活型及生活类群组成特征
Table 6 Life form composition of macrobenthos in different sampling sites
生活型及比值 life forms & ratio 筷子基 Fai Chi Kei 南湾湖 Nan Vam Lake 望德圣母湾 Wang de Notre Dame Bay 莲花大桥滩涂 Lotus Flower Bridge Flat 总数 total 底上附着型 benthic type (AD) 6 2 13 9 20 底内型 bottom type (IN) 11 12 2 26 35 穴居型 cave type (CA) 0 0 0 6 6 游泳底栖型 swimming benthic type (SW) 0 0 0 5 5 面上群 group of surface (GS) 6 2 13 15 25 面下群 group of surface below (GSB) 11 12 2 31 41 面上群/面下群 GS/GSB 0.55 0.17 6.50 0.48 0.61
下载: 导出CSV
-
[1] 胡知渊, 鲍毅新, 程宏毅, 等. 中国自然湿地底栖动物生态学研究进展[J]. 生态学杂志, 2009, 28(5): 959-968. [2] 梁华. 澳门路凼填海区湿地生物群落结构的动态变化及物种多样性研究[D]. 广州: 暨南大学, 2007. https://kns.cnki.net/kcms2/article/abstract?v=BkYFcxcfasA44XZxE433c79zmlCX0vJWL8UU63IH3DbeSPFKpB6mNkpPKi79xEgP70KoFQGhLlVF7Tz5IwDLTEW5V0r7MNXItbIVYh0j5F-ElIu9o3zfHHziEH5WcwcD3f69HE6fu3QEQdiDiVPp7_xolhtWY2CwA_5kE82Hr9eJ_FgGaWFhuqEAx0Xb7STr&uniplatform=NZKPT&language=CHS [3] 何伟添. 澳门海岸湿地生态系统的特征及变化趋势研究[D]. 广州: 暨南大学, 2008. https://kns.cnki.net/kcms2/article/abstract?v=BkYFcxcfasAbGO62Sym65pQSfJ-jAz8PToUsavong18HN4siU8gSbFmcEfn9egJqDBL-Ke28baohn6JomRixXgK1yjWMiuh_JOGaD7XCBmGGg51W6iIiRr-kLngkBGd94oImext6coygtNx7YsMDy7_MNBMWICxwRVHsmGXrN8wu6yZ8BUAbr09tgYtf5BYb&uniplatform=NZKPT&language=CHS [4] ZHANG L, HONG P I. A new species of Fissidens with remarkable rhizoidal tubers and gemmae from Macao, China[J]. J Bryol, 2011, 33(1): 50-53. doi: 10.1179/1743282010Y.0000000010
[5] 陈金秋. 珠江河口不同类型湿地底栖多毛类形态学及群落生态研究[D]. 广州: 中山大学, 2007. [6] 吴宝玲. 中国近海沙蚕科研究[M]. 北京: 海洋出版社, 1981: 1-228. https://xueshu.baidu.com/usercenter/paper/show?paperid=078d8d42c5e78aa9a4bb3ecca064ed22&site=xueshu_se&hitarticle=1 [7] 齐忠彦. 中国经济软体动物[M]. 北京: 中国农业出版社, 1998: 1-325. https://xueshu.baidu.com/usercenter/paper/show?paperid=83bba212e4c723f036562361bd10d5e4&site=xueshu_se&hitarticle=1 [8] 徐凤山, 张素萍. 中国海产双壳类图志[M]. 北京: 科学出版社, 2008: 1-336. https://kns.cnki.net/kcms2/article/abstract?v=BkYFcxcfasCT08PS3E93Vuu_ZRxGwcZyvfgmbOfPUfOryGevyRdsz4lWAZuy6_ictNS3sJc9lzs4mSFuI58UdtxoXJK4zRRj26GoOWkyPg-xZ4ERcxpBAWuQFzlyXGQ7owci9sA7zyCaX3muOeqcE0pbs8NUnTuyHIMxPreAbj25qV8dmDSxGZoAU1d0OsJYE9oviCpXhiY=&uniplatform=NZKPT&language=CHS [9] 杨文, 蔡英亚, 邝雪梅. 中国南海经济贝类原色图谱[M]. 