
Effects of dietary microalgae on growth and survival of larval development of sea urchin (Anthocidaris crassispina)
-
摘要:
文章采用湛江叉鞭金藻 (Dicrateria zhanjiangensis)、纤细角毛藻 (Chaetoceros gracilis)、绿色巴夫藻 (Pavlova viridis)、扁藻 (Platymonas subcordiformis)、小球藻 (Chlorella vulgaris)及混合藻 (金藻和角毛藻1∶1混合) 投喂紫海胆 (Anthocidaris crassispina),研究了微藻饵料对紫海胆浮游幼体生长发育的影响。结果表明,不同饵料对紫海胆浮游幼体生长发育的效果有差异,金藻是最佳饵料品种,角毛藻次之,其后分别为巴夫藻、小球藻和混合藻,扁藻不适于作为紫海胆浮游幼体的开口饵料。就饵料种类而言,金藻作为紫海胆浮游幼体饵料的成活率可达 (23.12±1.8)%,显著高于其他单胞藻类 (P<0.05)。就发育速度而言,以角毛藻和金藻混合喂食的海胆浮游幼体整体发育速度最快,幼体平均体长为 (885.25±30.49) μm,显著高于其他5种处理 (P<0.05);投喂巴夫藻的浮游幼体前期生长较慢,体长为 (337.98±24.56) μm,后期较其他单胞藻类生长快,体长可达 (580±32.95) μm,适宜作为紫海胆浮游幼体后期饵料。
Abstract:To investigate the growth and survival of sea urchin (Anthocidaris crassispina) larvae, we fed six microalgal diets [Dicrateria zhanjiangensis, Chaetoceros gracilis, Pavlova viridis, Platymonas subcordiformis, Chlorella vulgaris, and mixture of C. gracilis and D. zhanjiangensis (1∶1)]. The results show that different diets had different effects on the growth and development of the planktonic larvae of sea urchin. D. zhanjiangensis was the optimal microalgal diet. C. gracilis was the second, and then were P. viridis, C. vulgari, and mixture of C. gracilis and D. zhanjiangensis. However, P. subcordiformis was not suitable to be used as the opening bait for sea urchin larvae rearing. In terms of baits, D. zhanjiangensis was the best, since the survival rate of the larvae could reach (23.12 ±1.8)%, significantly higher than those of other unicellular algae (P<0.05). In terms of growth rate, mixture of C. gracilis and D. zhanjiangensis was the best, since the average length of the larvae could reach (885.25±30.49) μm, significantly higher than those of the other five treatments (P<0.05). The growth of the larvae fed with P. viridis was slow at early stage with body length of (337.98±24.56) μm, but it grew faster than the other unicellular algae at later stage with body length of (580±32.95) μm, so P. viridis is the suitable bait for planktonic larvae of A. crassispina at later stage.
-
Keywords:
- Anthocidaris crassispina /
- Microalgal /
