
Food habits study of Mystus guttatus juvenile based on water body analysis and DNA macro barcode technology for stomach contents
-
摘要:
为摸清斑鳠 (Mystus guttatus) 幼鱼的食性和生物学特征,为其资源保护和种群恢复提供科学依据,采用DNA宏条形码技术分析了贵州省罗甸县斑鳠保种场河道中的斑鳠幼鱼胃含物及水体的物种特征。结果显示:1) 在斑鳠幼鱼胃含物中共鉴定出109种生物,分属于15个门,12大类。从门水平看,节肢动物门相对丰度最高,其次是轮虫动物门、绿藻门、脊索动物门。从属水平看,中镖水蚤属 (Sinodiaptomus) 相对丰度最高,其次是臂尾轮虫属 (Brachionus)、鲃属 (Barbus) 和Paralamyctes。2) 在斑鳠幼鱼生存的河道中,共获得193个物种,分属于18个门。从门水平来看,节肢动物门相对丰度最高,其次是绿藻门。从属水平来看,中镖水蚤属的相对丰度最高,其次是衣藻属 (Chlamydomonas),占比较高的还有臂尾轮虫属、筒壳虫属 (Tintinnidium)和单壳缝藻属 (Monoraphidium)。节肢动物门、轮虫动物门和绿藻门在斑鳠幼鱼胃含物中和水体中都是相对丰度较高的类群。在水体中,脊索动物门相对丰度较低 (第15位),而在胃含物中列第4位。研究揭示了斑鳠幼鱼食性选择受水域环境食物的易得性及其喜好性两方面的影响。从能量收支效益的角度来看,斑鳠幼鱼在摄食过程中遵循以最小的能量投入获取最大收益的原则,利于幼鱼存活与生长。
Abstract:To characterize the diet and biology of juvenile Mystus guttatus, and to provide a scientific basis for its resource conservation and population restoration, we applied DNA macro barcode technology to analyze the species characteristics of stomach contents of M. guttatus juvenile and water body in the river channel of M. guttatus conservation farm in Luodian County, Guizhou Province. The results show that: 1) A total of 109 species of organisms were identified in the stomach contents of M. guttatus juvenile, belonging to 15 phyla and 12 major categories. On phylum level, the relative abundance of Arthropods was the highest, followed by Rotifera, Chlorophyta and Chordata. On genus level, the relative abundance of Sinodiaptomus was the highest, followed by Brachionus, Barbus and Paralamyctes. 2) A total of 193 species belonging to 18 phylum were obtained in the river channel where M. guttatus juvenile fish lived. On phylum level, the relative abundance of Arthropods was the highest, followed by Chlorophyta. On genus level, the relative abundance of Sinodiaptomus was the highest, followed by Chlamydomonas. Other genus with higher relative abundance were Brachionus, Tintinnidium and Monoraphidium. Arthropoda, Rotifera and Chlorophyta were relatively abundant groups in the stomach contents of M. guttatus juvenile and water body. The relative abundance of Chordate was low, ranking 15th in the water body, and 4th in the stomach contents. The results indicate that the dietary selection of M. guttatus juvenile is influenced by both the availability of food in water environment and the preference of juvenile. From the perspective of energy balance benefits, the feeding process of M. guttatus juvenile follows the principle of obtaining maximum benefits with minimal energy input, which is beneficial for its survival and growth.
