
Study on body-color polymorphism of Chlamys nobilis in Liusha Bay
-
摘要:
对雷州半岛西部流沙湾养殖的华贵栉孔扇贝(Chlamys nobilis)群体进行了体色多态性研究。结果显示,在调查的10个苗种来源不同、总抽样数量为5 954个个体的养殖群体中,壳色可分为6类,分别是橘黄色(62.31%)、枣褐色(14.51%)、紫白色(12.46%)、橘黄间紫色(6.43%)、紫顶枣褐色(3.46%)和黄顶紫黄色(0.82%);6类壳色群体的闭壳肌颜色均存在白、黄2色,且白色比例均显著高于黄色,其中黄色闭壳肌比例最高的是橘黄色(18.79%);华贵栉孔扇贝成熟的生殖腺颜色分为黄、乳白2色,分别代表雌性和雄性,性比与闭壳肌颜色相关。研究表明,华贵栉孔扇贝壳色以橘黄色为主(P < 0.05),闭壳肌颜色以白色为主(P < 0.05);壳色与闭壳肌颜色存在一定的相关性,带“黄”壳色个体黄色闭壳肌比例较高(P < 0.05);黄色闭壳肌群体雌性比例显著高于雄性(P < 0.01)。
Abstract:We studied the body-color polymorphism of cultured Chlamys nobilis populations in Liusha Bay, Leizhou Peninsula. The results show that there are 5 954 individuals from 10 different groups of different seed sources whose shell colors can be classified into 6 categories [orange (62.31%), bordeaux-brown (14.51%), purple-white (12.46%), orange-purple (6.43%), purple-bordeaux-brown (3.46%)and orange-purple-yellow (0.82%)]. The adductors of all categories have 2 colors (white and yellow) with significantly more white than yellow, and the highest ratio for yellow adductor is orange (18.79%). The gonads of mature C.nobilis has 2 colors of yellow (female) and ivory (male). The sex ratio is correlated with the color of adductor. It is revealed that orange is the primary shell color (P < 0.05) while white is the primary adductor color (P < 0.05) for C.nobilis; the colors of shell and adductor have certain relationship; individuals with yellow shell color have higher ratio of yellow adductor (P < 0.05);the ratio of female is significantly higher than male for population with yellow adductor (P < 0.01).
-
Keywords:
- Chlamys nobilis /
- shell color /
- adductor /
- gonad /
- polymorphism
-
四指马鲅 (Eleutheronema tetradactylum) 俗称马友、午鱼等,隶属于鲻形目、马鲅亚目、马鲅科、四指马鲅属,主要分布在印度洋至西太平洋,尤其在波斯湾至巴布亚新几内亚、澳大利亚北部和东亚 (中国、日本和越南等) 沿海分布密集[1]。四指马鲅肉质鲜美,营养价值高,已成为海水鱼类养殖的新品种。区又君等[2]开展四指马鲅规模化人工繁殖技术研究取得成功,并在广东、广西、海南、福建、浙江等沿海地区推广养殖,推动了马鲅科鱼类的养殖生产。
鳃是鱼类重要的呼吸器官,与外界水体环境直接接触,能最先感知水体溶氧的变化[3]。同时还是渗透压、离子调节及氨氮排泄器官,其功能主要与鳃小片上的线粒体丰富细胞有关。研究表明,广盐性鱼类的线粒体丰富细胞适应低渗和高渗环境时分别表现出淡水型线粒体丰富细胞和海水型线粒体丰富细胞[4-8]。鳃的基本结构和功能研究主要集中在波纹唇鱼 (Cheilinus undulatus)[9]、大黄鱼 (Pseudosciaena crocea)[10]、黄颡鱼 (Pelteobagrus fulvidraco)[11]、怒江裂腹鱼 (Schizothorax nukiangensis)[12]、梭鱼 (Liza haematocheila)[13]、斑马鱼 (Danio rerio)[14]等。有关鱼类鳃组织发育分化也有较多报道,区又君等[15]在卵形鲳鲹 (Trachinotus ovatus) 鳃的分化发育研究中,将其分为原基期、鳃丝分化和发育期以及鳃器官生长发育完善期共3个时期,并发现鳃的分化和发育与鱼的生长、形态发育、呼吸、运动、代谢以及身体各部位功能的完善等相一致;刘阳等[16]在花鲈 (Lateolabrax maculatus) 鳃的组织学研究中,将鳃的发育分为鳃原基出现、鳃丝分化、鳃小片分化和鳃器官完善4个时期,并发现鳃发育与其呼吸和渗透调节功能成正相关。目前有关四指马鲅鳃的基本组织结构、鳃早期发育和分期及鳃小片上线粒体丰富细胞的类型研究资料较少。通过观察线粒体丰富细胞的类型结构,以便更好地分析其对水体盐度的适应性变化,对阐明鱼类渗透压调节以及其适盐范围的研究有重要意义。本研究采用组织切片和透射电镜技术,对人工培育四指马鲅鳃的基本结构、鳃的发育和分期以及线粒体丰富细胞类型进行了系统研究,以期更好地了解四指马鲅鳃组织基本结构以及鳃早期发育分期特征与呼吸和渗透调节功能的关系,为养殖技术的改进及养殖水体的选择和调控提供理论参考。
1. 材料与方法
1.1 实验材料
实验鱼采自中国水产科学研究院南海水产研究所珠海试验基地。实验期间参照区又君等[2]的培育方法进行鱼苗培育。仔鱼孵化后第1天开始计算日龄,1—35日龄,每天取样1次,每次10尾,其中1—20日龄样本取全鱼,20日龄之后的样本取头部组织置于4%多聚甲醛溶液中固定备用;另采10月龄的四指马鲅,共10尾,取左侧第二对鳃弓鳃小块,将鳃组织剪成小块分别置于4%多聚甲醛溶液和电镜固定液 (Servicebio G1102) 中固定备用。
