
Underwater irradiance and optimal allocation of 2 kW straight type fish aggregation lamps
-
摘要:
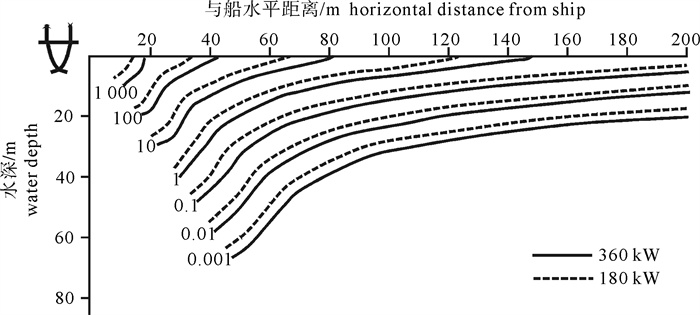
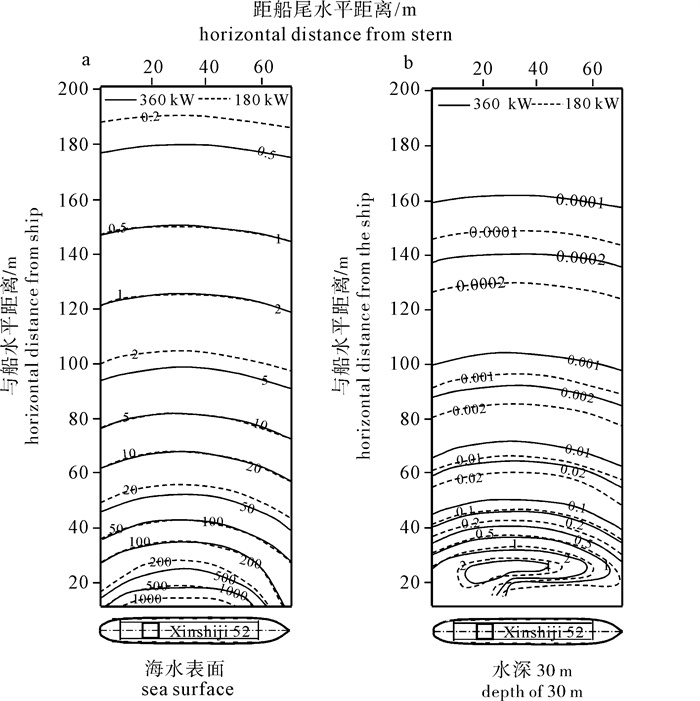
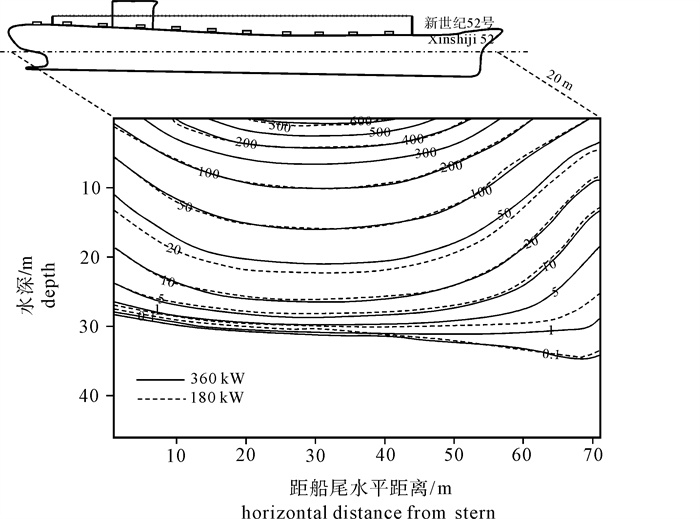
根据2 kW直筒型集鱼灯的实测配光分布,以大型光诱鱿钓渔船“新世纪52号”为例,对该船的水中照度分布进行了计算。结果表明:渔船集鱼灯总功率为360 kW时,10.000 lx的等照度曲线水平方向最远约为80 m,水深方向为32 m;0.100 lx照度最深可约至50 m,最远约为260 m;当总功率减为180 kW时,等照度曲线所在水层仅减少约5 m。以0.010 lx为诱集鱿鱼的最低照度,集鱼灯功率从180 kW增加到360 kW,有效诱集距离从620 m增加到750 m,约能增加21.1%;如能将该船的灯高增大0.5 m,则可增加12.0%的有效水体体积;如能将灯距增大0.05 m,则可增加4.4%的有效水体体积。
Abstract:Based on the actual measurement of 2 kW straight type fish aggregation lamps, we calculated the underwater illumination distribution of the squid jigging vessel Xinshiji No.52. The results show that when the total power was 360 kW, the iso-illuminance curve contours of 10.000 lx were at the depth of 32 m; in horizontal direction, it reached 80 m. The curve contours of 0.100 lx were at 50 m vertically and 260 m horizontally. When the total power was 180 kW, the curve contours reduced about 5 m in depth. Regarding 0.010 lx as the lowest limit to attract squid, if the power was increased from 180 kW to 360 kW, attracting distance could be increased from 620 m to 750 m, and 21.1% of effective attracting distance increased horizontally. If the lamps′ height increased for 0.5 m, 12.0% of the effective water volume increased. If the lamps′ space increased for 0.05 m, 4.4% of the effective water volume increased.
