
Study on habitat suitability of Coilia mystus in Pearl River Estuary, China
-
摘要: 探究珠江河口洄游性鱼类凤鲚 (Coilia mystus) 的生境适宜度,可为其渔业资源保护和利用提供科学依据。根据2019年冬季和2020年夏季在珠江河口进行的底拖网调查数据,选取水深、底层盐度、底层水温、底层溶解氧等作为指示因子,利用层次分析法 (Analytic hierarchy process, AHP) 计算各指示因子的权重,采用算数平均法 (Arithmetic Mean Model, AMM) 和几何平均法 (Geometric Mean Model, GMM) 计算栖息地适宜度指数 (Habitat Suitability Index, HSI),验证发现基于GMM且赋予权重的HSI模型精确度较高,能较好地反映珠江河口凤鲚的栖息地分布。最优模型结果显示,珠江河口冬季和夏季凤鲚适宜的底层盐度分别为21.03‰~30.55‰和20.25‰~25.82‰,溶解氧质量浓度分别为6.79~7.28和3.85~4.01 mg·L−1,水深分别为11.69~20.10和7.80~9.23 m,底层水温分别为19.84~19.95和28.36~29.44 ℃。珠江河口凤鲚HSI大于0.5的站点仅占总调查站点的17.39%,表明其栖息地适宜性较低,目前珠江河口适宜凤鲚的栖息地主要位于内伶仃海域的中华白海豚自然保护区。Abstract: Exploring the habitat suitability of Coilia mystus, a migratory fish in the Pearl River estuary, can provide a scientific basis for the protection and utilization of fishery resources of C. mystus. Based on the date collected by the bottom-trawl in the Pearl River Estuary during winter in 2019 and summer in 2020, we constructed the habitat suitability index (HSI) model of C. mystus, taking the water depth, bottom salinity, bottom temperature, as well as bottom dissolved oxygen as indicator environment indicators. The analytic hierarchy process (AHP) was used to calculate the weight of the indicators. The arithmetic mean (AMM) and the geometric mean (GMM) were used to build a model. Then a suitability curve was developed and the HSI of C. mystus was calculated. Results show that the optimal bottom salinities for C. mystus in the Pearl River Estuary in winter and summer were 21.03‰−30.55‰ and 20.25‰−25.82‰, respectively; the mass concentrations of suitable dissolved oxygen were 6.79–7.28 and 3.85–4.01 mg·L−1, respectively; the suitable water depths were 11.69–20.10 and 7.80–9.23 m, respectively; the suitable bottom temperatures were 19.84–19.95 and 28.36–29.44 ℃, respectively. The sites with HSI value greater than 0.5 accounted for 17.39% of the total survey sites. The study shows that the suitability of C. mystus was low, and its suitable habitats in the Pearl River Estuary are mainly located in the Sousa chinensis Nature Reserve in the Neilingding sea area.
