
Discussion of relationship between Collichthys lucidus and C. niveatus based on 16S rRNA and Cyt b gene sequences
-
摘要:
比较分析了棘头梅童鱼(Collichthys lucidus)和黑鳃梅童鱼(C. niveatus)的16S rRNA和Cyt b基因片段序列差异及遗传分化程度。在长度为526 bp的16S rRNA和379 bp的Cyt b基因片段的核苷酸序列中,2种间共检测到44处核苷酸替代。分析结果表明,2个基因片段的鸟嘌呤(G)含量较低,在Cyt b蛋白质编码基因第三密码子位点上表现尤为明显。基于16S rRNA和Cyt b基因片段分析结果显示,2种间平均遗传距离分别为0.012和0.111。构建的系统树显示2种梅童鱼在这2个基因片段上存在显著的遗传分化。根据Cyt b基因2%/百万年的进化速率推断,棘头梅童鱼与黑鳃梅童鱼的分化时间约为550万年,发生于上新世(Pliocene)早期。
Abstract:Collichthys lucidus and C. niveatus are commercially important fishery resources in China. Partial nucleotide sequences of mitochondrial DNA 16S rRNA and Cyt b gene fragments were sequenced and analyzed to study the relationship between C.lucidus and C. niveatus. Forty-four nucleotide substitutions were detected among 16S rRNA (526 bp) and Cyt b (379 bp) fragments. Nucleotide composition analysis indicated that the content of guanine (G) was relatively low in the two fragments, especially at the third codon position of Cyt b genes. Average genetic distances based on 16S rRNA and Cyt b between C.lucidus and C. niveatus were 0.012 and 0.111, respectively. The phylogenetic tree showed that there was significant genetic divergence between C.lucidus and C. niveatus. According to the evolutionary rate of 2% per one million years, the estimated divergence time between C.lucidus and C. niveatus was 5.5 million years, which indicated an early Pliocene divergence for them.
-
Keywords:
- Collichthys lucidu /
- C. niveatus /
- 16S rRNA /
- Cyt b /
- genetic differentiation
-
棘头梅童鱼(Collichthys lucidus)和黑鳃梅童鱼(C. niveatus)隶属于鲈形目(Perciformes)、石首鱼科(Sciaenidae)、梅童鱼属(Collichthys),均为中国重要的小型底栖鱼类资源[1]。棘头梅童鱼广泛分布于黄、渤海、东海和南海,营底栖生活;黑鳃梅童鱼常见于黄、渤海,主要营浮游生活[1-2]。每年的4~6月和9~10月为梅童鱼的渔汛旺期。近年来,由于过度捕捞和环境变化,梅童鱼资源严重衰退[3]。
国内外学者对梅童鱼的研究主要集中在摄食、生长、鱼卵和仔、稚鱼等渔业生物学方面的报道[4-7]。遗传学研究方面,田兰香等[8]基于Cyt b序列对国内7种石首鱼科鱼类系统发育进行了研究,发现大黄鱼(Pseudosciaena crocea)和棘头梅童鱼亲缘关系较近;蒙子宁等[9]研究了中国近海8种石首鱼科鱼类系统发育关系,结果表明,2种梅童鱼16S rRNA基因片段序列完全相同,但未对其进行深入探讨;陈泉梅[10]对中国石首鱼科鱼类的分子系统学研究结果显示,

1. 材料与方法
1.1 试验材料
选用2种梅童鱼样品各3尾。棘头梅童鱼样品于2005年10月采自海州湾121.0~121.5°E,34.5~35.0°N;黑鳃梅童鱼样品于2004年10月采自海州湾123.5~124.0°E,34.5~35.0°N。酒精固定的梅童鱼肌肉组织保存于中国海洋大学渔业生态研究室样品库,入库编号、GenBank下载的梅童鱼属鱼类16S rRNA序列以及用于外群的白姑鱼(Argyrosomus argentatus) 序列等信息见表 1。