-
Keywords:
- Mystus guttatus /
- Juvenile /
- Stomach contents /
- Feeding habits /
- DNA macro barcode technology
-
幼鱼是水生生态系统中的重要组成部分,了解幼鱼的摄食习性对理解其在生态系统中的功能至关重要。幼鱼的营养供应是决定其生长发育的关键因素[1],研究幼鱼的食物种类与组成,有助于了解其营养状况和稳定的食物偏好,并为其营养水平提供初步评估[2]。幼鱼的食性复杂、食物来源广泛,会根据营养需求、口裂大小等选择适合的饵料,并倾向于选择外形较大的饵料,从而减少消耗、获取更多的物质和能量,以提高单位时间的收益[3]。
学者们对海洋区域的幼鱼食性作了大量研究。波罗的海比目鱼 (Platichthys flesus) 幼鱼 [(37.0±18.6) mm] 主要以摇蚊类、桡足类和寡毛类为食[4];北海道南部沿海地区亚洲油鲽 (Microstomus achne) 幼鱼 (30.0~39.9 mm) 主要捕食小型甲壳类动物[5];日本霞光湖近海防波堤地区斑点叉尾鮰 (Ictalurus punctatus) 幼鱼 (20~56 mm) 的主要食物包括摇蚊幼虫和枝角类[6];金鼻眶灯鱼 (Diaphus chrysorhynchus) 幼鱼 (45~74 mm) 以浮游甲壳动物 (桡足类、磷虾类等) 为主要摄食对象[7]。对江河区域的幼鱼食性也有很多研究,例如,生活在松花江的花䱻 (Hemibarbus maculatus) 幼鱼 (94~141 mm) 主要摄食水生昆虫、底栖生物、浮游植物[8];黑河白甲鱼 (Onychostoma macrolepis) 幼鱼 (78~182 mm) 饵料生物来源主要是水生昆虫成虫、藻类及摇蚊幼虫[9];金沙河三级支流孙水河硬刺松潘裸鲤 (Gymnocypris potanini firmispinatus) 幼鱼 (94~103 mm) 的饵料生物分为着生藻类、浮游藻类、浮游动物、底栖动物和水生高等植物5大类[10]。
胃含物形态鉴定法在鱼类食性鉴定中的应用最早,也是目前应用最广泛的传统鉴定方法。但由于形态学鉴定的局限性,只能检测到食物大类,而幼鱼个体较小,摄食食物量甚微,很难通过胃内容物的视觉评估来识别高度消化的食物[11]。随着分子生物学技术的快速发展,DNA测序技术在短短十年内由传统的一代Sanger测序已快速发展到二代高通量测序及三代单分子测序,基于DNA的动物食性分析技术也从传统的Sanger测序拓展到高通量测序,使得基于高通量测序的DNA宏条形码技术被广泛用于动物食性分析的研究中[12]。目前,DNA宏条形码技术已应用于盲洞鳗 (Ophisternon candidum)[13]、横带刺尾鱼 (Acanthurus triostegus)[14]、沙丁鱼 (Sardina pilchardus)[15]和亮眶灯鱼 (Diaphus splendidus)[16]等鱼类和一些幼鱼的食性研究中[17]。