-
Keywords:
- Coilia mystus /
- Habitat suitability index /
- Environment factors /
- Pearl River Estuary
-
珠江河口是我国三大河口之一,其饵料丰富,是许多经济鱼、虾、蟹的产卵和索饵场,同时也是中华白海豚 (Sousa chinensis) 等多种珍稀水生动物的栖息地。主要经济种类有凤鲚 (Coilia mystus)、棘头梅童鱼 (Collichthys lucidus)、多齿蛇鲻 (Saurida tumbil)、日本金线鱼 (Nemipterus virgatus)等[1]。近年来,随着珠江三角洲经济的发展,环境污染、过度捕捞等人类活动导致鱼类栖息环境受到不同程度的影响,生物多样性降低,珠江河口海域游泳动物经济物种群落结构发生明显变化。研究发现,以2000年为时间界线,之前珠江河口海域的游泳动物经济物种群落结构差异明显,之后游泳动物经济物种种类数量显著低于之前,20世纪80年代的4次调查共记录珠江河口鱼类330种,2013—2016年调查中仅记录鱼类285种[2-4]。2015年在珠江河口南沙海域进行的渔业资源调查情况相较于2002年同海域,渔获物种类下降了80%,渔获物规格也发生明显变化[5]。
凤鲚俗称凤尾鱼,隶属鲱形目、鳀科,属河口洄游性小型鱼类,在我国黄海、南海、东海海域均有分布,其主要有3个地方种群,即长江型、闽江型、珠江型,每年春季集群在河口咸水或咸淡水区进行繁殖[6-7]。研究发现,2000年以前珠江河口凤鲚生物量占比超过10%,渔获率超过10%,2000年以后,其生物量和渔获率均低于10%。与历史资料相比,南沙海域及伶仃洋海域凤鲚的体长与体质量均显著降低[4,8]。目前对凤鲚的研究主要集中在长江口海域,探讨其食性[9-11]、繁殖生物学[12-13]及资源状况等[14-16]。尚未见有关珠江河口凤鲚的栖息地适宜性的研究报道。
栖息地适宜性指数 (Habitat Suitability Index, HSI) 模型是一种用于研究野生动物栖息质量的研究方法,具有简单、易理解、可清晰表达物种的最适环境条件等优势[17-18],被广泛用于探究环境因子对鱼类空间分布的影响及描述物种对环境的偏好程度[19-20]。水温、水深、盐度、溶解氧 (DO)、化学需氧量 (COD) 等非生物环境因子是鱼类栖息地适宜性研究中常用的关键环境因子[21-22]。凤鲚是典型的河口洄游性中下层鱼类,盐度和DO的变化对其空间分布影响较大[17]。毕雪娟[23]利用盐度、浊度、水深、温度建立长江口凤鲚HSI模型;胡丽娟等[12]利用盐度、水温和DO建立广义可加模型 (Generalized additive models, GAM)探究凤鲚仔稚鱼栖息地适宜度。本文以珠江河口优势种凤鲚为研究对象,结合以往凤鲚栖息地的相关研究,选取水深、底层盐度、底层DO和底层水温作为环境因子,构建珠江河口凤鲚的HSI模型,探究珠江河口凤鲚对栖息地的偏好,分析影响珠江河口凤鲚栖息地的关键环境因子,以期为珠江河口凤鲚的渔业资源保护和利用提供科学的理论基础。
1. 材料与方法
1.1 数据来源
凤鲚样品来源于2019年12月 (冬季) 和2020年8月 (夏季) 在珠江河口海域 (113.30°E—114.00°E、21.80°N—23.00°N) 进行的生态调查,根据纬度、盐度等差异,每个航次随机选取一定数量的站位进行底拖网渔业资源调查,2019年冬季和2020年夏季分别设置10和13个渔业资源站位 (图1),调查站位图采用ArcGis 10.6叠加标准地图绘制。调查船为143 kW的单拖渔船,拖速3~3.5 kn,拖拽时间0.5 h,调查网具网口宽度36 m,网囊网目30 mm。记录每个站位渔获物的种类、数量、生物量、体长等生物学信息。并在每个调查站位同步使用YSI 556 MPS现场测定底层盐度、底层DO、底层水温、水深等非生物环境数据并记录;所有样品的采集、测定均按照GB 12763—2007《海洋调查规范》要求进行。
1.2 数据分析
1.2.1 HSI