表 1 研究所用样本、分子标记和GenBank注册号Table 1 Species, molecular markers and GenBank accession No.种名species name 缩写编号abbreviation 16S rRNA Cyt b 棘头梅童鱼1 Collichthys lucidus C.lucidus 1 OUC_FELA1 OUC_FELA1 棘头梅童鱼2 C.lucidus 2 OUC_FELA2 OUC_FELA2 棘头梅童鱼3 C.lucidus 3 OUC_FELA3 OUC_FELA3 棘头梅童鱼4 AY336721 AY336721 - 黑鳃梅童鱼1 C. niveatus C. niveatus 1 OUC_FELA6 OUC_FELA6 黑鳃梅童鱼2 C. niveatus 2 OUC_FELA7 OUC_FELA7 黑鳃梅童鱼3 C. niveatus 3 OUC_FELA8 OUC_FELA8 黑鳃梅童鱼4 AY336722 AY336722 - 白姑鱼Argyrosomus argentatus A.argentatus AY336725 AB372025 1.2 试验方法
1.2.1 基因组DNA的提取
取约100 mg的梅童鱼肌肉组织,采用标准的酚-氯仿方法提取基因组DNA[14],将乙醇沉淀后的基因组DNA溶解于100 μL TE溶液中,4 ℃保存备用。
1.2.2 PCR扩增
扩增16S rRNA的引物为:L2510 (5′-CGCCTGTTTAACAAAAACAT-3′) 和H3059 (5′-CCGGTCTGAACTCAGATCATGT-3′)[15];在TaKaRa扩增仪上进行PCR反应:95 ℃变性3 min;94 ℃ 45 s,50 ℃ 45 s,72 ℃ 45 s,40个循环;72 ℃延伸10 min;PCR反应体积为25 μL,其中10×缓冲液2.5 μL,MgCl2 1.5 mmol ·L-1,各种dNTP 200 μmol · L-1,Ex Taq酶1.25 U,每个引物各0.2 mmol · L-1,模板DNA 1 μL,加灭菌蒸馏水至25 μL。扩增Cyt b基因片段的引物为:L14734(5′-AACCACCGTTGTTATTCAACT-3′)和Cyt b (5′-CTCAGAATGACATTTGTCCTCA-3′)[15];PCR反应条件为:95 ℃变性5 min;95 ℃ 1 min,50 ℃ 1 min,72 ℃ 1 min,35个循环;72 ℃延伸10 min。PCR反应体积为25 μL,其中10×缓冲液2.5 μL,MgCl2 1.5 mmol · L-1,各种dNTP 200 μmol · L-1,Ex Taq酶1.25 U,每个引物各0.2 mmol · L-1,模板DNA1 μL,加灭菌蒸馏水至25 μL。以上反应均采用阴性对照来检查是否存在DNA污染。
1.2.3 DNA序列测定
PCR产物电泳检测后,于紫外灯下切割目的条带,用UNIQ-10柱式DNA胶回收试剂盒(上海华舜出品)纯化回收,取适量回收产物做测序反应。用美国ABI公司3700型全自动序列分析仪进行双向测序。
1.2.4 数据分析
所有序列均由Dnastar软件包(DNASTAR, Inc., Madison, USA)进行编辑、校对和排序,并对排序结果进行分析和手工校正,然后用Modeltest 3.7[16]进行模型选择,得到2个目的基因片段的最佳核苷酸替代模型,并计算相关参数。根据核苷酸最适模型,应用MEGA 2.0[17]软件将Cyt b蛋白质编码基因核苷酸序列的密码子转化为氨基酸序列,分析核苷酸组成并计算种间遗传距离。应用ARLEQUIN Ver 2.000[18]软件计算分析棘头梅童鱼和黑鳃梅童鱼的DNA多态性。基于邻接法、最大简约法、最大似然法[19]和贝叶斯法[20]重建16S rRNA和Cyt b基因片段的系统树。
2. 结果
2.1 替代模型选择和参数估计
Modeltest 3.7分析结果显示,2种梅童鱼16S rRNA和Cyt b基因片段最适核苷酸替代模型分别为GTR和HKY+I,并得到5个模型相关参数(表 2),包括模型的似然值自然对数的负数(-ln L)、不变位点比率(I)和Gamma分布的形状参数(G)。
表 2 Modeltest检验得到的优化模型参数Table 2 Optimization model parameters estimated by Modeltest analyses基因gene 16S rRNA Cyt b 模型选择selected model GTR HKY+I -ln L=914.711 9 -ln L=818.320 6 K=8 K=5 AIC=1 845.423 8 AIC=1 646.641 1 (I)=0.761 1 (G)=Equal rates (G)=Equal rates 2.2 核苷酸组成分析
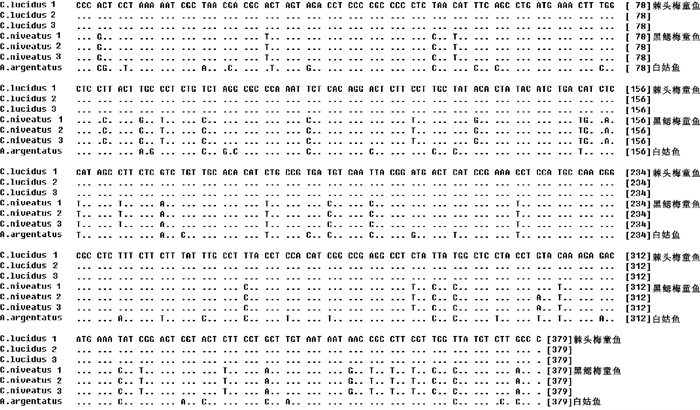