-
自然界由于食物存在空间上的分布不均匀、季节的更替、环境条件的剧变等原因,水产动物在生活周期的一定阶段将面临食物资源的缺乏而受到饥饿胁迫。很多水产动物都具备能够忍受较长时间饥饿的生理功能,为维持自身生命,它们将减少代谢率甚至通过消耗自身组织的贮存物质等行为来适应短期的食物不足问题[1]。
水产动物在胚胎期可以依靠卵黄或母体提供营养,但在生长繁殖期所需的食物和能量必须从外界环境中获得。食物短缺将严重制约水产动物的成活、生长、发育和繁殖等生命活动[2]。谢小军等[3]及胡麟和吴天星[4]综述了饥饿对鱼类代谢、身体组成、形态结构、行为、繁殖习性和成活的影响,并指出研究饥饿对鱼类生理学状况的影响有助于了解鱼类适应饥饿胁迫的生态对策,对水产动物自然资源的保护、苗种培育及人工养殖等方面具有重要的指导意义。研究饥饿对海产经济动物生理生态学的影响,不仅可以揭示其适应饥饿胁迫的生理学对策,也可以为研发高效投喂技术提供理论依据[5]。
近年来,国内外学者对乌贼类的种群变动、资源状况、洄游分布、环境耐受性、摄食、生长、营养价值、乌贼墨及其养殖等方面的研究已有报道[6-10],但对曾是中国四大渔业之一的曼氏无针乌贼(Sepiella maindrona)生理方面研究报道还较少。元冬娟等[11]指出脂肪酸在贝类的天然免疫反应、寒冷的适应性、繁殖和胚胎发育等生理过程中都起着关键的作用。文章研究了饥饿胁迫下曼氏无针乌贼肝脏与卵巢中的脂肪酸组成变化,以期了解饥饿这一特殊条件对曼氏无针乌贼生长、繁殖的影响,为曼氏无针乌贼营养需求、能量代谢及生理学研究提供理论资料。
1. 材料与方法
1.1 试验设计
试验于2007年4~5月在宁海双盘涂水产养殖基地进行。试验曼氏无针乌贼选自奉化市乌贼育苗养殖基地,初始平均胴长为(10.27±1.81)cm,平均体质量为(115.8±12.6)g且为卵巢成熟的雌性乌贼。暂养于设有遮阳布的网箱(4 m×2 m×2 m)中,每天早、晚测定水质指标,投喂鲜活小杂鱼并及时清除网箱内残饵,观察乌贼的活动情况。水温(18±1.5) ℃、盐度22.5±0.3、pH 8.2±0.1、溶氧(5.3±0.4)mg · L-1。适应15 d后,挑选规格整齐、体色正常的健康曼氏无针乌贼30只分3个网箱放养。将3组乌贼分别设计不同的禁食处理后同一天取样,即空腹处理组(禁食1 d)、饥饿12 d(禁食13 d)、饥饿24 d(禁食25 d),每个网箱内各取5只乌贼活体运至实验室并及时取其组织。曼氏无针乌贼的体质量和胴体长在运到双盘涂开始暂养前测定。由于曼氏无针乌贼离水后极容易喷墨而影响活力,因此,该试验的称量过程是先在烧杯中放入少量自然海水,在精确度为0.1 g的电子天平上去皮质量后再将曼氏无针乌贼移入该烧杯称量,记录湿质量即为体质量,用普通直尺测量乌贼胴体长,精确度为0.1 cm。
1.2 样本制备及测定
将饥饿处理的活体乌贼用吸水纸吸干体表水分,测定体质量及胴体长后迅速在冰盘上解剖,取出其肝脏、卵巢,用预冷生理盐水快速冲洗,并用脱脂棉小心揩干,称量,低温冷冻干燥后用于脂肪酸测定。测定工具同1.1。
脂肪酸(GC-MS)分析具体参数设定。1)气相色谱条件:进样口温度250 ℃,载气氦(He),柱流速0.81 mL · min-1,柱前压73.0 kPa,柱起始温度150 ℃,保持3.5 min,以20 ℃ · min-1升至200 ℃,保持5 min,再以5 ℃ · min-1升至280 ℃,保持15 min。分流进样1 μL,分流比50 : 1;2)质谱条件:电子轰击源(EI),电子能量70 eV,离子源温度200 ℃,接口温度250 ℃,全程离子碎片扫描(SCAN)模式,质量扫描范围40~650,溶剂延迟3.5 min。测定仪器及柱子:QP 2010GC-MS仪(日本SHIMADZU公司出品),30 m×0.25 mm×0.25 μm SPB-50色谱柱(美国SUPELCO公司出品)。
1.3 数据统计分析
所有数据均以平均值±标准差(Mean±SD)表示,用Excel进行整理统计,采用SPSS 13.0软件进行单因素方差分析(One-Way ANOVA),达显著差异后(P < 0.05),则采用Duncan检验进行组间差异的多重比较。
2. 结果
空腹处理组、饥饿12和24 d处理组的曼氏无针乌贼个体大小出现显著差异(P < 0.05),空腹处理组曼氏无针乌贼个体大,活力好,其生殖腔内可见大量分散、透明的成熟卵子;饥饿12 d组的乌贼个体较空腹处理组小,卵巢中的卵子颗粒也不饱满;饥饿24 d组的乌贼身体明显消瘦、弱小、反应迟钝,其卵巢严重萎缩,发育不良,具体形态学指标见表 1。曼氏无针乌贼在饥饿12和24 d后的体质量、胴体长、肝脏质量及卵巢质量均有显著下降(P < 0.05)。