斑鳠 (Mystus guttatus) 隶属于鲇形目、鲿科、鳠属,俗称芝麻剑、魽鱼、白须魽,主要分布于珠江水系[18],在贵州省所属珠江水系的河流中均有分布。因其全身无鳞、肉质细嫩、味道鲜美、营养价值高,被誉为珠江“四大名鱼”之一,享有“淡水鱼王”的美誉[19]。由于斑鳠资源量日渐减少,2021年2月,斑鳠 (野生种群) 被列入《国家重点保护野生动物名录》二级野生保护动物,具有重要的保护和利用价值。目前对斑鳠食性的研究较少,仅在珠江水系的西江干流和珠江三角洲河网开展过基于胃内容物分析法的斑鳠食性研究[20]。本研究以贵州省罗甸县斑鳠保种场河道内的斑鳠幼鱼为材料,利用DNA宏条形码技术研究其胃含物及河道水体中的饵料生物的物种组成,旨在进一步掌握斑鳠幼鱼的食性,深入了解其生物学特征,为斑鳠的资源保护及种群恢复提供科学依据。
1. 材料与方法
1.1 样品采集
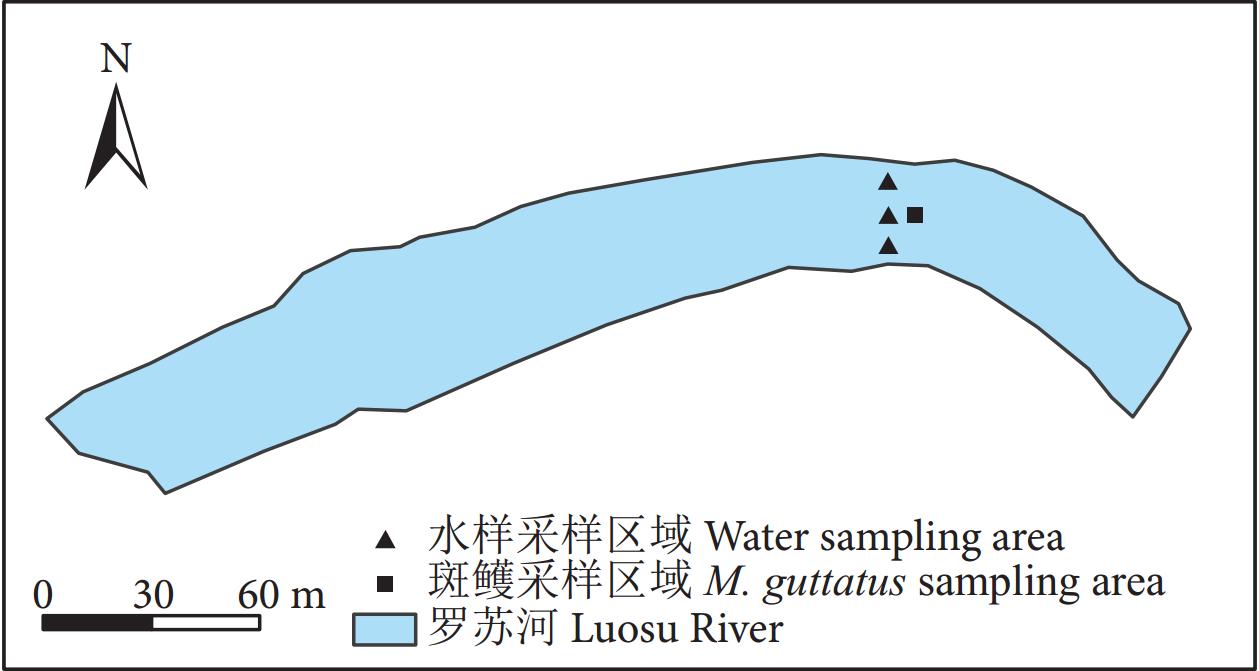
于2023年7月18日在贵州省罗甸县罗悃镇平榜村的斑鳠保种场内的河道 (106°33'36''E—106°33'38''E, 25°14'10''N—25°14'12''N) 进行采样。河中间水深6.5 m,靠近河岸水深4 m,河道长1 400 m,宽30~32 m。
在斑鳠保种场河道的中间使用抬网采集斑鳠幼鱼,随机选取9尾测量体长及体质量 [平均体质量 (30.97±3.04) g,平均体长 (13.97±0.62) cm],活体运输至贵州大学动物科学学院实验室。同时选取3个水样采样点:用抬网捕获斑鳠的位置、抬网左侧距河岸2 m处、抬网右侧距河岸2 m处 (图1)。每个采样点使用容量为1 L的采水器分别采集上层 (离水面0.5 m处)、中层 (离水面3 m处) 和下层 (离泥面0.5 m处) 水样,3层水样混合后保存至已消毒的5 L塑料桶中作为1个重复水样,每个采样点采集3个重复。5 L塑料桶在使用前均用10% (w) 的次氯酸钠溶液进行消毒,具体方法为:将塑料桶置于塑料收纳箱中,向收纳箱中倒入次氯酸钠溶液直至淹没塑料桶,浸泡30 min后,戴上塑胶手套将其取出,并用纯净水反复冲洗干净。
1.2 样品处理
用75% (φ) 的乙醇冲洗斑鳠鱼体表面,然后用无菌剪刀和镊子将胃从腹腔中分离出来,用PBS缓冲液冲洗,将取出的胃含物置于50 mL无菌离心管中。每3尾斑鳠的胃含物置于同一离心管中并混合均匀后作为1份样品,−80 ℃冰箱保存,共3份样品。
河道水样采集后,在24 h内使用0.45 μmol·L−1混合纤维素滤膜 (Whatman,英国) 进行真空抽滤,将每个采样点的3个重复水样抽滤后放入同一无菌离心管中作为1份样品,3个采样点共3份样品。结束后立即用液氮速冻并置于 −80 ℃ 冰箱保存。
1.3 DNA提取、PCR扩增及高通量测序