1.2 实验方法
1.2.1 鳃组织切片的制作
将固定好的样品进行流水冲洗12 h,经50%~100%梯度乙醇脱水 (50%乙醇2 h、70%乙醇4 h、80%乙醇2 h、85%乙醇2 h、95%乙醇45 min×2、100%乙醇45 min×2),二甲苯透明 (1/2无水乙醇+1/2二甲苯混合液1 h、二甲苯15 min×2),浸蜡 (石蜡2 h×2),石蜡包埋,切片 (切片厚度为5 μm),采用苏木精-伊红 (HE) 进行染色,中性树脂封片,并在ZEISS Axio Scope. A1型光学显微镜下观察并拍照分析。
1.2.2 透射电镜样品制备
将固定好的样品用0.1 mol·L−1磷酸缓冲液 (pH 7.4) 漂洗3次 (每次15 min)。依次放入 (50%→70%→80%→90%→95%→100%→100%) 乙醇→100%丙酮→100%丙酮进行脱水,每次15 min。渗透[V (丙酮)∶V (812包埋剂)=1∶1,2~4 h渗透过夜],包埋 (纯812包埋剂5~8 h),37 ℃烤箱过夜。超薄切片机切片后用铀铅双染色 (2%醋酸铀饱和乙醇溶液,枸橼酸铅,各染色15 min),切片室温干燥过夜最后用透射电子显微镜 (HITACHI HT7700)下观察,采集图像分析。
2. 结果
2.1 四指马鲅鳃基本结构观察
2.1.1 鳃的组织结构
四指马鲅全长 (27.84±0.06) cm,体质量 (198.48±0.18) g,全鳃4对,鳃弓表面粗糙,鳃弓的外缘着生鳃片,鳃耙位于鳃弓的内缘,呈列齿状排列,鳃耙的数量13~15个 (图1-a),鳃丝整体呈梳状排列在鳃弓上 (图1-b),末端为膨大的盲囊,有血管分布,由鳃丝软骨支持 (图1-c)。鳃小片相互平行且与鳃丝纵轴垂直,基部与鳃丝相连 (图1-d)。每侧鳃丝具鳃小片110~120片,平均长度为 (58.83±7.53) μm。鳃小片相互平行,鳃小片上可见扁平上皮细胞、柱细胞、血细胞、线粒体丰富细胞和黏液细胞 (图1-e)。
![]() 图 1 四指马鲅鳃的显微结构a. 鳃的解剖结构;b. 鳃的横切面;c. 鳃丝顶端;d. 鳃丝和鳃小片整体;e. 鳃小片放大;Gr. 鳃弓;Gb. 鳃耙;SF. 鳃小片;BF. 鳃丝;B. 血管;C. 软骨细胞;MC. 线粒体丰富细胞;PVC. 扁平上皮细胞;BC. 血细胞;SC. 柱细胞;CC. 黏液细胞Figure 1. Microstructure of gill tissue in E. tetradactyluma. Anatomical structure of gill; b. Cross section of gill; c. Top part of gill filament; d. Filament and secondary filament; e. Enlarge of secondary filament; Gr. Gill arch; Gb. Gill raker; SF. Secondary filament; BF. Filament; B. Blood vessel; C. Arch cell; MC. Mitochondria-rich cell; PVC. Pavement cell; BC. Blood cell; SC. Pillar cell; CC. Mucous cell
图 1 四指马鲅鳃的显微结构a. 鳃的解剖结构;b. 鳃的横切面;c. 鳃丝顶端;d. 鳃丝和鳃小片整体;e. 鳃小片放大;Gr. 鳃弓;Gb. 鳃耙;SF. 鳃小片;BF. 鳃丝;B. 血管;C. 软骨细胞;MC. 线粒体丰富细胞;PVC. 扁平上皮细胞;BC. 血细胞;SC. 柱细胞;CC. 黏液细胞Figure 1. Microstructure of gill tissue in E. tetradactyluma. Anatomical structure of gill; b. Cross section of gill; c. Top part of gill filament; d. Filament and secondary filament; e. Enlarge of secondary filament; Gr. Gill arch; Gb. Gill raker; SF. Secondary filament; BF. Filament; B. Blood vessel; C. Arch cell; MC. Mitochondria-rich cell; PVC. Pavement cell; BC. Blood cell; SC. Pillar cell; CC. Mucous cell2.1.2 鳃小片的超微结构
四指马鲅鳃小片主要由扁平上皮细胞、柱细胞、血细胞、线粒体丰富细胞、黏液细胞以及未分化细胞组成。
扁平上皮细胞呈长棒状,内有内质网、线粒体和高尔基体等细胞器,主要分布在鳃小片外侧,每片鳃小片上有扁平上皮细胞15~20个。鳃小片中有大量微血管网分布,内部由血细胞组成 (图2-a)。鳃小片上扁平细胞和微血管内皮中间形成基膜,微血管内皮延伸至柱细胞,在柱细胞和基膜之间有较多的胶原纤维束。柱细胞主要分布于鳃小片中部,数量较少,每片鳃小片上有柱细胞10~15个,细胞呈圆柱形 [(3.05±0.59) μm],细胞核较大[(2.05±0.62) μm],细胞质电子密度大 (图2-b)。在鳃小片基部可见到线粒体丰富细胞、黏液细胞以及未分化细胞 (图2-c)。根据线粒体的特征和细胞器的密度,将线粒体丰富细胞分为两类,分别是McI型 (图2-d) 和McII型 (图2-e)。McI型主要排列在鳃小片基部与鳃丝相连处。胞体和核均比较大 [ 胞体 (8.26±1.97) μm,细胞核 (4.36±0.82) μm],细胞核位于基部,呈圆形或卵圆形,细胞器致密,内含大量的线粒体、内质网和囊泡。细胞内的线粒体表现出多种形状特征,包括椭圆形、长条形以及弯曲的条形等,长椭圆形是其主要类型 (图2-d)。McII型主要排列在鳃小片上,胞体呈长椭圆形 [ 长径 (11.04±0.53) μm,短径 (3.96±0.28) μm],细胞核位于中部 [ 长径 (3.26±0.84) μm,短径 (1.27±0.33) μm],胞内细胞器稀少。线粒体主要呈圆形,微管系统不发达。黏液细胞主要分布在鳃小片的基部,和线粒体丰富细胞对生存在,细胞呈圆形 [(3.88±0.60) μm],胞内充满黏液泡,细胞器较少,只有少量内质网和线粒体,并被黏液泡挤到细胞基部。在鳃小片的基部可见少量未分化细胞 [(3.53±0.41) μm],是上皮细胞、线粒体丰富细胞等其他功能细胞的母体,细胞核较大 [(2.70±0.20) μm],呈圆形。