- Growth and development /
- Planktonic larvae
-
河豚毒素(TTX)为氨基全氢喹唑啉化合物,是一种典型的非蛋白类神经毒素,能造成神经麻痹,严重患者会因为中枢神经麻痹导致呼吸停止而死亡[1-2]。TTX的毒性是氰化物的1 250多倍[3],最初在河鲀体内发现,后陆续发现产TTX的细菌[4-5],在织纹螺(Nassarius)等多种动物体内也检出TTX[6-8]。织纹螺为腐食性生物,可通过食物链富集TTX,近年来,在中国台湾[9]、福建[10]、江苏[11]等沿海地区发生多起因食用织纹螺引起的中毒事件,导致多人死亡,引起了相关部门的高度关注,通过从织纹螺体内分离出产TTX菌株确认TTX为其主要毒素[12]。由于织纹螺中常含有较高浓度的TTX,中毒后没有特效解毒剂,因此需要建立织纹螺中TTX的检测方法,对螺类生物的TTX含量进行监控,减少食物中毒事件的发生。
TTX生物检测存在一定的局限性,其检测结果不稳定,重复性较差。免疫检测法主要有酶联免疫吸附法(ELISA)[13]和免疫层析法(ICA)[14]2种,在食品和农产品残留检测中较为常用[15]。目前液相色谱-质谱联用检测法(LC-MS/MS)是检测的热点[16-17],其灵敏度和准确性都要优于其他方法,但其对检测样品的纯化要求高。样品本底基质复杂,如果样品净化不理想,在上机检测过程易受基质干扰,会出现灵敏度低和回收率低等问题。严忠雍等[18]和Guo等[19]都采用免疫亲和柱对样品进行净化,显著降低了基质效应,后用LC-MS/MS检测,前者的定量限为0.3 μg·kg–1,回收率达到88.7%~102.3%;后者的定量限为1 μg·kg–1,回收率为75.8%~107%。虽然免疫亲和柱特异性强,作为前处理净化方法效果良好,但是价格昂贵,检测成本高[20]。
传统的TTX样品前处理方法较少,且净化效果一般,存在基质干扰严重和假阳性等问题,会对检测结果造成影响。功能性纳米材料能够弥补该不足,其高效、经济的优点让其成为食品检测领域的热点,并已经取得一定研究进展。碳纳米管具有特殊物理结构和化学性质,其表面积大,吸附力强,性质稳定,在检测领域有广泛前景[21]。Taghizadeh等[22]利用纳米管复合材料萃取鱼类、沉积物、土壤和水样中的重金属离子,对3种重金属离子的检测限分别达到了0.09 ng·mL–1、0.72 ng·mL–1和1.0 ng·mL–1,该方法适用于样品中微量重金属离子的萃取;Wang等[23]利用多壁碳纳米管(multi-walled carbon nanotubes,MWCNTs)萃取水样和牛奶样品中的多溴联苯醚,用气相色谱进行分析,检测限为3.6~8.6 ng·L–1,回收率为90%~119%,多壁碳纳米管对多溴联苯醚的吸附效率高;陈啟荣等[24]以乙腈提取茶叶样品中的26种农药成分,用多壁碳纳米管进行净化和富集,用气相色谱进行分析,采用外标法定量,检出限为0.005~0.05 mg·kg–1,定量限为0.015~0.15 mg·kg–1,回收率为66.6%~125.5%,与其他用于检测分析茶叶中残留农药的方法相比,该方法简单方便且回收率和准确性较高。同时,市面上还有多个有关多壁碳纳米管对样品进行检测分析的专利,通过多壁碳纳米管进行吸附,经过洗脱、浓缩,对组分进行定量分析;廖且根等[25]建立了多壁碳纳米管检测水样中的微囊藻毒素含量方法;陈黎等[26]建立了多壁碳纳米管检测烟草中残留农药的方法。这些专利方法均利用多壁碳纳米管材料吸附能力强的特点,且具有用量少、检测成本低等优点。
本研究建立了碳纳米管固相萃取-超高效液相色谱串联质谱法测定织纹螺中河豚毒素的方法,样品经过碳纳米管富集与净化后,经LC-MS/MS测定,用外标法定量。本方法前处理过程简单,结果灵敏度高、准确性好,适用于织纹螺中TTX的残留检测。
1. 材料与方法
1.1 主要仪器与装置
Acquity UPLC I-Class/Xevo TQS超高效液相色谱仪串联三重四级杆质谱仪(美国Waters公司);5810型台式离心机(德国Eppendorf公司);MS3旋涡混合器(德国IKA公司);Milli Q去离子水发生器(美国Millipore公司);N-EVAP氮吹仪(美国Organomation公司);PHS-25 pH计(上海仪电科学仪器股份有限公司)。
1.2 主要材料与试剂
实验材料为氨水(分析纯,广州化学试剂厂)、乙腈、甲醇(均为色谱纯或分析纯,美国Sigma公司),TTX标准品纯度≥99.0% (美国Sigma公司),羧基化碳纳米管CNT303 50 μm (北京德科岛科技有限公司),羟基化碳纳米管CNT203 10~30 μm (北京德科岛科技有限公司),河豚毒素免疫亲和柱3 mL(江苏美正生物科技有限公司)。织纹螺采自广东湛江。
