
Development of immune organs at early stages of Trachinotus ovatus
-
摘要:
对卵形鲳鲹(Trachinotus ovatus)仔、稚鱼期间3种免疫器官的发生和发育过程进行了研究。结果表明,3种免疫器官原基出现的先后顺序是头肾、脾脏和胸腺,出现时间分别为2日龄、3日龄和3日龄。其中,头肾的早期发育过程可分为原基形成期、造血功能发展期和泌尿功能退化期。进入稚鱼期后头肾的肾小管分布相对减少,表明头肾已不是主要的泌尿功能区。6~8日龄仔鱼脾脏中造血干细胞向红血细胞分化并产生大量未成熟红细胞。22日龄时脾脏组织体积进一步增大,内部血细胞参与血液循环。胸腺原基出现后胸腺细胞增多、体积增大,细胞嗜碱性不强。10~13日龄时胸腺细胞的嗜碱性增强,淋巴细胞开始增多,胸腺开始分区。20~22日龄稚鱼期结束时胸腺的内区和外区进一步分化,分区更为明显,表明胸腺进一步成熟。未发现胸腺和头肾之间淋巴细胞迁移的现象。
Abstract:A histological examination was conducted to study the ontogenetic development of 3 immune organs of Trachinotus ovatus at early developmental stages. The anlage of head kidney was the first organ to be observed at 2 day-post-hatch (dph), and then followed by spleen (3 dph) and thymus (3 dph). The developmental process for head kidney includes 3 stages which are anlage forming stage, developmental stage of hematopoiesis and degraded stage of urinary function. When the fish became postlarva (18~22 dph), the distribution of renal tubules became less and its urinary function degraded. At 6~8 dph, the hematopoietic stem cells began to differentiate into large amount of immature red cells. At 22 dph, the spleen became larger and more mature, and the inner blood cells were involved in blood circulation. The number and size of thymus cells with weak basophilia grew after anlage of thymus appeared. At 10~13 dph, the basophilia of thymus cells and the number of lymphocyte grew, and the thymus began to differentiate to inner zone and outer zone. During the end of juvenile stage (20~22 dph), the inner zone and outer zone became distinct, indicating the maturity of thymus. The transfer of lymphocyte between thymus and head kidney had not been observed.
-
Keywords:
- Trachinotus ovatus /
- immune organ /
- development
-
卵形鲳鲹(Trachinotus ovatus)隶属于鲈形目,鲈亚目,鲹科,鲳鲹属,俗称金鲳、黄腊鲳和红三等,分布于印度洋、印度尼西亚、澳洲、日本、美洲的热带及温带的大西洋海岸及中国黄渤海、东海、南海,体呈卵形。卵形鲳鲹的肉白色细嫩、含脂肪多、鲜美甜口,是高级食用海水鱼,人工养殖生长速度快,养殖半年多可达500 g,广东、福建和海南等地均有养殖[1]。目前国内外已有不少卵形鲳鲹的研究报道[1-9],但有关其免疫学方面的报道不多见[10],未见免疫系统发育的研究报道。鱼类在早期发育过程中经历变态过程,此期间鱼类的形态、组织器官等发生巨大变化并逐步成熟。卵形鲳鲹在早期发育过程中,18日龄左右即出现鳞片,22日龄体表布满鳞片[11],完成变态,标志着稚鱼期结束。笔者以0~22日龄的卵形鲳鲹仔、稚鱼为研究对象,运用连续石蜡切片技术,探讨卵形鲳鲹变态过程中(仔鱼和稚鱼期)3种免疫器官的发生和发育过程,为其发育生物学和免疫防病等提供基础资料。
1. 材料与方法
1.1 试验材料
试验材料为南海水产研究所深圳试验基地人工培育的卵形鲳鲹仔、稚鱼。苗种培育池面积约为1 000 m2,平均水深约1.2 m。苗种培育的水温为21.9~26.2 ℃(平均为24.7 ℃),盐度为29~31。苗种培育池中提前接种轮虫和桡足类,在卵形鲳鲹仔鱼开口后,投喂自然繁殖的轮虫和桡足类,补充池塘中卵形鲳鲹仔、稚鱼的饵料生物。
1.2 方法
从孵化出膜开始,每天小心随机捞取健康仔、稚鱼进行固定。固定的仔、稚鱼为0~22日龄。固定液为波恩氏液,样品固定12~24 h后,转至70%酒精中保存。用Leica系列组织切片机进行脱水、包埋,5~6 μm连续切片,展片,H.E.染色,中性树胶封片。用Leica DM LB2型显微镜观察仔、稚鱼的头肾、胸腺和脾脏组织结构,数码相机摄影。
2. 结果与分析
2.1 头肾
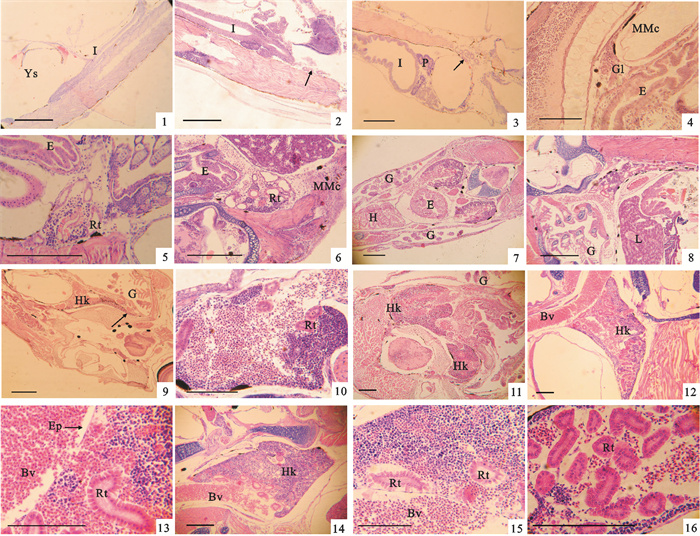
0日龄仔鱼:纵向切片中显示,仔鱼的肠管从肛门处向身体前端逐渐开通,但肠管背部和脊索下侧未发现原肾管(图 1-1)。