表 1 饥饿对曼氏无针乌贼形态学的影响Table 1 Effects of starvation on morphology of S.maindroni处理组group 体质量/g body weight 胴体长/cm carcass length 肝脏质量/g liver weight 卵巢质量/g ovary weight 空腹组fasting group 159.8±17.4a 11.14±2.31a 3.1±0.6a 14.5±2.9a 饥饿12 d starvation 123.4±25.9b 9.56±1.97b 2.5±0.7b 10.1±1.6b 饥饿24 d starvation 87.1±12.8c 7.42±1.75c 1.3±0.4c 5.6±2.1c 注:不同字母表示处理间差异显著(P < 0.05),下同
Note:Different letters represent significant difference (P < 0.05).The same below.2.1 饥饿前后曼氏无针乌贼肝脏脂肪酸变化
空腹处理组、饥饿12和24 d处理组的肝脏脂肪酸含量测定结果见表 2。饥饿12 d时乌贼肝脏中的饱和脂肪酸(saturated fatty acids,SFA)和单不饱和脂肪酸(monounsaturated fatty acid,MUFA)比率较空腹处理组显著下降(P < 0.05),而多不饱和脂肪酸(polyunsaturated fatty acid,PUFA)和高不饱和脂肪酸(highly-unsaturated fatty acid,HUFA)比率均有显著提高(P < 0.05);饥饿24 d时未检测到其肝脏中的C12:0,∑SFA、∑MUFA和∑n-3/n-6 PUFA的比率较空腹处理组显著下降(P < 0.05),而∑PUFA+∑HUFA、DHA/EPA比率较空腹处理组有显著上升(P < 0.05)。
表 2 饥饿对曼氏无针乌贼肝脏脂肪酸组成的影响Table 2 Effect of starvation on fatty acid composition of liver in S.maindroni脂肪酸fatty acids 空腹组fasting group 饥饿时间/d starvation time 12 24 12:0 0.042±0.002a 0.033±0.006b - 14:0 1.231±0.010b 3.533±0.012a 0.853±0.015c 15:0 1.860±0.009a 0.663±0.040b 0.373±0.025c 16:0 27.636±0.014a 24.373±0.021b 12.757±0.051c 17:0 3.344±0.017a 1.467±0.006b 1.780±0.010b 18:0 19.130±0.010a 12.457±0.049b 18.500±0.026a 19:0 0.308±0.008a 0.193±0.006b 0.357±0.006a 20:0 2.953±0.015a 0.663±0.006c 2.123±0.025b 21:0 - - 0.053±0.006 22:0 0.241±0.012b 0.063±0.006c 0.323±0.006a 23:0 0.099±0.009 - - ∑SFA 56.844±0.011a 43.445±0.025b 37.119±0.029c 16:1(n-7) 4.963±0.025a 0.510±0.010b 0.417±0.021b 18:1(n-9) 4.087±0.021a 3.530±0.017b 1.030±0.017c 18:1(n-7) 6.488±0.033a 1.767±0.012b 1.287±0.006b 20:1(n-9) 1.532±0.010a 0.290±0.010c 0.470±0.020b ∑MUFA 17.790±0.027a 6.097±0.014b 3.204±0.016c 18:3(n-3) - 0.070±0.010b 0.233±0.006a 20:5(n-3)EPA 8.886±0.044b 15.047±0.045a 16.827±0.025a 22:5(n-3) 0.765±0.004c 0.883±0.012b 1.810±0.010a 22:6(n-3)DHA 7.015±0.018c 24.440±0.072a 17.220±0.026b 20:4(n-6) 3.619±0.018b 3.747±0.021b 9.363±0.038a 22:4(n-6) - - 1.330±0.026 ∑PUFA+HUFA 20.285±0.015b 44.187±0.039a 46.783±0.029a DHA/EPA 0.789±0.002c 1.624±0.001a 1.023±0.002b ∑n-3 PUFA 16.666±1.228c 40.440±3.074a 36.090±2.419b ∑n-6 PUFA 3.619±0.010b 3.747±0.012b 10.693±1.796a ∑n-3/n-6 PUFA 4.603±0.002b 10.794±0.049a 3.375±0.001c 注:∑SFA. 