采用单位面积渔获物尾数作为相对资源密度指数 (尾·km−2),开发基于资源密度的适宜性指数 (Suitability Index, SI)。
单位面积渔获物尾数采用扫海面积法估算,公式为:
$$ P=C/(a·q) $$ (1) 式中:P为相对资源密度 (kg∙km−2);C为每小时渔获量 (kg∙h−1);a为拖网每小时的扫海面积 (km2∙h−1),为拖网的拖拽距离和网口水平扩张宽度的乘积;q为网具的捕捞率,参考以往凤鲚资源的相关研究结果,选取0.5[24]。
SI的计算公式为:
$$ {\mathrm{S}\mathrm{I}}_{i}=\dfrac{{Y}_{i}}{{\mathrm{Y}}_{i,\mathrm{m}\mathrm{a}\mathrm{x}}} $$ (2) 式中:SIi为该季节i站的适宜性指数;Yi为该季节i站单位面积渔获尾数;Yi,max为该季最大单位渔获物尾数。SI的取值范围为0~1。SI越接近1,表示适宜性指数越高;SI越接近0,表示适宜性越低;SI介于0.5~1.0时认为适宜鱼类栖息。
1.2.2 环境因子
鉴于凤鲚为中下层河口洄游性鱼类,结合以往关于凤鲚栖息地的研究结果[12,23],本研究选取水深、底层盐度、底层水温、底层DO为环境因子建立珠江河口凤鲚的HSI模型。珠江河口冬季和夏季盐度变化较大,且盐度是影响珠江河口鱼类群落结构的主要环境因子[25],因此认为底层盐度对珠江河口凤鲚空间分布的影响性最大;DO也是对河口鱼类群落丰富度影响较大的环境因子之一[23],赋予DO次于盐度的贡献率;水深对鱼类的生长具有直接影响,也是鱼类栖息地适宜性评价中常用的环境因子[26];水温直接影响鱼类的性腺发育,已有研究表明水深对凤鲚栖息地的影响大于水温[23]。因此,本研究各环境因子的相对重要性依次为底层盐度>底层DO>水深>底层水温。利用层次分析法 (Analytic hierarchy process, AHP) 计算各环境因子的权重,根据环境因子两两之间的相对重要性之比,构建指示因子的判别矩阵,利用软件MATLAB 9.7计算一致性比率 (Consistent ratio, CR),一般认为CR<0.1时判断矩阵通过一致性检验,即各环境因子的权重分配合理[27]。然后采用一元非线性函数拟合各环境因子与SI的关系。
1.2.3 HSI模型
采用算数平均法 (Arithmetic Mean Model, AMM) 和几何平均法 (Geometric Mean Model, GMM) 建立HSI模型,计算公式分别如下:
$$ {\mathrm{H}\mathrm{S}\mathrm{I}}_{\mathrm{A}\mathrm{M}\mathrm{M}}=\dfrac{1}{\displaystyle \sum _{i=1}^{n}{\omega }_{i}}·\displaystyle \sum _{i=1}^{n}{\mathrm{S}\mathrm{I}}_{i}{\omega }_{i} $$ (3) $$ {\mathrm{H}\mathrm{S}\mathrm{I}}_{\mathrm{G}\mathrm{M}\mathrm{M}}={\left(\prod _{i=1}^{n}{\mathrm{S}\mathrm{I}}_{i}^{{\omega }_{i}}\right)}^{\tfrac{1}{\sum _{i=1}^{n}{\omega }_{i}}} $$ (4) 式中:ωi为各环境因子的权重;SIi为各环境因子的SI值。
1.2.4 模型的验证
根据各因子的权重基于AMM和GMM建立HSI模型,分别计算2019年冬季、2020年夏季各站点的HSI,使用SPSS 19.0软件将各站位计算所得的HSI与实际调查获得的凤鲚资源密度进行比较分析,以验证模型的效果并选取最优模型。
2. 结果
2.1 环境因子
经多次调整判断矩阵,各环境因子的判断矩阵见表1。矩阵的CR为0.001 9,小于0.1,说明该判别矩阵具有良好的一致性,影响珠江河口凤鲚栖息地分布的各个环境因子的权重分配合理。底层盐度、底层DO、水深和底层水温的权重分别为0.387 1、0.272 8、0.182 8和0.157 4。
表 1 指示因子的判断矩阵Table 1. Judgment matrix of indicator factors指示因子Indicator factor 底层盐度Bottom salinity 底层溶解氧Bottom DO 水深Water depth 底层水温Bottom temperature 底层盐度Bottom salinity 1 3 5 5 底层溶解氧Bottom DO 0.33 1 3 4 水深Water depth 0.20 0.33 1 2 底层水温Bottom temperature 0.20 0.25 0.5 1 2.2 环境因子的适宜度曲线