按照E.Z.N.ATM Mag-Bind Soil DNA Kit试剂盒 (OMEGA,美国) 说明书提取斑鳠胃含物及水样的DNA。以提取的DNA为模板,使用通用引物18S V4F (5'-GGCAAGTCTGGTGCCAG-3')和18S V4R (5'-ACGGTATCTRATCRTCTTCG-3')[21]来扩增真核生物18S rRNA基因V4区。PCR产物经2% (w) 琼脂糖凝胶电泳纯化后,选择主带大小为400~450 bp的序列,使用QIAquick胶回收试剂盒割胶回收目标条带。PCR扩增产物使用Tru SeqDNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建,构建好的文库经过Qubit和Q-PCR定量,经检测合格后,将合格的文库在Illumina Msieq PE300测序平台 (Illumina,美国) 进行高通量测序 (上海生工生物工程股份有限公司)。
1.4 数据处理与分析
通过Illumina Miseq PE300测序平台双端测序得到下机数据,去除引物接头序列,根据overlap关系进行拼接,区分样本后对序列质量进行质控过滤,得到各样本的有效序列。应用Usearch (11.0.667) 软件对有效序列按照97%的序列相似度进行聚类得到OTU,选择OTU代表序列使用GenBank数据库比对进行物种注释,并人工去除非饵料信息 (主要是一些真菌类生物)、斑鳠自身序列、无法鉴定且数量较少的序列。若满足序列相似度 (per.identity) ≥97%,则认为鉴定到属水平,不满足条件的序列则被归为未分类。利用SPSS 26.0软件对注释到的饵料进行统计分析。
2. 结果
2.1 测序特征
斑鳠胃含物与水样测序结果中的有效序列数及OTU数统计见表1。经测序,河道斑鳠胃含物和水样分别获得46 116和84 413条原始序列,数据经拼接和质量过滤后,分别获得37 169和70 343条有效序列。
表 1 有效序列数及OTU数统计Table 1 Statistics of number of effective sequences and OTU样品
Sample原始序列数
Number of original sequences有效序列数
Number of valid sequences有效序列平均长度
Average length of effective sequence/bp最短序列长度
Minimum sequence length/bp最长序列长度
Longest sequence length/bpOTU总数
Total number of OTU斑鳠幼鱼胃含物
Stomach contents of juvenile M. guttatus46 116 37 169 438 310 474 239 河道水环境样品
Sample of river channel water84 413 70 343 416 303 475 441 斑鳠幼鱼胃含物及河道水样有效序列注释后共聚为466个OTUs,其中25个为斑鳠特有,227个为水样特有,214 个为斑鳠和水样共有。
2.2 斑鳠幼鱼胃含物组成
2.2.1 斑鳠幼鱼胃含物饵料生物种类
去除斑鳠自身序列、无法鉴定且数量较少的序列后,斑鳠胃含物共获得109个种属,分属15个门 (表2)。序列最丰富的是绿藻门 (31种),其次是纤毛虫门 (22种),节肢动物门和轮虫动物门各11种,丝足虫门、硅藻门、线虫动物门、脊索动物门和链型植物门也较丰富,分别为8、7、6、3和3种。