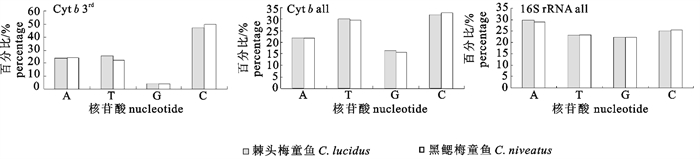
对棘头梅童鱼与黑鳃梅童鱼线粒体DNA 2个基因片段序列进行比对分析,得到长度分别为526 bp(16S rRNA) (图 1)和379 bp(Cyt b) (图 2)的目的片段。2种间在16S rRNA基因片段上共检测到了6个多态位点;在Cyt b基因片段上检测到了38个多态位点(表 3)。核苷酸组成分析结果表明,2种梅童鱼在这2个目的基因片段上的鸟嘌呤(G)含量均比较低,其中最低的为黑鳃梅童鱼的Cyt b基因片段(15.8%);G含量在Cyt b蛋白质编码基因的第三密码子位点上呈现明显的反G偏倚(<5%) (图 3)。棘头梅童鱼与黑鳃梅童鱼Cyt b基因片段中的G+C含量分别为48.1%和48.5%;16S rRNA基因片段G+C含量分别为47.34%和47.72%。
![]() 图 1 棘头梅童鱼与黑鳃梅童鱼16S rRNA基因片段核苷酸序列比对Fig. 1 Nucleotide sequence alignment of partial 16S rRNA of C.lucidus and C. niveatus
图 1 棘头梅童鱼与黑鳃梅童鱼16S rRNA基因片段核苷酸序列比对Fig. 1 Nucleotide sequence alignment of partial 16S rRNA of C.lucidus and C. niveatus![]() 图 2 棘头梅童鱼与黑鳃梅童鱼Cyt b基因片段核苷酸序列比对Fig. 2 Nucleotide sequence alignment of partial Cyt b gene of C.lucidus and C. niveatus
图 2 棘头梅童鱼与黑鳃梅童鱼Cyt b基因片段核苷酸序列比对Fig. 2 Nucleotide sequence alignment of partial Cyt b gene of C.lucidus and C. niveatus![]() 图 3 棘头梅童鱼和黑鳃梅童鱼2个基因片段核苷酸组成all. 整个片段;3rd. 蛋白质编码基因第三密码子位点Fig. 3 Nucleotide composition of two partial fragments of C.lucidus and C.nivertusall. the whole fragment; 3rd. the third codon of the gene
图 3 棘头梅童鱼和黑鳃梅童鱼2个基因片段核苷酸组成all. 整个片段;3rd. 蛋白质编码基因第三密码子位点Fig. 3 Nucleotide composition of two partial fragments of C.lucidus and C.nivertusall. the whole fragment; 3rd. the third codon of the gene2.3 遗传分化分析
棘头梅童鱼与黑鳃梅童鱼在16S rRNA和Cyt b基因片段的核苷酸序列以及Cyt b基因片段相对应的氨基酸序列上存在着不同程度的遗传分化。在526 bp的16SrRNA基因片段上,2种间检测到6个多态位点(包括6个信息简约位点),6个核苷酸替代(nucleotide substitution)中出现1个转换(transition)、5个颠换(transversion),转换颠换比(Ts/Tv)为0.2,基于Tamura-Nei模型得到2种间的遗传距离为0.012,棘头梅童鱼与黑鳃梅童鱼的种内遗传距离皆为0,种内个体间无差异,2种梅童鱼与白姑鱼的遗传距离分别为0.085和0.078;在379 bp的Cyt b基因片段上,检测到38个多态位点(包括38个信息简约位点),38个核苷酸替代包含了35个转换和3个颠换,转换颠换比(Ts/Tv)为11.67,2种梅童鱼的种内遗传距离分别为0和0.002,种间遗传距离为0.111,而与白姑鱼的遗传距离分别为0.153和0.155。
表 3 棘头梅童鱼与黑鳃梅童鱼16S rRNA和Cyt b基因片段遗传多样性Table 3 Genetic diversity information of 16S rRNA and Cyt b gene in C.lucidus and C.niveatus基因gene 片段长/bp fragment length 插入\缺失位点gapped site 多态位点polymorphic site 信息简约位点parsimony informative site 平均碱基差异average pairwise difference 16S rRNA 526 0 6 6 3.2 Cyt b 379 0 38 38 22.7 在蛋白质编码基因片段中,大部分的突变是同义突变(synonymous substitution),最普遍的核苷酸替代是发生在密码子第三位点上的转换,其次是密码子第三位点上的颠换和密码子第一位点上的无义转换[21]。Cyt b基因片段上的38处突变中,35处核苷酸替代发生在密码子第三位点上(2处为颠换),3处位于密码子第一位点上(1处为颠换)。氨基酸水平上,在Cyt b基因片段编码的126个氨基酸序列上,2种间共检测到2处氨基酸替代(amino acid substitution),是由第一密码子非同义替代造成的(转换、颠换各1处) (图 4)。