北京: 中国农业出版社, 2012: 1-271. https://kns.cnki.net/kcms2/article/abstract?v=BkYFcxcfasCT08PS3E93Vuu_ZRxGwcZyvfgmbOfPUfOryGevyRdsz4lWAZuy6_ictNS3sJc9lzs4mSFuI58UdtxoXJK4zRRj26GoOWkyPg-xZ4ERcxpBAWuQFzlyXGQ7owci9sA7zyCaX3muOeqcE0pbs8NUnTuyHIMxPreAbj25qV8dmDSxGZoAU1d0OsJYE9oviCpXhiY=&uniplatform=NZKPT&language=CHS [10] 何斌源, 赖廷和. 广西北部湾红树林湿地海洋动物图谱[M]. 北京: 科学出版社, 2013: 1-236. https://xueshu.baidu.com/usercenter/paper/show?paperid=25da657171c64dc744bc4eb49508c79b&site=xueshu_se&hitarticle=1 [11] 黄宗国, 林茂. 中国海洋生物图集第四册[M]. 北京: 海洋出版社, 2012: 1-348. https://xueshu.baidu.com/usercenter/paper/show?paperid=1d599adedbbc68cbfc46fb30c5a60e9f&site=xueshu_se&hitarticle=1 [12] THORP J H, ROGERS D C. Field guide to freshwater invertebrates of North America[M]. New York: Academic Press, 2010: 65-95. https://xueshu.baidu.com/usercenter/paper/show?paperid=1b5da39cdfc785c34024f3de9a48c767&site=xueshu_se
[13] 尤仲杰, 陶磊, 焦海峰, 等. 象山港大型底栖动物功能群研究[J]. 海洋与湖沼, 2011, 42(3): 431-435. doi: 10.11693/hyhz201103015015 [14] 鲍毅新, 胡知渊, 李欢欢, 等. 灵昆东滩围垦区内外大型底栖动物季节变化和功能群的比较[J]. 动物学报, 2008, 54(3): 416-427. [15] 王卉, 钟山, 方展强. 珠海鹤洲水道沿岸红树林湿地大型底栖动物群落特征[J]. 生态学报, 2013, 33(21): 6913-6924. doi: 10.5846/stxb201207181017 [16] 杨泽华, 童春富, 陆健健. 长江口九段沙潮间带底栖动物功能群[J]. 动物学研究, 2003, 24(5): 355-361. doi: 10.3321/j.issn:0254-5853.2003.05.006 [17] 范航清, 何斌源, 韦受庆. 海岸红树林地沙丘移动对林内大型底栖动物的影响[J]. 生态学报, 2000, 22(6): 811-819. doi: 10.3321/j.issn:1000-0933.2000.05.002 [18] 黄洪辉, 林燕棠, 李纯厚, 等. 珠江口底栖动物生态学研究[J]. 生态学报, 2002, 22(4): 603-607. doi: 10.3321/j.issn:1000-0933.2002.04.024 [19] 彭松耀, 赖子尼, 蒋万祥, 等. 珠江口大型底栖动物的群落结构及影响因子研究[J]. 水生生物学报, 2010, 34(6): 1179-1189. doi: 10.3321/j.issn:1000-0933.2002.04.024 [20] 张敬怀. 珠江口及邻近海域大型底栖动物多样性随盐度、水深的变化趋势[J]. 生物多样性, 2014, 22(3): 302-310. doi: 10.3724/SP.J.1003.2014.13141 [21] 袁兴中. 河口潮滩湿地底栖动物群落的生态学研究[D]. 上海: 华东师范大学, 2001. https://xueshu.baidu.com/usercenter/paper/show?paperid=2b66b260105f6379fc7a35106d5e4113 [22] 安传光, 赵云龙, 林凌, 等. 长江口九段沙潮间带大型底栖动物季节分布特征的初步研究[J]. 水产学报, 2007(增刊): 52-58. https://kns.cnki.net/kcms2/article/abstract?v=BkYFcxcfasApUZRcMHOODuO1rURdWjsBAGEV8oIHxGqCyzZ_RjmTz_1jdrKDeyE06O8t7LPa7m99Tl5as1ruDVEpi2X9T8ahTzwKUTXIklZI40dVdwGheG2Wpf0bUhr52yk0MpauStAb9Sf5X2FWFzvzO02UKQX4S5ZI_CHrVRf7hiRlTSUduIb9_E5tsb3B&uniplatform=NZKPT&language=CHS [23] 黄睿婧, 蔡立哲, 叶洁琼, 等. 广州南沙十四涌潮间带三种生境的大型底栖动物群落比较[J]. 生态学杂志, 2010, 29(6): 1187-1192. https://www.cje.net.cn/CN/abstract/abstract16717.shtml [24] THOMAS J D. Mutualistic interactions in freshwater modula systems with molluscan components[J]. Adv Ecol Res, 1990, 20: 125-178. http://www.sciencedirect.com/science/article/pii/S0065250408600549