![]() 图 2 四指马鲅鳃小片内部超微结构观察a. 鳃小片顶部结构;b. 柱细胞和窦状隙;c. 鳃小片底部结构;d. I型线粒体丰富细胞 (←. 微管);e. II型线粒体丰富细胞;PVC. 扁平上皮细胞;GS. 微血管;BC. 血细胞;SC. 柱细胞;BM. 基膜;CF. 胶原纤维束;McI. I型线粒体丰富细胞;McII. II型线粒体丰富细胞;CC. 黏液细胞;NDC. 未分化细胞;M. 线粒体;N. 细胞核;V. 囊泡;箭头. 微管Figure 2. Ultrastructure of gill filaments in E. tetradactyluma. Top part of lamellae; b. Pillar cell and sinusoid; c. Base of lamellae; d. Mitochondria-rich cell I (Arrow-Tubular); e. Mitochondria-rich cell II; PVC. Pavement cell; GS. Blood capillary; BC. Blood cell; SC. Pillar cell; BM. Basement membrane; CF. Collagenous fiber bundle; McI. Mitochondria-rich cell I; McII. Mitochondria-rich cell II; CC. Mucous cell; NDC. Non-differentiated cell; M. Mitochondrion; N. Nucleus; V. Vacuoles; arrow. Tubular
图 2 四指马鲅鳃小片内部超微结构观察a. 鳃小片顶部结构;b. 柱细胞和窦状隙;c. 鳃小片底部结构;d. I型线粒体丰富细胞 (←. 微管);e. II型线粒体丰富细胞;PVC. 扁平上皮细胞;GS. 微血管;BC. 血细胞;SC. 柱细胞;BM. 基膜;CF. 胶原纤维束;McI. I型线粒体丰富细胞;McII. II型线粒体丰富细胞;CC. 黏液细胞;NDC. 未分化细胞;M. 线粒体;N. 细胞核;V. 囊泡;箭头. 微管Figure 2. Ultrastructure of gill filaments in E. tetradactyluma. Top part of lamellae; b. Pillar cell and sinusoid; c. Base of lamellae; d. Mitochondria-rich cell I (Arrow-Tubular); e. Mitochondria-rich cell II; PVC. Pavement cell; GS. Blood capillary; BC. Blood cell; SC. Pillar cell; BM. Basement membrane; CF. Collagenous fiber bundle; McI. Mitochondria-rich cell I; McII. Mitochondria-rich cell II; CC. Mucous cell; NDC. Non-differentiated cell; M. Mitochondrion; N. Nucleus; V. Vacuoles; arrow. Tubular2.2 四指马鲅鳃结构发育的组织学观察
2.2.1 鳃原基发生
1日龄仔鱼出现口咽腔原基,其鳃原基呈半椭圆形排列 (图3-a)。3日龄仔鱼出现原始鳃弓,其上皮细胞形成鳃丝原基,开始出现血管系统,血管中有血细胞 (图3-b)。
![]() 图 3 四指马鲅鳃组织的早期发育a. 1日龄仔鱼;b. 3日龄仔鱼;c. 5日龄仔鱼;d. 5日龄仔鱼 (c放大);e. 7日龄仔鱼;f. 9日龄仔鱼;g. 16日龄仔鱼;h. 17日龄仔鱼;i. 18日龄;j. 22日龄;k. 22日龄 (j的部分放大);l. 30日龄;m. 35日龄;B. 脑;OE. 食道;O. 口咽腔;GP. 鳃原基; BF. 鳃丝;BC. 血细胞;PVC. 扁平细胞;GA. 鳃弓;GAC. 鳃弓软骨;SF. 鳃小片;MC. 线粒体丰富细胞;SC. 柱细胞;CC. 黏液细胞;BFC. 鳃丝软骨Figure 3. Early development of gill tissue in E. tetradactyluma. 1 DAH; b. 3 DAH; c. 5 DAH; d. 5 DAH (Enlargement of c); e. 7 DAH; f. 9 DAH; g. 16 DAH; h. 17 DAH; i. 18 DAH; j. 22 DAH; k. 22 DAH (Enlargement of the box in j); l. 30 DAH; m. 35 DAH; B. Brain; OE. Esophagus; O. Oropharynx; GP. Gill primordium; BF. Filament; BC. Blood cell; PVC. Pavement cell; GA. Gill arches; GAC. Gill arch cartilage; SF. Secondary filament; MC. Mitochondria-rich cell; SC. Pillar cell; CC. Mucous cell; BFC. Filament cartilage
图 3 四指马鲅鳃组织的早期发育a. 1日龄仔鱼;b. 3日龄仔鱼;c. 5日龄仔鱼;d. 5日龄仔鱼 (c放大);e. 7日龄仔鱼;f. 9日龄仔鱼;g. 16日龄仔鱼;h. 17日龄仔鱼;i. 18日龄;j. 22日龄;k. 22日龄 (j的部分放大);l. 30日龄;m. 35日龄;B. 脑;OE. 食道;O. 口咽腔;GP. 鳃原基; BF. 鳃丝;BC. 血细胞;PVC. 扁平细胞;GA. 鳃弓;GAC. 鳃弓软骨;SF. 鳃小片;MC. 线粒体丰富细胞;SC. 柱细胞;CC. 黏液细胞;BFC. 鳃丝软骨Figure 3. Early development of gill tissue in E. tetradactyluma. 1 DAH; b. 3 DAH; c. 5 DAH; d. 5 DAH (Enlargement of c); e. 7 DAH; f. 9 DAH; g. 16 DAH; h. 17 DAH; i. 18 DAH; j. 22 DAH; k. 22 DAH (Enlargement of the box in j); l. 30 DAH; m. 35 DAH; B. Brain; OE. Esophagus; O. Oropharynx; GP. Gill primordium; BF. Filament; BC. Blood cell; PVC. Pavement cell; GA. Gill arches; GAC. Gill arch cartilage; SF. Secondary filament; MC. Mitochondria-rich cell; SC. Pillar cell; CC. Mucous cell; BFC. Filament cartilage2.2.2 鳃结构分化、发育
