
Comparison of seven metallic elements in prismatic layer and nacreous layer of shells from four shell color strains in pearl oyster (Pinctada fucata)
-
摘要:
利用电感耦合等离子体发射光谱法(ICP-AES)对合浦珠母贝(Pinctada fucata)黑、金、红、白F5代选育群体贝壳棱柱层和珍珠质层的7种金属元素质量分数进行比较分析。结果显示, 合浦珠母贝贝壳棱柱层中铁(Fe)、钾(K)、镁(Mg)和锰(Mn)的质量分数明显高于珍珠质层, 而钠(Na)、Sr(锶)和锌(Zn)的质量分数则低于珍珠质层。4种壳色棱柱层和珍珠质层中的金属元素质量分数均存在差异: Mg、Sr、K、Zn在黑壳合浦珠母贝棱柱层中质量分数最低, Mn、Fe在金壳棱柱层中最低, 而Na却在红壳棱柱层中最低; 单因素方差分析显示, Fe、Mg、Zn可能分别会影响合浦珠母贝红壳、金壳、白壳的形成。白壳贝珍珠质层的6种金属元素(Mg除外)质量分数都最少, 更有可能培育出优质珍珠。
Abstract:We analyzed the contents of seven metallic elements in prismatic layer and nacreous layer of shells from four shell color strains [(B), gold (G), red (R) and white (W)] in pearl oyster (Pinctada fucata) by ICP-AES. The Fe, K, Mg and Mn contents in prismatic layer were obviously higher than those in nacreous layer. The elements of Mg, Sr, K and Zn in prismatic layer of black shell were the lowest. Similarly, the Mn and Fe contents in prismatic layer of gold shell were lower than those in the other color shells, but Na had minimum content in red shell. Fe, Mg and Zn might have influence on the formation of red, gold and white shells. Besides, the nacreous layer of white shell had the lowest level in the content of analytical elements except Mg, which indicates that the white shell in pearl oysters is more likely to produce pearls of better quality.
-
Keywords:
- Pinctada fucata /
- shell color /
- metallic elements /
- prismatic layer /
- nacreous layer
-
合浦珠母贝(Pinctada fucata)是暖水性贝类,是中国培育海水珍珠最主要的贝种,所培养珍珠占海水珍珠总产量90%以上。然而,目前合浦珠母贝养殖业出现育珠质量日益下降的问题,亟需通过有效的手段提高珍珠质量。海水珍珠是通过人工向育珠母贝进行细胞小片移植,刺激育珠母贝的外套膜形成珍珠囊分泌珍珠质沉积在人工插入的核上而成[1],因而人工培育海水珍珠的质量同时受到小片贝和育珠贝的影响[2-6]。目前的研究认为不同壳色育珠贝的育珠性能指标(留核率、珠层厚度和育珠绩效值)存在显著差异[3];珍珠的大小、珠层厚度均与育珠贝的壳长、壳高、壳质量、体质量等可量性状间存在极显著的相关性[4];珍珠颜色和光泽主要受小片贝壳色的影响,而可能与育珠贝无关[2, 5-6]。因此,要从根本上提高海水珍珠质量,优质小片贝和育珠贝的选育是关键。
KY等[7-8]已证明可通过对黑蝶贝(P.margari-tifera)小片贝的选育来改善其所产珍珠的质量(光泽、颜色和形状)。ACOSTA-SALMÓN等[9]研究认为提供细胞小片贝的贝壳珍珠层颜色决定珍珠的颜色。符韶等[3]的研究发现,黑、金、红、白4个壳色合浦珠母贝作为插核贝育珠的留核率、珠层厚度和育珠绩效值均存在显著性差异(P<0.05),其中黑壳色具有最高的留核率、珠层厚度和育珠绩效值。因此,开展合浦珠母贝壳色选育研究具有重要的现实意义。笔者以合浦珠母贝的黑、金、红、白壳色作为遗传标记,并通过定向选育使壳色不断纯化,建立了合浦珠母贝的4个壳色选育群体。