![]() 图 4 棘头梅童鱼与黑鳃梅童鱼Cyt b氨基酸序列比对Fig. 4 Amino acid sequence alignment of partial Cyt b of C.lucidus and C. niveatus
图 4 棘头梅童鱼与黑鳃梅童鱼Cyt b氨基酸序列比对Fig. 4 Amino acid sequence alignment of partial Cyt b of C.lucidus and C. niveatus根据分子钟假说,某一特定的大分子(蛋白质或DNA分子)在所有的世系(Lineage)中,核苷酸替代速率在时间上是稳定的。BROWN等[13]将2%/百万年的核苷酸分歧速率应用于Cyt b基因片段来推算硬骨鱼的分化时间(divergence time),此核苷酸分歧速率被广泛应用于鱼类分化年代的计算。该研究将此分歧速率应用于棘头梅童鱼与黑鳃梅童鱼Cyt b基因片段,基于Tamura-Nei模型计算的净遗传距离为0.110,推测棘头梅童鱼与黑鳃梅童鱼的分歧时间约为550万年,推算2种间的分化事件发生于上新世(Pliocene)早期。
2.4 系统发育分析
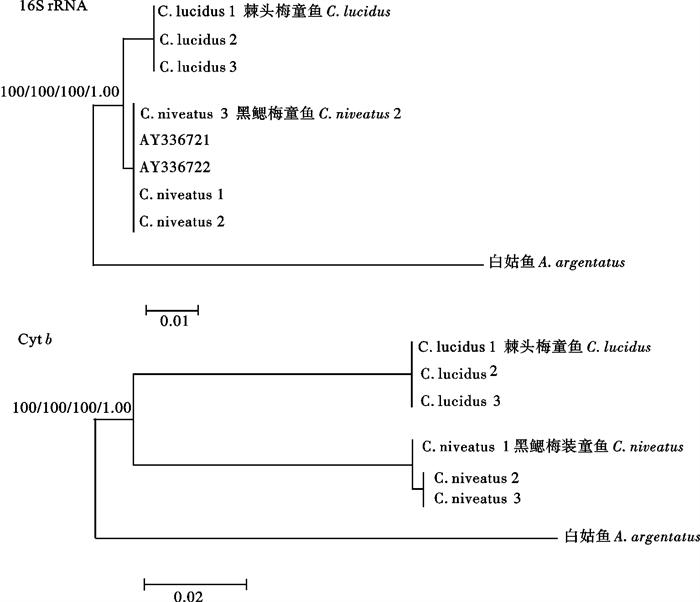
以石首鱼科的白姑鱼作为外群,基于邻接法、最大简约法、最大似然法和贝叶斯法构建的16S rRNA系统发育树基本一致(图 5)。该研究得到的黑鳃梅童鱼序列同GenBank中的棘头梅童鱼序列(AY336721)和黑鳃梅童鱼序列(AY336722)完全相同,三者聚为一支,形成一个单系类群。基于邻接法、最大简约法、最大似然法和贝叶斯法构建的Cyt b系统发育树也基本一致(图 5)。棘头梅童鱼与黑鳃梅童鱼明显分为2支,节点支持率均为100%。
![]() 图 5 基于邻接法、最大简约法、最大似然估计法和贝叶斯法构建的棘头梅童鱼与黑鳃梅童鱼系统发育树(支持率及后验概率标注在分支上)Fig. 5 Phylogenetic trees of C.lucidus and C. niveatus based on neighbor-joining analyses, maximum parsimony analyses and Bayes analyses (Numbers on the trees represent the supporting value and posterior probability)
图 5 基于邻接法、最大简约法、最大似然估计法和贝叶斯法构建的棘头梅童鱼与黑鳃梅童鱼系统发育树(支持率及后验概率标注在分支上)Fig. 5 Phylogenetic trees of C.lucidus and C. niveatus based on neighbor-joining analyses, maximum parsimony analyses and Bayes analyses (Numbers on the trees represent the supporting value and posterior probability)3. 讨论
3.1 2种梅童鱼的分类地位
蒙子宁等[9]对中国近海8种石首鱼科鱼类16S rRNA基因片段序列的研究结果显示,2种梅童鱼的16S rRNA序列相同。而该研究得到的黑鳃梅童鱼序列与棘头梅童鱼序列(AY336721)和黑鳃梅童鱼序列(AY336722)完全相同,共享一个单倍型。基于邻接法、最大简约法、最大似然法和贝叶斯法构建的16S rRNA系统发育树显示,该研究得到的黑鳃梅童鱼序列与蒙子宁等得到的序列(AY336721和AY336722)聚为一支,形成一个单系群,且棘头梅童鱼与黑鳃梅童鱼明显分为2个单系群,节点支持率均为100%。根据16S rRNA基因高度保守性的特点,推测蒙子宁等研究中所用的“棘头梅童鱼”应为黑鳃梅童鱼。基于Cyt b基因片段序列构建的系统树与16S rRNA系统树的结果一致,也支持棘头梅童鱼与黑鳃梅童鱼为2种不同的梅童鱼。
16S rRNA比Cyt b基因进化速率要慢,2个基因片段的序列分析结果都显示棘头梅童鱼与黑鳃梅童鱼存在显著的遗传分化,2种间遗传距离分别为0.012和0.111,属于属内种间差异[22],与笔者对棘头梅童鱼与黑鳃梅童鱼在鳃腔、头部小棘数目、臀鳍第一鳍棘等部位存在显著形态学差异的研究结果(未发表)相一致。
3.2 2种梅童鱼序列分析
4种核苷酸在线粒体基因组中分布不均是动物线粒体基因组的共性[13]。该研究得到的16S rRNA和Cyt b基因片段核苷酸组成中的鸟嘌呤(G)含量普遍较低。棘头梅童鱼和黑鳃梅童鱼的Cyt b蛋白质编码基因第三密码子位点的核苷酸组成存在较大偏倚(Bias),C占46.8%和50%,而G仅占4.2%和4%。在其他鱼类中,Cyt b基因片段核苷酸组成分析的结果也显示G含量普遍较低,这一点在密码子第三位点上尤为明显[23-25]。对于编码重要功能蛋白质的Cyt b基因来说,由于在DNA水平上受自然选择压力的影响,密码子第三位点的突变率高于第一和第二位点,而且密码子第三位点的突变绝大多数是同义突变,受自然选择压力较小,突变后易固定,所以密码子第三位点能够更清晰地表明线粒体基因组核苷酸组成的不均一性[26],而第一、第二位点则相反。