5日龄仔鱼出现鳃弓软骨,并出现鳃小片结构,鳃丝短,鳃丝上有1~2片鳃小片,其基部有线粒体丰富细胞,细胞核着色较深,胞质染色较浅,细胞呈不规则形。柱细胞出现在鳃小片上,细胞质为淡红色,胞核着色较深。扁平上皮细胞分布在鳃小片边缘 (图3-c、图3-d)。7日龄鳃弓软骨呈横向生长,鳃小片数量增多并逐渐变长。在鳃丝基部可观察到与线粒体丰富细胞相邻的黏液细胞,细胞呈长椭圆形,细胞核呈长棒状 (图3-e)。9日龄鳃丝两边的鳃小片数量增多,长度增加 (表1)。黏液细胞内可观察到较多的黏液颗粒,细胞核被挤到细胞基部 (图3-f)。16日龄鳃弓软骨明显,骨质致密。鳃丝软骨进一步发育,每根鳃丝上有5~10片鳃小片,鳃小片长度为 (13.04±3.06) μm (图3-g)。17日龄鳃丝呈梳状排列,末端膨大。鳃小片排列紧密,鳃丝软骨明显致密,基本结构逐渐完整 (图3-h)。
表 1 四指马鲅鳃发育时鱼体全长、鳃小片数量和鳃小片长度变化Table 1. Total length, number and length change of secondary gill lamella during gill development of E. tetradactylum日龄
Days after hatching/d鱼体全长
Total length/mm鳃小片数量
Number of secondary gill lamella/片鳃小片长度
Length of secondary gill lamella/μm1 4.20±0.21 − − 3 4.33±0.03 − − 5 4.54±0.02 1~2 11.99±1.49 7 5.38±0.11 1~4 12.81±1.46 9 6.41±0.06 3~7 13.02±3.29 16 10.15±0.03 5~10 13.04±3.06 17 12.09±0.03 5~12 13.06±3.08 18 13.18±0.02 7~14 15.02±1.84 22 17.23±0.13 16~20 18.16±2.03 30 26.65±0.06 22~26 21.27±1.52 35 28.61±0.07 24~28 28.71±3.24 2.2.3 鳃结构完善
18日龄鳃丝软骨发育完全,形态完整,鳃结构已经基本形成 (图3-i)。22日龄鳃进一步发育完善,鳃丝上鳃小片数量增多,每根鳃丝上有16~20片鳃小片,扁平上皮细胞和血细胞呈规则相间紧密排列在鳃小片上。在鳃小片的基部可见形态规则的线粒体丰富细胞和黏液细胞 (图3-j、图3-k)。30—35日龄鳃小片平行紧密排列,鳃结构和成鱼的基本相同 (图3-l、图3-m)。
3. 讨论
3.1 四指马鲅鳃结构与功能的适应性
四指马鲅鳃的显微结构与大部分硬骨鱼类基本相似,主要由鳃弓、鳃耙、鳃丝和鳃小片组成[17-20]。李霞[21]研究发现鳃丝两侧靠近边缘处各有1条血管,并在鳃小片内分支形成毛细血管网。鳃小片上水血屏障的双层结构在很大程度上参与了水与血之间的交流,与鳃血管中的血红蛋白释放出氧并带走二氧化碳功能联系密切[22-24]。扁平上皮细胞紧密连接,分布在鳃小片外围,起到保护鳃丝不受外界伤害的作用[25-26]。四指马鲅鳃小片上微血管内皮和扁平上皮中间存在基膜,猜测基膜参与形成鳃小片上水血屏障的双层结构。四指马鲅鳃小片上柱细胞与基膜之间存在较多胶原纤维束,使鳃小片具有一定的收缩性,这与纳氏鲟 (Acipenser naccarii) 相同[27]。柱细胞主要维持鳃小片的形状,起一定支撑作用。鳃组织上黏液细胞数量较少,成熟的黏液细胞可以分泌黏液形成一个保护膜,起到保护鳃组织的作用。
3.2 鳃小片线粒体丰富细胞的分类及其生理功能
硬骨鱼类鳃小片线粒体丰富细胞 (泌氯细胞) 具有调节渗透压、进行离子分泌和吸收的功能[28]。根据线粒体丰富细胞在鳃丝上的位置分布、结构的区别以及适应淡海水时的变化,可以把线粒体丰富细胞分为2种不同类型:α型和β型[11, 29]。α型线粒体丰富细胞与本研究中四指马鲅Ⅰ型线粒体丰富细胞形态结构特点相一致,主要分布在鳃丝和鳃小片基部。β型线粒体丰富细胞与本研究中四指马鲅鲅Ⅱ型线粒体丰富细胞形态相对应,主要存在于鳃小片上。区又君等[30]研究遮目鱼 (Chanos chanos) 幼鱼鳃发现,在不同盐度条件下,遮目鱼幼鱼线粒体丰富细胞表现出2种不同的类型。四指马鲅兼备Ⅰ型和Ⅱ型线粒体丰富细胞,表明其对水体盐度的变化在生理结构上有较强的适应能力。
3.3 四指马鲅鳃发育分期及其功能完善
四指马鲅仔稚鱼鳃发育从组织学方面可以分为3个阶段:第1阶段 (0—3日龄)为鳃原基的出现期。鳃原基初步形成,但没有进行分化。此阶段仔鱼运动能力较弱,主要漂流在水层中。仔鱼主要靠皮肤、鳍和卵黄囊丰富的微血管吸收水中的溶氧进行呼吸[15]。未出现线粒体丰富细胞,仔鱼主要通过卵黄囊、皮肤、肾以及肠道等器官进行渗透调节来维持体内外渗透压的平衡[16]。第2阶段 (4—17日龄)为鳃结构的发育、分化期。此期鳃耙、鳃弓、鳃丝、鳃小片逐渐开始形成,鳃具备了基本的结构和形态特点。在第5日龄时鳃小片基部出现线粒体丰富细胞,与短盖巨脂鲤 (Piaractus brachypomum)[31]和条石鲷 (Oplegnathus fasciatus)[32]类似,与花鲈[16]比相对较早。仔鱼开始摄食,从内源性营养过渡到外源性营养。鳃丝上出现血管网,鳃开始行使呼吸功能,但主要还是以皮肤呼吸为主[33]。鳃开始进行渗透压调节,但功能尚未完善。第3阶段 (18—35日龄)为鳃结构的发育完善期。鳃的整体结构进一步发育和完善,此后,鳃组织的发育主要体现在数量形状的变化。此阶段,四指马鲅能进行完善的渗透调节功能,很好地维持体内外渗透压的平衡。鳃内血管系统发育完善,鳃的呼吸能力增强,成为四指马鲅的主要呼吸器官[34]。线粒体丰富细胞随鳃小片数量增加不断增多,其鳃的呼吸和渗透调节能力也逐渐增强,鳃小片及线粒体丰富细胞多少是决定鳃行使渗透和呼吸力强弱的决定性因素。不同鱼类鳃发育时期有所差异,可能与生活环境、生长繁殖季节以及种属差异有一定联系。
-
![]()
图 1 华贵栉孔扇贝的6种不同壳色
a. 橘黄色; b. 橘黄间紫色; c. 黄顶紫黄色; d. 紫白色; e. 紫顶枣褐色; f. 枣褐色
Figure 1. Six kinds of different shell colors of C.nobilis
a~f represent that shell colors are orange, orange-purple, orange-purple-yellow, purple-white, purple-bordeax-brown and bordeaux-brown, respectively.
