
Effect of different molecular weight chitosan coating on preservation of fish fillets during refrigerated storage
-
摘要: 为探究不同分子量 (50、200和500 kD) 壳聚糖涂膜对冷藏草鱼 (Ctenopharyngodon idella) 鱼片品质保持效果的差异性,比较了不同涂膜保鲜样品贮藏期间的感官评分、微生物计数 [菌落总数和假单胞菌(Pseudomonas)数] 和理化指标 [剪切力、汁液流失率、pH、挥发性盐基氮 (TVB-N)、TCA-溶解肽和生物胺] 的变化情况。结果表明,相较于对照组,3组壳聚糖涂膜处理均能有效抑制鱼肉品质劣化。结合感官评价、TVB-N与菌落总数分析结果,对照组样品冷藏货架期少于6 d,而涂膜处理能使产品货架期延长2 d以上,其中200 kD分子量壳聚糖涂膜鱼片贮藏期间的感官品质保持最佳,货架期延长效果更为明显。通过比较3组涂膜样品的微生物和理化指标,200 kD分子量壳聚糖涂膜鱼片在贮藏第8天时的菌落总数 (TVC)、假单胞菌数、剪切力、汁液流失率、TVB-N和腐胺含量等品质指标均优于另两组涂膜样品,200 kD分子量壳聚糖可作为优选的涂膜基料用于生鲜鱼肉的保鲜。Abstract: In order to explore the differences in the effect of different molecular weight (Mw) chitosan coatings (50, 200 and 500 kD) on the quality of refrigerated grass carp (Ctenopharyngodon idella) fillets, we applied the biological, physical and chemical methods to detect the fresh-keeping quality of the fish fillets, and compared the changes in the sensory scores, microbial enumeration [total viable count (TVC) and Pseudomonas count], and physicochemical indicators [shear force, water loss rate, pH, total volatile base nitrogen (TVB-N), TCA-soluble peptides and biogenic amines] during the storage of different coating samples. The results show that compared with the control group, the coating treatments with all the three Mw chitosan coatings could inhibit the deterioration of fillets quality effectively. Combining with the results of sensory evaluation, TVB-N and TVC, the shelf life of the control group was less than 6 d, while the coating treatment could extend the shelf life more than at least 2 d, and especially the sensory quality of fillets with 200 kD Mw chitosan coating remained the best and had the longest shelf life during the storage. By comparing the microbial and physicochemical indexes of the three coated groups, we found that the total viable count (TVC), Pseudomonas count, shear force, juice loss rate, TVB-N and putrescine content of 200 kD Mw chitosan coating on the 8th day of storage were better than those of the other two groups. Thus, 200 kD Mw chitosan coating could be used as a preferred coating material for the preservation of fresh fish meat.
-
Keywords:
- Coating preservation /
- Chitosan /
- Molecular weight /
- Fillet
-
钙调蛋白(calmodulin,CaM)是广泛存在于真核生物细胞中的、结构和功能高度保守的一种钙离子(Ca2+)结合蛋白,由148个氨基酸组成,含4个EF手结构域,每个EF手能与1个Ca2+结合。4个EF手结构中EF1和EF3、EF2和EF4两两之间相似性最高,具有明显的内部同源性[1]。WATTERSON等[2]和NOJIMA[3]曾提出假说,编码4个EF手结构域的现代CaM基因可能是由编码单个EF手结构域的原始基因经过几轮复制进化而来。钙调蛋白在真核生物细胞中担当胞内钙受体角色,介导Ca2+对环腺苷酸代谢和糖原代谢的调控,在细胞分泌、细胞运动和Ca2+转运方面起重要作用[4]。已克隆得到的哺乳动物和鸟类、蛙等脊椎动物的钙调蛋白的氨基酸序列几乎完全一致,并且低等无脊椎动物居蟹皮海绵(Suberites domuncula)的CaM与脊椎动物CaM之间存在差异的氨基酸位点也仅5个[5-8]。钙调蛋白这种高保守性可以由它参与细胞活动之多、在细胞功能行使中地位之重要来解释。
到目前为止,钙调蛋白是真核细胞所有Ca2+结合蛋白中研究得最为透彻的一种。研究者们在牛(Bos taurus)、小鼠(Mus musculus)、拟南芥(Arabidopsis thaliana)等模式生物中,对CaM参与调控神经发育、心肌收缩、植物免疫等的研究工作已经进行得相当深入[9-11]。在海洋无脊椎动物中关于CaM参与调控机制的研究还不多,但也有一些进展。GAO等[12]研究发现在克氏原螯虾(Procambarus clarkii)蜕皮期间CaM和Ca2+-ATPase存在某种调控关联。在合浦珠母贝(Pinctada fucata)等贝类动物中,CaM被认为在贝类钙代谢中扮演重要角色,参与贝壳的形成[13]。LUAN等[14]的研究表明,CaM可能参与文昌鱼(Branchiostoma belcheri tsingtauense)消化道的分化。CHEN等[15]的研究表明CaM可能参与纹藤壶(Balanus amphitrite)幼体的附着。然而,海参中有关CaM的研究尚未见报道。