密码子第三位点上的突变很少导致氨基酸替代(同义替代),所以比导致氨基酸替代的突变(非同义替代)积累要快得多,最常见的突变是密码子第三位点上的转换,其次是第一密码子位点上的无义转换和密码子第三位点上的颠换[26]。该研究中Cyt b作为编码重要蛋白质的基因片段,在编码的126个氨基酸序列上,由于第一密码子的非同义替代造成2处氨基酸替代,与先前的研究结果一致[26]。
3.3 2种梅童鱼遗传分化
用一个时间尺度来估计分子进化率和有机体形态变化率以解释生物大进化和生物地理模式是非常必要的[27]。HEWITT[28]认为物种成种事件(speciation event)主要发生在上新世(Pliocene)和更新世(Pleistocene)。目前,很多研究者将2%/百万年的核苷酸分歧速率应用于Cyt b基因片段来推算硬骨鱼类的分化时间[28-30]。根据2种梅童鱼的Cyt b基因片段的净遗传距离为0.110,推算棘头梅童鱼与黑鳃梅童鱼的分歧时间约为550万年,分化事件发生在上新世(Pliocene)早期,与HEWITT的研究结果相符。
-
![]()
图 1 棘头梅童鱼与黑鳃梅童鱼16S rRNA基因片段核苷酸序列比对
Figure 1. Nucleotide sequence alignment of partial 16S rRNA of C.lucidus and C. niveatus
![]()
图 2 棘头梅童鱼与黑鳃梅童鱼Cyt b基因片段核苷酸序列比对
Figure 2. Nucleotide sequence alignment of partial Cyt b gene of C.lucidus and C. niveatus
![]()
图 3 棘头梅童鱼和黑鳃梅童鱼2个基因片段核苷酸组成
all. 整个片段;3rd. 蛋白质编码基因第三密码子位点
Figure 3. Nucleotide composition of two partial fragments of C.lucidus and C.nivertus
all. the whole fragment; 3rd. the third codon of the gene
![]()
图 4 棘头梅童鱼与黑鳃梅童鱼Cyt b氨基酸序列比对
Figure 4. Amino acid sequence alignment of partial Cyt b of C.lucidus and C. niveatus
![]()
图 5 基于邻接法、最大简约法、最大似然估计法和贝叶斯法构建的棘头梅童鱼与黑鳃梅童鱼系统发育树(支持率及后验概率标注在分支上)
Figure 5. Phylogenetic trees of C.lucidus and C. niveatus based on neighbor-joining analyses, maximum parsimony analyses and Bayes analyses (Numbers on the trees represent the supporting value and posterior probability)
表 1 研究所用样本、分子标记和GenBank注册号
Table 1 Species, molecular markers and GenBank accession No.
种名species name 缩写编号abbreviation 16S rRNA Cyt b 棘头梅童鱼1 Collichthys lucidus C.lucidus 1 OUC_FELA1 OUC_FELA1 棘头梅童鱼2 C.lucidus 2 OUC_FELA2 OUC_FELA2 棘头梅童鱼3 C.lucidus 3 OUC_FELA3 OUC_FELA3 棘头梅童鱼4 AY336721 AY336721 - 黑鳃梅童鱼1 C. niveatus C. niveatus 1 OUC_FELA6 OUC_FELA6 黑鳃梅童鱼2 C. niveatus 2 OUC_FELA7 OUC_FELA7 黑鳃梅童鱼3 C. niveatus 3 OUC_FELA8 OUC_FELA8 黑鳃梅童鱼4 AY336722 AY336722 - 白姑鱼Argyrosomus argentatus A.argentatus AY336725 AB372025  下载: 导出CSV
下载: 导出CSV
表 2 Modeltest检验得到的优化模型参数
Table 2 Optimization model parameters estimated by Modeltest analyses