![]()
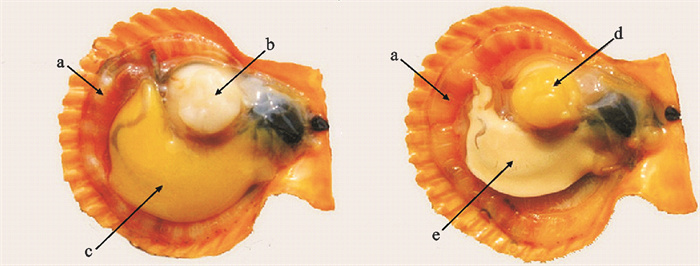
图 2 华贵栉孔扇贝内面观
a. 外套膜;b. 白色闭壳肌;c. 雌性生殖腺;d. 黄色闭壳肌;e. 雄性生殖腺
Figure 2. Internalaspect of C.nobilis
a. mantle; b. white adductor; c. female gonad; d. yellow adductor; e. male gonad
![]()
图 3 华贵栉孔扇贝外套膜颜色
a. 雄性黄肌;b. 雌性黄肌;c. 雄性白肌;d. 雌性白肌
Figure 3. Color of mantle of C.nobilis
a. male with yellow adductor; b. female with yellow adductor; c. male with white adductor; d. female with white adductor
表 1 样品来源
Table 1 Source of samples
养殖群体编号
No. of cultured population采样地点
sampling site苗种来源
source of seed取样数量/个
quantity of sample1 下郁村 英楠村 574 2 下郁村 后洪村 525 3 下郁村 下海村 645 4 下郁村 海尾村 589 5 下郁村 企水村 716 6 下郁村 后港村 685 7 下郁村 流沙村 571 8 海边村 那灵村 509 9 海边村 英岭村 514 10 下郁村 海边村 626  下载: 导出CSV
下载: 导出CSV
表 2 不同群体的华贵栉孔扇贝的壳色组成
Table 2 Composition of shell colors for different C.nobilis populations
个 组别
population橘黄色
(O)枣褐色
(BB)紫白色
(PW)橘黄间紫色
(OP)紫顶枣褐色
(PBB)黄顶紫黄色
(OPY)1 319 112 137 0 0 6 2 400 35 68 6 9 7 3 277 82 184 71 5 26 4 206 134 109 117 16 7 5 517 71 84 41 3 0 6 542 57 9 28 49 0 7 256 209 41 33 31 1 8 376 54 29 18 32 0 9 326 77 56 23 30 2 10 491 33 25 46 31 0 总计 total 3 710 864 742 383 206 49 平均值±标准差 X±SD 371.00a±115.39 86.40b±53.44 74.20b±55.27 38.30bc±34.35 20.60c±16.24 4.90c±7.99 注:同一行数据右上角不同字母表示差异显著(P < 0.05),相同字母表示差异不显著(P>0.05),后表同此
Note: Values with different superscripts in the same line are significantly different (P < 0.05),and those with the same superscripts are not significantly different (P>0.05). The same case in the following tables.