合浦珠母贝贝壳棱柱层的颜色最常见的有黑色、金色、红色和白色4种。壳色在过去仅作为一种分泌产物被忽视,事实上珍珠贝的壳色不仅与其生长、成活等性状有关[9-10],还与优质珠的培育及产量息息相关[3, 9-11]。因此,贝类壳色致色因素的研究引起了国内外学者的广泛关注。近年来,对贝壳壳色致色机理的研究认为,金属元素的存在和含量与贝壳、珍珠的颜色有一定的关系。金属卟啉是自然界中最常见的一种色素,血红素、叶绿素均是卟啉与不同金属离子的配合物,且卟啉类是存在于双壳类贝壳中重要的一类色素[12-14]。此外,张蕴韬[14]认为海水珍珠中的金属离子一部分以离子形式储存于文石(珍珠质层)中,一部分与有机色素中卟啉类结合形成金属卟啉,因其结合的金属种类与含量各有不同而形成不同颜色。然而目前对金属元素的研究仍多集中于其与珍珠呈色的关系上[14-15],而关于金属元素种类和含量变化对贝壳棱柱层和珍珠质层颜色形成的影响则鲜见报道。
金属元素的研究方法有很多种,目前常使用方法有原子吸收光谱(AAS)、电感耦合等离子体质谱(ICP-MS)、电感耦合等离子体原子发射光谱(ICP-AES)等技术。ICP-AES法适用范围广,不仅灵敏度高,精密度好(0.15%~2.00% RSD),而且可以同时分析多种元素,因此在环境、生命科学相关样品的元素分析中的应用逐渐增多[16-18]。此研究用ICP-AES法测定了黑、金、红、白4种壳色合浦珠母贝贝壳棱柱层和珍珠质层中7种重要的微量金属元素质量分数,以比较分析珍珠质层和棱柱层金属元素质量分数差异对其形成的影响并了解合浦珠母贝4种壳色选育群体贝壳棱柱层中金属元素的质量分数,以期为合浦珠母贝壳色形成与金属元素之间的关系提供有价值的试验数据。此外,贝壳珍珠质层及珍珠都是由贝类外套膜组织通过生物矿化形成,二者的组成成分基本类似,均由95%以上的碳酸钙(CaCO3)和低于5%的有机质组成,育珠贝贝壳珍珠质层的金属元素质量分数间接反映出其所产珍珠的珍珠质中金属元素的质量分数。杨磊等[16]研究认为镁(Mg)、铁(Fe)、锌(Zn)和锰(Mn)的含量过高会影响海水珍珠的质量,因此对贝壳珍珠质层进行金属元素质量分数的比较分析,还有助于合浦珠母贝壳色选育过程中优质育珠贝的培育。
1. 材料与方法
1.1 试验样品
试验所用样品是笔者课题组选育出来的合浦珠母贝F5代黑色(B)、金色(G)、红色(R)和白色(W)4个群体的1龄贝。试验用贝为同批次、培育环境一致的选育群体。每个合浦珠母贝壳色选育群体中随机采样,把贝体表面附着物及淤泥洗刷干净后,4 ℃避光保存备用。
1.2 试剂及仪器
试剂有硝酸(优级纯,广州化学试剂厂出品),超纯水。
仪器有微波消解仪(Mars 5,美国CEM公司出品),电感耦合等离子体发射光谱仪(ICP-AES,美国ThermoFisher公司出品)。
1.3 样品的消解和测定
将样品用5%氢氧化钠(NaOH)溶液浸泡15 min除去贝壳表面的有机质及杂质,再用蒸馏水和超纯水分别冲洗2~3次后,置于60 ℃真空干燥24 h,后用机械打磨的方法分开贝壳的棱柱层和珍珠质层,置于玛瑙研钵中研磨,过200目筛。棱柱层和珍珠质层粉末置于105 ℃干燥3 h备用。
准确称取已干燥好的样品约0.1 g(精确至0.000 1 g),放进聚四氟乙烯消化管底部,加入8~10 mL的硝酸消解液,盖上内盖,装好外套,放入微波消解系统的转盘上,选择适当的工作条件进行消解。消解结束后,待冷却后取出消解管,慢慢拧开外套,打开内盖,在通风橱放气约15 min。再置于电热板上150 ℃加热赶酸,至消解管中仅剩约1 mL无色透明的液体时,转移至50 mL容量瓶中定容并摇匀后过滤,滤液送至中山大学测试中心检测Fe、K(钾)、Mg、Mn、Na(钠)、Sr(锶)和Zn 7种金属元素的质量分数。每个壳色群体取6个贝做平行试验,且每个元素测3次取平均值。
2. 结果与讨论
2.1 棱柱层与珍珠质层金属元素质量分数的差异
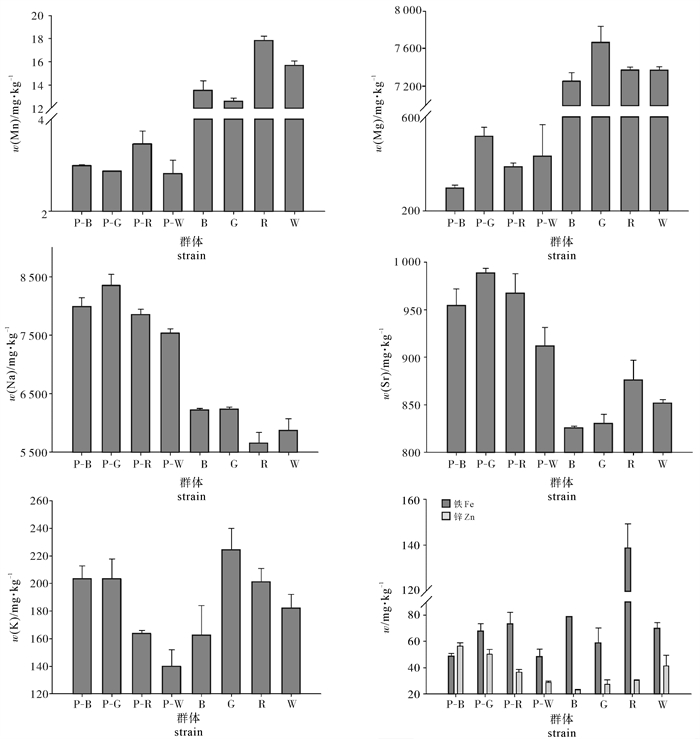
合浦珠母贝贝壳棱柱层和珍珠质层金属元素质量分数分析的结果显示,Na、Mg、Sr的质量分数高,分别大于5 600 mg · kg-1、290 mg · kg-1和820 mg · kg-1(表 1、表 2和图 1)。木士春等[19]对海水珍珠微量元素的质量分数分析认为,海水养殖珍珠多生长于浅海或海湾,生长水域通常具有弱还原性,容易富集Na、Mg、Sr等微量元素。此研究中合浦珠母贝的养殖是在浅海区域进行的,较容易富集Na、Mg、Sr这3种微量元素而导致其在贝壳中的质量分数比其他元素高。在所分析的这7种金属元素中,Fe、K、Mg和Mn的质量分数在棱柱层中明显高于珍珠质层,其中Mn和Mg在合浦珠母贝贝壳棱柱层和珍珠质层中的质量分数在数量级上就已出现差异,在棱柱层中的质量分数与珍珠质层中的质量分数相差10~40倍(图 1)。然而,棱柱层Na、Sr和Zn的质量分数明显低于珍珠质层,其中棱柱层中的质量分数分别为(5 997.15±108.00)mg · kg-1、(845.96±9.05)mg · kg-1和(30.50±3.12)mg · kg-1,珍珠质层的分别为(7 935.67±121.67)mg · kg-1、(955.71±15.50)mg · kg-1和(43.07±2.22)mg · kg-1(表 1和表 2)。综上所述,贝壳棱柱层和珍珠质层间对金属元素的沉积能力存在差异,且两者间的无机物质量分数不同。