试剂配制:1%乙酸,1%乙酸甲醇,0.1%氨水,0.1%甲酸-乙腈(1∶1,V/V )。流动相为0.1%甲酸水;TTX标准贮备溶液(100 μg·mL–1)为取TTX冻干标准品1.00 mg,加入0.1%甲酸-乙腈(1∶1,V/V )定容至10 mL,于4 ℃保存;碳纳米管洗脱液为0.1%乙酸-乙腈(8∶2,V/V );标准工作液为取TTX标准储备液,用0.1%甲酸-乙腈(1∶1,V/V )逐级稀释成质量浓度分别为0.5 ng·mL–1、1 ng·mL–1、5 ng·mL–1、10 ng·mL–1和50 ng·mL–1的梯度标准工作液。
1.3 实验条件
1.3.1 色谱条件
色谱柱为Waters ACQUITY UPLC BEH Amide柱(100 mm×2.1 mm,1.7 μm);柱温40 ℃;进样量为5 μL;流速为0.3 mL·min–1;流动相为乙腈(A)和0.1%甲酸水(B)。梯度洗脱条件见表1。
表 1 梯度洗脱条件Table 1. Gradient elution conditiont/min 流动相A/%
mobile phase A流动相B/%
mobile phase B0 95 5 1.0 40 60 3.0 40 60 3.2 95 5 5.0 95 5 1.3.2 质谱条件
电喷雾离子源,正离子扫描;检测方式为多反应监测( MRM );毛细管电压为3.0 kV;离子源温度150 ℃;脱溶剂气温度400 ℃;锥孔气流量为150 L·h–1;脱溶剂气流量为900 L·h–1;TTX的MRM离子对见表2。
表 2 TTX的多反应监测条件Table 2. Conditions of multiple reaction monitoring for tetrodotoxin分析物
analyte母离子/(m/z)
parent ion子离子/(m/z)
daughter ion锥孔电压/V
cone voltage碰撞能量/eV
collision energyTTX 320.1 161.9 35 35 302.0* 35 20 注:*. 定量离子 Note: *. quantitative ion 1.4 样品处理
1.4.1 提取
用自来水清洗织纹螺2~3遍,洗去表面泥沙。用钳子夹碎织纹螺外壳,取织纹螺全部组织充分均质备用。取织纹螺肉匀浆样品2.00 g于50 mL离心管中,加入5 mL 1%乙酸甲醇溶液,在60 ℃水浴中超声15 min,待其冷却至室温,在6 000 r·min–1下离心5 min,转移上清液,在残渣中加入5 mL 1%乙酸甲醇,重复提取1次,合并上清液。取5 mL上清液,在样液中逐滴加入氨水并充分搅拌,调节pH至8.5,过碳纳米管小柱净化。
1.4.2 碳纳米管净化
取250 mg羧基化多壁碳纳米管,湿法装填于空的6 mL小柱中,依次加入5 mL 0.1%乙酸-乙腈(8∶2,V/V )、5 mL 0.1%氨水活化和平衡小柱,挤干柱中残留液体,将5 mL提取液全部上样,挤干柱中液体后用5 mL甲醇、5 mL纯水依次淋洗,挤干小柱,用3 mL 0.1%乙酸-乙腈(8∶2,V/V )洗脱。收集3 mL洗脱液,涡旋后过0.22 μm滤膜,供LC-MS/MS分析。
1.4.3 免疫亲和柱净化
取出冷藏保存的免疫亲和柱,自然恢复至室温,取下柱头,放出柱内保存的PBS溶液。将5 mL提取液全部上样,用10 mL超纯水淋洗,挤干小柱后用5 mL 2%乙酸甲醇溶液洗脱,洗脱液于45 ℃下氮吹至干,残渣用1 mL 0.1%甲酸-乙腈(1∶1,V/V )复溶,过0.22 μm滤膜,供LC-MS/MS分析(参照GB 5009.206—2016)。
2. 结果与分析
2.1 提取条件的优化
织纹螺样品中蛋白质、色素和脂肪等杂质多,TTX不溶于有机试剂,易溶于酸性水溶液或酸性醇溶液,因此需要选取适当的萃取溶剂提取样品中的TTX。分别用1%乙酸水溶液和1%乙酸甲醇溶液提取样品中的TTX,回收率相近,均能有效提取TTX。用乙酸水溶液提取织纹螺肉时会提取出部分蛋白质成分,对仪器检测造成干扰。乙酸甲醇溶液提取能力强,能有效穿透组织,使蛋白质发生沉降,可降低上机检测时蛋白质对仪器产生的累积性干扰效应。在用乙酸甲醇溶液提取样品后,可用二氯甲烷除去脂肪和蛋白质。而吴佳俊等[27]认为用二氯甲烷处理并没有降低基质效应,也不能提高回收率。由于织纹螺样品蛋白质和脂肪含量低,因此本实验在用1%乙酸甲醇提取后,经高速离心吸取上清液过柱,上清液澄清,上样过程速率适当,没有出现堵柱情况。
2.2 碳纳米管的选择
碳纳米管吸附力强,对样品的净化效果好[22-23]。本实验比较了2种碳纳米管对样液中TTX的吸附和净化能力,发现羟基化碳纳米管在上样和淋洗阶段不能很好保留样品中的TTX,均有不同程度的丢失,造成回收率下降;羧基化碳纳米管在上样、淋洗阶段能很好保留TTX,洗脱后其回收率达到85%~90%。在选择装填小柱规格时,若选用3 mL空小柱装填,装填后碳纳米管间会出现空隙使得吸附效果减弱,因此选用6 mL的小柱进行湿法装填。
2.3 上样条件优化