下载: 导出CSV
表 3 不同群体华贵栉孔扇贝的壳色出现频率
Table 3 Frequency of different shell colors of different C.nobilis populations
% 组别
population橘黄色
(O)枣褐色
(BB)紫白色
(PW)橘黄间紫色
(OP)紫顶枣褐色
(PBB)黄顶紫黄色
(OPY)1 55.57 19.51 23.87 0 0 1.05 2 76.19 6.67 12.95 1.14 1.71 1.33 3 42.95 12.71 28.53 11.01 0.78 4.03 4 34.97 22.75 18.51 19.86 2.72 1.19 5 72.21 9.92 11.73 5.73 0.42 0 6 79.12 8.32 1.31 4.09 7.15 0 7 44.83 36.60 7.18 5.78 5.43 0.18 8 73.87 10.61 5.70 3.54 6.29 0 9 63.42 14.98 10.89 4.47 5.84 0.39 10 78.43 5.27 3.99 7.35 4.95 0 平均值±标准差 X±SD 62.15a±16.46 14.73b±9.50 12.46b±8.81 6.30bc±5.68 3.53c±2.70 0.82c±1.25
下载: 导出CSV
表 4 各群体华贵栉孔扇贝各壳色2种颜色闭壳肌数量
Table 4 Quantity of adductor of 2 colors for different shell colors of different C.nobilis populations
个 组别
population橘黄色
O橘黄间紫色
OP枣褐色
BB紫白色
PW黄顶紫黄色
OPY紫顶枣褐色
PBBY W Y W Y W Y W Y W Y W 1 131 188 0 0 33 79 17 120 4 2 0 0 2 18 382 0 6 0 35 1 67 0 7 0 9 3 9 268 1 70 0 82 4 180 0 26 0 5 4 31 175 9 108 3 131 6 103 0 7 0 16 5 42 475 6 35 0 71 1 83 0 0 0 3 6 140 402 5 23 1 56 0 9 0 0 0 49 7 50 206 2 31 0 209 0 41 0 1 0 31 8 340 36 12 6 0 54 1 28 0 0 0 32 9 273 53 15 8 7 70 3 53 1 1 0 30 10 85 406 4 42 0 33 0 25 0 0 1 30 总计 total 1 119 2 591 54 329 44 820 33 709 5 44 1 205 平均值±标准差 X±SD 111.90a±112.81 259.10b±153.11 5.40a±5.17 32.90b±33.91 4.40a±10.30 82.00b±52.57 3.30a±5.21 70.90b±52.17 0.50a±1.27 4.40a±8.07 0.10a±0.32 20.50b±16.17 注:Y. 黄色闭壳肌;W. 白色闭壳肌
Note:Y. yellow adductor;W. white adductor
下载: 导出CSV
表 5 不同群体华贵栉孔扇贝的闭壳肌颜色统计
Table 5 Quantity and frequency of adductor colors for different C.nobilis populations
组别
population黄肌数量/个
quantity of yellow adductor白肌数量/个
quantity of white adductor总数/个
total黄肌频率/%
frequency of yellow adductor白肌频率/%
frequency of white adductor1 185 389 574 32.23 67.77 2 19 506 525 3.62 96.38 3 14 631 645 2.17 97.83 4 49 540 589 8.32 91.68 5 49 667 716 6.84 93.16 6 146 539 685 21.31 78.69 7 52 519 571 9.11 90.89 8 353 156 509 69.35 30.65 9 299 215 514 58.17 41.83 10 90 536 626 14.38 85.62 总计 total 1 256 4 698 5 954 - - 平均值±标准差 X±SD 125.60a±119.24 469.80b±167.5 595.40±71.6 21.10 78.90
下载: 导出CSV
表 6 华贵栉孔扇贝各壳色中黄色闭壳肌出现频率
Table 6 Frequency of yellow adductor color in C.nobilis with different shell colors
% population 橘黄色
O橘黄间紫色
OP黄顶紫黄色
OPY紫白色
PW枣褐色
BB紫顶枣褐色
PBB1 41.07 - 66.67 12.41 29.46 - 2 4.50 0 0 1.47 0 0 3 3.25 1.41 0 2.17 0 0 4 15.05 7.69 0 5.50 2.24 0 5 8.12 14.63 - 1.19 0 0 6 25.83 17.86 - 0 1.75 0 7 19.53 6.06 - 0 0 0 8 90.43 66.67 - 3.45 0 0 9 83.74 65.22 50.00 5.36 9.09 0 10 17.31 8.70 - 0 0 3.23 总计 total 30.16 14.10 10.20 4.45 5.09 0.49 平均值±标准差 X±SD 30.16a±31.64 14.10ab±26.14 10.20abc±32.49 4.45bc±3.85 5.09bc±9.30 0.49c±1.08
下载: 导出CSV
表 7 各群体的不同壳色的华贵栉孔扇贝的性别分布
Table 7 Gender of of C.nobilis of different shell colors for different populations
个 组别
population橘黄色
O枣褐色
BB紫白色
PW橘黄间紫色
OP紫顶枣褐色
PBB黄顶紫黄色
OPY♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂ 1 189 130 74 38 74 63 0 0 0 0 4 2 2 239 161 20 15 40 28 3 3 6 3 5 2 3 161 161 38 44 119 65 32 39 2 3 16 10 4 124 82 66 68 63 46 64 53 10 6 5 2 5 317 200 45 26 35 49 21 20 2 1 0 0 6 243 299 29 28 3 6 17 11 31 18 0 0 7 114 142 65 144 11 30 19 14 13 18 0 1 8 216 160 11 43 6 23 12 6 4 28 0 0 9 196 130 41 36 23 33 18 5 15 15 2 0 10 258 233 16 17 10 15 23 23 12 19 0 0 合计 total 2 060 1 650 405 459 384 358 209 174 95 111 32 17 平均值±标准差 X±SD 206.00a±62.29 165.00a±63.61 40.50a±22.17 45.90a±37.70 38.40a±37.24 35.80a±19.60 20.90a±17.79 17.40a±17.06 9.50a±9.17 11.10a±9.67 3.20a±4.98 1.70a±3.06 ♀: sex ratio 1:0.80 1:1.13 1:0.93 1:0.83 1:1.17 1:0.53
下载: 导出CSV
表 8 不同养殖户群体的华贵栉孔扇贝的性比
Table 8 Sex ratio for different C.nobilis populations
组别
population雄性数量/个
quantity of male雌性数量/个
quantity of female总数/个
total雄性百分比/%
percentage of male雌性百分比/%
percentage of female1 233 341 574 40.59 59.41 2 212 313 525 40.38 59.62 3 274 371 645 42.48 57.52 4 258 331 589 43.80 56.20 5 296 420 716 41.34 58.66 6 362 323 685 52.85 47.15 7 349 222 571 61.12 38.88 8 260 249 509 51.08 48.92 9 219 295 514 42.61 57.39 10 307 319 626 49.04 50.96 总计 total 2 770 3 184 5 954 46.52 53.48 平均值±标准差 X±SD 277.00a±51.48 318.40a±56.24 595.40±71.58 46.50a±6.82 53.50a±6.82 注:其中雌性与雄性个数间进行比较,雌雄百分比与雄性百分比进行比较
Note:Comparisons are between quantity of male and female as well as between percentage of male and female.