棘皮动物与脊索动物同属后口动物,是无脊椎动物中与高等脊椎动物进化上亲缘关系最近的类群,在进化上拥有独特的地位。同时,海参具有的吐脏、内脏再生和自溶等这些独特的生物现象,长期以来倍受研究人员关注。花刺参(Stichopus monotuberculatus)俗称“黄肉参”,隶属于棘皮动物门,海参纲,楯手目,刺参科,在中国台湾、广东、广西以及海南省的海域均有分布[16]。花刺参是中国热带海参中具有代表性的高经济价值种类,不仅是名贵的食用种类,也是重要的食品和医药原料,特别用作香脂、擦剂油、奶油、牙膏和化妆品等的原料[17]。该研究在花刺参中克隆了CaM的cDNA全长序列,并分析其在花刺参不同组织中的表达特点,以期为后续进一步研究CaM在海参生命活动中所起的作用奠定基础。
1. 材料与方法
1.1 试验材料
研究所用花刺参采自广西涠洲岛,在实验室水族箱中暂养一周。水族箱为过滤循环水系统,24 h充气,光周期12 h : 12 h。从水族箱中取出花刺参冰上解剖,体腔液经2 000 g离心15 min,弃上清得到体腔细胞沉淀,加入Trizol后放入-80 ℃保存;体壁、呼吸树和肠组织等经液氮迅速研磨成粉末,加入Trizol后放入-80 ℃保存。
1.3 总RNA提取及cDNA第一链合成
Trizol(invitrogen)法提取海参体壁、呼吸树、肠以及体腔细胞的总RNA,按照TaKaRa cDNA第一链合成试剂盒的说明进行反转录,-20 ℃保存。其中肠样品同时用于CaM基因cDNA全长序列克隆。
1.4 花刺参钙调蛋白(StmCaM)基因的克隆
基于已获得的花刺参EST序列设计引物G-F/G-R,用以扩增CaM基因中间片段。扩增程序为94 ℃ 5 min,预变性;94 ℃ 15 s,52 ℃ 15 s,72 ℃ 1 min,共35个循环;72 ℃ 10 min延伸。PCR产物经回收、连接、转化后挑选阳性克隆送测序。
根据获得的CaM中间片段序列,用Primer 5软件设计3′-RACE引物(F3-1和F3-2)和5′-RACE引物(R5-1和R5-2)。根据3′/5′-Full RACE Kit(Takara)说明书的指导,结合巢式PCR技术扩增CaM基因3′末端和5′末端序列。PCR产物用上面所述的方法进行回收、连接、转化克隆并测序。此研究所用的引物序列见表 1。
表 1 基因克隆与荧光定量PCR使用的引物及其序列Table 1. Primers used in gene cloning and real-time PCR引物 primer 序列 sequence(5′→3′) G-F AGACAGACAGTGAGGAAGA G-R TCATAGTTGACCTGACCATC F3-1 GATGGCAAGGAAGATGAAGGA F3-2 CTTGGAGAAAAATTGACAGATGA R5-1 TGAAACCATTGCCATCTTTG R5-2 AGCAAGTCAGGGTTAAGAGGT qF1 AATGGTTTCATCGGTGCTG qR1 AGGGTTAAGAGGTGGTGGAT Actin-F ACGAAGTTCAAGCCCTGGTTA Actin-R TCTTCTCCATATCATCCCAGTTTG 1.5 序列分析
将测序得到的5′末端序列、中间片段序列和3′末端序列拼接起来,获得花刺参CaM基因的cDNA全长序列。利用NCBI比对所获得的cDNA全长序列,并做同源性分析。应用BioEdit软件寻找ORF并翻译成蛋白质。使用SMART在线软件分析蛋白质结构和功能。使用SWISS-MODEL分析蛋白质的三维结构。用ClustalX及GeneDoc软件做蛋白质序列比对分析。利用Mega 5.0软件根据不同物种的CaM ORF序列以最小进化法构建系统进化树。
1.6 实时荧光定量PCR(real-time PCR)的表达分析
设计荧光定量PCR特异引物qF1和qR1及内参引物Actin F和Actin R。qRT-RCR反应体系包括SYBR Rremix Ex TaqⅡ(12.5 μL)、qF1(1.0 μL)、qR1(1.0 μL)、cDNA模板(2 μL)和灭菌蒸馏水(8.5 μL)。采用2步法扩增标准程序:1)预变性95 ℃ 30 s;2)95 ℃ 5 s,60 ℃ 30 s,40个循环。反应在RG-3000荧光实时定量PCR仪(Applied Biosystems)进行,每种组织cDNA样品及内参分别做3个重复。用RG-3000自带分析软件进行溶解曲线分析以确定单一扩增,用2-ΔCT法分析花刺参CaM基因的组织表达水平[18],ΔCT为目的基因CT值与内参基因CT值的差值,然后用公式2-ΔCT计算相对表达量。
2. 结果
2.1 花刺参CaM cDNA克隆与结构分析
克隆得到的StmCaM cDNA全长1 394 bp(GenBank登录号KJ624993),其中包括138 bp的5′UTR,806 bp的3′-UTR和450 bp的ORF。3′-UTR端聚腺苷酸加尾信号(AAATAG)位于poly A尾上游第21个碱基位点处(图 1)。由ORF推导出的StmCaM含148个氨基酸,预测分子量约16.7 kD,理论等电点为4.1。与其他物种CaM一样花刺参CaM含4个EF手Ca2+结合域(图 1)。利用SWISS-MODEL在线软件分析花刺参CaM的三维结构(图 2),可见CaM含4个螺旋-环-螺旋(helix-loop-helix)结构,即代表 4个EF手。每个EF手包括2个螺旋和连接2个螺旋的环状结构。使用MegAlign软件分析花刺参CaM4个EF手结构域之间的相似度,EF1与EF3相似度为51.7%,EF2与EF4相似度为48.3%;这2个组合间相似度比其他任何两两组合之间,EF1和EF2(31%)、EF1和EF4(37.9%),EF2和EF3(31%)都要高。
![]() 图 1 StmCaM cDNA序列及推导的氨基酸序列下划线为聚腺苷酸加尾信号;*代表终止密码子;加框的序列为4个EF手结构;GenBank序列号KJ624993Figure 1. Nucleotide sequence of Stm CaM cDNA and deduced amino acid sequencesThe polyadenylation signal in 3′-UTR was underlined; the stop codon was marked by an asterisk, and four EF-Hand domains were included in rectangles. GenBank accession No. is KJ624993.