表 2 斑鳠幼鱼胃含物饵料生物种类Table 2 Types of stomach content of M. guttatus juvenile门
Phylum属水平种类数
Number of species on genus level绿藻门 Chlorophyta 31 纤毛虫门 Ciliophora 22 轮虫动物门 Rotifera 11 节肢动物门 Arthropoda 11 丝足虫门 Cercozoa 8 硅藻门 Bacillariophyta 7 线虫动物门 Nematoda 6 脊索动物门 Chordata 3 链型植物门 Streptophyta 3 Endomyxa 2 腹毛动物门 Gastrotricha 1 Haptista 1 Imbricatea 1 扁形动物门 Platyhelminthes 1 环节动物门 Annelida 1 总计Total 109 2.2.2 斑鳠幼鱼胃含物门水平相对丰度
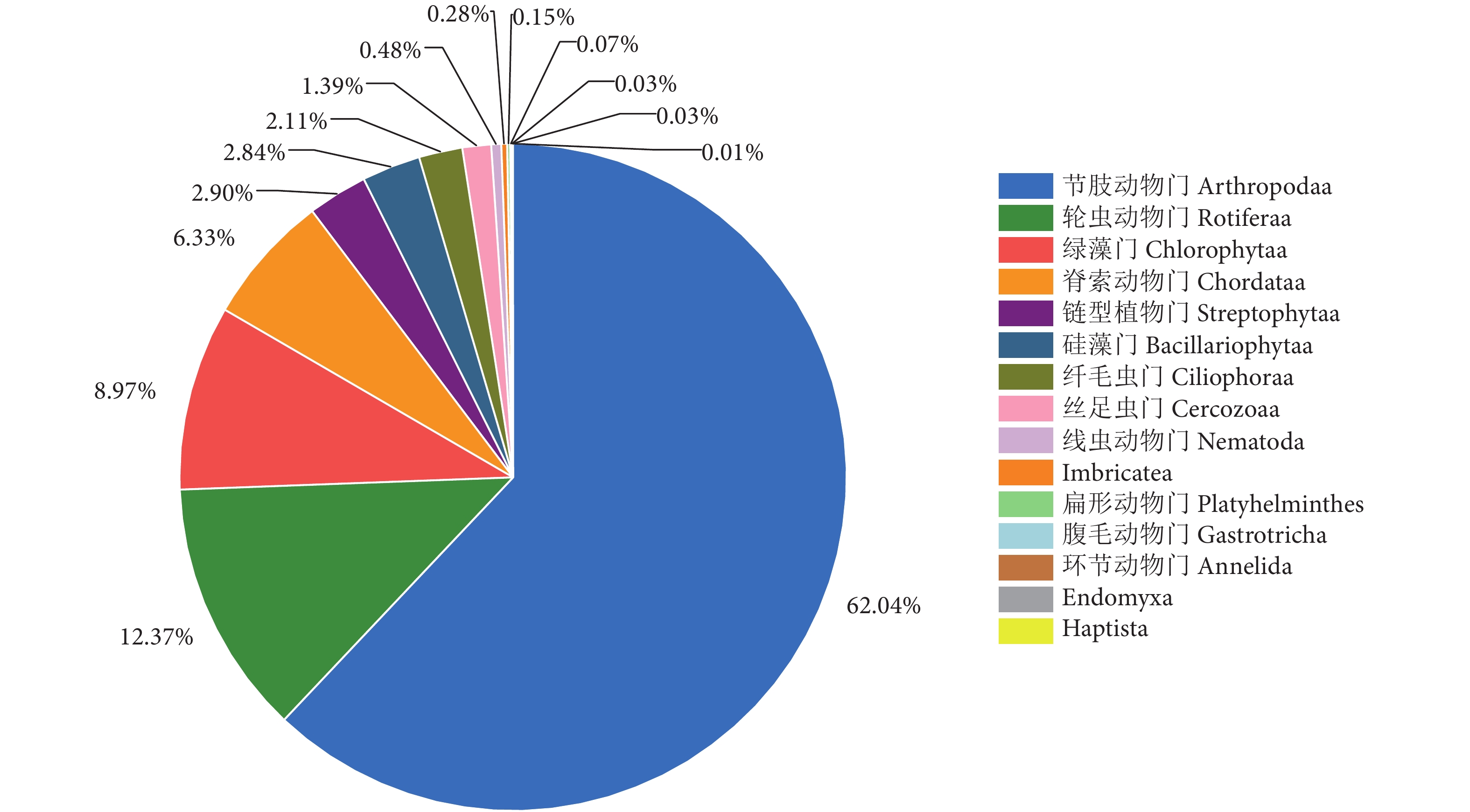
斑鳠幼鱼胃含物门水平相对丰度见图2。节肢动物门的相对丰度最高,占食物序列总数的62.04%;其次为轮虫动物门、绿藻门、脊索动物门,分别占12.37%、8.97%、6.33%;链型植物门、硅藻门和纤毛虫门的占比也较高,分别为2.90%、2.84%和2.11%。
![]() 图 2 斑鳠幼鱼胃含物门水平相对丰度Fig. 2 Relative abundance of stomach contents of M. guttatus juvenile fish on phylum level
图 2 斑鳠幼鱼胃含物门水平相对丰度Fig. 2 Relative abundance of stomach contents of M. guttatus juvenile fish on phylum level2.2.3 斑鳠幼鱼胃含物属水平相对丰度
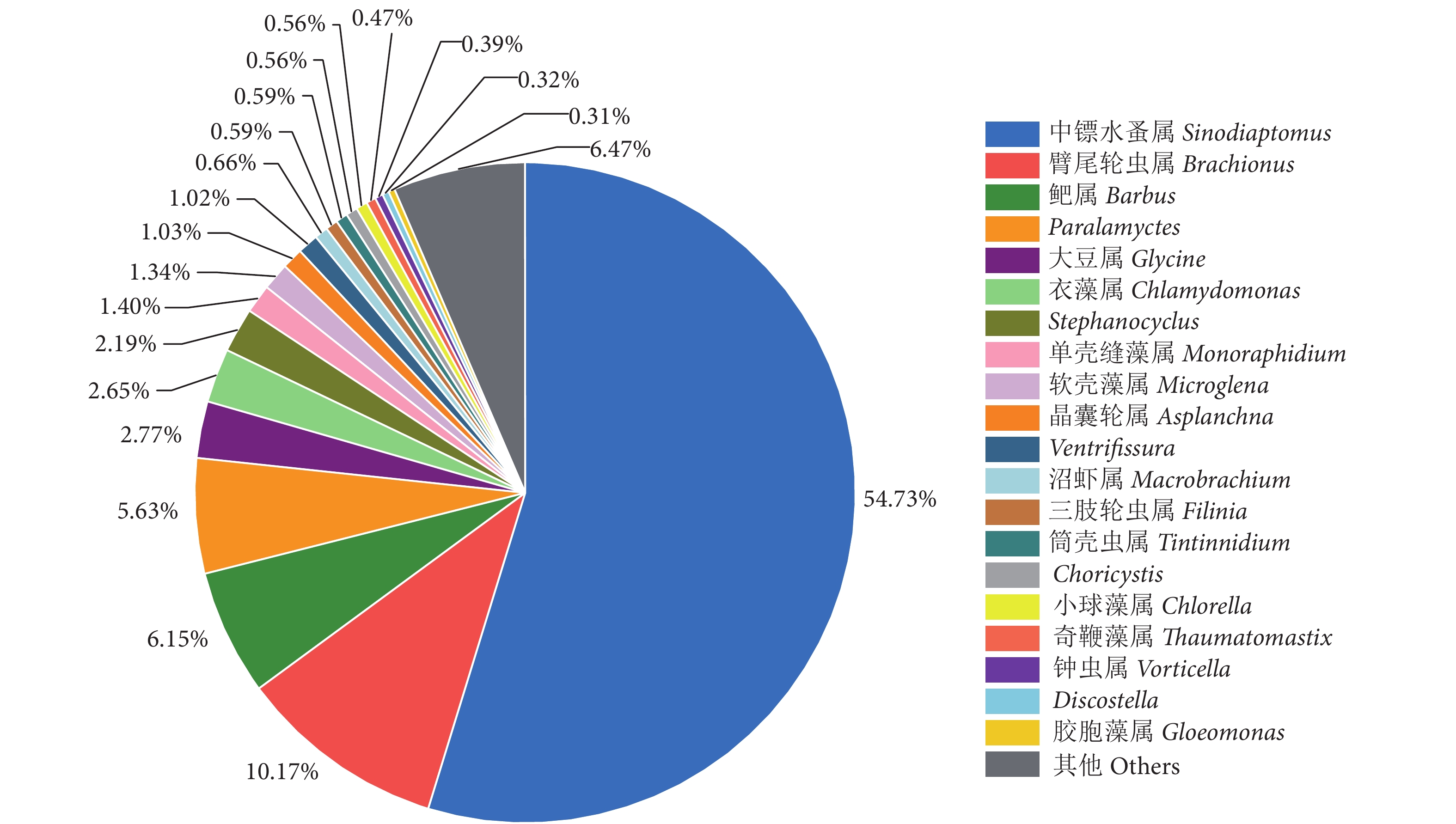
斑鳠幼鱼胃含物属水平相对丰度见图3。中镖水蚤属 (Sinodiaptomus) 占食物序列的相对丰度最高 (54.73%),其次为臂尾轮虫属 (Brachionus)、鲃属 (Barbus) 和Paralamyctes,分别为10.17%、6.15%和5.63%。
![]() 图 3 斑鳠幼鱼胃含物属水平相对丰度Fig. 3 Relative abundance of stomach contents of M. guttatus juvenile fish on genus level
图 3 斑鳠幼鱼胃含物属水平相对丰度Fig. 3 Relative abundance of stomach contents of M. guttatus juvenile fish on genus level2.2.4 斑鳠幼鱼胃含物饵料生物大类分析