下载: 导出CSV
表 9 各群体闭壳肌颜色与性别的相关性
Table 9 Correlation between adductor color and gender for different C.nobilis populations
个 组别
population黄色闭壳肌 yellow adductor 白色闭壳肌 white adductor ♀ ♂ ♀ ♂ 1 121 64 220 169 2 14 5 299 207 3 13 1 358 273 4 37 12 295 245 5 37 12 383 284 6 75 71 248 291 7 29 23 193 326 8 210 143 39 117 9 195 104 100 115 10 61 29 258 278 总计 total 792 464 2 393 2 305 平均值±标准差 X±SD 79.20a±72.52 46.40b±47.80 239.30a±107.43 230.50a±74.89
下载: 导出CSV
-
[1] 张跃环, 闫喜武, 张澎, 等. 贝类壳色多态的研究概况及展望[J]. 水产科学, 2008, 27(12): 680-683. doi: 10.3969/j.issn.1003-1111.2008.12.020 ZHANG Yuehuan, YAN Xiwu, ZHANG Peng, et al. Research advances and prospects on shell color polymorphism of mollusks[J]. Fish Sci, 2008, 27(12): 680-683. (in Chinese) doi: 10.3969/j.issn.1003-1111.2008.12.020
[2] 管云雁, 何毛贤. 海产经济贝类壳色多态性的研究进展[J]. 海洋通报, 2009, 28(1): 108-114. doi: 10.3969/j.issn.1001-6392.2009.01.017 GUAN Yunyan, HE Maoxian. Progress on shell coloration polymorphism in seashells[J]. Mar Sci Bull, 2009, 28(1): 108-114. (in Chinese) doi: 10.3969/j.issn.1001-6392.2009.01.017
[3] SOKOLOVA I M, BERGER V J. Physiological variation related to shell colour polymorphism in White Sea Littorina saxatilis[J]. J Exp Mar Biol Ecol, 2000, 245(1): 1-23. doi: 10.1016/S0022-0981(99)00132-X
[4] WINKLER F M, ESTEVEZ B F, JOLLAN L B, et al. Inheritance of the general shell color in the scallop Argopecten purpuratus (Bivalvia: Pectinidae)[J]. J Hered, 2001, 92(6): 521-525. doi: 10.1093/jhered/92.6.521
[5] 郑怀平, 许飞, 张国范, 等. 海湾扇贝壳色与数量性状之间的关系[J]. 海洋与湖沼, 2008, 39(4): 328-333. doi: 10.3321/j.issn:0029-814X.2008.04.005 ZHENG Huaiping, XU Fei, ZHANG Guofan, et al. Relationships between shell colors and quantitative traits in the bay scallop, Argopecten irradians irradians (Lamarck, 1819)[J]. Oceanologia et Limnologia Sinica, 2008, 39(4): 328-333. (in Chinese) doi: 10.3321/j.issn:0029-814X.2008.04.005
[6] 邬思荣, 刘志刚, 王辉. 湛江北部湾马氏珠母贝红壳色选育系生长模型研究[J]. 广东海洋大学学报, 2011, 31(3): 16-24. doi: 10.3969/j.issn.1673-9159.2011.03.004 WU Sirong, LIU Zhigang, WANG Hui. Study on growth model of the selected lines of red shell colored Pinctada martensii Dunker in Beibu Bay in Zhanjiang[J]. J Guangdong Ocean Univ, 2011, 31(3): 16-24. (in Chinese) doi: 10.3969/j.issn.1673-9159.2011.03.004
[7] 张涛, 郑怀平, 孙泽伟, 等. 华贵栉孔扇贝不同壳色后代早期发育阶段性状比较[J]. 中国农学通报, 2009, 25(23): 478-484. https://xueshu.baidu.com/usercenter/paper/show?paperid=9e2a463a27c124793cd7ae1f678dec96&site=xueshu_se&hitarticle=1 ZHANG Tao, ZHENG Huaiping, SUN Zewei, et al. Comparison of traits among offspring deriving from different shell colors in noble scallop Chlamys nobilis Reeve at early developmental stage[J]. Chin Agric Sci Bull, 2009, 25(23): 478-484. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=9e2a463a27c124793cd7ae1f678dec96&site=xueshu_se&hitarticle=1
[8] 陈炜, 孟宪治, 陶平. 2种壳色皱纹盘鲍营养成分的比较[J]. 中国水产科学, 2004, 11(4): 367-370. doi: 10.3321/j.issn:1005-8737.2004.04.016 CHEN Wei, MENG Xianzhi, TAO Ping. Comparative studies on nutritional composition of abalone Haliotis discus hannai between two shell-color stocks[J]. J Fish Sci of China, 2004, 11(4): 367-370. (in Chinese) doi: 10.3321/j.issn:1005-8737.2004.04.016
[9] 汤娇雯, 张富, 陈兆波. 中国海水养殖种类遗传育种进展与发展趋势[J]. 南方水产, 2009, 5(4): 77-84. doi: 10.3969/j.issn.1673-2227.2009.04.015 TANG Jiaowen, ZHANG Fu, CHEN Zhaobo. Achievement and development trend of genetics and breeding of mariculture varieties in China[J]. South China Fish Sci, 2009, 5(4): 77-84. (in Chinese) doi: 10.3969/j.issn.1673-2227.2009.04.015
[10] 王琦, 袁涛, 何毛贤. 华贵栉孔扇贝养殖及遗传育种研究进展[J]. 南方水产科学, 2011, 7(5): 73-80. doi: 10.3969/j.issn.2095-0780.2011.05.012 WANG Qi, YUAN Tao, HE Maoxian. Research progress of culture and genetic breeding of noble scallop Chlamys nobilis[J]. South China Fish Sci, 2011, 7(5): 73-80. (in Chinese) doi: 10.3969/j.issn.2095-0780.2011.05.012
[11] 张涛. 华贵栉孔扇贝养殖群体数量性状及壳色遗传的初步研究[D]. 汕头: 汕头大学, 2010. 10.7666/d.d171270 ZHANG Tao. The preliminary study on gentics of quantitative traits and shell color in the culture stock of noble scallop Chlamys nobilis Reve[D]. Shantou: Shantou University, 2010. (in Chinese) 10.7666/d.d171270