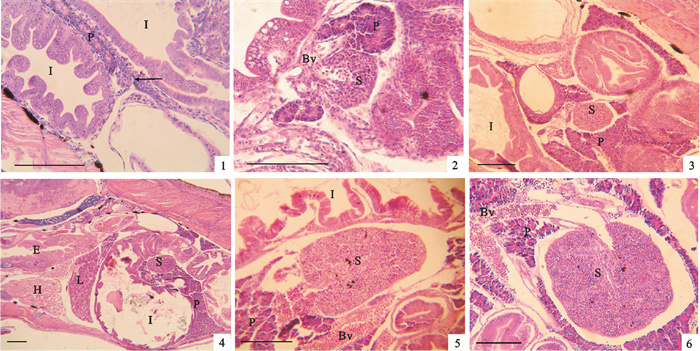
![]() 图 1 卵形鲳鲹头肾的早期发育1. 0日龄仔鱼;2. 2日龄仔鱼;3. 3日龄仔鱼;4~5. 4~5日龄仔鱼;6. 6~7日龄仔鱼;7~8. 10~12日龄仔鱼;9~10. 10~12日龄仔鱼;11. 14~15日龄仔鱼;12~13. 17~18日龄稚鱼;14~16. 20~22日龄稚鱼;E.食道;G.鳃;Gl.肾小球;H.心脏;I.肠;L.肝;MMc.黑色素-巨噬细胞中心;P.胰脏;Rt.肾小管;Ys.卵黄囊;图 1-2~3箭头示头肾原基;Bv.血管;Ep.上皮细胞;Hk.头肾;图 1-9箭头示头肾最前端;标尺=100 μmFig. 1 Development of head kidney at early stages of T.ovatus1. 0 dph larva; 2.2 dph larva; 3. 3 dph larva; 4~5. 4~5 dph larva; 6. 6~7 dph larva; 7~8. 10~12 dph larva; 9~10.10~12 dph larva; 11.14~15 dph larva; 12~13. 17~18 dph juvenile; 14~16. 20~22 dph juvenile; E.esophagus; G.gill; Gl.glomerulus; H.heart; I.intestine; L.liver; MMc.melanomacrophage center; P.pancrease; Rt.renal tubule; Ys.yolk sac; arrows in Fig. 1-2~3 show anlage of head kidney; Bv.vessle; Ep.epithelium cell; Hk.head kidney; arrows in Fig. 1-9 show the top end of head kidney; bar=100 μm
图 1 卵形鲳鲹头肾的早期发育1. 0日龄仔鱼;2. 2日龄仔鱼;3. 3日龄仔鱼;4~5. 4~5日龄仔鱼;6. 6~7日龄仔鱼;7~8. 10~12日龄仔鱼;9~10. 10~12日龄仔鱼;11. 14~15日龄仔鱼;12~13. 17~18日龄稚鱼;14~16. 20~22日龄稚鱼;E.食道;G.鳃;Gl.肾小球;H.心脏;I.肠;L.肝;MMc.黑色素-巨噬细胞中心;P.胰脏;Rt.肾小管;Ys.卵黄囊;图 1-2~3箭头示头肾原基;Bv.血管;Ep.上皮细胞;Hk.头肾;图 1-9箭头示头肾最前端;标尺=100 μmFig. 1 Development of head kidney at early stages of T.ovatus1. 0 dph larva; 2.2 dph larva; 3. 3 dph larva; 4~5. 4~5 dph larva; 6. 6~7 dph larva; 7~8. 10~12 dph larva; 9~10.10~12 dph larva; 11.14~15 dph larva; 12~13. 17~18 dph juvenile; 14~16. 20~22 dph juvenile; E.esophagus; G.gill; Gl.glomerulus; H.heart; I.intestine; L.liver; MMc.melanomacrophage center; P.pancrease; Rt.renal tubule; Ys.yolk sac; arrows in Fig. 1-2~3 show anlage of head kidney; Bv.vessle; Ep.epithelium cell; Hk.head kidney; arrows in Fig. 1-9 show the top end of head kidney; bar=100 μm2~3日龄仔鱼:2 d仔鱼在肠管背侧发现1对细长的肾管,在肾管前端发现初步卷曲的肾小管(图 1-2)。这个肾小管出现的地方即为头肾原基。3 d仔鱼在胸部脊椎下缘可见更加弯曲的肾小管(图 1-3)。
4~5日龄仔鱼:头肾部分可见肾小球(图 1-4),肾小管周围分布有红血细胞等,有的管内壁有被伊红染成红色的分泌细胞(图 1-5)。头肾部分布有一些染成蓝色的肾细胞,还有一些未分化的干细胞,呈圆形、核较大,其细胞质染色较浅,偶尔可观察到分裂现象,可能是造血干细胞。此时,头肾已经开始出现造血功能。黑色素-巨噬细胞中心(melanomacrophage center,MMc)仅分布在头肾的外周(图 1-5)。仔鱼头肾部后面是2个细细的集合管,通向体后部。
6~7日龄仔鱼:头肾部分的肾小管增多,肾细胞和造血干细胞成簇出现(图 1-6)。头肾部有大量的血细胞,以红血细胞为主,在前部汇集到血管中。这些红细胞形状不太规则,呈梭形、圆形和新月形等,是未成熟红细胞。头肾组织未见明显的被膜。
10~12日龄仔鱼:头肾部分的肾小管数量未见明显增加。两侧的头肾中含有许多血细胞,以红细胞为主,其汇聚的血管向下延伸,绕过食道后相连在一起(图 1-7)。头肾部分的肾细胞和造血干细胞主要成簇分布于周围的远血管端(图 1-8~10)。头肾部分最前端可以到达鳃腔前缘背部(图 1-9)。
14~15日龄仔鱼:头肾前部的肾小管有逐步退化的趋势。头肾部除了血管处之外的组织相对更加致密,造血干细胞、肾细胞和血细胞等混合分布(图 1-11)。血管处的血细胞含有红血细胞及其他淋巴细胞等。
17~18日龄稚鱼:头肾前部被染成蓝色,淋巴细胞的分化也越来越多(图 1-12)。头肾部分仍然分布有部分肾小管,但大部分逐步退化,头肾部分已经不再是泌尿的主要器官。头肾前部与血管相接之处可见极薄的被膜,由1层单层扁平上皮细胞构成,细胞核被染成蓝色,形状细长(图 1-13)。该被膜并未完全包被头肾,其若干区域有孔洞,血细胞可以穿过并进入血管。
20~22日龄稚鱼:头肾部分染色为蓝色,其后缘为中肾,其中含有大量的肾小管和肾小球(图 1-14~16)。但肾小管周围分布有较多的血细胞。中肾背部和腹部的边缘分布有肾细胞和一些造血干细胞,仍然具有造血功能,为造血和泌尿功能的混合区。但随着向中肾后部的延伸,造血干细胞数量的分布减少,血细胞的量也减少。整个肾脏的造血功能区和泌尿功能区没有明显的界限。