图 1 StmCaM cDNA序列及推导的氨基酸序列下划线为聚腺苷酸加尾信号;*代表终止密码子;加框的序列为4个EF手结构;GenBank序列号KJ624993Figure 1. Nucleotide sequence of Stm CaM cDNA and deduced amino acid sequencesThe polyadenylation signal in 3′-UTR was underlined; the stop codon was marked by an asterisk, and four EF-Hand domains were included in rectangles. GenBank accession No. is KJ624993.2.2 花刺参CaM进化与保守性分析
MegAlign软件对不同物种来源钙调蛋白进行相似度分析的结果,表中显示花刺参CaM与其他物种CaM相似度从95.9%到98%(表 2)。其中作为脊椎动物代表的小鼠的钙调蛋白与花刺参钙调蛋白之间的相似度最低为95.9%。另外,花刺参CaM与真海鞘(Halocynthia roretzi)、白氏文昌鱼(Branchiostoma lanceolatum)、中间球海胆(Strongylocentrotus intermedius)的相似度最高(均为98%);与秀丽隐杆线虫(Caenorhabditis elegans)、绣球海葵(Metridium senile)、居蟹皮海绵(Suberites domuncula)的相似度分别为96.6%、97.3%和96.6%。图 3比对了不同物种来源钙调蛋白之间发生氨基酸替换的全部氨基酸位点,从低等无脊椎海绵到高等脊椎动物小鼠一共有9个位点发生氨基酸替换。表 3具体列举了发生氨基酸替换的位点。花刺参CaM与中间球海胆CaM相比有3个氨基酸位点发生变换,同合浦珠母贝相比也仅有4个氨基酸位点发生变换。
表 2 花刺参CaM和其他物种CaM相似性比对Table 2. Similarity between StmCaM and other CaMs物种
species登录号
Accession No.相似度/%
similarity小鼠(M.musculus) NP_033920.1 95.9 真海鞘(H.roretzi) BAA33967.1 98.0 白氏文昌鱼(B.lanceolatum) P62148 98.0 中间球海胆(S.intermedius) BAB89360.1 98.0 克氏原螯虾(P.clarkii) ACI15835.1 97.3 加利福尼亚海兔(A.californica) NP_001191509 98.0 合浦珠母贝(P.fucata) GI: 46517823 96.6 黑腹果蝇(D.melanogaster) NP_523710 98.0 秀丽隐杆线虫(C.elegans) O16305 96.6 绣球海葵(M.senile) BAB61794.1 97.3 居蟹皮海绵(S.domuncula) CAA77069 96.6 ![]() 图 3 不同物种钙调蛋白氨基酸比对图Figure 3. Alignment of amino acid sequences of CaM from different species表 3 不同物种CaM之间氨基酸差异位点Table 3. Variable residues among different CaMs
图 3 不同物种钙调蛋白氨基酸比对图Figure 3. Alignment of amino acid sequences of CaM from different species表 3 不同物种CaM之间氨基酸差异位点Table 3. Variable residues among different CaMs物种