[12] 刘志刚, 朱晓闻, 王辉, 等. 华贵栉孔扇贝(Chlamys nobilis)壳色遗传规律的研究[J]. 海洋与湖沼, 2012, 43(1): 62-66. https://xueshu.baidu.com/usercenter/paper/show?paperid=8cfa29314c5bfb1abc39e3bdb928afe1 LIU Zhigang, ZHU Xiaowen, WANG Hui, et al. Inheritance of shell color in Chlamys nobilis[J]. Oceanologia et Limnologia Sinica, 2012, 43(1): 62-66. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=8cfa29314c5bfb1abc39e3bdb928afe1
[13] EVANS S, CAMARA M D, LANGDON C J. Heritability of shell pigmentation in the Pacific oyster, Crassostrea gigas[J]. Aquaculture, 2009, 286(3/4): 211-216. doi: 10.1016/j.aquaculture.2008.09.022
[14] INNES D J, HALEY L E. Inheritance of a shell-color polymorphism in the mussel[J]. J Hered, 1977, 68: 203-204. doi: 10.1093/OXFORDJOURNALS.JHERED.A108814
[15] ADAMKEWICZ L, CASTAGNA M. Genetics of shell color and pattern in the bay scallop Argopecten irradians[J]. J Hered, 1988, 79(1): 14-17. doi: 10.1093/oxfordjournals.jhered.a110436
[16] BRAKE J, EVANS F, LANGDON C. Evidence for genetic control of pigmentation of shell and mantle edge in selected families of Pacific oysters, Crassostrea gigas[J]. Aquaculture, 2004, 229(1/2/3/4): 89-98. doi: 10.1016/S0044-8486(03)00325-9
[17] BOETTIGER A, ERMENTROUT B, OSTER G. The neural origins of shell structure and pattern in aquatic mollusks[J]. Proc Natl Acad Sci, 2009, 106(16): 6837-6842. doi: 10.1073/pnas.0810311106
[18] 苏天凤. 华南沿海养殖近江牡蛎的分类研究[J]. 南方水产, 2006, 2(6): 72-75. doi: 10.3969/j.issn.2095-0780.2006.06.014 SU Tianfeng. Taxonomic studies of the cultured species of Crassosstrea sp. in South China waters[J]. South China Fish Sci, 2006, 2(6): 72-75. (in Chinese) doi: 10.3969/j.issn.2095-0780.2006.06.014
[19] MAOKA T. Carotenoids in marine animals[J]. Mar Drugs, 2011, 9(2): 278-293. doi: 10.3390/md9020278
[20] 李宁. 虾夷扇贝橘红色闭壳肌产生的原因及其在育种中的应用[D]. 青岛: 中国海洋大学, 2009. 10.7666/d.y1503915 LI Ning. What accounting for orange color of Yesso scallop muscle (Patinopecten yessoensis) and its application in breeding[D]. Qingdao: Ocean University of China, 2009. (in Chinese) 10.7666/d.y1503915
[21] BELTRAN-LUGO A I, MAEDA-MARTINEZ A N, PACHECO-AGUILAR R, et al. Physical, textural, and microstructural properties of restructured adductor muscles of 2 scallop species using 2 cold-binding systems[J]. J Food Sci, 2005, 70(2): 78-84. https://dialnet.unirioja.es/servlet/articulo?codigo=5747804
[22] KANTHA S S. Carotenoids of edible molluscs: a review[J]. J Food Biochem, 1989, 13(6): 429-442. doi: 10.1111/j.1745-4514.1989.tb00410.x
[23] MATSUNO T. Aquatic animal carotenoids[J]. Fish Sci, 2001, 67(5): 771-783. doi: 10.1046/j.1444-2906.2001.00323.x
[24] 吕文刚. 华贵栉孔扇贝早期生活史温盐效应与选择育种及颜色性状遗传规律研究[D]. 湛江: 广东海洋大学, 2010. 10.7666/d.y1804071 LV Wengang. Study on effects of temperature and salinity during early life histroy, selective breeding and genetic law of color character in Chlamys nobilis (Reeve)[D]. Zhanjiang: Guangdong Ocean University, 2010. (in Chinese) 10.7666/d.y1804071
[25] ZHENG Huaiping, LIU Helu, ZHANG Tao, et al. Total carotenoid differences in scallop tissues of Chlamys nobilis (Bivalve: Pectinidae) with regard to gender and shell colour[J]. Food Chem, 2010, 122(4): 1164-1167. doi: 10.1016/j.foodchem.2010.03.109
[26] 刘合露. 华贵栉孔扇贝橙色闭壳肌个体富集类胡萝卜素原因的初步研究[D]. 汕头: 汕头大学, 2011. https://xueshu.baidu.com/usercenter/paper/show?paperid=dd4ce004ca88e578f6c2e5ec4e06e7b5&site=xueshu_se&hitarticle=1 LIU Helu. Preliminary study on cause of carotenoids enriched in orange adductor individuals in the noble scallops Chlamys nobilis Neeve[D]. Shantou: Shantou University, 2011. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=dd4ce004ca88e578f6c2e5ec4e06e7b5&site=xueshu_se&hitarticle=1
[27] 胡建兴, 黎辉, 郭澄联, 等. 华贵栉孔扇贝雌雄同体及其性转换[M]//金启增. 华贵栉孔扇贝育苗与养殖生物学. 北京: 科学出版社, 1996: 120-126. HU Jianxing, LI Hui, GUO Chenglian, et al. The hermaphrodites and sex-reversal of Chlamys nobilis(Reeve)[M]//JIN Qizeng. The biology and mariculture of scallop Chlamys nobilis (Reeve). Beijing: Science Press, 1996: 120-126. (in Chinese)
-
期刊类型引用(2)
1. 徐思琪,张世勇,张文平,刘洪岩,王明华,钟立强,边文冀,陈校辉. 温度诱导斑点叉尾鮰雌性化研究. 福建农业学报. 2022(06): 741-747 .  百度学术
百度学术
2. 周胜杰,胡静,杨蕊,杨其彬,马振华. 三斑海马早期发育观察. 南方农业学报. 2021(08): 2302-2310 . 百度学术
其他类型引用(1)





计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 3
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