卵形鲳鲹头肾的早期发育过程可以分为原基形成期、造血功能发展期和泌尿功能退化期3个时期。原基形成期主要包括仔鱼孵化出膜后至头肾原基的形成,包括0~3日龄仔鱼。随后头肾原基中的肾小管逐步发育增多增长,造血干细胞增多并逐步分化为各种血细胞。造血干细胞首先主要分化为红细胞(4~12日龄),随后逐渐增加淋巴细胞的分化(14日龄以后),造血功能逐步发展。头肾的肾小管在发育过程中首先增多,但14日龄以后开始逐步退化,进入泌尿功能退化期。进入稚鱼期以后头肾部分的肾小管分布减少,头肾已经不是主要的泌尿功能区。头肾后部的中肾可见大量密集的肾小管,中肾逐渐成为主要的泌尿功能区。但稚鱼期中,卵形鲳鲹的头肾和中肾都可见肾小管和红血细胞相间分布,说明头肾和中肾都有造血功能和泌尿功能,只是头肾部分造血功能更强,后面的中肾泌尿功能更强。
2.2 脾脏
3日龄仔鱼:在仔鱼腹部的肠管之间发现脾脏原基,由数个实质细胞构成一个近圆形结构,被胰脏组织包围。脾脏原基中有十分细小的毛细血管,红血细胞可见(图 2-1)。
![]() 图 2 卵形鲳鲹脾脏的早期发育1. 3日龄仔鱼;2. 6~8日龄仔鱼;3~4. 10~12日龄仔鱼;5. 17~18日龄稚鱼;6. 20~22日龄稚鱼;Bv.血管;E.食道;H.心脏;I.肠;L.肝脏;P.胰脏;S.脾脏;箭头示脾脏原基;标尺=100 μmFig. 2 Development of spleen at early stages of T.ovatus1. 3 dph larva; 2. 6~8 dph larva; 3~4. 10~12 dph larva; 5. 17~18 dph juvenile; 6. 20~22 dph juvenile; Bv.vessle; E.esophagus; H.heart; I.intestine; L.liver; P.pancrease; S.slpeen; arrows show spleen anlage; bar=100 μm
图 2 卵形鲳鲹脾脏的早期发育1. 3日龄仔鱼;2. 6~8日龄仔鱼;3~4. 10~12日龄仔鱼;5. 17~18日龄稚鱼;6. 20~22日龄稚鱼;Bv.血管;E.食道;H.心脏;I.肠;L.肝脏;P.胰脏;S.脾脏;箭头示脾脏原基;标尺=100 μmFig. 2 Development of spleen at early stages of T.ovatus1. 3 dph larva; 2. 6~8 dph larva; 3~4. 10~12 dph larva; 5. 17~18 dph juvenile; 6. 20~22 dph juvenile; Bv.vessle; E.esophagus; H.heart; I.intestine; L.liver; P.pancrease; S.slpeen; arrows show spleen anlage; bar=100 μm6~8日龄仔鱼:脾脏呈桃形,附在肠外壁上,周围有胰脏组织等包被(图 2-2)。体积明显增大,内部由毛细血管形成的窦状隙增多增大,其中含有丰富的血细胞以及少量微嗜碱性细胞或造血干细胞。这些细胞以红细胞为主,形状不规则,有的细胞核偏位,细胞形状有的呈近圆形,多为未成熟的红细胞。这些红细胞最终汇聚到附近的毛细血管中。
10~12日龄仔鱼:脾脏椭圆形,上面紧贴中肠外壁,下半部分由胰脏组织包被。脾脏内窦状隙扩大,充满红色的血细胞(图 2-3)。同时可见淋巴细胞增多,均匀地分布在脾脏组织中(图 2-3~4)。
17~18日龄稚鱼:脾脏切面呈心形,与腹部血管联系紧密。脾脏内部窦状隙中分布有网状细胞、实质细胞、红血细胞和一些淋巴细胞等。淋巴细胞的细胞核染色为淡蓝色,数量不多,不易区分。脾脏组织内开始出现几个MMc(图 2-5)。
20~22日龄稚鱼:脾脏体积进一步增大,周围由胰脏组织包被,可见有较大的血管与脾脏连通(图 2-6)。脾脏中间部分比较疏松,分布有网状细胞和纤维组织。MMc数量较少,体积较小,主要分散分布于脾脏组织内部。脾脏周围血管中的血细胞仍然以红细胞为主。
卵形鲳鲹在仔、稚鱼发育阶段,脾脏原基从数个实质细胞逐渐发育到窦状隙增多、发达,脾脏内部主要表现为造血干细胞向红血细胞分化并产生大量未成熟红细胞。这在6~8日龄的仔鱼脾脏中就已经出现,因此,脾脏已经开始参与造血功能。22日龄稚鱼体表布满鳞片,稚鱼期结束,此时脾脏组织体积相对进一步增大,内部造血干细胞和未成熟红细胞十分丰富,造血功能逐步成熟。此时的脾脏与附近血管相连,大量血细胞进入血管中,参与血液循环。
2.3 胸腺
3日龄仔鱼:在3日龄仔鱼的鳃腔后上方出现胸腺原基(图 3-1)。胸腺原基是一个对称的器官,由数个细胞组成一团,位于各鳃腔后缘背角附近的上皮组织下方,被染成蓝色。胸腺原基和鳃腔之间由一层较薄的上皮组织间隔。这些细胞团呈短棒状,在鳃腔上皮中形成突起。
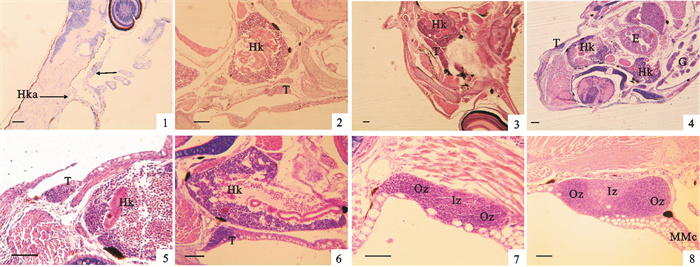
![]() 图 3 卵形鲳鲹胸腺的早期发育1. 3日龄仔鱼;2. 7日龄仔鱼;3~6. 10~13日龄仔鱼;7. 16~18日龄稚鱼;8. 20~22日龄稚鱼;E.食道;G.鳃;Hk.头肾;Hka.头肾原基;Iz.内区;MMc.黑色素-巨噬细胞中心;Oz.外区;T.胸腺;箭头示胸腺原基;标尺=50 μmFig. 3 Development of thymus at early stages of Trachinotus ovatus1. 3 dph larva; 2. 7 dph larva; 3~6. 10~13 dph larva; 7. 16~18 dph juvenile; 8. 20~22 dph juvenile; E.esophagus; G.gill; Hk.hed kidney; Hka.head kidney anlage; Iz.inner zone; MMc.melanomacrophage center; Oz.outerzone; T.thymus; arrows show thymus anlage; bar=50 μm