饱和脂肪酸总百分比;∑MUFA. 单不饱和脂肪酸总百分比;∑PUFA. 多不饱和脂肪酸总百分比;∑HUFA. 高不饱和脂肪酸总百分比;下同
Note:∑SFA. total percentage of saturated fatty acid;∑MUFA. total percentage of monounsaturated fatty acid;∑PUFA. total percentage of polyunsaturated fatty acid;∑HUFA. total percentage of highly-unsaturated fatty acid. The same below.2.2 饥饿前后曼氏无针乌贼卵巢脂肪酸变化
空腹处理组、饥饿12和24 d处理组的卵巢脂肪酸含量测定结果见表 3。与肝脏相比,乌贼卵巢中的各脂肪酸含量在饥饿12 d时与空腹处理组相比变化程度较小,其中∑SFA、∑MUFA、∑n-3/n-6 PUFA比率及组成在饥饿12 d时与空腹处理组无显著变化(P > 0.05),但∑PUFA+∑HUFA、∑n-3/n-6 PUFA和∑n-6 PUFA比率较空腹处理组有显著下降(P < 0.05),且未检测到EPA;饥饿24 d卵巢中的∑SFA、DHA/EPA、∑n-3/n-6 PUFA比率较空腹处理组有极显著下降(P < 0.01),而∑MUFA、∑PUFA+∑HUFA和∑n-6 PUFA比率显著高于空腹处理组(P < 0.05)。可见饥饿12 d时,曼氏无针乌贼卵巢内的各脂肪酸比率变化程度较小,而饥饿24 d时,曼氏无针乌贼卵巢内的脂肪酸比率变化趋势跟肝脏内的变化趋势一致。
表 3 饥饿对曼氏无针乌贼卵巢脂肪酸组成的影响Table 3 Effect of starvation on fatty acid composition of ovary in S.maindroni脂肪酸fatty acids 空腹组fasting group 饥饿时间/d starvation time 12 24 12:0 0.053±0.003a 0.023±0.006 0.043±0.006b 14:0 3.067±0.015a 3.600±0.046a 1.317±0.021b 15:0 0.294±0.005b - 0.477±0.025a 16:0 28.156±0.014a 25.547±0.050a 1.213±0.015b 17:0 0.997±0.006b 1.837±0.015a 0.257±0.006 18:0 11.737±0.006a 13.153±0.050a 0.090±0.010b 19:0 0.105±0.005b 0.273±0.006a 0.293±0.006a 20:0 0.856±0.005a 0.787±0.012a 0.123±0.006b 22:0 0.076±0.005c 0.103±0.015b 0.163±0.006a ∑SFA 45.341±0.009a 45.323±0.027a 3.976±0.012b 16:1(n-7) - 0.420±0.017b 1.130±0.026a 18:1(n-9) 3.806±0.020b 2.837±0.032b 18.047±0.047a 18:1(n-7) 1.191±0.010b 1.733±0.023a 1.203±0.006b 20:1(n-9) 0.197±0.011b 0.393±0.006a 0.323±0.006a ∑MUFA 5.194±0.013b 5.776±0.021b 20.703±0.019a 18:3(n-3) 0.176±0.005b 0.063±0.006c 6.067±0.042a 20:5(n-3) 10.652±0.053b - 14.153±0.046a 22:5(n-3) 0.584±0.003b 0.950±0.010a 0.836±0.006a 22:6(n-3) 28.441±0.079a 26.210±0.010a 18.653±0.137b 20:4(n-6) 4.461±0.022b 3.070±0.044b 10.580±0.069a 22:4(n-6) - - 0.657±0.025 ∑PUFA+HUFA 44.314±0.035b 30.293±0.017c 50.946.