基因gene 16S rRNA Cyt b 模型选择selected model GTR HKY+I -ln L=914.711 9 -ln L=818.320 6 K=8 K=5 AIC=1 845.423 8 AIC=1 646.641 1 (I)=0.761 1 (G)=Equal rates (G)=Equal rates
下载: 导出CSV
表 3 棘头梅童鱼与黑鳃梅童鱼16S rRNA和Cyt b基因片段遗传多样性
Table 3 Genetic diversity information of 16S rRNA and Cyt b gene in C.lucidus and C.niveatus
基因gene 片段长/bp fragment length 插入\缺失位点gapped site 多态位点polymorphic site 信息简约位点parsimony informative site 平均碱基差异average pairwise difference 16S rRNA 526 0 6 6 3.2 Cyt b 379 0 38 38 22.7
下载: 导出CSV
-
[1] 朱元鼎, 罗云林, 伍汉霖. 中国石首鱼类分类系统的研究和新属新种的叙述[M]. 上海: 上海科学技术出版社, 1963. [2] 杨纪明. 渤海鱼类的食性和营养级研究[J]. 现代渔业信息, 2001, 16(10): 10-19. doi: 10.3969/j.issn.1004-8340.2001.10.002 [3] 程济生. 东海、黄海冬季底层鱼类群落结构及其多样性[J]. 海洋水产研究, 2000, 21(3): 1-8. doi: 10.3969/j.issn.1000-7075.2000.03.001 [4] 单乐州, 邵鑫斌, 闫茂仓. 棘头梅童鱼幼鱼生物学特性的初步观察[J]. 水产养殖, 2007, 28(6): 4-5. doi: 10.3969/j.issn.1004-2091.2007.06.002 [5] 单乐州, 邵鑫斌, 谢起浪, 等. 棘头梅童鱼人工繁殖及育苗技术研究[J]. 浙江海洋学院学报: 自然科学版, 2006, 25(3): 266-271. https://xueshu.baidu.com/usercenter/paper/show?paperid=a9573d454dc8a3d427bf988d0b22a5a9&site=xueshu_se&hitarticle=1 [6] 吴振兴, 吴常文. 浙江近海棘头梅童鱼生长规律与群体组成的研究[J]. 水产科技情报, 1990, 17(6): 170-174. https://xueshu.baidu.com/usercenter/paper/show?paperid=50069dd52378130192beab1a98db1c39&site=xueshu_se&hitarticle=1 [7] 谢起浪, 单乐州, 王铁杆, 等. 浙南飞云江口棘头梅童鱼的资源调查和研究[J]. 渔业现代化, 2006(1): 35-36. doi: 10.3969/j.issn.1007-9580.2006.01.015 [8] 田兰香, 梁冰, 张树义, 等. 细胞色素b基因序列与7种石首鱼类的系统进化[J]. 台湾海峡, 2004, 23(4): 436-443. doi: 10.3969/j.issn.1000-8160.2004.04.006 [9] 蒙子宁, 庄志猛, 丁少雄, 等. 中国近海8种石首鱼类的线粒体16S rRNA基因序列变异及其分子系统进化[J]. 自然科学进展, 2004, 14(5): 514-521. doi: 10.3321/j.issn:1002-008X.2004.05.006 [10] 陈泉梅. 中国石首鱼科鱼类分子系统学研究[D]. 广州: 暨南大学, 2007. https://xueshu.baidu.com/usercenter/paper/show?paperid=a8bd7dfa85e3f0700b675e062953355e&site=xueshu_se&hitarticle=1 [11] 肖武汉, 张亚平. 鱼类线粒体DNA的遗传与分化[J]. 水生生物学报, 2000, 24(4): 384-391. doi: 10.3321/j.issn:1000-3207.2000.04.014 [12] AVISE J C. Phylogeography[M]. London: Harvard University Press, 2000: 1-36. doi: 10.2307/j.ctv1nzfgj7
[13] BROWN W M. Evolution of genes and proteins[M]. Sunderland: Sinaue, 1983: 62-88.
[14] SAMBROOK J, FRITSCH E F, MANIATIS T, et al. Molecular cloning: a laboratory manual[M]. New York: Cold Spring Harbor Laboratory Press, 1996: 464-468.