表 1 4种壳色合浦珠母贝贝壳棱柱层金属元素质量分数Table 1. Metallic element contents in prismatic layer of shells from four shell color strains of P.fucatamg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) 黑壳(B) 78.32±0.54a 162.57±21.47a 7 259.69±87.27a 13.45±0.81a 6 226.10±20.93a 825.75±1.86a 23.07±0.47a 金壳(G) 58.72±11.51a 224.38±15.44b 7 674.63±165.29b 12.57±0.28a 6 233.30±35.18a 830.27±9.66a 27.33±3.28ab 红壳(R) 138.66±10.56b 200.92±9.97b 7 378.63±31.03a 17.85±0.36b 5 657.00±179.97b 876.11±20.84b 30.31±0.47bc 白壳(W) 69.99±4.31a 182.07±10.02c 7 374.81±36.14a 15.67±0.40c 5 872.20±195.92b 851.69±3.82c 41.28±8.27d 平均值average 86.42±6.73 192.49±14.23 7 421.94±79.93 14.91±0.46 5 997.15±108.00 845.96±9.05 30.50±3.12 注:同列不同小写字母表示差异显著( P<0.05)
Note:Values with different letters in the same column have significant difference (P<0.05).表 2 4种壳色合浦珠母贝贝壳珍珠质层金属元素质量分数Table 2. Metallic element contents in nacreous layer of shells from four shell color strains of P.fucatamg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) P-B 48.56±2.36a 203.36±9.28a 297.33±12.22a 2.99±0.015a 7 992.04±150.69a 954.75±17.16a 56.44±2.62a P-G 67.77±5.68b 203.16±14.69a 516.95±38.18b 2.88±0.002b 8 354.21±181.37a 988.45±5.01b 50.22±3.65a P-R 73.41±8.80c 163.62±2.44b 387.18±16.01c 3.47±0.268c 7 853.86±88.11b 967.71±20.24ab 36.73±1.73b P-W 48.26±5.92a 139.99±11.94c 432.85±133.36abc 2.81±0.295ab 7 542.58±66.52b 911.93±19.58c 28.90±0.87c 平均值average 59.50±5.69 177.53±9.59 408.58±49.94 3.04±0.145 7 935.67±121.67 955.71±15.50 43.07±2.22 注:P-B、P-G、P-R和P-W分别代表合浦珠母贝黑壳、金壳、红壳和白壳珍珠质层。同列不同小写字母表示差异显著(P<0.05)
Note:P-B,P-G,P-R and P-W represent nacreous layers of black,gold,red,white shells in pearl oyster,respectively. Values with different letters in the same column have significant difference (P<0.05).![]() 图 1 4个壳色合浦珠母贝珍珠质层和棱柱层的金属元素质量分数分布Figure 1. Distribution of each metallic element in prismatic layer and nacreous layer of shells from four shell color strains of P.fucata
图 1 4个壳色合浦珠母贝珍珠质层和棱柱层的金属元素质量分数分布Figure 1. Distribution of each metallic element in prismatic layer and nacreous layer of shells from four shell color strains of P.fucata此试验所分析的7种金属元素是生物体进行正常生理活动不可缺少的微量元素。在贝壳形成过程中,金属元素可以通过与蛋白质(酶)结合而影响棱柱层和珍珠质层的生物矿化过程,而且CaCO3结晶形成的方式(珍珠层或棱柱层)受到不同金属元素影响[20-22]。已有研究发现,贝壳中的金属离子Mg能与金属酶ALP结合而影响珍珠质层形成的速度[20];Mg、Fe和Mn等金属离子含量较高,可能导致基质蛋白类有机质发生变性,影响CaCO3的生长结晶[16]。此外,Mg、Fe和Mn等金属离子含量较高时,可以进入方解石(棱柱层)的晶格取代钙离子的位置影响方解石的相变,却无法进入文石(珍珠质层)的晶格,阻碍文石CaCO3晶体的形成[22-23]。