TTX与碳纳米管的结合作用力与样液的pH有关,在酸性或中性条件下结合能力较弱,需要在碱性条件下上样。本实验用氨水调节样液的pH,通过比较不同pH下加标样品的回收率,优化上样液的pH (图1)。上样液pH 为8.5~9.0时,其吸附能力最强,pH≥9.0时TTX不稳定,易分解,回收率下降,因此确定上样液的pH为8.5~9.0。
![]() 图 1 不同上样液的pH对回收率的影响Figure 1. Influence of pH on recovery of tetrodotoxin in sample solution
图 1 不同上样液的pH对回收率的影响Figure 1. Influence of pH on recovery of tetrodotoxin in sample solution2.4 不同小柱吸附净化效果比较
与WCX柱和MCX柱相比,多壁碳纳米管小柱对生物样品中TTX的承载量更大,净化效果更好。之前的研究[28-29]用WCX小柱净化河鲀鱼肉样品时,加标回收率为75%~86%,但实际上样量仅为1 mL,在之后的淋洗过程中,甲醇和氨水均会造成样品中TTX的损失。黄清发等[30]用2%乙酸甲醇提取TTX,样液经过KOH碱解处理,最终上样液只有0.3 mL,之后过MCX柱,用BSTFA衍生,采用气相色谱进行分析。使用MCX柱与WCX柱净化样品时,均存在上样量偏少的问题,而加大实际样品的上样量又会造成上样过程中TTX漏穿,对检测的灵敏度和定量的准确性造成影响。羧基化多壁碳纳米管小柱相较于WCX柱和MCX柱,对样液中的TTX吸附量大、结合能力强,上样时能有效吸附TTX,在淋洗过程中也没有丢失。洗脱时用3 mL 0.1%乙酸-乙腈(8∶2,V/V)即可将TTX完全洗脱,洗脱液过膜后可直接上机检测,减少浓缩步骤,节省检测时间,提高工作效率。
2.5 基质效应的影响
生物样品基质复杂,净化效果不佳时会产生较强的基质抑制效应,造成灵敏度降低和定量不准确。本方法评价了基质效应的影响,在空白提取液中加入一定浓度的TTX标准溶液,比较其与相应浓度标准溶液的色谱峰面积,比值为0.41~0.67。免疫亲和柱特异性强,经过净化处理,能有效除去基质干扰[18]。骆和东等[31]用C18柱净化织纹螺样品,检测结果为0.50 mg·kg–1和0.38 mg·kg–1,小鼠生物法结果为0.73 mg·kg–1和0.58 mg·kg–1;吴佳俊等[27]用乙酸溶液提取河鲀鱼肉中的TTX后上机,对比4根色谱柱的绝对回收率,TSK-GEL Amide 80柱和ACQUITY UPLC BEH Amide柱的回收率达到50%左右,Kinetex HILIC柱与ZIC-HILIC柱的回收率仅为5%左右。上述2种方法的回收率均偏低,这是由样品中的基质抑制效应造成的。而羧基化多壁碳纳米管在碱性条件下能很好地吸附结合样品中的TTX,且承载量高,之后用甲醇和纯水淋洗去除杂质,用LC-MS/MS分析时可减少因基质干扰对检测结果造成的影响。
2.6 方法学研究
2.6.1 线性范围和定量限
用0.1%甲酸-乙腈(1∶1,V/V )将TTX标准工作液逐级稀释成质量浓度分别为0.5 ng·mL–1、1 ng·mL–1、5 ng·mL–1、10 ng·mL–1和50 ng·mL–1的梯度标准工作液。以峰面积为纵坐标,标准溶液浓度为横坐标,绘制工作曲线,线性回归方程为y=58 264.5x–4 145.33 (R2=0.999 7),TTX在0.3~50 ng·mL–1范围内表现良好。采用阴性样品加标方法,以3倍信噪比确定检出限(LOD)为0.3 μg·kg–1,以10倍信噪比确定定量限(LOQ)为1 μg·kg–1,空白样品与加标样品的MRM图谱见图2。
![]() 图 2 空白样品和10 μg·kg–1加标阴性样品经碳纳米管净化后的MRM图谱a. 空白样品;b. 加标样品Figure 2. MRM chromatograms of 10 μg·kg–1 spiked sample cleaned up by carbon nanotubesa. blank sample; b. spiked sample
图 2 空白样品和10 μg·kg–1加标阴性样品经碳纳米管净化后的MRM图谱a. 空白样品;b. 加标样品Figure 2. MRM chromatograms of 10 μg·kg–1 spiked sample cleaned up by carbon nanotubesa. blank sample; b. spiked sample2.6.2 回收率和相对标准偏差
本研究的阴性织纹螺样品均采用GB 5009.206—2016的液相色谱-串联质谱法测定。称取2.00 g阴性织纹螺分别添加水平为1.00 μg·kg–1、10.00 μg·kg–1和100.00 μg·kg–1的TTX标准溶液,每个平行测定5次,其回收率见表3。加标样品在1.00~100.00 μg·kg–1添加水平的平均回收率为83.7%~91.4%,相对标准偏差为2.3%~8.6%。