冬季珠江河口凤鲚分布海域的底层盐度介于3.40‰~30.20‰,适宜的底层盐度介于21.03‰~30.55‰;凤鲚分布海域的底层DO质量浓度介于5.91~8.16 mg·L−1,适宜的DO质量浓度介于6.79~7.28 mg·L−1;分布海域的水深介于4.50~24.90 m,适宜的水深介于11.69~20.10 m;分布海域的底层水温介于19.10~20.50 ℃,适宜的底层水温介于19.84~19.95 ℃。
夏季珠江河口凤鲚分布海域的底层盐度介于3.24‰~34.66‰,适宜的底层盐度介于20.25‰~25.82‰;分布海域的底层DO质量浓度介于3.09~8.59 mg·L−1,适宜的DO质量浓度介于3.85~4.01 mg·L−1;分布海域的水深介于7.00~26.70 m,适宜的水深介于7.80~9.23 m;分布海域的底层水温介于24.30~31.80 ℃,适宜的底层水温介于28.36~29.44 ℃ (图2)。
![]() 图 2 珠江河口凤鲚冬、夏季底层盐度、底层溶解氧、深度、底层水温的适宜性指数曲线Figure 2. Suitability index curves of bottom salinity, bottom DO, depth and bottom water temperature of C. mystus in Pearl River Estuary during winter and summer
图 2 珠江河口凤鲚冬、夏季底层盐度、底层溶解氧、深度、底层水温的适宜性指数曲线Figure 2. Suitability index curves of bottom salinity, bottom DO, depth and bottom water temperature of C. mystus in Pearl River Estuary during winter and summer2.3 HSI模型验证
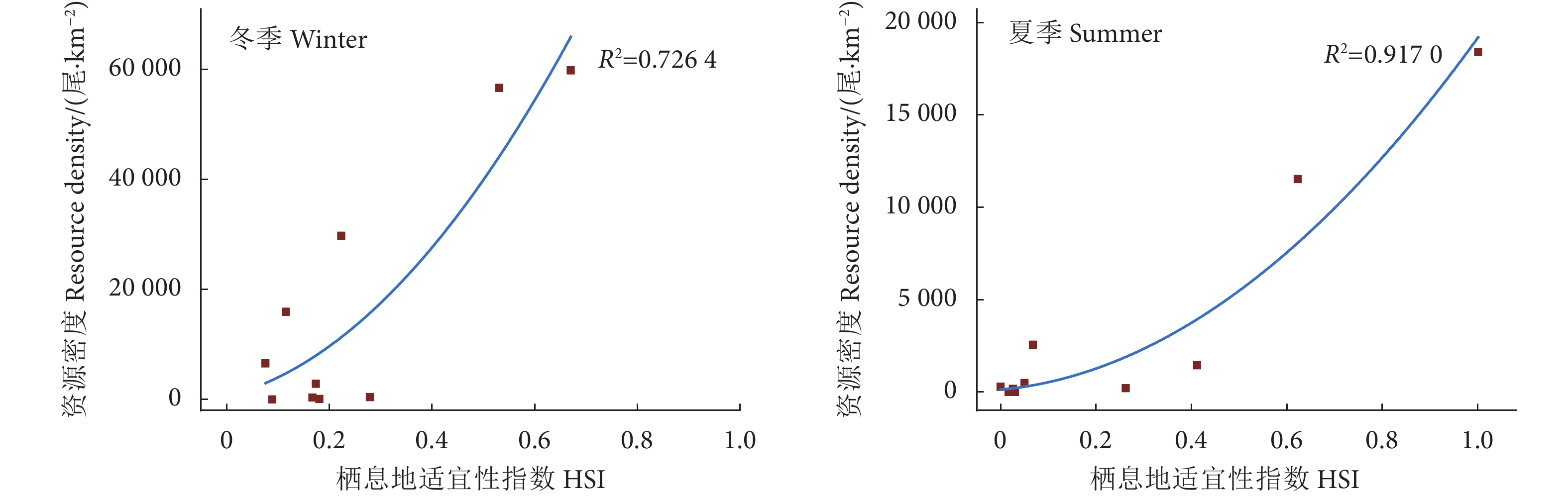
利用AMM和GMM,结合层次分析法计算的环境因子相应的权重计算得到的各站点HSI,将各站点的HIS值与实际凤鲚资源密度进行相关性分析,结果显示,两种计算方法各站点的HSI均与凤鲚实际丰度具有显著相关性 (P<0.05)。但赋予权重的GMM算法的计算结果与凤鲚实际资源的相关性更为显著。因此本研究选取基于GMM算法并赋予权重的HSI模型研究珠江河口凤鲚的栖息地适宜性 (图3)。
![]() 图 3 珠江河口凤鲚冬、夏季实际资源密度与栖息地适宜性指数的相关性Figure 3. Pearson correlation between actual resource density of C. mystus and HSI in Pearl River Estuary during winter and summer
图 3 珠江河口凤鲚冬、夏季实际资源密度与栖息地适宜性指数的相关性Figure 3. Pearson correlation between actual resource density of C. mystus and HSI in Pearl River Estuary during winter and summer2.4 珠江河口凤鲚最适栖息地的分布
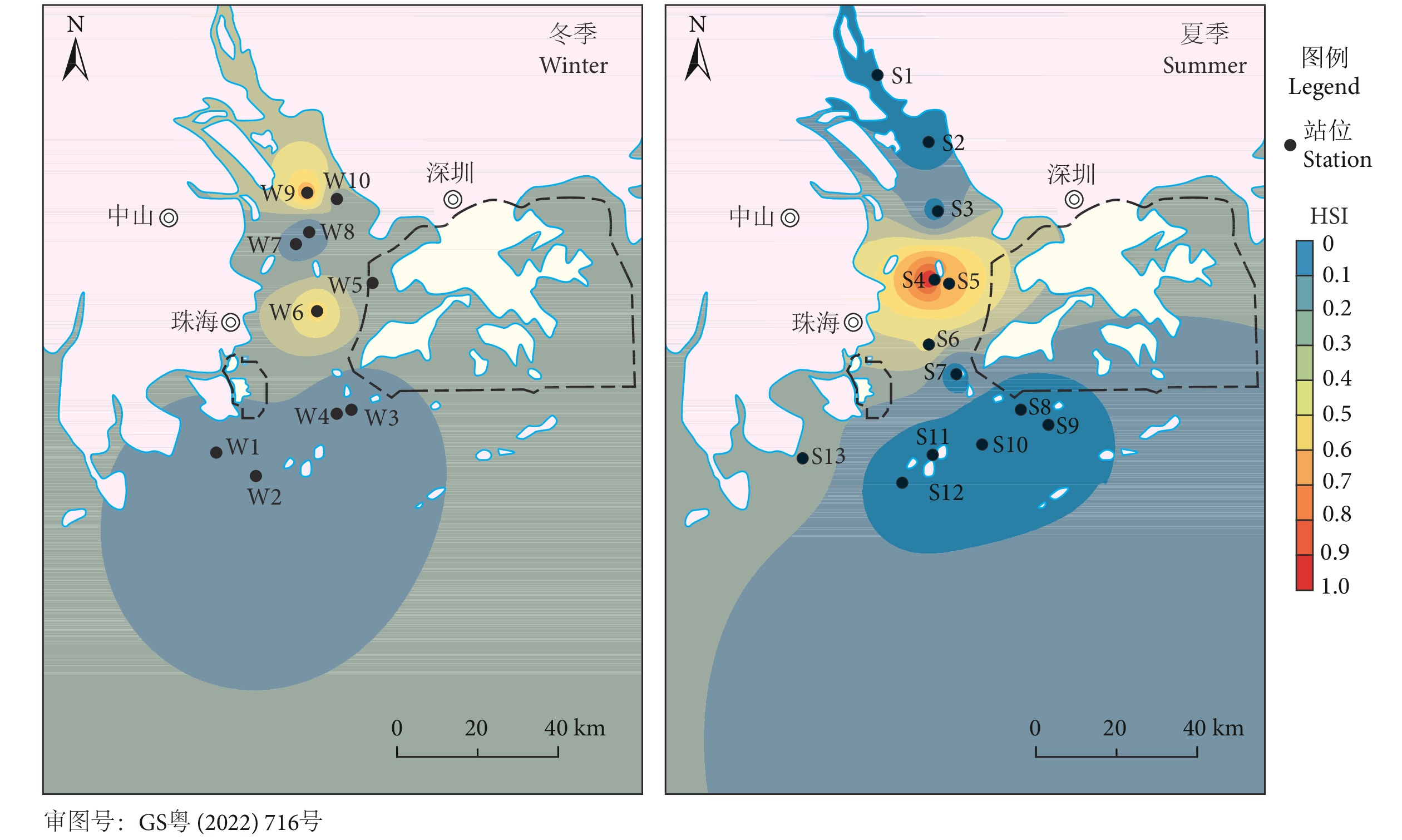
选取最优模型计算各站点的HSI,绘制珠江河口凤鲚的HSI分布图 (图4)。结果显示,珠江河口海域凤鲚空间分布季节差异较大,整体栖息地适宜性相对较低。冬季10个调查站点,HSI大于0.5的仅有W6和W9站点,W6站点的HSI为0.53,位于珠江河口的中华白海豚自然保护区港珠澳大桥中段北侧。W9站点的HSI为0.67,位于南沙的龙穴岛东南方向的近岸海域。夏季13个调查站点中仅有S4和S5站点的HSI大于0.5,其中S4站点的HSI为1,位于珠海淇澳岛和内伶仃岛连线之间的海域,S5站点的HSI为0.62,与S4站点毗邻。本研究调查海域内冬季和夏季凤鲚HSI大于0.5的站点共计4个,占17.39%,其中3个均位于内伶仃海域的中华白海豚自然保护区。