斑鳠幼鱼胃含物饵料生物见表3,共分为12大类,包括甲壳类、轮虫、藻类、鱼类、多足类、原生动物、水生植物、线虫、扁形动物、腹毛动物、昆虫类和环节动物。其中甲壳类相对丰度最高 (56.38%),其次是轮虫 (12.37%) 和藻类 (11.83%),相对丰度大于1%的还有鱼类、多足类、原生动物和水生植物,分别为6.33%、5.64%、4.00%和2.90%。
表 3 斑鳠幼鱼胃含物饵料生物表Table 3 Biota of bait in stomach contents of M. guttatus juvenile饵料类别 (数量百分比、种类)
Bait category
(Percentage, Category)门水平
Phylum level纲水平
Class level属水平
Genus level属水平相对丰度
Relative abundance on genus level/%甲壳类 Crustacean
(56.38%, 9)节肢动物门 甲壳纲 明镖水蚤属 Heliodiaptomus 0.01 中剑水蚤属 Mesocyclops 0.03 新鳋属 Neoergasilus 0.28 美丽猛水蚤属 Nitokra 0.10 中镖水蚤属 Sinodiaptomus 54.85 温剑水蚤属 Thermocyclops 0.21 日猛水蚤属 Tisbe 0.22 软甲纲 沼虾属 Macrobrachium 0.66 对虾属 Penaeus 0.02 轮虫 Rotifer
(12.37%, 11)轮虫动物门 真轮纲 Adineta 0.03 晶囊轮属 Asplanchna 1.04 臂尾轮虫属 Brachionus 10.19 Collotheca 0.01 狭甲轮虫属 Colurella 0.21 聚花轮虫属 Conochilus 0.13 三肢轮虫属 Filinia 0.59 巨腕轮属 Hexarthra 0.04 旋轮虫属 Philodina 0.04 多肢轮虫属 Polyarthra 0.02 异尾轮属 Trichocerca 0.07 藻类 Algae
(11.83%, 39)Haptista Centroplasthelida Choanocystis 0.01 硅藻门 硅藻纲 菱形藻属 Nitzschia 0.03 圆筛藻纲 Acanthoceras 0.02 直链藻属 Aulacoseira 0.01 碟星藻属 Discostella 0.32 Stephanocyclus 2.20 尾管藻属 Urosolenia 0.25 间藻纲 无管眼藻属 Extubocellulus 0.01 绿藻门 绿藻纲 负蝗属 Atractomorpha 0.06 荀球藻属 Bracteacoccus 0.01 四鞭藻属 Carteria 0.22 衣藻属 Chlamydomonas 2.66 绿梭藻属 Chlorogonium 0.04 空星藻属 Coelastrum 0.27 Dangeardinia 0.04 迪森藻属 Deasonia 0.07 链带藻属 Desmodesmus 0.14 异形藻属 