[15] INOUE J G, MIYA M, TSUKAMOTO K. A mitogeneomic perspective on the basal teleostean phylogeny: resolving higher-level relationships with longer DNA sequences[J]. Mol Phyl Evol, 2001, 20(2): 275-285. doi: 10.1006/mpev.2001.0970
[16] POSADA D, CRANDALL K A. Model test: testing the model of DNA substitution[J]. Bioinformatics, 1998, 14(9): 817-818. doi: 10.1093/bioinformatics/14.9.817
[17] KUMAR S, TAMURA K, JAKOBSEN I B, et al. MEGA 2.0: Molecular evolutionary genetics analysis software[J]. Bioinformatics, 2001, 17(12): 1244-1245. doi: 10.1093/bioinformatics/17.12.1244
[18] SCHNEIDER S, ROESSLI D, EXCOFFIER L. ARLEQUIN, Version 2, 000: a software of population genetic data analysis[M]. Geneva: University of Geneva, 2000. https://www.researchgate.net/publication/245491063_ARLEQUIN_A_software_for_population_genetics_data_analysis
[19] SWOFFORD D L. PAUP*. Phylogenetic analysis using parsimony (* and other methods), Version 4[Z]. Sunderland, MA: Sinauer Associates, 2002. 10.1111/j.0014-3820.2002.tb00191.x
[20] RONQUIST F, HUELSENBECK J P. Mrbayes 3: Bayesian phylogenetic inference under mixed models[J]. Bioinformatics, 2003, 19(12): 1572-1574. doi: 10.1093/bioinformatics/btg180
[21] SBISà E, TANZARIELLO F, REYES A, et al. Mammalian mitochondrial D-loop region structure analysis: identification of new conserved sequences and their functional and evolutionary implications[J]. Gene, 1997, 205(1/2): 125-140. doi: 10.1016/s0378-1119(97)00404-6
[22] JOHNS G C, AVISE J C A. Comparative summary of genetic distances in the vertebrates from the mitochondrial Cyt b gene[J]. Mol Phyl Evol, 1998, 15(11): 1481-1490. doi: 10.1093/oxfordjournals.molbev.a025875
[23] 高天翔, 陈省平, 韩志强, 等. 大银鱼和小齿日本银鱼线粒体细胞色素b和16S rRNA部分序列分析[J]. 中国海洋大学学报, 2004, 34(5): 791-794. doi: 10.3969/j.issn.1672-5174.2004.05.008 [24] 高天翔, 陈四清, 刘进贤, 等. 圆斑星鲽与条斑星鲽线粒体DNA部分片段的序列比较研究[J]. 高技术通讯, 2004(14): 329-334. [25] 张俊丽, 高天翔, 韩志强, 等. 3种白鲑线粒体细胞色素b和16S rRNA基因片段序列分析[J]. 中国水产科学, 2007, 14(1): 8-14. doi: 10.3321/j.issn:1005-8737.2007.01.002 [26] MEYER A. Evolution of mitochondrial DNA in fishes[M]//HOCHACHKA P W, MOMMSEN P. Biochemistry and molecular biology of fishes. Amsterdam: Elsevier Press, 1993: 1-36.