![]() 图 4 珠江河口凤鲚冬、夏季HSI分布平面图Figure 4. Distribution of HSI of C. mystus in Pearl River Estuary during winter and summer
图 4 珠江河口凤鲚冬、夏季HSI分布平面图Figure 4. Distribution of HSI of C. mystus in Pearl River Estuary during winter and summer3. 讨论
3.1 影响珠江河口凤鲚栖息地分布的环境因子
本研究中,影响珠江河口凤鲚栖息地分布的环境因子按重要性排序依次为底层盐度、底层DO、水深、底层水温。盐度可通过影响鱼类渗透压及体内酶活性,进而影响鱼类的生长发育和新陈代谢,已有研究发现盐度是影响河口鱼类群落分布的重要因素[28-29]。本研究中珠江河口夏季凤鲚适宜的盐度介于20.25‰~25.82‰,与李开枝等[8]珠江河口伶仃洋海域夏季凤鲚适宜的盐度低于25‰的结论基本一致,冬季凤鲚适宜的盐度介于21.03‰~30.55‰,略高于夏季,这可能与珠江河口夏季受珠江径流影响,大量淡水入海,造成盐度偏低有关。另外,晁眉等[30]研究表明冬季凤鲚会游向咸水区生活,这也可能是冬季珠江河口凤鲚适宜的盐度高于夏季的原因之一。已有研究表明水深和温度是影响海州湾皮氏叫姑鱼 (Johnius belangerii)[31]和浙江南部黄鲫 (Setipinna tenuifilis)[32]空间分布的重要环境因子。水深会影响水体的透明度和初级生产力,水温则会影响鱼类的性腺发育、产卵和生长等[33],本研究发现珠江河口凤鲚夏季适宜水深介于7.80~9.23 m,适宜水温介于20.25~25.82 ℃,与毕雪娟[23]和詹海刚[34]的研究结果也较为一致,因此本研究选取水深和底层水温作为研究珠江河口凤鲚栖息地分布的环境因子具有一定的合理性。有研究发现珠江河口夏、冬季桡足类的丰度分布具有明显的季节和地理差异,且从河口上游至万山群岛海域,桡足类种类逐渐增多[35],凤鲚以浮游动物为食,夏、冬季均主要摄食桡足类[9],因此珠江河口桡足类丰度和种类的分布差异也可能影响珠江河口凤鲚栖息地分布。DO对鱼类的生长至关重要,冬季珠江河口凤鲚适宜的DO质量浓度介于6.79~7.28 mg·L−1,这与胡丽娟等[12]的研究结果一致,夏季适宜的DO质量浓度低于冬季 (3.85~4.01 mg·L−1),这可能与冬季水温低,水体DO质量浓度高有关。
3.2 珠江河口凤鲚的栖息地分布特征
珠江河口同时受径流和外海高盐海水的影响,从虎门水域至担杆岛海域,盐度分布呈淡水、咸淡水、高盐水分布的特征[36]。冬季的W6站点和夏季HSI较大的两个站点 (S4和S5) 均位于淇澳岛与内伶仃岛联线之间的海域,该海域位于珠江河口内伶仃洋海域的中华白海豚国家级自然保护区。李开枝等[8]研究发现夏、冬季凤鲚主要分布在伶仃洋水域;詹海刚[34]研究发现在丰水期伶仃洋水域凤鲚的个体数及生物量占绝对优势;肖尤盛等[36]、黄吉万等[37]对中华白海豚保护区的调查研究发现凤鲚是夏季该海域的优势种。这可能是因为中华白海豚保护区属于典型的咸淡水交汇区,且受径流影响,营养物质丰富,有利于凤鲚在该水域栖息生长;另外保护区内的水质等环境条件也相对较好[34],也可能是此海域适宜凤鲚栖息的原因之一。凤鲚的繁殖时间在5—8月[13],凤鲚群体繁殖后在该海域索饵也可能是造成该海域凤鲚分布较多的原因。冬季HSI最大的W9站点位于南沙龙穴岛的东南方向的离岸海域,推测可能是由于该海域处于珠江虎门和蕉门的出水口交界处,咸淡水交替,营养物质丰富,更适宜凤鲚这类咸淡水洄游的河口鱼类生长,该海域的浮游动物种类较为丰富,其中小型桡足类种类占绝对优势[38],因此该海域丰富的饵料生物更有利于凤鲚觅食。
3.3 HSI模型的构建
利用HSI模型评估鱼类的栖息地适宜性,环境因子的权重、环境因子的适宜性曲线对于模型的建立至关重要[39]。本研究选取水深、底层盐度、底层DO、底层水温4个环境因子构建珠江河口凤鲚的栖息地指数模型,既考虑了水质,又考虑了水文因素,更全面地评估了生物与非生物因子对珠江河口凤鲚栖息地的影响;相比仅参考以往相关文献而确定环境因子的权重,本研究根据凤鲚渔业资源密度与各环境因子的相关性分析结果构建了判断矩阵,并使用AHP计算各环境因子的权重,更具有科学性。