棱柱层中Mg、Fe和Mn的质量分数分别为(7 421.94±79.93)mg ·kg-1、(86.42±6.73)mg · kg-1和(14.91±0.46)mg · kg-1,明显高于珍珠质层(表 1和表 2),棱柱层的基质蛋白发生变性的可能比珍珠质层大,且其可影响棱柱层方解石的相变却无法影响珍珠质层的文石相变。因此,合浦珠母贝棱柱层和珍珠质层的形成也受到Mg、Fe和Mn 3种元素质量分数的影响。王慧等[24]也已证明,造成贝壳珍珠质层和棱柱层结晶差异的原因可能是金属元素含量的差异,而非金属元素的种类。此研究中贝壳棱柱层和珍珠质层7种金属元素质量分数的差异显著(图 1),因此推测金属元素相对质量分数的差异可能影响贝壳的形成,这也许是棱柱层和珍珠质层产生差异的原因之一。
2.2 4个壳色选育群体贝壳棱柱层金属元素的质量分数差异
贝壳或者珍珠中金属元素的质量分数与珍珠贝的种类、养殖环境及其分布的地理位置有密切关系[25-26]。此研究所用的样品是课题组培育的同批次、养殖环境一致的合浦珠母贝黑、金、红、白4种选育群体,试验所用选育群体间的差异仅是壳色差异,因此选育群体间金属元素质量分数差异可能与合浦珠母贝的壳色(棱柱层颜色)差异存在相关性。比较4种壳色合浦珠母贝贝壳棱柱层的各金属元素质量分数差异显示(图 1),w(Mn)为R(红壳)>W(白壳)>B(黑壳)>G(金壳);w(Mg)为G>R>W>B;w(Na)为G>B>W>R;w(Sr)为R>W>G>B;w(K)为G>R>W>B;Fe为R>B>W>G;w(Zn)为W>R>G>B。综上所述,4种壳色的合浦珠母贝贝壳棱柱层中金属元素质量分数存在差异,金属元素质量分数与合浦珠母贝壳色的形成存在关联;且金属元素对壳色的影响并非是单一的金属元素造成,而是多种金属元素共同作用的结果,与逯云召[27]对黑、黄、红、白4种壳色合浦珠母贝贝壳中金属元素分析的结果类似。
目前对贝壳中金属元素的研究认为,金属元素一部分以离子形式存在于方解石(棱柱层)中影响棱柱层的形成;一部分金属元素可与棱柱层的有机色素中的卟啉结合形成金属卟啉,卟啉结合的金属元素不同表现出来不同的颜色,且随着各种金属元素的质量分数不同,金属卟啉显示出来的颜色色调也不同[14]。由此可知,金属元素的质量分数差异可能反映出某种金属卟啉的质量分数差异。表 1的单因素方差分析显示,红壳贝中Fe的质量分数最高,且与其余3种壳色贝之间存在显著差异,因而Fe可能与合浦珠母贝红壳的形成有关;Mg在金壳贝中的质量分数最高并与其余壳色贝之间存在显著差异,可能与金壳的形成存在关联;Zn在白壳合浦珠母贝中的质量分数最高,且与其余壳色贝间存在显著差异,表明Zn可能与白壳的形成有关。同种金属元素在4种壳色合浦珠母贝棱柱层中的质量分数不同且在不同壳色合浦珠母贝之间存在显著差异(表 1和图 1),导致贝壳棱柱层中的金属卟啉显示出同种颜色的不同色调。然而,多种金属元素在合浦珠母贝的同一个体贝壳棱柱层中共同存在,表明棱柱层中多种金属卟啉共同作用可能是合浦珠母贝个体间产生颜色差异的因素之一。此外,在同一养殖水域中,不同壳色合浦珠母贝棱柱层中金属元素质量分数的差异,表明不同壳色选育群体贝壳对金属元素的沉积能力也不同。此研究第一次探讨了合浦珠母贝贝壳棱柱层金属元素质量分数对壳色形成的影响。
2.3 4个壳色选育群体贝壳珍珠质层金属元素的质量分数差异
杨磊等[16]对优质珍珠(即珍珠圆润、皎洁、色泽鲜艳、光彩耀目,珠心不含有泥沙、污物等杂质)和劣质珍珠(即乌珠—珠面光滑,但珠心含有泥沙、污物,从外观上看珠质发黑,色深而杂)的研究发现,劣质珍珠的珍珠质层中w(Mg)、w(Fe)、w(Zn)和w(Mn)分别为(645±7.43)mg · kg-1、(67±1.50)mg · kg-1、(460±39.69)mg · kg-1和(18.5±1.50)mg · kg-1,优质珍珠分别为(282±51.83)mg ·kg-1、(34±4.19)mg · kg-1、(74±10.35)mg · kg-1和(6.9±1.10)mg · kg-1。此研究结果显示,w(Mg)为G>W>R>B;w(Fe)为R>G>B>W;w(Zn)为B>G>R>W;w(Mn)为R>B>G>W(图 1)。合浦珠母贝白壳珍珠质层中w(Fe)[(48.26±5.92)mg ·kg-1]、w(Zn)[(28.90±0.87)mg · kg-1]和w(Mn)[(2.81±0.295)mg · kg-1]均最低,w(Mg)[(432.85±133.36)mg · kg-1]也并未达到劣质珍珠的质量分数指标(表 2)。育珠贝的贝壳珍珠质层中金属元素质量分数差异可间接反映出其所育珍珠的珍珠质层金属元素质量分数差异,因而较之其余壳色合浦珠母贝,白壳的合浦珠母贝更有可能培育出优质珍珠。