图 3 卵形鲳鲹胸腺的早期发育1. 3日龄仔鱼;2. 7日龄仔鱼;3~6. 10~13日龄仔鱼;7. 16~18日龄稚鱼;8. 20~22日龄稚鱼;E.食道;G.鳃;Hk.头肾;Hka.头肾原基;Iz.内区;MMc.黑色素-巨噬细胞中心;Oz.外区;T.胸腺;箭头示胸腺原基;标尺=50 μmFig. 3 Development of thymus at early stages of Trachinotus ovatus1. 3 dph larva; 2. 7 dph larva; 3~6. 10~13 dph larva; 7. 16~18 dph juvenile; 8. 20~22 dph juvenile; E.esophagus; G.gill; Hk.hed kidney; Hka.head kidney anlage; Iz.inner zone; MMc.melanomacrophage center; Oz.outerzone; T.thymus; arrows show thymus anlage; bar=50 μm7日龄仔鱼:胸腺组织增大,胸腺实质细胞未见明显分化(图 3-2)。胸腺细胞团埋于鳃腔上皮下方,胸腺细胞嗜碱性不强,染色较淡,可能含有一些干细胞。此时胸腺与头肾相隔较近,但中间有少量肌肉组织和纤维组织膜等间隔。
10~13日龄仔鱼:胸腺组织呈长棒状,胸腺细胞被染成较为均匀的蓝色,多为典型的淋巴细胞。这些淋巴细胞圆形、近圆形,细胞核相对较大,细胞嗜碱性较强,染色较深。胸腺组织中间部分的细胞染色略微变淡,预示着胸腺组织开始分区,但分区不明显(图 3-3)。胸腺组织和头肾组织相隔不远,但中间有网状组织和肌肉组织间隔,未发现有淋巴细胞迁移的现象(图 3-4~6)。
16~18日龄仔稚鱼:胸腺长形,位于鳃腔后缘背角处。胸腺紧贴着肌肉组织,其鳃腔一侧的表面为一层不规则的上皮细胞,上皮细胞中分布有大量个体较大的分泌样细胞(图 3-7)。此时胸腺开始分区。胸腺的中部且靠近肌肉组织的部分染蓝色较淡,淋巴细胞分布较为稀疏,为内区。前后两侧部分染蓝色较深,淋巴细胞较小,分布较为致密,为外区。内区外区之间的分界并不明显。
20~22日龄稚鱼:胸腺的体积进一步增大,在背角一侧出现MMc。内区和外区的分辨更加明显。胸腺中间的内区细胞分布更加稀疏,多为网状细胞和类肌细胞等,细胞核质比减小,细胞核染色变淡;两侧的外区淋巴细胞的核质比较大,细胞核染色较深。胸腺外侧的上皮组织中,分布有较多的分泌样细胞,在胸腺的表层附近也出现少数分泌样细胞,其深陷在胸腺组织的表层(图 3-8)。
卵形鲳鲹的胸腺原基出现在3日龄,但在随后的一周中其发育主要表现为细胞的增多和体积的增大,细胞嗜碱性不强。10~13日龄胸腺细胞的嗜碱性增强,淋巴细胞开始增多,此时胸腺开始分区。20~22日龄稚鱼期结束时胸腺的内区和外区进一步分化,分区更为明显,表明胸腺进一步成熟。对卵形鲳鲹早期发育切片进行连续观察,胸腺和头肾之间一直有网状组织和肌肉组织等分隔,并未发现胸腺和头肾之间有淋巴细胞迁移现象。
3. 讨论
卵形鲳鲹免疫器官在早期发育阶段中出现的顺序是头肾(2日龄)、脾脏和胸腺(均为3日龄),与大黄鱼(Pleudosciaena crocea)相似[12],但早于大黄鱼、斜带石斑鱼(Epinephelus coioides)[13]和军曹(Rachycentron canadum)[14]等。另外,大西洋鳕(Gadus morhua)的头肾和脾脏在孵化之时出现,早于卵形鲳鲹,胸腺则在9 mm仔鱼时出现,迟于卵形鲳鲹[15]。而大西洋庸鲽(Hippoglossus hippoglossus)的脾脏形成较晚,在孵化后49 d才形成[16]。
研究表明,不同鱼类的初始血细胞形成的时期及造血位点不尽相同[17]。根据卵形鲳鲹头肾的早期发育特征,发育过程基本分为原基形成期、造血功能发展期和泌尿功能退化期。造血功能发展期始于4日龄。4~5日龄卵形鲳鲹仔鱼的头肾部分布有一些染成蓝色的肾细胞,还有一些未分化的干细胞,呈圆形、核较大,其细胞质染色较浅,体积比淋巴细胞大,偶尔可观察到分裂现象,可能是造血干细胞。这与斜带石斑鱼中的类似[13]。随后在6~7日龄仔鱼的头肾中发现大量的血细胞,以红血细胞为主,形状不太规则,呈梭形、圆形和新月形等,是未成熟红细胞。而脾脏也是在6~8日龄左右发现窦状隙中有未成熟红细胞,表明此时卵形鲳鲹的头肾和脾脏均已经开始具备造血功能。卵形鲳鲹肾脏和脾脏的淋巴化程度与牙鲆(Paralichthys olivaceus)早期发育相似[18]。
卵形鲳鲹胸腺在10~13日龄左右淋巴细胞开始增多,标志着胸腺淋巴化开始。而头肾中的淋巴细胞逐渐增多是在进入稚鱼期以后(17~18日龄),其淋巴化迟于胸腺。而在仔、稚鱼期,未发现脾脏淋巴化。因此,3种免疫器官淋巴化的顺序依次是胸腺、头肾和脾脏,与斜带石斑鱼相同[13]。另外,有的鱼类在早期发育阶段出现胸腺消失的现象[18]。此研究主要进行了卵形鲳鲹变态期完成前免疫器官的发育观察,其胸腺在随后的发育中是否会消失,还需进一步的观察研究。
PADROS和CRESPO[19]在大菱鲆(Scophthalmus maximus)刚孵化仔鱼头肾中发现造血干细胞,认为在头肾早期发育阶段有同样的干细胞移向未发育完全的胸腺中。大黄鱼[12]和金鲷(Sparus aurata)[20]等硬骨鱼类在早期发育过程中,均在头肾和胸腺之间发现有淋巴细胞“桥”,推测头肾和胸腺之间存在细胞的迁移。TATNER[21]曾用同位素氚原位标记了6月龄的胸腺细胞,发现胸腺细胞迁移到外周淋巴器官中,如脾脏和肾脏中,但其在免疫器官的早期发生和发育中是否存在细胞迁移则未可知。此试验对卵形鲳鲹早期发育切片进行了连续观察,胸腺和头肾之间一直有网状组织和肌肉组织等分隔,并未发现胸腺和头肾之间有淋巴细胞迁移现象,与斜带石斑鱼类似[13]。
鱼类在免疫系统发育成熟之前其免疫防御主要依靠非特异性免疫,包括细胞免疫和体液因子免疫等。卵形鲳鲹在变态完成之前免疫器官逐渐发育成熟,此时头肾部分造血干细胞已经可以分化为红细胞、淋巴细胞和粒细胞等,脾脏也已经具备造血功能。胸腺则最早进行了淋巴化,变态完成时内外区分区明显,胸腺进一步成熟。这表明卵形鲳鲹非特异性免疫系统发育迅速,在早期发育阶段中对机体免受细菌等的感染起到重要的保护作用。
-
![]()
图 1 卵形鲳鲹头肾的早期发育