±0.074a DHA/EPA 2.670±0.007a - 1.318±0.007b ∑n-3 PUFA 39.853±3.456a 27.223±4.286b 39.710±2.087a ∑n-6 PUFA 4.461±0.013b 3.070±0.025c 11.237±2.219a ∑n-3/n-6 PUFA 8.924±0.005a 8.869±0.129a 3.534±0.011b 3. 讨论
面对食物缺乏,动物将调整自身的代谢过程以适应饥饿胁迫,许多水产动物都能忍受较长时间的饥饿。饥饿状态下水产动物的一些外部形态特征和内部解剖性状产生某些特定的变化,如动物体逐渐消瘦、头大体小、背薄尾尖、肠管变细、肝胰脏缩小和卵巢发育不良等[12-13]。该试验中,饥饿24 d后的曼氏无针乌贼在形态上就发生了明显变化,个体明显小于空腹组,体质量显著下降(P < 0.05),解剖内脏也发现其肝脏组织明显萎缩和卵巢组织严重发育不良,这与很多学者研究的结果相似[14-15]。说明曼氏无针乌贼至少能够忍受24 d内的饥饿胁迫,但在饥饿胁迫情况下,曼氏无针乌贼必须动用体内的储存物质来维持自身生命,已不能保证其生长发育。关于饥饿胁迫下曼氏无针乌贼各组织的组织学研究将有待于进一步深入。
饥饿胁迫时,水产动物代谢发生适应性生理变化,多数水产动物以脂肪和糖元作为能源物质,调节身体各种酶活性,以达到积极利用体内的贮存物质,得以维持生命[16]。一般情况下,作为主要贮能物质的糖元、脂肪和蛋白质在饥饿过程中将会不同程度地被消耗。通过对性腺发育成熟的曼氏无针乌贼作不同程度的饥饿处理,测定其肝脏和卵巢中各脂肪酸含量比率,可以分析其相应的生化组成,获得饥饿期间动物使用身体贮能物质的变化资料。
饥饿期间动物体动用机体内能源物质有先有后,而对不同器官,动用贮能物质也有先后次序。该试验结果发现,饥饿过程中曼氏无针乌贼肝脏中的∑SFA和∑MUFA作为维持生命的能源物质首先被利用,并且肝脏中的∑SFA和∑MUFA其中一部分也可能被转移到卵巢中,致使其比率在饥饿12 d时明显降低(P < 0.05)。此外,卵巢的营养由于无法完成繁殖活动而逐步被吸收,其∑n-3 PUFA和∑n-6 PUFA可能是首先被转移的对象,同时从肝脏中获得了∑SFA和∑MUFA来保证卵巢的正常发育,因而导致卵巢中∑n-3 PUFA和∑n-6 PUFA比率有所下降,而∑SFA和∑MUFA组成在饥饿12 d时受饥饿胁迫的影响较小,其比率及组成没有明显变化(P > 0.05)。这一结果同时表明,曼氏无针乌贼各器官组织的功能不同,肝脏组织起贮存能源物质的作用,对饥饿等外界条件敏感,饥饿时优先动用其中的能源物质,而性腺(卵巢)在饥饿等恶劣的条件下,首先会得到保护,也有可能是肝脏储存的脂肪一部分用于能量被消耗,而另一部分被用于卵巢的发育,即卵巢的脂肪积累在没有外界食物来源时主要来源于肝脏中储存脂肪的转运。但随着饥饿胁迫时间的延长,因肝脏中的脂肪将不足以提供给卵巢,为达到维持自身生命的目的,当饥饿24 d后曼氏无针乌贼只能动用卵巢中的∑SFA来保护自己,故∑SFA比率明显降低,这可能是因为曼氏无针乌贼肝脏中所贮存的能量已经提供到了极限,卵巢的营养由于无法完成繁殖活动,逐步被吸收来维持自身的生命活动。JEZIERSKA等[17]研究虹鳟(Salmo gairdneri)指出,饥饿胁迫降低了单不饱和脂肪酸的含量,从而导致了饱和脂肪酸含量的升高。
谢小军等[3]研究报道了饥饿过程中鱼类对其体内不同种类脂肪酸的利用顺序有一定的规律,即首先利用SFA,然后MUFA,最后才动用HUFA。FALK等[18]和MOURENTE[19]研究也发现,n-6 PUFA和n-3 PUFA在各自组织中被优先保存下来,而SFA和MUFA作为仔鱼吸收卵黄内源营养发育阶段的重要能源被首先利用。KOVEN等[20]发现金鲷(Sparus aurata)仔鱼在饥饿过程中脂肪酸损失速率的排列顺序为∑n-6 PUFA>∑n-9 PUFA>∑n-3 PUFA。SATOH等[21]对饥饿胁迫下尼罗罗非鱼(Tilapia nilotica)脂肪酸含量变化的研究中也发现各种类脂肪酸损失速率有类似排列顺序。DESLIVA等[22]提出饥饿影响鱼体组织特别是肝脏和肌肉的脂肪酸组成及内脏的脂肪积累,且不同鱼类脂肪酸的动用也存在着差异。该试验结果也证明以上学者的观点,即饥饿过程中曼氏无针乌贼肝脏中∑SFA和∑MUFA的比率在饥饿12 d时含量降低明显,而∑PUFA+∑HUFA在饥饿12 d时的比率显著升高。当肝脏中的∑SFA和∑MUFA被利用到一定程度后(饥饿24 h),卵巢中的∑SFA和∑MUFA比率也出现极显著下降(P < 0.01)。至于曼氏无针乌贼若继续长时间饥饿,其组织内的∑MUFA的比率是否将相应下降还有待于作进一步深入研究。