表 3 样品加标回收率与相对标准偏差Table 3. Recovery of standard addition and relative standard deviation for tetrodotoxinn=5 添加水平/μg·kg–1
added amount回收率/%
recovery相对标准偏差/%
RSD1 91.4 2.3 10 83.7 3.4 100 84.8 8.6 2.7 实际样品测定
本文用碳纳米管固相萃取-超高效液相色谱串联质谱法对采自广东省湛江市的30个织纹螺样品进行检测,TTX毒素的质量分数为1.87~3.34 mg·kg–1,与TTX免疫亲和柱作对比参照结果相近,本方法回收率高,精密度好,适用于螺肉样品TTX的检测。
3. 结论
在TTX样品检测分析工作中,通常采用SPE柱对样品进行上机前处理,但是TTX样品成分复杂,SPE净化效果并不理想[27]。本研究建立了一种高效、灵敏的检测织纹螺中TTX的LC-MS/MS方法。以1%乙酸甲醇溶液作为萃取剂,结合超声水浴法,对织纹螺样品中的TTX进行提取,在碱性条件下样品经过羧基化多壁碳纳米管净化,洗脱液用量少,可直接过滤膜上机检测,结合LC-MS/MS准确定量分析样品中的TTX。本方法步骤简单方便,洗脱液用量少,减少了洗脱液浓缩步骤,大大节省了样品处理和净化时间,在保证灵敏度的同时降低了实验成本,适用于对织纹螺样品中的TTX进行定量分析,是一种安全可靠的检测手段,为织纹螺的TTX检测提供新方法。但碳纳米管应用于TTX检测的研究还不多,许多条件仍需进一步深入探究。
-
![]()
图 1 紫海胆胚胎及幼体发育过程
a. 受精卵;b. 2细胞期;c. 4细胞期;d. 8细胞期;e. 多细胞期;f. 囊胚期;g. 原肠期;h. 棱柱幼体期;i. 二腕幼体;j. 四腕幼体;k. 六腕幼体;l、m. 八腕幼体;n. 稚胆
Figure 1. Embryonic development of A. crassispina
a. Fertilized eggs; b. Two-cell stage; c. Four-cell stage; d. Eight-cell stage; e. Multicellular stage; f. Blastula; g. Gastrula; h. Prism; i. Two-armed larva; j. Four-armed larva; k. Six-armed larva; l and m. Eight-armed larva; n. Juvenile
![]()
图 2 投喂6种不同藻类时浮游幼体的存活状况
Figure 2. Survival rate of A. crassispina fed with six different microalgae diets
![]()
图 3 投喂6种不同藻类时浮游幼体的体长、躯干部骨针长度和口后腕骨针长度
Figure 3. Larval length, body-rod length and post-oral arm length of A. crassispina larvae fed with six different microalgae diets
表 1 紫海胆胚胎和幼体发育过程
Table 1 Embryonic and larval development process of A. crassispina
$\overline {\mathit{\boldsymbol{X}}}{\bf \pm {{SD}}} $ 发育阶段
Developmental stage受精后时间
Time after fertilization体长
Body length/μm图示
Figure受精卵 Fertilized eggs 0 h 83.90±1.5 图1-a 2细胞期 Two-cell stage 1 h 30 min 84.70±1.3 图1-b 4细胞期 Four-cell stage 1 h 45 min 93.40±4.1 图1-c 8细胞期 Eight-cell stage 2 h 93.50±4.4 图1-d 多细胞期 Multicellular stage 3 h 95.20±4.2 图1-e 囊胚期 Blastula 4 h 92.90±0.8 图1-f 原肠期 Gastrula 10 h 103.27±2.3 图1-g 棱柱幼体 Prism 15 h 116.50±11.3 图1-h 二腕幼体 Two-armed larva 17 h 138.50±9.9 图1-i 四腕幼体 Four-armed larva 1 d 7 h 329.70±39.7 图1-j 六腕幼体 Six-armed larva 5 d 514.80±43.1 图1-k 八腕幼体 Eight-armed larva 8 d 761.10±83.2 图1-l 稚海胆 Juvenile 22 d 373.12±12.9 图1-n  下载: 导出CSV
下载: 导出CSV
表 2 不同海胆胚胎和幼体发育过程比较