Dysmorphococcus 0.15 胶胞藻属 Gloeomonas 0.32 Gungnir 0.08 Hamakko 0.04 软壳藻属 Microglena 1.34 单针藻属 Monoraphidium 1.40 麦可属 Mychonastes 0.03 Neochlorosarcina 0.14 润衣藻属 Oogamochlamys 0.01 介壳藻属 Phacotus 0.11 杂球藻属 Pleodorina 0.17 Rotundella 0.01 四角藻属 Tetraedron 0.04 维衣藻属 Vitreochlamys 0.01 平藻纲 单鞭藻属 Pedinomonas 0.01 共球藻纲 小球藻属 Chlorella 0.56 索囊藻属 Choricystis 0.56 Jaagichlorella 0.03 Myrmecia 0.22 卵囊藻属 Oocystis 0.11 微绿藻属 Picochlorum 0.07 石莼纲 石莼属 Ulva 0.06 鱼类 Fish
(6.33%, 3)脊索动物门 鱼纲 鲃属 Barbus 6.16 Eutropiichthys 0.10 翠鳞鱼属 Herklotsichthys 0.07 多足类Myriapods
(5.64%, 1)节肢动物门 唇足纲 Paralamyctes 5.64 原生动物Protozoa
(4.00%, 33)Endomyxa norank_Endomyxa 浮霉菌属 Planctomyxa 0.01 Thalassomyxa 0.02 Imbricatea norank_Imbricatea 奇鞭藻属 Thaumatomastix 0.48 丝足虫门 norank_Cercozoa Cavernomonas 0.01 无吻虫属 Clautriavia 0.12 Massisteria 0.01 Neocercomonas 0.11 Paracercomonas 0.01 Tremula 0.06 Thecofilosea Trachyrhizium 0.04 Ventrifissura 1.02 纤毛虫门 Armophorea Brachonella 0.01 异毛纲 Linostomella 0.04 核残迹纲 Loxodes 0.03 叶口纲 Balantidion 0.02 串须茶属 Cyclotrichium 0.10 Lacrymaria 0.04 寡膜纲 草履虫属 Paramecium 0.08 伪钟虫属 Pseudovorticella 0.04 钟虫属 Vorticella 0.39 叶咽纲 吸管虫属 Acineta 0.03 壳吸管虫属 Paracineta 0.10 前口纲 Balanion 0.07 Hexasterias 0.03 裸口虫属 Holophrya 0.01 Levicoleps 0.03 尾毛虫属 Urotricha 0.04 旋毛纲 异列虫属 Anteholosticha 0.04 纤毛属 Hypotrichidium 0.04 Limnostrombidium 0.09 侠盗虫属 Strobilidium 0.07 筒壳虫属 Tintinnidium 0.59 拟铃虫属 Tintinnopsis 0.22 水生植物 Hydrophyte