species位置 posion 27 57 78 99 101 130 143 146 147 小鼠(M.musculus) I A D Y S I Q T A 真海鞘(H.roretzi) I A D F S I T T S 白氏文昌鱼(B.lanceolatum) I A D F S I T T S 中间球海胆(S.intermedius) I A D F S I T T S 花刺参(S.monotuberculatus) T A E F G I T T S 克氏原螯虾(P.clarkii) I A D F S I R T S 加利福尼亚海兔(A.californica) I A D F S I T T S 合浦珠母贝(P.fucata) I A D F S I K M S 黑腹果蝇(D.melanogaster) I A D F S I T T S 秀丽隐杆线虫(C.elegans) I T E F S T G T S 绣球海葵(M.senile) I A D F S I K T S 居蟹皮海绵(S.domuncula) I T E F S T G T S 注:A. 丙氨酸;D. 天冬氨酸;F. 苯丙氨酸;I. 异亮氨酸;K. 赖氨酸;M. 蛋氨酸t;Q. 谷氨酰胺;R. 精氨酸;S. 丝氨酸;T. 苏氨酸
Note:A. Ala;D. Asp;F. Phe;I. Ile;K. Lys;M. Met;Q. Gln;R. Arg;S. Ser;T. Thr利用Mega 4软件,采用最小进化法构建CaM系统发育树。结果显示,花刺参CaM与囊舌虫(Saccoglossus kowalevskii)、绣球海葵、加利福尼亚海兔(Aplysia californica)三者的CaM亲缘关系较近,聚为一簇。另外,不同脊椎动物的CaM聚为一簇,居蟹皮海绵、黑腹果蝇(Drosophila melanogaster)和秀丽隐杆线虫聚为一簇;这2簇则与StmCaM亲缘关系较远(图 4)。
2.3 花刺参CaM基因在各组织中表达的差异
采用荧光定量PCR检测CaM基因在花刺参不同组织中的表达情况。结果显示体壁中CaM相对表达量最低,2-ΔCT值为0.9。呼吸树中最高,2-ΔCT为113.8,其次是体腔细胞和肠(图 5)。
![]() 图 5 荧光定量PCR检测CaM基因在花刺参不同组织中的表达Figure 5. Expression patterns of CaM mRNA in different tissues of S.monotuberculatus by real-time PCR
图 5 荧光定量PCR检测CaM基因在花刺参不同组织中的表达Figure 5. Expression patterns of CaM mRNA in different tissues of S.monotuberculatus by real-time PCR3. 讨论
前人已经克隆并描述了真海鞘、白氏文昌鱼、凡纳滨对虾(Litopenaeus vannamei)、绣球海葵和居蟹皮海绵等不同进化地位的海洋无脊椎动物的钙调蛋白[19-22]。海参作为棘皮动物中代表性的一个类群,其钙调蛋白尚未被研究发表过。该研究克隆了花刺参CaM全长cDNA序列,由其ORF推导出的花刺参CaM与其他物种CaM的相似度从95.9%到98%,同样含有4个EF手结构域。比对花刺参CaM内部4个EF手结构相互之间相似性的结果显示EF1与EF3相似度为51.7%,EF2与EF4相似度为48.3%;表明花刺参CaM中EF1与EF3、EF2与EF4之间内具有很高的内同源性。这为WATTERSON等[2]和NOJIMA[3]曾提出的关于CaM基因进化的假说提供了支撑证据。