饥饿或限食对于控制因投饵不当引起的自身污染具有十分现实的意义。随着养殖规模的扩大,养殖水体富营养化日趋严重,导致水质恶化,溶氧减少,病害频发,对养殖产业构成了严重威胁。而在养殖过程中因投喂不合理导致残饵、动物粪便和排泄物等产生的自身污染是产生养殖水体恶化的主要原因之一。如果能够利用补偿生长现象进行科学投喂,减少饥饿期间排泄量及增加恢复生长期间食欲来减少养殖过程中的残饵量,从源头控制自身污染,就能促进水产养殖业的可持续健康发展。该试验为进一步研究曼氏无针乌贼的补偿生长奠定了基础,也为曼氏无针乌贼的高效投喂模式提供了技术支持。
-
![]()
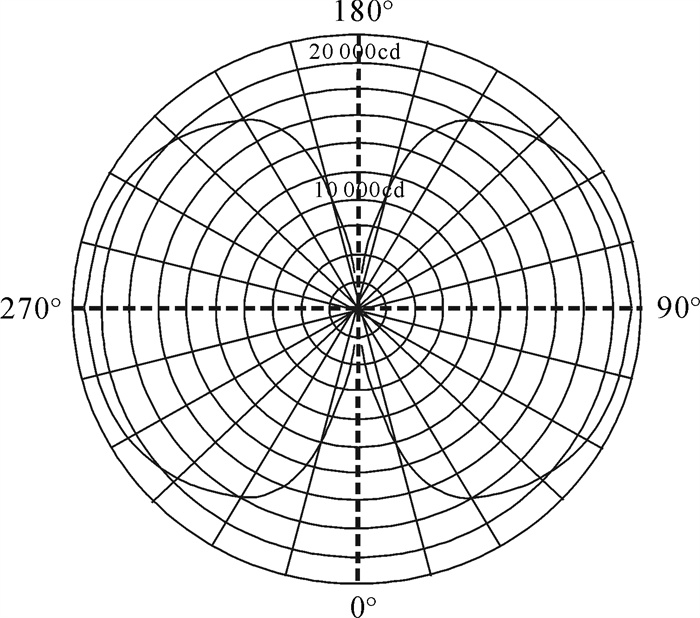
图 1 2 kW直筒型金属卤化物灯的辐射量分布
Figure 1. Distribution of radiation of 2 kW straight type metal halide lamp
![]()
图 2 2 kW直筒型金属卤化物灯(DC-2000TT)配光曲线
Figure 2. Distribution of luminous intensity of straight type metal halide lamp (DC-2000TT, 2 kW)
![]()
图 5 不同集鱼灯功率时的等照度曲线分布(水平距离20 m)
Figure 5. Contours of irradiance by different powers (horizontal distance of 20 m from boat)
表 1 2 kW型金属卤化物灯的基本参数
Table 1 Basic parameters of 2 kW metal halide lamp
型号
type功率/W
power电源电压/V
voltage光通量/Lm
luminous flux经济寿命/h
economic life全长×直径/mm
full length×diameter风阻系数
drag coefficientDC-2000TT 2 000 220 220 000 3 000 465×90 0.846×10-4  下载: 导出CSV
下载: 导出CSV
表 2 不同灯高灯距时的水中照度V0.01和V10
Table 2 V0.01 and V10 with different heights and space between of fishing lamps
灯高
height of lamp/m灯距 space
between oflamps/mV10/m3 V0.01/m3 V0.01-10/m3 6.0 0.45 30 282.52 238 156.47 207 873.95 0.50 31 932.58 255 314.90 223 382.32 0.55 31 786.55 254 725.82 222 939.27 0.60 30 599.33 252 346.96 221 747.63 0.65 32 556.37 251 757.66 219 201.29 0.70 35 674.85 249 607.23 213 932.38 6.5 0.45 30 510.27 254 464.89 223 954.63 0.50 33 672.49 269 781.34 236 108.85 0.55 36 256.37 282 792.92 246 536.56 0.60 34 094.65 274 357.40 240 262.75 0.65 34 160.46 287 137.22 252 976.76 0.70 35 985.17 272 821.19 236 836.02 7.0 0.45 38 126.38 258 974.13 220 847.74 0.50 46 011.98 310 402.34 264 390.36 0.55 41 239.93 276 618.89 235 378.96 0.60 40 220.00 300 959.21 260 739.21 0.65 40 269.49 282 190.72 241 921.23 0.70 37 064.15 276 859.17 239 795.02