1. 0日龄仔鱼;2. 2日龄仔鱼;3. 3日龄仔鱼;4~5. 4~5日龄仔鱼;6. 6~7日龄仔鱼;7~8. 10~12日龄仔鱼;9~10. 10~12日龄仔鱼;11. 14~15日龄仔鱼;12~13. 17~18日龄稚鱼;14~16. 20~22日龄稚鱼;E.食道;G.鳃;Gl.肾小球;H.心脏;I.肠;L.肝;MMc.黑色素-巨噬细胞中心;P.胰脏;Rt.肾小管;Ys.卵黄囊;图 1-2~3箭头示头肾原基;Bv.血管;Ep.上皮细胞;Hk.头肾;图 1-9箭头示头肾最前端;标尺=100 μm
Figure 1. Development of head kidney at early stages of T.ovatus
1. 0 dph larva; 2.2 dph larva; 3. 3 dph larva; 4~5. 4~5 dph larva; 6. 6~7 dph larva; 7~8. 10~12 dph larva; 9~10.10~12 dph larva; 11.14~15 dph larva; 12~13. 17~18 dph juvenile; 14~16. 20~22 dph juvenile; E.esophagus; G.gill; Gl.glomerulus; H.heart; I.intestine; L.liver; MMc.melanomacrophage center; P.pancrease; Rt.renal tubule; Ys.yolk sac; arrows in Fig. 1-2~3 show anlage of head kidney; Bv.vessle; Ep.epithelium cell; Hk.head kidney; arrows in Fig. 1-9 show the top end of head kidney; bar=100 μm
![]()
图 2 卵形鲳鲹脾脏的早期发育
1. 3日龄仔鱼;2. 6~8日龄仔鱼;3~4. 10~12日龄仔鱼;5. 17~18日龄稚鱼;6. 20~22日龄稚鱼;Bv.血管;E.食道;H.心脏;I.肠;L.肝脏;P.胰脏;S.脾脏;箭头示脾脏原基;标尺=100 μm
Figure 2. Development of spleen at early stages of T.ovatus
1. 3 dph larva; 2. 6~8 dph larva; 3~4. 10~12 dph larva; 5. 17~18 dph juvenile; 6. 20~22 dph juvenile; Bv.vessle; E.esophagus; H.heart; I.intestine; L.liver; P.pancrease; S.slpeen; arrows show spleen anlage; bar=100 μm
![]()
图 3 卵形鲳鲹胸腺的早期发育
1. 3日龄仔鱼;2. 7日龄仔鱼;3~6. 10~13日龄仔鱼;7. 16~18日龄稚鱼;8. 20~22日龄稚鱼;E.食道;G.鳃;Hk.头肾;Hka.头肾原基;Iz.内区;MMc.黑色素-巨噬细胞中心;Oz.外区;T.胸腺;箭头示胸腺原基;标尺=50 μm
Figure 3. Development of thymus at early stages of Trachinotus ovatus
1. 3 dph larva; 2. 7 dph larva; 3~6. 10~13 dph larva; 7. 16~18 dph juvenile; 8. 20~22 dph juvenile; E.esophagus; G.gill; Hk.hed kidney; Hka.head kidney anlage; Iz.inner zone; MMc.melanomacrophage center; Oz.outerzone; T.thymus; arrows show thymus anlage; bar=50 μm
-
[1] 区又君, 李加儿. 卵形鲳鲹的早期胚胎发育[J]. 中国水产科学, 2005, 12(6): 786-789. doi: 10.3321/j.issn:1005-8737.2005.06.019 OU Youjun, LI Jia′er. Embryonic development in Trachinotus ovatus[J]. J Fish Sci China, 2005, 12(6): 786-789. (in Chinese) doi: 10.3321/j.issn:1005-8737.2005.06.019
[2] 张邦杰, 梁仁杰, 王晓斌, 等. 卵形鲳鲹Trachinotus ovatus (Linnaeus)的引进、咸、海水池养与越冬[J]. 现代渔业信息, 2001, 16(3): 16-20. https://cstj.cqvip.com/Qikan/Article/Detail?id=5033244&from=Qikan_Article_Detail ZHANG Bangjie, LIANG Renjie, WANG Xiaobing, et al. Introduing Trachinotus ovatus (Linnaeus) and rearing and overwintering them in the ponds with brackish water & sea water[J]. Mod Fish Inf, 2001, 16(3): 16-20. (in Chinese) https://cstj.cqvip.com/Qikan/Article/Detail?id=5033244&from=Qikan_Article_Detail
[3] 方永强, 戴燕玉, 洪桂英. 卵形鲳鲹早期卵子发生显微及超微结构的研究[J]. 台湾海峡, 1996, 15(4): 407-414. https://qikan.cqvip.com/Qikan/Article/Detail?id=2415473 FANG Yongqiang, DAI Yanyu, HONG Guiying. Study on micro-and ultrastructure of early oogenesis in Trachinotus ovatus[J]. J Oceanogr Taiwan Strait, 1996, 15(4): 407-414. (in Chinese) https://qikan.cqvip.com/Qikan/Article/Detail?id=2415473