[27] 彭作刚, 何舜平, 张耀. 细胞色素b基因序列变异与东亚鳄科鱼类系统发育[J]. 自然科学进展, 2002, 12(6): 596-600. https://xueshu.baidu.com/usercenter/paper/show?paperid=255a47ea18f667ffb1fdbcdbff9bbf09&site=xueshu_se [28] HEWITT G M. Some genetic consequences of ice ages, and their role in divergence and speciation[J]. Biol J Linn Soc, 1996, 58(3): 247-276. doi: 10.1098/rstb.2003.1388
[29] 毕潇潇, 高天翔, 肖永双, 等. 4种鳕鱼线粒体16S rRNA、COI和Cyt b基因片段序列的比较研究[J]. 南方水产, 2009, 5(3): 46-52. doi: 10.3969/j.issn.1673-2227.2009.03.008 [30] 张岩, 肖永双, 高天翔, 等. 两种黄盖鲽线粒体DNA部分片段比较分析[J]. 水产学报, 2009, 33(2): 201-207. https://xueshu.baidu.com/usercenter/paper/show?paperid=6f682f8bb71aff8c26e3ef0b9657a233&site=xueshu_se&hitarticle=1 -
期刊类型引用(8)
1. 刘沅,张玉波,杨琳,陈祥盛. 基于16S rDNA与Cytb序列的疏广蜡蝉属三个近似种的分子鉴定(半翅目:蜡蝉总科:广翅蜡蝉科). 环境昆虫学报. 2016(03): 557-564 .  百度学术
百度学术
2. YUAN JunXia,SHENG GuiLian,HOU XinDong,SHUANG XiaoYan,YI Jian,YANG Hong,LAI XuLong. Ancient DNA sequences from Coelodonta antiquitatis in China reveal its divergence and phylogeny. Science China(Earth Sciences). 2014(03): 388-396 . 必应学术
3. 高天翔,韩刚,马国强,宋娜. 黑鳃梅童鱼和棘头梅童鱼的形态学比较研究. 中国海洋大学学报(自然科学版). 2013(01): 27-33 . 百度学术
4. 宋国华,高继萍,王裕,王春芳,刘田福. 中国地鼠线粒体DNA 16S rRNA基因序列分析及分子系统发育研究. 中国实验动物学报. 2013(02): 52-55 . 百度学术
5. 陈林,周文玉,潘桂平. 盐度对棘头梅童鱼胚胎发育和仔鱼存活的影响. 海洋渔业. 2012(04): 388-392 . 百度学术
6. 江艳娥,林昭进,邱永松,方展强. 中国海域棘头梅童鱼头棘和鳞片轮纹形态的初步研究(英文). 江西农业学报. 2012(04): 157-159+162 . 百度学术
7. 郑德锋,赵金良,周文玉. 棘头梅童鱼线粒体控制区的序列变异与群体遗传结构. 渔业科学进展. 2011(02): 34-40 . 百度学术
8. 薛泰强,杜宁,高天翔. 基于线粒体COI及Cytb基因的4种鱚科鱼类系统发育研究. 中国海洋大学学报(自然科学版). 2010(S1): 91-98 . 百度学术
其他类型引用(13)




计量
- 文章访问数: 4924
- HTML全文浏览量: 200
- PDF下载量: 2659
- 被引次数: 21
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