[25] JONES J I, SAYER C D. Does the fish-invertebrate-periphyton cascade precipitate plant loss in shallow lakes?[J]. Ecology, 2003, 84(8): 2155-2167. doi: 10.1890/02-0422
[26] 杜飞雁, 王雪辉, 李纯厚, 等. 大亚湾大型底栖动物物种多样性现状[J]. 南方水产, 2008, 4(6): 34-41. [27] 王宝强, 薛俊增, 庄骅, 等. 洋山港潮间带大型底栖动物群落结构及多样性[J]. 生态学报, 2011, 31(20): 5865-5874. [28] 袁兴中, 陆健健. 长江口潮滩湿地大型底栖动物群落的生态学特征[J]. 长江流域资源与环境, 2002, 11(5): 414-420. doi: 10.3969/j.issn.1004-8227.2002.05.005 [29] 蔡立哲, 林鹏, 佘书生. 深圳河口泥滩多毛类动物的生态研究[J]. 海洋环境科学, 1998, 17(1): 42-48. [30] 蔡立哲, 厉红梅, 林鹏, 等. 深圳河口潮间带泥滩多毛类的数量变化及环境影响[J]. 厦门大学学报: 自然科学版, 2001, 40(3): 741-750. doi: 10.3321/j.issn:0438-0479.2001.03.016 [31] 黄建荣, 刘启智, 赵一臣, 等. 珠海淇澳岛红树林蟹类区系[J]. 生态环境学报, 2011, 20(4): 730-736. doi: 10.3969/j.issn.1674-5906.2011.04.026 [32] 周晓, 葛振鸣, 施文彧, 等. 长江口新生湿地大型底栖动物群落时空变化格局[J]. 生态学杂志, 2007, 26(3): 372-377. doi: 10.3321/j.issn:1000-4890.2007.03.015 [33] 安传光, 赵云龙, 林凌, 等. 崇明岛潮间带夏季大型底栖动物多样性[J]. 生态学报, 2008, 28(2): 577-586. doi: 10.3321/j.issn:1000-0933.2008.02.01649|2005||| [34] 蔡永久, 姜加虎, 张路, 等. 长江中下游湖泊大型底栖动物群落结构及多样性[J]. 湖泊科学, 2010, 18(1): 50-59. doi: 10.18307/2010.0602 [35] 刘健康. 东湖生态学研究[M]. 北京: 科学出版社, 1990: 147-148. http://ir.ihb.ac.cn/handle/342005/11046 [36] 任淑智. 北京地区河流中大型底栖无脊椎动物与水质关系的研究[J]. 环境科学学报, 1991, 11(1): 31-46. -
期刊类型引用(8)
1. 李凤玉,解绶启,贠彪,钱雪桥. 饲料蛋白质水平对斑节对虾(Penaeus monodon)生长性能、消化酶活性和血清生化指标的影响. 饲料工业. 2023(08): 76-82 .  百度学术
百度学术
2. 李钊丞,项盛羽,沈梦庭,王秀秀,张日新,曹正良. 南美白对虾快速游动发声特征及其信息利用研究. 水生生物学报. 2023(06): 997-1006 . 百度学术
3. 王学习,陈友,王德娟,郑运宗,陈政榜,王余鑫,朱文博,邵建春. 鸡肝粉部分替代鱼粉对凡纳滨对虾生长性能、饲料利用、消化、抗氧化和免疫能力的影响. 饲料研究. 2023(20): 57-62 . 百度学术
4. 王建,曾本和,徐兆利,张忭忭,刘海平,王万良,王金林,周建设,黄莉萍. 饲料蛋白水平对拉萨裸裂尻幼鱼肠道和肝脏的消化酶活性及组织结构的影响. 南方水产科学. 2019(06): 112-119 . 本站查看
5. 路晶晶,郭冉,夏辉,解伟,王美雪,杨品贤,李娜. 家禽副产物酶解肽对凡纳滨对虾生长性能、消化指标和非特异性免疫指标的影响. 动物营养学报. 2018(02): 797-806 . 百度学术
6. 苟妮娜,王开锋. 两种盐度下饲料蛋白水平对凡纳滨对虾消化酶活力的比较研究. 基因组学与应用生物学. 2017(08): 3229-3236 . 百度学术
7. 斯烈钢,邹李昶,申屠基康,朱卫东. 饲料添加不同脂肪及蛋白质水平对日本沼虾(Macrobrachium nipponensis)生长性能、体成分及消化酶活力的影响. 海洋与湖沼. 2014(02): 400-408 . 百度学术
8. 周凡,王亚琴,林玲,马文君,丁雪燕,何丰,薛辉利. 饲料蛋白水平对中华鳖稚鳖生长和消化酶活性的影响. 浙江农业学报. 2014(06): 1442-1447 . 百度学术
其他类型引用(7)
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 15
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