[4] 王刚, 李加儿, 区又君, 等. 卵形鲳鲹幼鱼耗氧率和排氨率的初步研究[J]. 动物学杂志, 2010, 45(3): 116-121. http://dwxzz.ioz.ac.cn/dwxb/article/abstract/20100318?st=article_issue WANG Gang, LI Jia′er, OU Youjun, et al. Oxygen consumption and ammonia excretion of the juvenile Trachinoms ovatus[J]. Chin J Zool, 2010, 45(3): 116-121. (in Chinese) http://dwxzz.ioz.ac.cn/dwxb/article/abstract/20100318?st=article_issue
[5] 陈伟洲, 许鼎盛, 王德强, 等. 卵形鲳鲹人工繁殖及育苗技术研究[J]. 台湾海峡, 2007, 26(3): 435-441. doi: 10.3969/j.issn.1000-8160.2007.03.019 CHEN Weizhou, XU Dingsheng, WANG Deqiang, et al. Study on the spawning and hatching technique for Trachinotus ovatus[J]. J Oceanogr Taiwan Strait, 2007, 26(3): 435-441. (in Chinese) doi: 10.3969/j.issn.1000-8160.2007.03.019
[6] 区又君, 罗奇, 李加儿, 等. 卵形鲳鲹消化酶活性的研究Ⅰ. 成鱼和幼鱼消化酶活性在不同消化器官中的分布及其比较[J]. 南方水产科学, 2011, 7(1): 50-55. doi: 10.3969/j.issn.2095-0780.2011.01.008 OU Youjun, LUO Qi, LI Jia′er, et al. Study on digestive enzymes activities of Trachinotus ovatusⅠ. Comparison of their distribution in different digestive organs of adult and young fish[J]. South China Fish Sci, 2011, 7(1): 50-55. (in Chinese) doi: 10.3969/j.issn.2095-0780.2011.01.008
[7] 许晓娟, 李加儿, 区又君. 盐度对卵形鲳鲹胚胎发育和早期仔鱼的影响[J]. 南方水产, 2006, 5(6): 31-35. doi: 10.3969/j.issn.2095-0780.2006.06.006 XU Xiaojuan, LI Jia′er, OU Youjun. Efects of salinity on embryonic development and early larvae in ovate pompano Trachinotus ovatus[J]. South China Fish Sci, 2006, 5(6): 31-35. (in Chinese) doi: 10.3969/j.issn.2095-0780.2006.06.006
[8] 许海东, 区又君, 郭志勋, 等. 神经坏死病毒对卵形鲳鲹的致病性及外壳蛋白基因序列分析[J]. 上海海洋大学学报, 2009, 19(4): 482-488. https://xueshu.baidu.com/usercenter/paper/show?paperid=a9cf7410a36e7fcef229e7f446488fd1&site=baike XU Haidong, OU Youjun, GUO Zhixun, et al. The pathogenicity of nervous necrosis virus to Trachinotus ovatus and sequence analysis of the coat protein gene[J]. J Shanghai Ocean Univ, 2010, 18(4): 482-488. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=a9cf7410a36e7fcef229e7f446488fd1&site=baike
[9] 王江勇, 郭志勋, 黄剑南, 等. 一起卵形鲳鲹幼鱼死亡原因的调查[J]. 南方水产, 2006, 2(3): 54-56. doi: 10.3969/j.issn.2095-0780.2006.03.009 WANG Jiangyong, GUO Zhixun, HUANG Jiannan, et al. Investigation on mass mortality of young fish Trachinotus ovatus[J]. South China Fish Sci, 2006, 2(3): 54-56. (in Chinese) doi: 10.3969/j.issn.2095-0780.2006.03.009
[10] 周永灿, 张本, 陈雪芬, 等. 嗜麦芽假单胞菌脂多糖的制备及其在卵形鲳鲹中的免疫效应[J]. 水产学报, 2002, 26(2): 143-148. doi: 10.3321/j.issn:1000-0615.2002.02.009 ZHOU Yongcan, ZHANG Ben, CHEN Xuefen, et al. Preparation of Pseudomonas maltophilia lipopolysaccharidae and immunological analysis of the lipopolysaccharide against Trachinotus ovatus[J]. J Fish China, 2002, 26(2): 143-148. (in Chinese) doi: 10.3321/j.issn:1000-0615.2002.02.009