本研究采用非线性函数拟合各环境因子的适宜度曲线,除夏季水深外,其余各环境因子适宜度曲线的决定系数R2均大于0.70,说明各环境因子的适宜度曲线拟合效果较好。本研究使用实际调查获得的凤鲚资源密度数据用于模型验证,将计算所得的各站点HSI与两个季度珠江河口渔业资源调查中实际的凤鲚资源密度进行相关性分析,结果显示强相关 (R2=0.7264, R2=0.9170),说明本研究基于GMM构建的HSI模型对于珠江河口凤鲚的栖息地适宜性模型具有较好的拟合度和精度 (图3)。因此,本研究结果在探究影响珠江河口凤鲚的栖息地适宜性的关键环境因子及空间分布等方面具有一定的指导作用。
-
![]()
图 2 珠江河口凤鲚冬、夏季底层盐度、底层溶解氧、深度、底层水温的适宜性指数曲线
Figure 2. Suitability index curves of bottom salinity, bottom DO, depth and bottom water temperature of C. mystus in Pearl River Estuary during winter and summer
![]()
图 3 珠江河口凤鲚冬、夏季实际资源密度与栖息地适宜性指数的相关性
Figure 3. Pearson correlation between actual resource density of C. mystus and HSI in Pearl River Estuary during winter and summer
![]()
图 4 珠江河口凤鲚冬、夏季HSI分布平面图
Figure 4. Distribution of HSI of C. mystus in Pearl River Estuary during winter and summer
表 1 指示因子的判断矩阵
Table 1 Judgment matrix of indicator factors
指示因子Indicator factor 底层盐度Bottom salinity 底层溶解氧Bottom DO 水深Water depth 底层水温Bottom temperature 底层盐度Bottom salinity 1 3 5 5 底层溶解氧Bottom DO 0.33 1 3 4 水深Water depth 0.20 0.33 1 2 底层水温Bottom temperature 0.20 0.25 0.5 1  下载: 导出CSV
下载: 导出CSV
-
[1] 林坤, 麦广铭, 王学锋. 2015—2018年珠江口近岸海域鱼类群落结构及其稳定性[J]. 水产学报, 2020, 44(11): 1841-1850. [2] 庄平, 李桂峰. 珠江口鱼类资源多样性与资源保护 [M]. 北京: 中国农业出版社, 2018: 19-47. [3] 费鸿年, 何宝全, 陈国铭. 南海北部大陆架底栖鱼群聚的多样度以及优势种区域和季节变化[J]. 水产学报, 1981(1): 1-20. [4] 徐鹏, 谢木娇, 周卫国, 等. 近30年珠江口海域游泳动物经济物种群落结构变化特征[J]. 应用海洋学学报, 2021, 40(2): 239-250. doi: 10.3969/J.ISSN.2095-4972.2021.02.007 [5] 袁梦, 汤勇, 徐姗楠, 等. 珠江口南沙海域秋季渔业资源群落结构特征[J]. 南方水产科学, 2017, 13(2): 18-25. doi: 10.3969/j.issn.2095-0780.2017.02.003 [6] 袁传宓, 秦安舲. 我国近海鲚鱼生态习性及其产量变动状况[J]. 海洋科学, 1984(5): 35-37. [7] JIANG T, LIU H B, HUANG H H, et al. Migration patterns and habitat use of the tapertail anchovy Coilia mystus in the Oujiang River Estuary and the Zhujiang River Estuary, China[J]. Acta Oceanol Sin, 2019, 38(8): 35-40. doi: 10.1007/s13131-019-1436-0