在海水珍珠的人工育珠过程中,珍珠贝的壳色和贝壳珍珠质层的颜色均对海水珍珠的颜色存在影响。MCGINTY等[28]研究发现,当将白蝶贝(P.maxima)作细胞小片贝时,主要产生基色为白色的珍珠,而用黑蝶贝作细胞小片贝时,则主要产生黑色珍珠。张根芳等[29]研究发现,提供外套膜细胞小片的珍珠贝贝壳珍珠质层的颜色会影响到珍珠的颜色。珍珠的颜色是决定珍珠品质和价格的主要因素之一,所以,贝壳珍珠质层颜色被列为合浦珠母贝育种的重要性状。近年来的研究表明,珍珠颜色与微量元素有着密不可分的关系[30-33],不同质量分数的金属元素与卟啉结合可导致珍珠质层颜色差异;不同颜色珍珠的珍珠质层文石板片厚度差异较大,且Mg、Fe、Zn和Mn质量分数越高珍珠单个文石板片就越厚[16]。单因素方差分析的结果显示(表 2),经过连续5代人工选育,7种金属元素的质量分数在4种壳色合浦珠母贝珍珠质层中存在显著差异,4个壳色合浦珠母贝珍珠质层均可能产生颜色差异。红壳珍珠质层中Fe、K、Mn、Zn的质量分数均与其余壳色珍珠质层间存在显著差异,且Fe和Mn在红壳珍珠质层中质量分数最高(表 2),因而红壳合浦珠母贝更有可能产生与其余3种壳色贝不同颜色的珍珠质层。
-
![]()
图 1 4个壳色合浦珠母贝珍珠质层和棱柱层的金属元素质量分数分布
Figure 1. Distribution of each metallic element in prismatic layer and nacreous layer of shells from four shell color strains of P.fucata
表 1 4种壳色合浦珠母贝贝壳棱柱层金属元素质量分数
Table 1 Metallic element contents in prismatic layer of shells from four shell color strains of P.fucata
mg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) 黑壳(B) 78.32±0.54a 162.57±21.47a 7 259.69±87.27a 13.45±0.81a 6 226.10±20.93a 825.75±1.86a 23.07±0.47a 金壳(G) 58.72±11.51a 224.38±15.44b 7 674.63±165.29b 12.57±0.28a 6 233.30±35.18a 830.27±9.66a 27.33±3.28ab 红壳(R) 138.66±10.56b 200.92±9.97b 7 378.63±31.03a 17.85±0.36b 5 657.00±179.97b 876.11±20.84b 30.31±0.47bc 白壳(W) 69.99±4.31a 182.07±10.02c 7 374.81±36.14a 15.67±0.40c 5 872.20±195.92b 851.69±3.82c 41.28±8.27d 平均值average 86.42±6.73 192.49±14.23 7 421.94±79.93 14.91±0.46 5 997.15±108.00 845.96±9.05 30.50±3.12 注:同列不同小写字母表示差异显著( P<0.05)
Note:Values with different letters in the same column have significant difference (P<0.05). 下载: 导出CSV
下载: 导出CSV
表 2 4种壳色合浦珠母贝贝壳珍珠质层金属元素质量分数
Table 2 Metallic element contents in nacreous layer of shells from four shell color strains of P.fucata
mg · kg-1 群体
strain元素element 铁(Fe) 钾(K) 镁(Mg) 锰(Mn) 钠(Na) 锶(Sr) 锌(Zn) P-B 48.56±2.36a 203.36±9.28a 297.33±12.22a 2.99±0.015a 7 992.04±150.69a 954.75±17.16a 56.44±2.62a P-G 67.77±5.68b 203.16±14.69a 516.95±38.18b 2.88±0.002b 8 354.21±181.37a 988.45±5.01b 50.22±3.65a P-R 73.41±8.80c 163.62±2.44b 387.18±16.01c 3.47±0.268c 7 853.86±88.11b 967.71±20.24ab 36.73±1.73b P-W 48.26±5.92a 139.99±11.94c 432.85±133.36abc 2.81±0.295ab 7 542.58±66.52b 911.93±19.58c 28.90±0.87c 平均值average 59.50±5.69 177.53±9.59 408.58±49.94 3.04±0.145 7 935.67±121.67 955.71±15.50 43.07±2.22 注:P-B、P-G、P-R和P-W分别代表合浦珠母贝黑壳、金壳、红壳和白壳珍珠质层。同列不同小写字母表示差异显著(P<0.05)
Note:P-B,P-G,P-R and P-W represent nacreous layers of black,gold,red,white shells in pearl oyster,respectively. Values with different letters in the same column have significant difference (P<0.05).