研究对CaM在花刺参不同组织中的mRNA表达水平作了定量分析。结果显示,StmCaM mRNA在呼吸树中表达量最高,其次为体腔细胞和肠,而体壁中则几乎检测不到其表达。刘晓云等[23]使用光学显微镜和电子显微镜观察刺参呼吸树超微结构,发现刺参呼吸树由外向内依次为体腔上皮、肌层、血腔、内皮细胞和中央腔。内皮细胞与流经中央腔的海水进行气体交换,同时里面含有丰富的囊泡(吞饮小泡和质膜小泡)可以吸收海水中营养物质。体腔上皮细胞游离面长有许多微绒毛,胞内同样含有丰富的囊泡,能与体腔液进行物质交换。前人对钙调蛋白功能的研究表明,钙调蛋白在细胞分泌、细胞运动和Ca2+中起重要作用。SATIR等[24]在草履虫(Paramecium tetraurelia)、四膜虫(Tetrahymena thermophila)和淡水蚌(Elliptio)的纤毛基部都发现有CaM存在,JAMIESON等[25]也证明钙调蛋白是纤毛轴丝复合物的组成成分;这些研究表明钙调蛋白可能参与纤毛的摆动[24, 26]。LINDEN等[27]研究发现Ca2+/calmodulin复合体能特异的结合到有被囊泡上,参与受体介导的细胞内吞作用。因此,从CaM在花刺参呼吸树中的高表达来推断,CaM可能介导花刺参呼吸树表皮细胞中囊泡吸收营养物质的过程,同时也可能参与微绒毛的摆动和Ca2+吸收。
脊椎动物和无脊椎动物天然免疫细胞中均存在由Toll样受体介导的活性氮(reactive nitrogen species,RNS)抗菌免疫机制[28]。RNS中主要有效成分一氧化氮(NO)由一氧化氮合成酶(Nitric oxide synthase,NOS)合成。紫海胆(Strongylocentrotus purpuratus)的基因组测序发现,棘皮动物中存在NOS基因,并存在多达220个Toll样受体基因,表明这种抗菌免疫机制在棘皮动物中也是存在的[29]。NOS蛋白的结构上有2个CaM结合域,需要与2个Ca2+/CaM结合形成复合体才能合成NO[30]。因此,花刺参肠道和体腔细胞中较高的CaM表达量,可能与其参与活性氮相关的抗菌免疫机制有关。
-
![]()
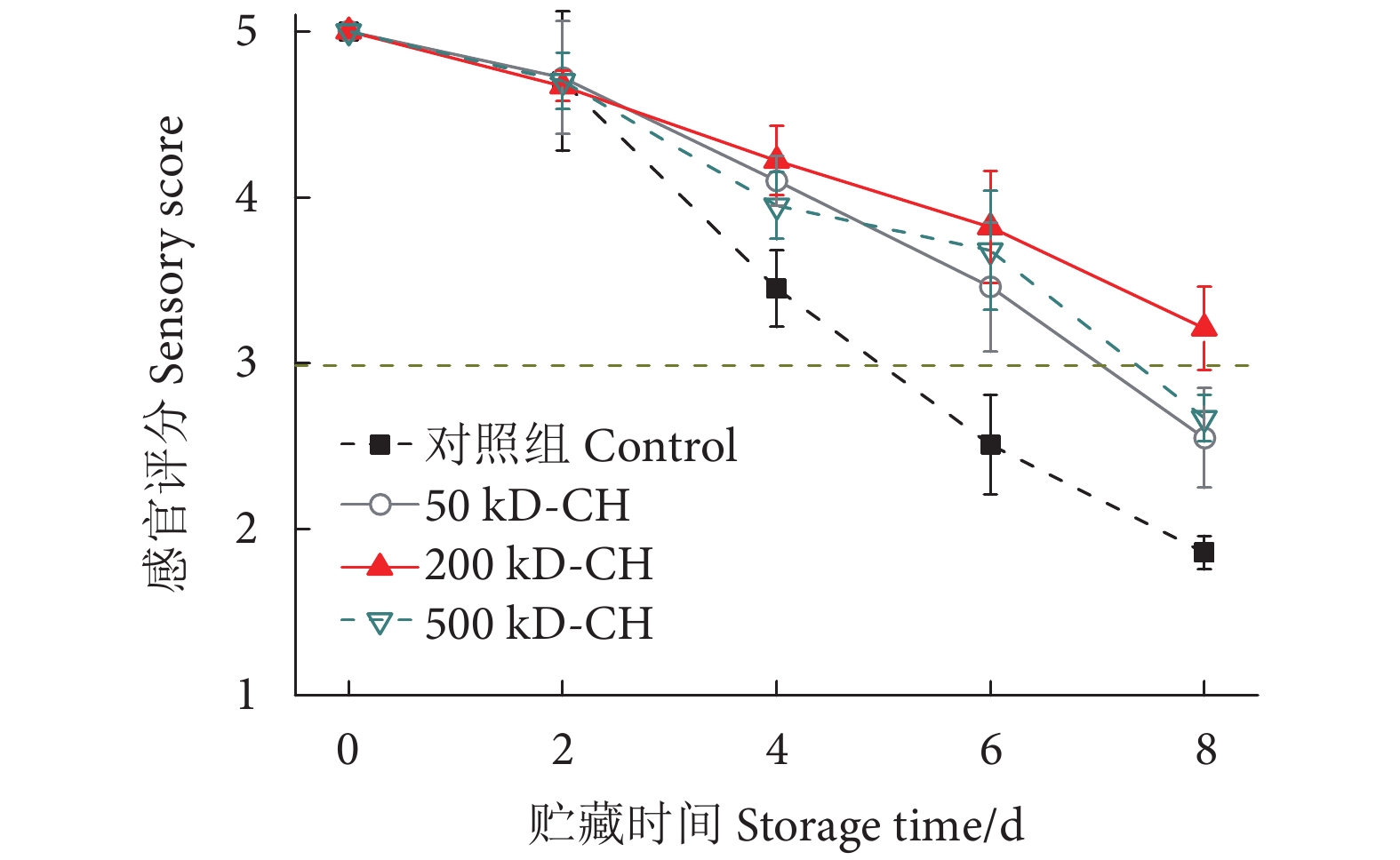
图 1 不同分子量壳聚糖涂膜对冷藏鱼片感官品质的影响
Figure 1. Effect of different molecular weight chitosan coatings on sensory quality of fillets during refrigerated storage
![]()
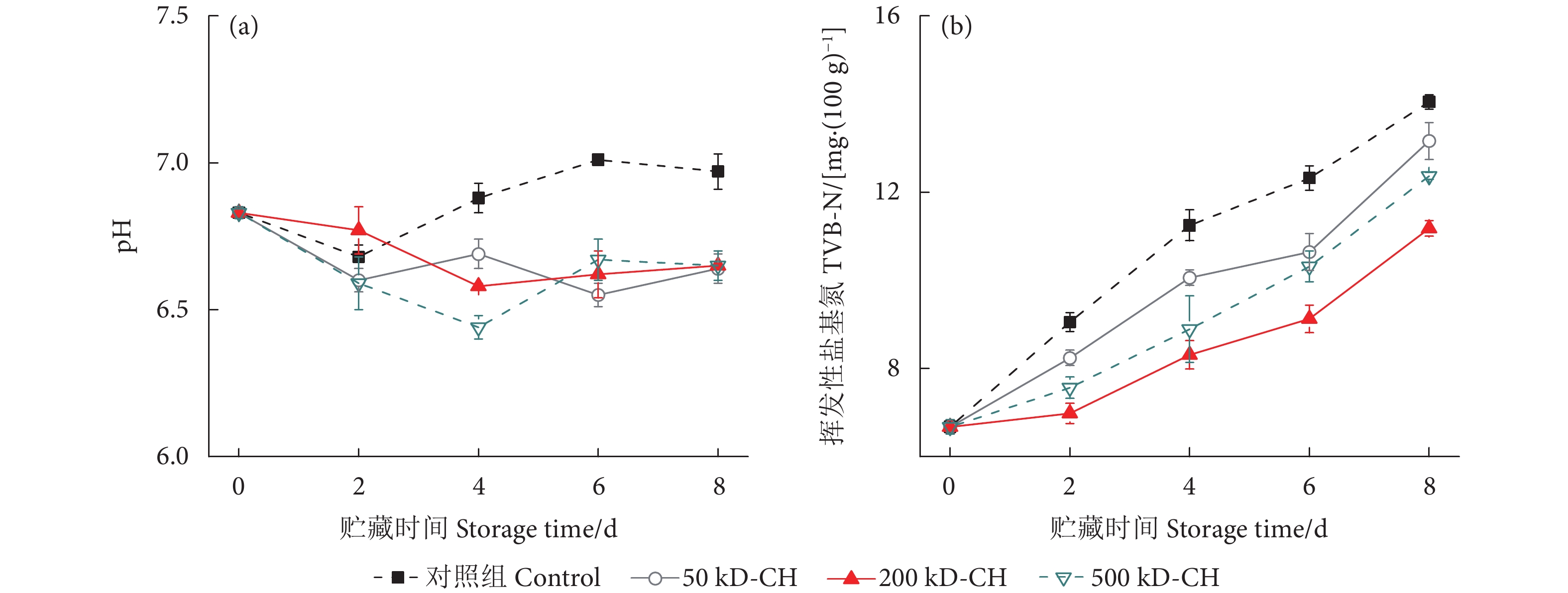
图 2 不同分子量壳聚糖涂膜对冷藏鱼片pH和挥发性盐基氮的影响
Figure 2. Effect of different molecular weight chitosan coatings on pH and TVB-N of fillets during refrigerated storage
![]()
图 3 不同分子量壳聚糖涂膜对冷藏鱼片TCA-溶解肽的影响
Figure 3. Effect of different molecular weight chitosan coatings on TCA-soluble peptides of fillets during refrigerated storage
![]()
图 4 不同分子量壳聚糖涂膜对冷藏鱼片剪切力的影响