[8] 李开枝, 尹健强, 黄良民, 等. 珠江口伶仃洋海域底层游泳动物的季节变化[J]. 生态科学, 2012, 31(1): 2-8. doi: 10.3969/j.issn.1008-8873.2012.01.001 [9] 刘守海, 徐兆礼, 田丰歌. 长江口及附近水域凤鲚摄食习性的分析[J]. 上海海洋大学学报, 2012, 21(4): 589-597. [10] 郭爱, 陈峰, 金海卫, 等. 东、黄海凤鲚的食物组成及其食性的季节变化[J]. 海洋渔业, 2014, 36(5): 402-408. doi: 10.3969/j.issn.1004-2490.2014.05.003 [11] CUI Y, WU Y, XU Z L, et al. Potential dietary influence on the stable isotopes and fatty acid composition of migratory anchovy (Coilia mystus) around the Changjiang Estuary[J]. J Mar Biol Assoc UK, 2014, 95(1): 1-13.
[12] 胡丽娟, 宋超, 耿智, 等. 长江口凤鲚仔稚鱼主要繁殖季节的时空分布特征[J]. 中国水产科学, 2021, 28(9): 1152-1161. [13] 洪波, 周轩, 王淼. 杭州湾北部海域凤鲚个体繁殖力初步研究[J]. 水产科技情报, 2021, 48(6): 335-341. doi: 10.16446/j.fsti.20201200123 [14] 倪健夫, 郭弘艺, 唐文乔, 等. 长江口汛期凤鲚捕捞量年际变化研究[J]. 海洋渔业, 2020, 42(2): 192-204. [15] WANG L L, LIN L, LI Y, et al. Sustainable exploitation of dominant fishes in the largest estuary in southeastern China[J]. Water, 2020, 12: 3390. doi: 10.3390/w12123390
[16] WANG C, KYNARD B, WEI Q, et al. Spatial distribution and habitat suitability indices for non-spawning and spawning adult Chinese sturgeons below Gezhouba Dam, Yangtze River: effects of river alterations[J]. J Appl Ichthyol, 2013, 29(1): 1-10. doi: 10.1111/jai.12157
[17] 周为峰, 李英雪, 程田飞, 等. 栖息地适宜性指数模型在鱼类生境评价中的应用进展[J]. 渔业信息与战略, 2020, 35(1): 48-54. doi: 10.13233/j.cnki.fishis.2020.01.007 [18] GONG C X, CHEN X J, GAO F, et al. Effect of spatial and temporal scales on habitat suitability modeling: a case study of Ommastrephes bartramii in the northwest Pacific Ocean[J]. J Ocean Univ China, 2014, 13(6): 1-11.
[19] ZHANG Z P, ZHOU J, SONG J J, et al. Habitat suitability index model of the sea cucumber Apostichopus japonicus (Selenka): a case study of Shandong Peninsula, China[J]. Mar Pollut Bull, 2017, 122(1/2): 1-12.
[20] 范秀梅, 杨胜龙, 张胜茂, 等. 基于栖息地指数的阿拉伯海鲐鱼渔情预报模型构建[J]. 南方水产科学, 2020, 16(4): 8-17. doi: 10.12131/20190255 [21] 邹易阳, 薛莹, 麻秋云, 等. 应用栖息地适宜性指数研究海州湾小黄鱼的空间分布特征[J]. 中国海洋大学学报, 2016, 46(8): 54-63. [22] BROWN S K, BUJA K R, JURY S H, et al. Habitat Suitability Index models for eight fish and invertebrate species in Casco and Sheepscot Bays, Maine[J]. N Am J Fish Manag, 2011, 20(2): 1-28.