下载: 导出CSV
-
[1] 龙敏明, 黄桂菊, 邹记兴, 等. 育珠对合浦珠母贝N 19和Prismalin-14基因表达水平的影响[J]. 南方水产科学, 2013, 9(5): 58-63. doi: 10.3969/j.issn.2095-0780.2013.05.010 [2] 张根芳, 许式见, 方爱萍. 三角帆蚌(Hyriopsis cumingii)外套膜无核珍珠颜色成因的育珠实验[J]. 海洋与湖沼, 2013, 44(2): 531-536. doi: 10.11693/hyhz201302042042 [3] 符韶, 谢绍河, 邓岳文, 等. 利用马氏珠母贝4个壳色系F3培育厚层优质珍珠[J]. 水产学报, 2012, 36(9): 1418-1424. doi: 10.3724/SP.J.1231.2012.27905 [4] 王庆恒, 逯云召, 邓岳文, 等. 马氏珠母贝生长性状与珍珠质量和珍珠层厚度的相关分析[J]. 广东海洋大学学报, 2013, 33(3): 18-21. doi: 10.3969/j.issn.1673-9159.2013.03.003 [5] TAYALE A, GUEGUEN Y, TREGUIER C, et al. Evidence of donor effect on cultured pearl quality from a duplicated grafting experiment on Pinctada margaritifera using wild donors[J]. Aquat Living Resour, 2012, 25(3): 269-280. doi: 10.1051/alr/2012034
[6] 王海. 珍珠与马氏珠母贝表型性状相关性以及矿化基因在珍珠囊和外套膜中的定量表达研究[D]. 海口: 海南大学, 2013. [7] KY C L, BLAY C, SHAM-KOUA M, et al. Family effect on cultured pearl quality in black-lipped pearl oyster Pinctada margaritifera and insights for genetic improvement[J]. Aquat Living Resour, 2013, 26(2): 133-145. doi: 10.1051/alr/2013055
[8] KY C L, BLAY C, SHAM-KOUA M, et al. Indirect improvement of pearl grade and shape in farmed Pinctada margaritifera by donor "oyster" selection for green pearls[J]. Aquaculture, 2014, 432(S1): 154-162. https://www.sciencedirect.com/science/article/pii/S0044848614002178
[9] ACOSTA-SALMÓN H, MARTÍNEZ-FERNÁNDEZ E, SOUTHGATE P C. Use of relaxants to obtain saibo tissue from the blacklip pearl oyster (Pinctada margaritifera) and the Akoya pearl oyster (Pinctada fucata)[J]. Aquaculture, 2005, 245(1/2/3/4): 167-172. https://www.sciencedirect.com/science/article/pii/S0044848604007264
[10] 王照旗, 韩学凯, 白志毅, 等. 三角帆蚌紫色选育系1龄阶段内壳色及生长性状的遗传参数估计[J]. 水产学报, 2014, 38(5): 644-650. https://www.cnki.com.cn/Article/CJFDTotal-SCKX201405004.htm [11] 张文府. 马氏珠母贝四种壳色选育系中育珠贝家系和小片贝家系的选育研究[D]. 湛江: 广东海洋大学, 2013. [12] 侯巍巍, 白金泉, 吴翠敏. 卟啉及金属卟啉化合物的应用研究新进展[J]. 广东化工, 2014, 41(2): 44-45. doi: 10.3969/j.issn.1007-1865.2014.02.023 [13] 张根芳, 叶容晖, 方爱萍. 珍珠颜色和贝壳珍珠层颜色研究进展[J]. 