[11] 何永亮, 区又君, 李加儿. 卵形鲳鲹早期发育的研究[J]. 上海海洋大学学报, 2009, 18(4): 428-434. https://xueshu.baidu.com/usercenter/paper/show?paperid=e1629e0f6b1d49e7257336eff4062d9b&site=xueshu_se&hitarticle=1 HE Yongliang, OU Youjun, LI Jia′er. Early development of Trachinotus ovatus[J]. J Shanghai Ocean Univ, 2009, 18(4): 428-434. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=e1629e0f6b1d49e7257336eff4062d9b&site=xueshu_se&hitarticle=1
[12] 徐晓津, 翁朝红, 王军, 等. 大黄鱼早期发育过程中免疫器官的发生[J]. 海洋学报, 2007, 29(3): 105-113. https://xueshu.baidu.com/usercenter/paper/show?paperid=46dd48fff75af0b262fe66740b9f6037&site=xueshu_se XU Xiaojin, WENG Chaohong, WANG Jun, et al. Ontogeny of immune organs during early development stage of Pseudosciaena crocea[J]. Acta Oceanologyica Sinica, 2007, 29(3): 105-113. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=46dd48fff75af0b262fe66740b9f6037&site=xueshu_se
[13] 吴金英, 林浩然. 斜带石斑鱼淋巴器官个体发育的组织学[J]. 动物学报, 2003, 49(6): 819-828. http://www.irtree.cn/3233/articles/8927280/article_detail.aspx WU Jinying, LIN Haoran. Ontogeny of lymphoid organs in the orange-spotted grouper (Epinephelus coioides)[J]. Acta Zoologica Sinica, 2003, 49(6): 819-828. (in Chinese) http://www.irtree.cn/3233/articles/8927280/article_detail.aspx
[14] 苏友禄, 冯娟, 郭志勋, 等. 军曹鱼淋巴器官发育的形态学研究[J]. 海洋水产研究, 2008, 29(4): 7-14. SU Youlu, FENG Juan, GUO Zhixun, et al. Morphological studies on the development of lymphoid organs in cobia Rachycentron canadum[J]. Mar Fish Res, 2008, 29(4): 7-14. (in Chinese)
[15] SCHRØDER M B, VILLENA A J, JØRGENSEN T O. Ontogeny of lymphoid organs and immunoglobulin producing cells in Atlantic cod (Gadus morhua L.)[J]. Dev Comp Immunol, 1998, 22(5-6): 507-517. doi: 10.1016/s0145-305x(98)00030-5
[16] PATEL S, SØRHUS E, FIKSDAL I U, et al. Ontogeny of lymphoid organs and development of IgM-bearing cells in Atlantic halibut (Hippoglossus hippoglossus L.)[J]. Fish Shellfish Immunol, 2009, 26(3): 385-395. doi: 10.1016/j.fsi.2008.11.018
[17] ZAPATA A, DIEZ B, CEJALVO T, et al. Ontogeny of the immune system of fish[J]. Fish Shellfish Immunol, 2006, 20(2): 126-136. doi: 10.1016/j.fsi.2004.09.005
[18] LIU Y, ZHANG S, JIANG G, et al. The development of the lymphoid organs of flounder, Paralichthys olivaceus, from hatching to 13 months[J]. Fish Shellfish Immunol, 2004, 16(5): 621-632. doi: 10.1016/j.fsi.2003.10.001
[19] PADROS F, CRESPO S. Ontogeny of the lymphoid organs in the turbot Scophthalmus maximus: a light and electron microscope study[J]. Aquaculture, 1996, 144(1/2/3): 1-16. doi: 10.1016/S0044-8486(96)01326-9