[23] 毕雪娟. 长江口凤鲚繁殖生物学及HSI评估 [D]. 上海: 上海海洋大学, 2015: 43-63. [24] 施炜纲, 王博. 长江河口区凤鲚的资源现状[J]. 水生生物学报, 2002, 26(6): 648-653. doi: 10.3321/j.issn:1000-3207.2002.06.012 [25] 莫伟均, 何用, 王建平, 等. 珠江河口鱼类分布现状及栖息水域特征[J]. 应用与环境生物学报, 2022, 28(5): 1-10. doi: 10.19675/j.cnki.1006-687x.2021.05048 [26] WU J H, DAI L B, DAI X J, et al. Comparison of generalized additive model and boosted regression tree in predicting fish community diversity in the Yangtze River Estuary, China[J]. J Appl Ecol, 2019, 30(2): 1-9.
[27] 邓雪, 李家铭, 曾浩健. 等. 层次分析法权重计算方法分析及其应用研究[J]. 数学的实践与认识, 2012, 42(7): 93-100. doi: 10.3969/j.issn.1000-0984.2012.07.012 [28] 王云峰, 朱鑫华. 盐度对鱼类生态生理学特征的影响[J]. 海洋科学集刊, 2002(1): 151-158. [29] 孙艳秋, 刘鉴毅, 庄平, 等. 温度、盐度和pH对多纹钱蝶鱼胚胎发育的影响[J]. 南方水产科学, 2021, 17(6): 122-129. doi: 10.12131/20210109 [30] 晁眉, 黄良敏, 李军, 等. 福建九龙江口凤鲚的生物学特征[J]. 集美大学学报(自然科学版), 2016, 21(1): 16-20. [31] 张云雷, 薛莹, 于华明, 等. 海州湾春季皮氏叫姑鱼栖息地适宜性研究[J]. 海洋学报, 2018, 40(6): 83-91. [32] 柳晓雪, 高春霞, 田思泉, 等. 基于栖息地适宜指数的浙江南部近海黄鲫最适栖息地分布[J]. 中国水产科学, 2020, 27(12): 1485-1495. [33] 邓吉河. 浅谈水温与鱼类的关系[J]. 黑龙江水产, 2019(1): 25-27. doi: 10.3969/j.issn.1674-2419.2019.01.009 [34] 詹海刚. 珠江口及邻近水域鱼类群落结构研究[J]. 海洋学报, 1998, 20(3): 91-97. [35] 黄彬彬, 郑淑娴, 蔡伟叙, 等. 珠江口枯水期和丰水期中小型桡足类种类组成、丰度分布及其与环境因子的关系[J]. 厦门大学学报(自然科学版), 2017, 56(6): 852-858. [36] 肖尤盛, 陈海亮, 王雪辉, 等. 珠江口中华白海豚的食物变化[J]. 海洋与渔业, 2018(10): 72-73. [37] 黄吉万, 孙典荣, 刘岩, 等. 珠江口中华白海豚自然保护区鱼类群落多样性分析[J]. 南方农业学报, 2018, 49(5): 1000-1007. doi: 10.3969/j.issn.2095-1191.2018.05.25 [38] 徐姗楠, 龚玉艳, 詹凤娉, 等. 珠江口海域南沙段浮游动物群落生态特征[J]. 海洋湖沼通报, 2017(6): 82-90. [39] 官文江, 田思泉, 朱江峰, 等. 渔业资源评估模型的研究现状与展望[J]. 中国水产科学, 2013, 20(5): 1112-1120. -
期刊类型引用(4)
1. 郑秋实,刘永,李纯厚,吴鹏,肖雅元,林琳,刘玉,邹剑. 季节变化对珠江口棘头梅童鱼肠道和水体中细菌群落结构的影响. 南方水产科学. 2024(04): 77-87 .  本站查看
本站查看
2. 武智,李跃飞,朱书礼,杨计平,夏雨果,李捷. 基于渔业声学调查的珠江东塔产卵场鱼类栖息地适宜性研究. 南方水产科学. 2023(03): 11-18 . 本站查看
3. 李纯厚,齐占会. 中国渔业生态环境学科研究进展与展望. 水产学报. 2023(11): 132-147 . 百度学术
4. 郑秋实,刘永,吴鹏,李纯厚,肖雅元,唐广隆,郭智成,吴佳鹏,洪义国. 珠江口优势种鱼类肠道与水体中的微生物组成关系. 农业环境科学学报. 2023(11): 2531-2540 . 百度学术
其他类型引用(2)

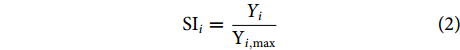
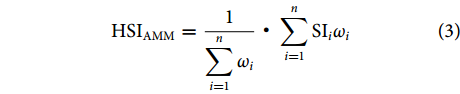
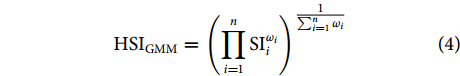


计量
- 文章访问数: 571
- HTML全文浏览量: 167
- PDF下载量: 69
- 被引次数: 6
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