Table 2 Combined embrryonic and larval development process of different A. crassispina
发育阶段
Developmental stage紫海胆
A. crassispina高腰海胆[22]
M. globulus光棘球海胆[21]
M. nudus中间球海胆[21]
S. intermedius受精卵 Fertilized eggs 0 h 0 h 0 h 0 h 2细胞期 Two-cell stage 1 h 30 min 1 h 1 h 1 h 4细胞期 Four-cell stage 1 h 45 min 1 h 30 min 1 h 30 min 1 h 30 min 8细胞期 Eight-cell stage 2 h 2 h 2 h 2 h 30 min 多细胞期 Multicellular stage 3 h 4 h 30 min 5 h 5 h 囊胚期 Blastula 4 h 8 h 10 h 11 h 原肠期 Gastrula 10 h 15 h 15 h 18 h 四腕幼体 Eight-armed larva 1 d 7 h 2 d 10 h 1 d 2 h 1 d 3 h 六腕幼体 Six-armed larva 5 d 7 d 13 h 8 d 16 h 9 d 18 h 八腕幼体 Eight-armed larva 8 d 9 d 18 h 19 d 1 h 21 d 2 h
下载: 导出CSV
-
[1] 陈锤. 紫海胆的生物学与养殖[J]. 海洋与渔业, 2007(7): 32. [2] 郭文场. 中国海胆种质资源养殖与保护[J]. 特种经济动植物, 2013, 16(9): 9-11. [3] 冯永勤. 紫海胆人工育苗技术研究[J]. 海洋科学, 2006, 30(1): 5-8, 48. doi: 10.3969/j.issn.1000-3096.2006.01.002 [4] 杨章武, 李正良, 郑雅友, 等. 紫海胆人工育苗技术的研究[J]. 台湾海峡, 2001, 20(1): 32-36. [5] 陈颜锋, 游小艇. 紫海胆养殖试验[J]. 科学养鱼, 2003(10): 29. [6] 聂永康, 陈丕茂, 周艳波, 等. 南方紫海胆增殖放流对虾类和蟹类行为的影响[J]. 安徽农业科学, 2016, 44(21): 7-11. doi: 10.3969/j.issn.0517-6611.2016.21.003 [7] CARBONI S, VIGNIER J, CHIANTORE M, et al. Effects of dietary microalgae on growth, survival and fatty acid composition of sea urchin Paracentrotus lividus throughout larval development[J]. Aquaculture, 2011, 324: 250-258.
[8] BRUNDU G, VIAN MONLEÓN L, VALLAINC D, et al. Effects of larval diet and metamorphosis cue on survival and growth of sea urchin post-larvae (Paracentrotus lividus; Lamarck, 1816)[J]. Aquaculture, 2016, 465: 265-271. doi: 10.1016/j.aquaculture.2016.09.014
[9] 聂永康, 陈丕茂, 周艳波, 等. 南方紫海胆摄食习性的初步研究[J]. 南方水产科学, 2016, 12(3): 1-8. doi: 10.3969/j.issn.2095-0780.2016.03.001 [10] 莫宝霖, 秦传新, 陈丕茂, 等. 基于碳、氮稳定同位素技术的大亚湾紫海胆食性分析[J]. 中国水产科学, 2017, 24(3): 566-75. [11] 冯雪, 陈丕茂, 秦传新, 等. 温度和体重对南海野生紫海胆(Anthocidaris crassispina)呼吸代谢的影响[J]. 广东农业科学, 2012, 39(23): 123-5, 31. doi: 10.3969/j.issn.1004-874X.2012.23.040 [12] 孙勉英. 大连紫海胆食性的研究Ⅱ幼海胆期铒料的初步探讨[J]. 水产科学, 1990, 9(4): 31-34. [13] 张鹏, 曾晓起, 尤凯. 盐度及饵料对马粪海胆幼体生长发育及变态的影响[J]. 海洋湖沼通报, 2004(4): 75-80. doi: 10.3969/j.issn.1003-6482.2004.04.013 [14] 常亚青, 丁君, 宋坚, 等. 海参、海胆生物学研究与养殖 [M]. 北京: 海洋出版社, 2004: 245-269. [15] SCHOLTZ R, BOLTON J J, MACEY B M. Effects of different microalgal feeds and their influence on larval development in the white-spined sea urchin Tripneustes gratilla[J]. S Afr J Mar Sci, 2013, 35(1): 25-34. doi: 10.2989/1814232X.2013.769902