Figure 4. Effect of different molecular weight chitosan coatings on shear force of fillets during refrigerated storage
![]()
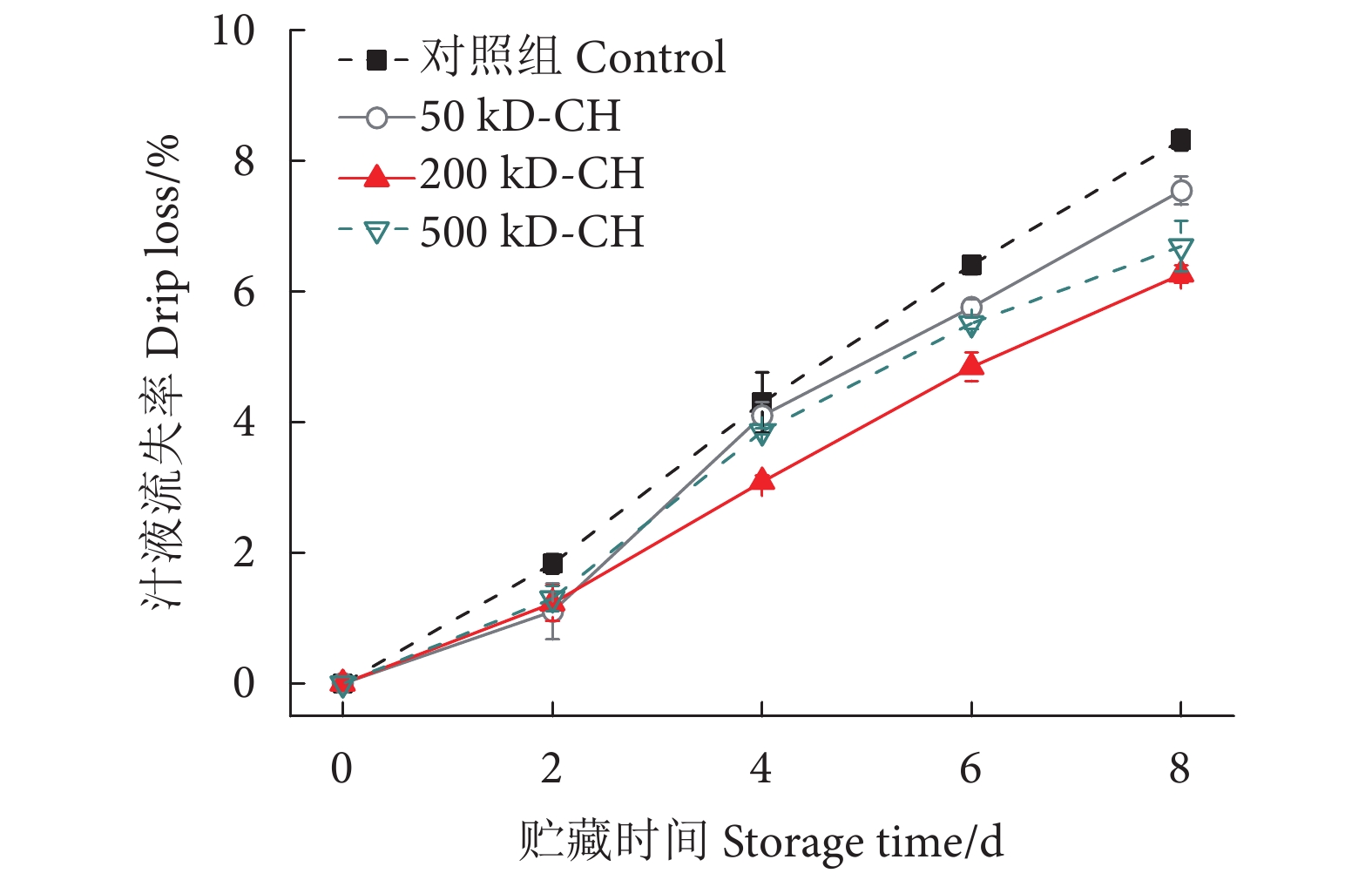
图 5 不同分子量壳聚糖涂膜对冷藏鱼片汁液流失率的影响
Figure 5. Effect of different molecular weight chitosan coatings on drip loss of fillets during refrigerated storage
表 1 感官评分标准
Table 1 Criterion for sensory evaluation
气味 Odor 色泽 Color 质地 Texture 得分 Score 鱼肉新鲜的特有气味 色泽明亮,光泽度好 坚实有弹性,指压立即复原 5 无明显气味 色泽正常,光泽度较好 较有弹性,指压较快复原 4 轻微腥味 光泽稍暗 稍有弹性,指压复原较慢 3 轻微腥臭味 表面部分发黄 弹性不足,指压复原很慢 2 严重的腥臭味 失色,表面大部分发黄 无弹性,指压复原很慢 1  下载: 导出CSV
下载: 导出CSV
表 2 不同分子量壳聚糖涂膜对冷藏鱼片微生物计数的影响
Table 2 Effect of different molecular weight chitosan coatings on microbial enumeration of fillets during refrigerated storage
log10 CFU·g−1 项目
Item组别
Group贮藏时间 Storage time/d 0 2 4 6 8 菌落总数
Total viable count对照组 Control 4.33±0.05Ea 4.98±0.05Da 5.41±0.07Ca 8.39±0.02Ba 9.28±0.13Aa 50 kD-CH 4.33±0.05Da 4.18±0.08Dbc 4.87±0.06Cb 6.97±0.11Bb 8.60±0.21Ab 200 kD-CH 4.33±0.05Da 4.25±0.03Db 4.74±0.18Cb 6.33±0.05Bd 8.18±0.10Ac 500 kD-CH 4.33±0.05Ea 4.04±0.07Dc 4.85±0.12Cb 6.61±0.05Bc 8.44±0.02Abc 假单胞菌数
Pseudomonas count对照组 Control — 3.43±0.03D 3.98±0.07Ca 4.85±0.07Ba 6.23±0.06Aa 50 kD-CH — — 2.91±0.10Cb 3.99±0.13Bb 5.03±0.07Ab 200 kD-CH — — 2.80±0.07Cb 3.51±0.09Bc 4.48±0.05Ad 500 kD-CH — — 2.74±0.08Cb 3.72±0.08Bc 4.80±0.06Ac 注:—. 未检出;同列中不同小写字母间存在显著性差异 (P<0.05);同行中不同大写字母间存在显著性差异 (P<0.05);表3同此。 Note: —. Undetected. Different lowercase letters within the same column or different capital letters within the same line indicate significant difference (P<0.05). The same case in Table 3.