动物学杂志, 2014, 49(1): 137-144. https://www.cnki.com.cn/Article/CJFDTotal-BIRD201401024.htm [14] 张蕴韬. 卟啉及金属卟啉对珍珠颜色的贡献及致色机理研究[D]. 北京: 中国地质大学, 2006. [15] 严俊, 胡仙超, 王巨安, 等. 不同颜色的淡水养殖珍珠呈色机理研究[J]. 岩矿测试, 2013, 32(2): 263-268. doi: 10.3969/j.issn.0254-5357.2013.02.014 [16] 杨磊, 欧阳茜茜, 李思东, 等. 中国海水珍珠Mg、Fe、Zn和Mn含量变化及其对珍珠微观结构的影响[J]. 矿物学报, 2014, 34(3): 416-420. https://www.cnki.com.cn/Article/CJFDTotal-KWXB201403019.htm [17] 隋欣, 吴海铭, 王宝辉, 等. ICP-AES应用进展及展望[J]. 牡丹江师范学院学报: 自然科学版, 2014(1): 25-27. doi: 10.3969/j.issn.1003-6180.2014.01.013 [18] 王茂林, 张秀梅, 高天翔, 等. 水体钙离子质量浓度对褐牙鲆幼鱼鱼体和耳石元素成分的影响[J]. 南方水产科学, 2013, 9(3): 31-38. doi: 10.3969/j.issn.2095-0780.2013.03.006 [19] 木士春, 马红艳. 养殖珍珠微量元素特征及其对珍珠生长环境的指示意义[J]. 矿物学报, 2001, 21(3) : 551-553. doi: 10.3321/j.issn:1000-4734.2001.03.069 [20] 林静瑜, 谢莉萍. EDTA对合浦珠母贝碱性磷酸酶活性的影响[J]. 集美大学学报, 2004, 9(4): 313. doi: 10.3969/j.issn.1007-7405.2004.04.006 [21] 高玲, 曹洪玉, 刘阁. 4种金属卟啉配合物与血清白蛋白作用的光谱研究[J]. 化学试剂, 2012, 34(2): 108-112. doi: 10.3969/j.issn.0258-3283.2012.02.004 [22] MICHIO S, HIROMICHI N. Mollusk shell structures and their formation mechanism[J]. Can J Zool, 2013, 91(6): 349-366. doi: 10.1139/cjz-2012-0333
[23] JACOB D E, SOLDATI A L, WIRTH R, et al. Nanostructure, composition and mechanisms of bivalve shell growth[J]. Geochimicaet Cpsmochimica Acta, 2008, 72(22): 5401-5415. doi: 10.1016/j.gca.2008.08.019
[24] 王慧, 封余军, 郑碧华, 等. 等离子体发射光谱分析珍珠贝贝壳结晶的微量元素[J]. 湖北农学院学报, 1998, 18(2): 142-144. https://www.cqvip.com/qk/91582X/199802/3080694.html [25] 夏静芬, 钱国英, 陈亮, 等. 珍珠粉和贝壳粉的化学成分和结构特征分析[J]. 化学研究与应用, 2010, 22(11): 1467-1471. doi: 10.3969/j.issn.1004-1656.2010.11.025 [26] 孙维萍, 潘建明, 刘小涯, 等. 浙江沿海贝类体内重金属元素含量水平与评价[J]. 海洋学研究, 2010, 28(4): 43-49. doi: 10.3969/j.issn.1001-909X.2010.04.006 [27] 逯云召. 金属离子对马氏珠母贝珍珠层颜色和金属硫蛋白基因表达的影响[D]. 湛江: 广东海洋大学, 2012. [28] MCGINTY E L, EVANS B S, TAYLOR T J, et al. Xenografts and pearl product ion in two pearl oyster species, P. maxima and P. margaritifera: effect on pearl quality and a key to understanding genetic contribution[J]. Aquaculture, 2010, 302(3/4): 175-181.