[16] BRUNDU G, VALLAINC D, BAROLI M, et al. Effects of on-demand feeding on sea urchin larvae (Paracentrotus lividus; Lamarck, 1816), development, survival and microalgae utilization[J]. Aquacult Res, 2017, 48(4): 1550-1560. doi: 10.1111/are.12990
[17] CASTILLA-GAVILÁN M, BUZIN F, COGNIE B, et al. Optimising microalgae diets in sea urchin Paracentrotus lividus larviculture to promote aquaculture diversification[J]. Aquaculture, 2018, 490: 251-259. doi: 10.1016/j.aquaculture.2018.02.003
[18] QI S, ZHAO X, ZHANG W, et al. The effects of 3 different microalgae species on the growth, metamorphosis and MYP gene expression of two sea urchins, Strongylocentrotus intermedius and S. nudus[J]. Aquaculture, 2018, 492: 123-131. doi: 10.1016/j.aquaculture.2018.02.007
[19] SONNENHOLZNER-VARAS J I, TOURON N, ORRALA M M P. Breeding, larval development, and growth of juveniles of the edible sea urchin Tripneustes depressus: a new target species for aquaculture in Ecuador[J]. Aquaculture, 2018, 496: 134-145. doi: 10.1016/j.aquaculture.2018.07.019
[20] 蒲利云, 吴洪流, 冯永勤, 等. 白棘三列海胆生殖系统的组织学研究[J]. 海南大学学报(自然科学版), 2009, 27(2): 147-153. [21] 常亚青, 王子臣, 宋坚, 等. 四种海胆杂交的可行性及子代的早期发育[J]. 水产学报, 2000, 24(3): 211-216. [22] 王宝锋. 高腰海胆胚胎及幼体发育过程[J]. 大连海洋大学学报, 2019, 34(4): 526-530. [23] 孙勉英. 大连紫海胆浮游幼体期的饵料试验[J]. 水产科学, 1989, 8(2): 23-26. [24] DUY N D Q, PIROZZI I, SOUTHGATE P C. Ingestion and digestion of live microalgae and microalgae concentrates by sandfish, Holothuria scabra, larvae[J]. Aquaculture, 2015, 448: 256-261. doi: 10.1016/j.aquaculture.2015.06.009
[25] 邓正华, 陈明强, 王雨, 等. 合浦珠母贝幼虫培育的饵料优化研究[J]. 水产科学, 2018, 37(6): 721-727. [26] 邓正华, 叶乐, 吴开畅, 等. 黄边糙鸟蛤幼虫对9种单胞藻的摄食与消化研究[J]. 南方水产科学, 2016, 12(6): 91-98. doi: 10.3969/j.issn.2095-0780.2016.06.012 [27] 常亚青, 高绪生. 中间球海胆的人工育苗及增养殖技术(二)[J]. 水产科学, 2004(7): 45-46. doi: 10.3969/j.issn.1003-1111.2004.07.016 [28] CAMERON R, HINEGARDNER R. Initiation of metamorphosis in laboratory cultured sea urchins[J]. Biol Bull, 1974, 146(3): 335-342. doi: 10.2307/1540409