下载: 导出CSV
表 3 不同分子量壳聚糖涂膜对冷藏鱼片生物胺质量分数的影响
Table 3 Effect of different molecular weight chitosan coatings on mass fraction of biogenic amine of fillets during refrigerated storage mg·kg−1
生物胺
Biogenic amines组别
Group贮藏时间 Storage time/d 0 2 4 6 8 腐胺 PUT 对照组 Control 4.77±1.69Ba 4.84±0.18Ba 6.16±0.38Ba 6.94±0.12Ba 20.55±0.74Aa 50 kD-CH 4.77±1.69Ba 4.49±0.22BCab 3.76±0.90Cc 6.60±1.14Ba 11.93±0.55Ab 200 kD-CH 4.77±1.69Ba 3.88±0.36Bb 5.28±1.30ABab 5.69±0.38ABa 7.23±1.16Ac 500 kD-CH 4.77±1.69Ba 2.64±0.23Bc 4.26±0.30Bab 4.80±1.11Ba 14.21±2.11Ab 尸胺 CAD 对照组 Control 0.15±0.08Ca 0.05±0.06Ca 0.26±0.14Ca 5.91±0.26Ba 18.60±1.34Ab 50 kD-CH 0.15±0.08Ca 0.14±0.01Ba 1.75±1.37Ba 1.76±0.64Bb 4.35±0.84Aa 200 kD-CH 0.15±0.08Ca 0.05±0.04Ca 1.21±1.10BCa 2.36±0.82ABbc 3.88±0.08Aa 500 kD-CH 0.15±0.08Ca 1.02±1.18Ba 0.66±0.17Ba 0.68±0.14Bd 3.19±0.93Aa 组胺 HIS 对照组 Control 1.26±0.02Ba 0.96±0.44Ba 1.55±0.02Bb 1.25±0.12Ba 64.20±13.07Aa 50 kD-CH 1.26±0.02Ba 0.94±0.70Aa 2.12±0.22Aa 0.89±0.19Aa 2.06±0.86Ab 200 kD-CH 1.26±0.02Ba 1.12±0.35Ba 1.82±0.01Bab 0.97±0.34Ba 8.08±5.21Ab 500 kD-CH 1.26±0.02Ba 0.91±0.77Ba 1.15±0.07Bc 0.95±0.32Ba 6.46±2.71Ab 酪胺 TYR 对照组 Control 6.82±1.30Ca 5.41±1.43Ca 4.63±1.62Ca 13.19±1.59Ba 58.90±2.66Aa 50 kD-CH 6.82±1.30Ca 5.83±0.29Ca 7.07±1.31Ca 8.28±0.61Bb 29.25±0.21Ac 200 kD-CH 6.82±1.30Ca 5.46±0.33Ba 5.30±1.32Ba 7.64±0.14Bb 41.89±1.14Ab 500 kD-CH 6.82±1.30Ca 5.26±0.36Ba 3.63±1.57Ba 8.11±0.04Bb 20.41±3.40Ad
下载: 导出CSV
-
[1] 农业农村部渔业渔政管理局. 2020中国渔业统计年鉴 [M]. 北京: 中国农业出版社, 2020 : 24. [2] 刘大松. 草鱼肉在微冻和冰藏保鲜中的品质变化及其机理 [D]. 无锡: 江南大学, 2012: 4-7. [3] DEHGHANI S, HOSSEINI S V, REGENSTEIN J M. Edible films and coatings in seafood preservation: a review[J]. Food Chem, 2018, 240: 505-513. doi: 10.1016/j.foodchem.2017.07.034
[4] 李婷婷, 励建荣, 胡文忠. 可食性壳聚糖涂膜保鲜大黄鱼品质控制研究[J]. 中国食品学报, 2013, 13(6): 147-152. [5] 李婷婷, 励建荣, 赵崴. 壳聚糖涂膜对冷藏美国红鱼品质的影响[J]. 食品科学, 2013, 34(10): 299-303. doi: 10.7506/spkx1002-6630-201310066 [6] ZHAO W, YU D, XIA W. Vacuum impregnation of chitosan coating combined with water-soluble polyphenol extracts on sensory, physical state, microbiota composition and quality of refrigerated grass carp slices[J]. Int J Biol Macromol, 2021, 193: 847-855. doi: 10.1016/j.ijbiomac.2021.10.190
[7] LI T, SUN X, CHEN H, et al. Effect of the combination of vanillin and chitosan coating on the microbial diversity and shelf-life of refrigerated turbot (Scophthalmus maximus) filets[J]. Front Microbiol, 2020, 11: 462. doi: 10.3389/fmicb.2020.00462
[8] YU D, REGENSTEIN J M, XIA W. Bio-based edible coatings for the preservation of fishery products: a review[J]. Crit Rev Food Sci Nutr, 2019, 59(15): 2481-2493. doi: 10.1080/10408398.2018.1457623
[9] KONG M, CHEN X G, XING K, et al. Antimicrobial properties of chitosan and mode of action: a state of the art review[J]. Int J Food Microbiol, 2010, 144(1): 51-63. doi: 10.1016/j.ijfoodmicro.2010.09.012
[10] CHANG S H, LIN H T, WU G J, et al. pH Effects on solubility, zeta potential, and correlation between antibacterial activity and molecular weight of chitosan[J]. Carbohydr Polym, 2015, 134: 74-81. doi: 10.1016/j.carbpol.2015.07.072
[11] JONGSRI P, WANGSOMBOONDEE T, ROJSITTHISAK P, et al. Effect of molecular weights of chitosan coating on postharvest quality and physicochemical characteristics of mango fruit[J]. LWT, 2016, 73: 28-36. doi: 10.1016/j.lwt.2016.05.038
[12] DREVINSKAS T, NAUJOKAITYTE G, MARUSKA A, et al. Effect of molecular weight of chitosan on the shelf life and other quality parameters of three different cultivars of Actinidia kolomikta (kiwifruit)[J]. Carbohydr Polym, 2017, 173: 269-275. doi: 10.1016/j.carbpol.2017.06.002
[13] ZHAO X, ZHOU Y, ZHAO L, et al. Vacuum impregnation of fish gelatin combined with grape seed extract inhibits protein oxidation and degradation of chilled tilapia fillets[J]. Food Chem, 2019, 294: 316-325. doi: 10.1016/j.foodchem.2019.05.054