下载: 导出CSV
-
[1] 孙满昌. 海洋渔业技术学[M]. 2版. 北京: 中国农业出版社, 2012. [2] 钱卫国, 官文江, 陈新军. 1 kW国产金属卤化物灯光学特性及其应用[J]. 上海海洋大学学报, 2012, 21(3): 439-444. https://xueshu.baidu.com/usercenter/paper/show?paperid=4a39b2e81342bc1239e03b734f634ee1&site=xueshu_se [3] 沙锋, 钱卫国, 吴仲琪, 等. 鲐鱼灯光围网渔船水上集鱼灯水中照度分布及优化配置的理论计算[J]. 海洋学研究, 2013, 31(1): 85-90. doi: 10.3969/j.issn.1001-909X.2013.01.011 [4] 侍炯, 钱卫国, 杨卢明. 鲐鱼灯光围网渔船合适作业间距的理论研究[J]. 南方水产科学, 2013, 9(4): 82-86. [5] 肖启华, 张丽蕊. 光诱渔业中光强分布的理论研究及其应用[J]. 上海水产大学学报, 2007, 16(6): 613-617. [6] 王飞, 钱卫国. 智利外海茎柔鱼渔场集鱼灯灯光的配置[J]. 水产学报, 2008, 32(2): 279-286. doi: 10.3321/j.issn:1000-0615.2008.02.016 [7] 戴天元, 沈长春, 冯森, 等. 光诱渔船集鱼灯的光照度分布及其适渔性能分析[J]. 福建水产, 2007, 26(1): 27-31. doi: 10.3969/j.issn.1006-5601.2007.01.008 [8] 崔浙珍, 荒川久幸, 中村善彦, 等. 日本海のィカ钓り渔场における海水の光学的水型と集鱼灯の透过特性[J]. 日本水産學會誌, 1998, 64(4): 650-657. doi: 10.2331/suisan.64.650 [9] 崔淅珍, 荒川久幸, 有远贵文, 等. 线光源モデルを用ぃた小型ィカ钓り渔船集鱼灯の水中照度分布解析[J]. 日本水産學會誌, 2003, 69(1): 44-51. doi: 10.2331/suisan.69.44 [10] 钱卫国, 陈新军, 钱雪龙, 等. 300W型LED集鱼灯光学特性及其节能效果展望[J]. 海洋渔业, 2011, 33(1): 99-105. doi: 10.3969/j.issn.1004-2490.2011.01.015 [11] 钱卫国, 陈新军, 雷林. 300W型绿光LED集鱼灯光学特性初探[J]. 大连海洋大学学报, 2012, 27(5): 471-476. doi: 10.3969/j.issn.2095-1388.2012.05.016 [12] 崔雪亮, 张伟星. 新型LED集鱼灯节能效果实船验证及推广[J]. 浙江海洋学院学报: 自然科学版, 2013, 32(2): 169-172. doi: 10.3969/j.issn.1008-830X.2013.02.015 [13] SHEN S C, HUANG H J, CHAO C C, et al. Design and analysis of a high-intensity LED lighting module for underwater illumination[J]. Appl Ocean Res, 2012, 39: 89-96. doi: 10.1016/j.apor.2012.10.006
[14] OKAMOTO T, TAKAHASHI K, OHSAWA H, et al. Application of LEDs to fishing lights for Pacific saury[J]. J Light Vis Environ, 2008;32(2): 88-92. doi: 10.2150/jlve.32.88
[15] CHOI J S, CHOI S K, KIM S J, et al. Photoreaction analysis of squids for the development of a LED-fishing lamp[C]//Proceedings of the 2nd International Conference on Maritime and Naval Science and Engineering (MN′09), Transilvania University of Brasov, Romania, September 24-26, 2009. WSEAS Press, 2009: 92-96.