[29] 王笑月. 杂交海胆浮游幼体期的饵料效果研究[J]. 齐鲁渔业, 2005, 22(4): 1-3. [30] 王波, 张春利, 高倩, 等. 饵料对中间球海胆生长发育的影响初探[J]. 黄渤海海洋, 2002, 20(1): 54-59. -
期刊类型引用(19)
1. 王文兵,张钰爽,余倩云,唐文君. 日粮添加复合酶制剂对断奶仔猪生长性能、免疫功能及养分消化率的影响. 中国饲料. 2023(12): 44-47 .  百度学术
百度学术
2. 裘泓杰,何向中,陈乃松,李松林. 低鱼粉饲料中添加外源酶制剂对大口黑鲈生长性能、体组成以及饲料利用率的影响. 上海海洋大学学报. 2023(04): 730-740 . 百度学术
3. 申亮,于志文,韩佳乐,何立彬,郭冉. 复合酶制剂对凡纳滨对虾生长性能、血浆生化指标和免疫性能的影响. 河北渔业. 2022(02): 16-20+46 . 百度学术
4. 谭俊杰,邓岳松,潘奕霖,何托宇. 黄颡鱼配合饲料研究进展. 湖南饲料. 2022(04): 33-38 . 百度学术
5. 杨明容,李达,周美玉. 饲用抗生素替代物在水产养殖中的应用研究进展. 安徽农学通报. 2020(08): 62-66+102 . 百度学术
6. 郭静文,陈山多,冯雨薇,郭子华,吴丹,邢程顺,侯丽萍,舒琥. 饲料中添加海洋红酵母对宝石鲈生长性能的影响. 饲料工业. 2020(10): 44-49 . 百度学术
7. 曲木,张宝龙,暴丽梅,白东清,赵子续,翟胜利. 饲料叶酸和VB_(12)水平对黄颡鱼生长、肉质及抗氧化能力的影响. 饲料工业. 2018(12): 17-23 . 百度学术
8. 张志强,张琼文,孙思琪,王淑荷,王嘉丰,张云龙. 禁食对大鳞副泥鳅形态学及各型白细胞百分比的影响. 安徽农业大学学报. 2018(04): 621-625 . 百度学术
9. 刘龙镇,田相利,王明阳,李海东,李丽,董双林,马家好. 不同复合微生态制剂添加方式对凡纳滨对虾生长、非特异性免疫及抗病力的影响. 中国海洋大学学报(自然科学版). 2018(12): 23-31 . 百度学术
10. 窦勇,姚妙爱. 复合酶制剂对黄颡鱼生长性能、表观消化率及氮排放的影响. 江苏农业科学. 2018(24): 190-193 . 百度学术
11. 张宝龙,曲木,赵国营,王云祥,赵子续,翟胜利. 饲料中添加不同含量的小球藻对黄颡鱼生长及抗氧化能力的影响. 科技资讯. 2018(19): 239-243 . 百度学术
12. 王国霞,王敏,孙育平,胡俊茹,黄文庆,陈晓瑛,邱世殿,黄燕华. 外源脂肪酶对花鲈生长性能、体组成、血清生化指标、消化酶活性及营养物质表观消化率的影响. 动物营养学报. 2017(12): 4542-4553 . 百度学术
13. 陈钱,张滕闲,张宝龙,何建武,张曼,曾小理,白东清. 岩藻黄质对黄颡鱼的生长、消化和抗氧化能力的影响. 饲料工业. 2017(12): 25-31 . 百度学术
14. 王国霞,曹俊明,牛凤池,黄文庆,胡俊茹,陈冰,范怡杰,黄燕华. 外源酶制剂对黄颡鱼免疫抗氧化指标和肠道形态结构的影响. 饲料工业. 2017(16): 17-21 . 百度学术
15. 王国霞,孙育平,牛凤池,何飞,莫文艳,朱喜锋,曹俊明,黄燕华. 外源酶制剂对黄颡鱼稚鱼消化酶活性、消化率及粪便氮磷含量的影响. 广东海洋大学学报. 2017(06): 19-25 . 百度学术
16. 蔡云川,赵书燕,林黑着,黄忠,周传朋,马志洲. 不同蛋白水平下添加抗菌肽对赤点石斑鱼影响的研究. 上海海洋大学学报. 2017(03): 383-391 . 百度学术
17. 田芊芊,徐树德,胡毅,李昭林,赵清海. 不同发酵来源的木聚糖酶对芙蓉鲫幼鱼消化酶及部分血液指标的影响. 中国饲料. 2016(09): 31-33+38 . 百度学术
18. 牛凤池,黄燕华,曹俊明,王国霞,赵红霞,孙育平,李永娟,马永萍. 5种添加剂对黄颡鱼生长性能、体成分及血清生化指标的影响. 动物营养学报. 2015(07): 2176-2183 . 百度学术
19. 卢静,黎中宝,陈强,李文静,黄永春. 复合酶制剂对欧洲鳗鲡(Anguilla anguilla)生长性能、消化酶及非特异性免疫的影响. 海洋与湖沼. 2015(02): 420-425 . 百度学术
其他类型引用(7)




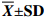

计量
- 文章访问数: 5090
- HTML全文浏览量: 2320
- PDF下载量: 88
- 被引次数: 26
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