[14] KHAZANDI M, DEO P, FERRO S, et al. Efficacy evaluation of a new water sanitizer for increasing the shelf life of Southern Australian King George whiting and Tasmanian Atlantic salmon fillets[J]. Food Microbiol, 2017, 68: 51-60. doi: 10.1016/j.fm.2017.06.008
[15] LI Y, ZHUANG S, LIU Y, et al. Effect of grape seed extract on quality and microbiota community of container-cultured snakehead (Channa argus) fillets during chilled storage[J]. Food microbiol, 2020, 91: 103492. doi: 10.1016/j.fm.2020.103492
[16] YU D, XIA W, XU Y, et al. The effects of chitosan coating on biogenic amines inhibition and microbial succession of refrigerated grass carp (Ctenopharyngodon idellus) fillets[J]. J Aquat Food Prod T, 2017, 26(10): 1266-1279. doi: 10.1080/10498850.2016.1233473
[17] SUN X, HONG H, JIA S, et al. Effects of phytic acid and lysozyme on microbial composition and quality of grass carp (Ctenopharyngodon idellus) fillets stored at 4 °C[J]. Food Microbiol, 2020, 86: 103313. doi: 10.1016/j.fm.2019.103313
[18] HUANG Z, LIU X, JIA S, et al. The effect of essential oils on microbial composition and quality of grass carp (Ctenopharyngodon idellus) fillets during chilled storage[J]. Int J Food Microbiol, 2017, 266: 52-59.
[19] ICMSF. Micro-organisms in foods: a publication of the International Commission on Microbiological Specifications for Foods (ICMSF) of the international association of microbiological societies [M]. Toronto: University of Toronto Press, 1986: 181-196.
[20] ALISHAHI A, AÏDER M. Applications of chitosan in the seafood industry and aquaculture: a review[J]. Food Bioproc Tech, 2012, 5(3): 817-830. doi: 10.1007/s11947-011-0664-x
[21] HOSSEINNEJAD M, JAFARI S M. Evaluation of different factors affecting antimicrobial properties of chitosan [J]. Int J Biol Macromol 2016, 85: 467-475.
[22] LIU X, HUANG Z, JIA S, et al. The roles of bacteria in the biochemical changes of chill-stored bighead carp (Aristichthys nobilis): proteins degradation, biogenic amines accumulation, volatiles production, and nucleotides catabolism[J]. Food Chem, 2018, 255: 174-181. doi: 10.1016/j.foodchem.2018.02.069
[23] YU D, XU Y, JIANG Q, et al. Freshness assessment of grass carp (Ctenopharyngodon idellus) fillets during storage at 4°C by physicochemical, microbiological and sensorial evaluations[J]. J Food Saf, 2017, 37(2): 1-9.
[24] 余达威. 壳聚糖涂膜对冷藏草鱼片的品质影响研究 [D]. 无锡: 江南大学, 2019: 6. [25] 葛黎红. 内源蛋白酶在低温保鲜草鱼质构劣化中的作用与控制研究 [D]. 无锡: 江南大学, 2017: 44. [26] SINGH A, BENJAKUL S. Proteolysis and its control using protease inhibitors in fish and fish products: a review[J]. Compr Rev Food Sci F, 2018, 17(2): 496-509. doi: 10.1111/1541-4337.12337
[27] WANG L L, YU Q L, HAN L, et al. Study on the effect of reactive oxygen species-mediated oxidative stress on the activation of mitochondrial apoptosis and the tenderness of yak meat[J]. Food Chem, 2018, 244: 394-402. doi: 10.1016/j.foodchem.2017.10.034
[28] LI Y, WU T, WU C, et al. Eugenol-loaded chitosan emulsion holds the texture of chilled hairtail (Trichiurus lepturus) better: mechanism exploration by proteomic analysis[J]. Food Funct, 2020, 11(9): 7509-7522. doi: 10.1039/D0FO01135E
[29] 洪惠. 鳙脂肪酸组成及贮藏过程中品质变化规律与控制技术的研究 [D]. 北京: 中国农业大学, 2015: 90-91. [30] 王航. 草鱼贮藏过程中品质变化规律及特定腐败菌的研究 [D]. 北京: 中国农业大学, 2016: 44-48. -
期刊类型引用(4)
1. 陈淑吟, 张志勇, 吉红九, 李鹏, 赵永超, 张志伟. 黑鲷×真鲷杂交子代与真鲷的Calmodulin基因克隆与表达分析. 海洋渔业. 2018(04): 435-446 .  百度学术
百度学术
2. 邬晓勇, 孙雁霞, 梁勇, 陈月星, 苟小军. 基于CODEHOP的欧李钙调蛋白基因片段的克隆. 烟台大学学报(自然科学与工程版). 2017(04): 292-300 . 百度学术
3. 陈淑吟, 李鹏, 张志勇, 许津, 祝斐, 贾超峰, 王思婷, 任忠宏. 黑鲷♀×真鲷♂反交子代与黑鲷的CaM基因克隆与mRNA表达分析. 中国水产科学. 2017(06): 1193-1202 . 百度学术
4. 李彦, 李北平, 蔡永琴, 岳俊杰, 邱正良. 用高斯网络模型研究钙调蛋白的构象转变. 生物技术通讯. 2016(06): 799-803 . 百度学术
其他类型引用(1)







计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 5
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