[29] 张根芳, 许式见, 方爱萍. 组织小片对三角帆蚌外套膜无核珍珠颜色成因的影响[J]. 水生生物学报, 2013, 37(3): 581-587. doi: 10.7541/2013.66 [30] 郝之奎, 王嫣, 顾志峰, 等. 珍珠颜色的研究进展[J]. 安徽农业科学, 2007, 35(11): 3260-3261. doi: 10.3969/j.issn.0517-6611.2007.11.061 [31] 刘娟花, 马孝甜, 苗东亮, 等. 珍珠致色因素研究概况[J]. 江苏农业科学, 2011(1): 289-292. doi: 10.3969/j.issn.1002-1302.2011.01.105 [32] 高珊. 广西马氏贝珍珠颜色特征研究和染色实验[D]. 北京: 中国地质大学, 2013. [33] 严俊, 邓小琼, 胡丹静, 等. 淡水养殖珍珠棱柱层的微结构形貌特征[J]. 南方水产科学, 2013, 9(1): 48-52. doi: 10.3969/j.issn.2095-0780.2013.01.008 -
期刊类型引用(20)
1. 贺文聪,黄庚胤,梁健,陈丽梅,郭永军. 四角蛤蜊(Mactra veneriformis)壳色分布及贝壳金属元素含量特征分析. 天津农业科学. 2024(05): 35-41 .  百度学术
百度学术
2. 李杰,荣克明,杨丹荣,李雁宜,纪沛杉,马良骁,马徐发. 酸化牡蛎壳粉对铜绿微囊藻和小球藻生长的影响. 中国环境科学. 2023(S1): 197-204 . 百度学术
3. 魏海军,邓正华,陈明强,邓林琪,谭春明,李有宁,王雨,王爱民. 不同壳色合浦珠母贝生产性能评估及其雌雄群体间的差异. 南方农业学报. 2020(02): 412-420 . 百度学术
4. 张善发,贺承章,李俊辉,邓岳文,杨创业. 马氏珠母贝金黄壳色选育群体与养殖群体不同组织中矿物质元素的分析与评价. 海洋通报. 2020(03): 357-362 . 百度学术
5. 金武,闻海波,马学艳,何义进,邴旭文. 2种壳色梨形环棱螺贝壳7种金属元素质量分数比较分析. 中国农学通报. 2019(21): 148-151 . 百度学术
6. 赵永强,王安凤,陈胜军,杨贤庆,李来好,吴燕燕,杨少玲. 米曲霉和鲁氏酵母协同发酵优化合浦珠母贝肉酶解液风味. 食品与发酵工业. 2019(15): 115-120 . 百度学术
7. 唐何娜,杨磊,刘艳茹,陈进. 纯水和棕榈酸体系中Mg~(2+)/Sr~(2+)对碳酸钙结晶行为的影响. 电子显微学报. 2019(06): 639-649 . 百度学术
8. ZHU Yijing,LI Qi,YU Hong,KONG Lingfeng. Biochemical Composition and Nutritional Value of Different Shell Color Strains of Pacific Oyster Crassostrea gigas. Journal of Ocean University of China. 2018(04): 897-904 . 必应学术
9. 朱怡静,李琪,张景晓. 5种壳色长牡蛎不同组织中金属元素的分析与评价. 水产学报. 2018(09): 1358-1366 . 百度学术
10. 刘先进,陈胜军,李来好,杨贤庆,黄卉. 四种鲍鱼肌肉营养成分分析与品质评价. 食品与发酵工业. 2018(05): 227-231 . 百度学术
11. 张磊,王锦旭,杨贤庆,魏涯,杨少玲. 海洋动物多糖的研究进展. 食品工业. 2018(01): 211-215 . 百度学术
12. 王祎哲,邓正华,王雨,李有宁,于刚,黄桂菊,陈明强. 合浦珠母贝不同壳色选育系F6数量性状的相关性和通径分析. 中国水产科学. 2018(05): 988-997 . 百度学术
13. 张磊,王锦旭,杨贤庆,魏涯,杨少玲. 合浦珠母贝糖胺聚糖泡腾片制备工艺优化及其质量分析. 大连海洋大学学报. 2018(05): 644-650 . 百度学术
14. 王德清,李海梅,黄桂菊,张博,范嗣刚,刘宝锁,苏家齐,喻达辉. 合浦珠母贝章鱼胺受体(OAR)基因的克隆与表达分析. 南方水产科学. 2017(01): 58-65 . 本站查看
15. 王安凤,赵永强,陈胜军,李来好,杨贤庆,吴燕燕,杨少玲,李春生. 响应面法优化合浦珠母贝肉水解工艺. 食品与发酵工业. 2017(11): 165-171 . 百度学术
16. 战欣,陈琼,顾志峰,石耀华,王爱民. 马氏珠母贝贝壳珍珠质层厚度性状与微卫星标记的相关性分析. 热带生物学报. 2016(03): 290-295 . 百度学术
17. 周代志,黄桂菊,刘宝锁,张博,姜松,邓正华,喻达辉. 合浦珠母贝骨形态发生蛋白10(BMP10)基因的克隆与表达分析. 南方水产科学. 2016(06): 83-90 . 本站查看
18. 陈明强,刘宝锁,严俊贤,郭华阳,吴开畅,李有宁. 合浦珠母贝4种壳色选育系主要性状的比较分析. 南方水产科学. 2016(05): 118-122 . 本站查看
19. 王安凤,赵永强,陈胜军,李来好,杨贤庆,吴燕燕,胡晓. 栅栏技术制备高水分即食合浦珠母贝肉工艺的研究. 食品工业科技. 2016(21): 183-188 . 百度学术
20. 程珊,侯俊杰,肖凌,聂晶. 珍珠明目滴眼液中16种元素的测定与分析. 中国实验方剂学杂志. 2016(19): 53-58 . 百度学术
其他类型引用(18)
计量
- 文章访问数: 3326
- HTML全文浏览量: 146
- PDF下载量: 2001
- 被引次数: 38
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