[16] GUTTSAIT E M. Analysis of the illuminance provided by LED modules placed at large distances from illuminated objects[J]. J Comm Tech Electron, 2009, 54(1): 107-118. doi: 10.1134/S1064226909010100
[17] 钱卫国. 鱿钓渔业中集鱼灯的优化配置研究[D]. 上海: 上海水产大学, 2005. https://xueshu.baidu.com/usercenter/paper/show?paperid=193739b4f198e0126b37d5773f5fcf1c&site=xueshu_se&hitarticle=1 [18] 官文江, 钱卫国, 陈新军. 应用Monte Carlo方法计算水上集鱼灯向下辐照度在一类海水中的分布[J]. 水产学报, 2010, 34(10): 140-151. doi: 10.3724/SP.J.1231.2010.06723 [19] 上海海洋大学. 水上集鱼灯水下光场计算系统V1.0(登记号: 2010SR042147)[CP]. 2010. https://www.hanspub.org/reference/Reference?ReferenceID=113311 [20] 何大仁, 肖金华, 罗会明, 等. 杜氏枪乌贼趋光行为的研究[J]. 厦门大学学报: 自然科学版, 1979, 18(3): 99-104. [21] 郑美丽, 肖金华, 郑微云, 等. 曼氏无针乌贼的趋光特性[J]. 厦门大学学报: 自然科学版, 1980, 19(3): 91-99. https://www.doc88.com/p-1098606448787.html [22] 铃木恒由. スルメィカの光对すゐ生理反应と集鱼灯の省ェネ对策にっぃて[J]. 日本水产资源保护协会月报, 1986(266): 11-26. [23] 郑微云, 柴敏娟. 头足类视觉特性的电生理研究——Ⅰ. 无针乌贼视网膜电图的特性[J]. 海洋学报: 中文版, 1981, 3(3): 472-476. http://www.hyxbocean.cn/article/id/19810310 [24] 柴敏娟, 郑微云. 头足类视觉特性的电生理研究——Ⅱ. 无针乌贼视网膜电图的光谱敏感性[J]. 海洋学报: 中文版, 1982, 4(6): 784-787. http://www.hyxbocean.cn/article/id/19820615 [25] HISAYUKI A, SOKJIN C, TAKEFUMI A, et al. Relationship between underwater irradiance and distribution of Japanese common squid under fishing light of a squid jigging boat[J]. Fish Sci, 1998, 64(4): 553-557. doi: 10.2331/fishsci.64.553
[26] SUKREE H. Effect of light on squid behaviour and its application for squid net fishing in Malaysia[D]. Serdang: Universiti Putra Malaysia, 1996. https://www.semanticscholar.org/paper/Effect-Of-Light-On-Squid-Behaviour-And-Its-For-Net-Hajisamae/a22b8a56d2bdc08ca3525a6145e96e6ad204812e
-
期刊类型引用(7)
1. LI Shuang,HAO Qing,QIU Jiayin,ZHOU Xu,CHI Changfeng,ZHENG Libing. The First Non-Chordates QRFP-Like Peptide Receptor Gene in the Cephalopod Sepiella japonica: Identification, Characterization and Possible Role in Food Intake. Journal of Ocean University of China. 2025(01): 195-208 .  必应学术
必应学术
2. 尉小强,王江,周爱民,王鸿泽,罗晓林,安添午,官久强,王之盛. 饥饿对牦牛生理生化指标的影响. 中国奶牛. 2023(11): 16-22 . 百度学术
3. 戴远棠,谢晓晖,黄国光,梁伟峰,沈南南,赵会宏. 虎斑乌贼幼体对盐度及pH值耐受力的研究. 河北渔业. 2012(10): 8-11+16 . 百度学术
4. 吴常文,周超,郭宝英,张建设. 浙江近海曼氏无针乌贼(Sepiella maindroni)繁殖生物学特性变化研究. 海洋与湖沼. 2012(04): 689-694 . 百度学术
5. 张建设,夏灵敏,迟长凤,吴常文. 人工养殖曼氏无针乌贼(Sepiella maindroni)繁殖生物学特性研究. 海洋与湖沼. 2011(01): 55-59 . 百度学术
6. 张建设,迟长凤,吴常文. 曼氏无针乌贼胚胎发育生物学零度和有效积温的研究. 南方水产科学. 2011(03): 45-49 . 本站查看
7. 范帆,尹飞,王建,范帆,李杰. 曼氏无针乌贼(Sepiella maindroni)生物学研究现状及展望. 现代渔业信息. 2011(06): 6-9+19 . 百度学术
其他类型引用(7)


计量
- 文章访问数: 3124
- HTML全文浏览量: 121
- PDF下载量: 1701
- 被引次数: 14
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