[20] JOSEFSSON S, MARY F T. Histogenesis of the lymphoid organs in sea bream (Sparus aurata L.)[J]. Fish & Shellfish Immunol, 1993, 3(1): 35-49. doi: 10.1006/fsim.1993.1004
[21] TATNER M F. The migration of labeled thymocytes to the peripheral lymphoid organs in the rainbow trout, Salmo gairdneri Richardson[J]. Dev & Comp Immunol, 1985, 9(1): 85-91. doi: 10.1016/0145-305X(85)90062-X
-
期刊类型引用(25)
1. 孙丽莹,刘晓彤,孙敏敏,王娜,齐洁,贺艳. 卵形鲳鲹幼鱼期肌肉生长发育规律探究. 中国海洋大学学报(自然科学版). 2025(03): 125-132 .  百度学术
百度学术
2. 朱玲,高心明,刘思琪,吕耀平,戴庆敏,胡则辉,梅祖飞. 唇(鱼骨)成鱼脾和肾的解剖学与组织学特征. 上海海洋大学学报. 2024(03): 635-645 . 百度学术
3. 杨蕊,周胜杰,方伟,马振华. 营养强化对卵形鲳鲹仔、稚鱼骨骼发育基因表达的影响. 水产科技情报. 2021(03): 126-131 . 百度学术
4. 周胜杰,胡静,杨蕊,杨其彬,马振华. 三斑海马早期发育观察. 南方农业学报. 2021(08): 2302-2310 . 百度学术
5. 谭晓晨,麻艳群,董升辉,司楠,曲照球. 中草药复方制剂对团头鲂免疫组织形态及血液指标的影响. 饲料研究. 2021(23): 54-58 . 百度学术
6. 蓝军南,李俊伟,区又君,温久福,李加儿,李活,周慧. 四指马■泌尿系统胚后发育组织学研究. 海洋渔业. 2020(01): 35-44 . 百度学术
7. 蓝军南,温久福,李俊伟,区又君,周慧,李加儿,李活. 四指马鲅淋巴器官发育组织学观察. 渔业科学进展. 2020(03): 70-77 . 百度学术
8. 周慧,李加儿,区又君,温久福,王鹏飞,谢木娇,刘奇奇,赵彦花. 四指马鲅视网膜早期发育的组织学研究. 动物学杂志. 2017(03): 458-467 . 百度学术
9. 杨其彬,马振华,程大川,江世贵,李有宁,陈明强. 卵形鲳鲹仔稚鱼异速生长的研究. 水产科学. 2017(03): 259-266 . 百度学术
10. 宋沙沙,马红玲,冯娟,苏友禄,程长洪,郭志勋. 卵形鲳鲹Tf、TNFα和C-Lys的组织分布及对美人鱼发光杆菌感染的响应. 南方水产科学. 2017(02): 77-84 . 本站查看
11. 陈世喜,王鹏飞,区又君,温久福,李加儿,王雯,谢木娇. 急性和慢性低氧胁迫对卵形鲳鲹幼鱼肝组织损伤和抗氧化的影响. 动物学杂志. 2016(06): 1049-1058 . 百度学术
12. 区又君,李加儿,柳琪. 褐毛鲿早期发育阶段行为选择和游泳能力的研究. 生态科学. 2016(04): 1-11 . 百度学术
13. 罗俊标,谢木娇,区又君,温久福,李加儿,王鹏飞. 四指马鲅头肾和脾脏组织学研究. 生物学杂志. 2016(04): 43-47 . 百度学术
14. 区又君,刘江华,李加儿,吴水清,谢木娇. 驼背鲈头肾和脾脏的形态组织学观察. 南方农业学报. 2015(11): 2034-2039 . 百度学术
15. 区又君,李加儿,蔡文超. 卵形鲳鲹肾脏的显微和超微结构观察. 海洋渔业. 2015(05): 434-441 . 百度学术
16. 区又君,吉磊,李加儿,范春燕,王刚. 相同养殖条件下卵形鲳鲹3个选育群体生长特性的比较. 应用海洋学学报. 2015(02): 177-182 . 百度学术
17. 廖经球,杨小立,庞强,杜民,张玉麟,周振锋. 卵形鲳鲹繁殖生物学及人工繁育技术研究进展. 红河学院学报. 2014(05): 21-25 . 百度学术
18. 郑攀龙,马振华,郭华阳,李有宁,张殿昌,江世贵. 卵形鲳鲹尾部骨骼胚后发育研究. 南方水产科学. 2014(05): 45-50 . 本站查看
19. OU You-jun,LI Jia-er,CAI Wen-chao. Effect of Cu toxicity at different salinity on selective group of juvenile pompano Trachinotus ovatus. 热带海洋学报. 2014(04): 26-32 . 百度学术
20. 区又君,范春燕,李加儿,于娜,苏慧. 急性低氧胁迫对卵形鲳鲹选育群体血液生化指标的影响. 海洋学报(中文版). 2014(04): 126-131 . 百度学术
21. 区又君,陈四海,李加儿. 珠江口池养梭鱼肾脏显微和超微结构观察. 生态科学. 2014(04): 686-690 . 百度学术
22. 区又君,范春燕,李加儿,于娜. 盐度对卵形鲳鲹幼鱼渗透压调节和饥饿失重的影响. 生态学报. 2014(24): 7436-7443 . 百度学术
23. 刘汝建,区又君,李加儿,苏慧,曹守花,王永翠. 盐度、温度对卵形鲳鲹选育群体肝抗氧化酶活力的影响. 动物学杂志. 2013(03): 428-436 . 百度学术
24. 区又君,刘汝建,李加儿,曹守花. 不同盐度下人工选育卵形鲳鲹(Trachinotus ovatus)子代鳃线粒体丰富细胞结构变化. 动物学研究. 2013(04): 411-416 . 百度学术
25. 区又君,吉磊,李加儿,范春燕,王刚. 卵形鲳鲹不同月龄选育群体主要形态性状与体质量的相关性分析. 水产学报. 2013(07): 961-969 . 百度学术
其他类型引用(16)
 下载:
下载:


计量
- 文章访问数: 3955
- HTML全文浏览量: 198
- PDF下载量: 2550
- 被引次数: 41
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