(2.90%, 3)链型植物门 木兰纲 Camellia 0.07 大豆属 Glycine 2.77 双星藻纲 新月藻属 Closterium 0.06 线虫Nematode
(0.27%, 6)线虫动物门 色矛纲 滑刃线虫属 Aphelenchoides 0.01 Chromadorina 0.01 异小杆属线虫 Heterorhabditis 0.10 单宫线虫属 Monhystera 0.01 Pellioditis 0.01 Enoplea Viscosia 0.13 扁形动物Platyhelminth
(0.15%, 1)扁形动物门 被杆体纲 Phaenocora 0.15 腹毛动物Gastrotricha
(0.07%, 1)腹毛动物门 norank_Gastrotricha 鼬虫属 Chaetonotus 0.07 昆虫类Hexapeopoda
(0.03%, 1)节肢动物门 昆虫纲 大白蚁属 Macrotermes 0.03 环节动物 Annelid
(0.03%, 1)环节动物门 环带纲 瓢体虫属 Aeolosoma 0.03 2.3 河道水环境生物组成
2.3.1 河道水环境生物种类
河道水环境生物种类见表4。在除去无法鉴定且数量较少的序列后,共获得193种属,分属18个门。生物种类最多的是绿藻门 (52种),其次是纤毛虫门 (45种),种类大于10的还有硅藻门、轮虫动物门、节肢动物门和线虫动物门,分别为15、14、13和10种。
表 4 水样生物种类Table 4 Biological species of water sample门
Phylum属水平种类数
Number of species on genus level绿藻门 Chlorophyta 52 纤毛虫门 Ciliophora 45 硅藻门 Bacillariophyta 15 轮虫动物门 Rotifera 14 节肢动物门 Arthropoda 13 线虫动物门 Nematoda 10 丝足虫门 Cercozoa 9 Endomyxa 6 Tubulinea 6 Haptista 5 腹毛动物门 Gastrotricha 4 链型植物门 Streptophyta 3 环节动物门 Annelida 2 顶复动物亚门 Apicomplexa 2 脊索动物门 Chordata 2 Imbricatea 2 扁形动物门 Platyhelminthes 2 Microsporidia 1 总计 Total 193 2.3.2 河道水样门水平相对丰度
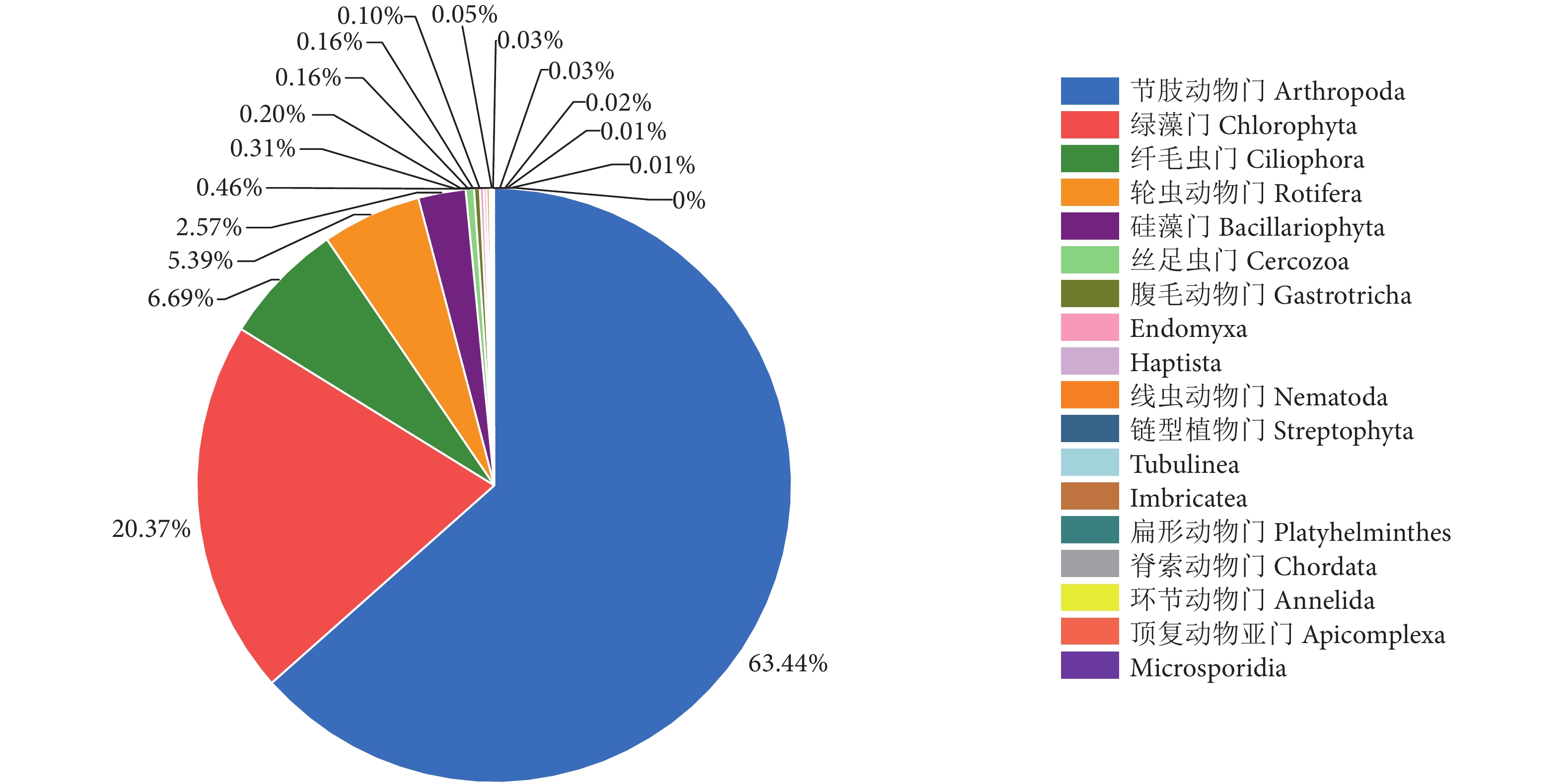
河道水样门水平相对丰度见图4。节肢动物门相对丰度最高,占食物序列总数的63.44%;其次是绿藻门,占20.37%;相对丰度较高的还有纤毛虫门、轮虫动物门和硅藻门,分别占6.69%、5.39%和2.57%。
2.3.3 河道水样属水平相对丰度
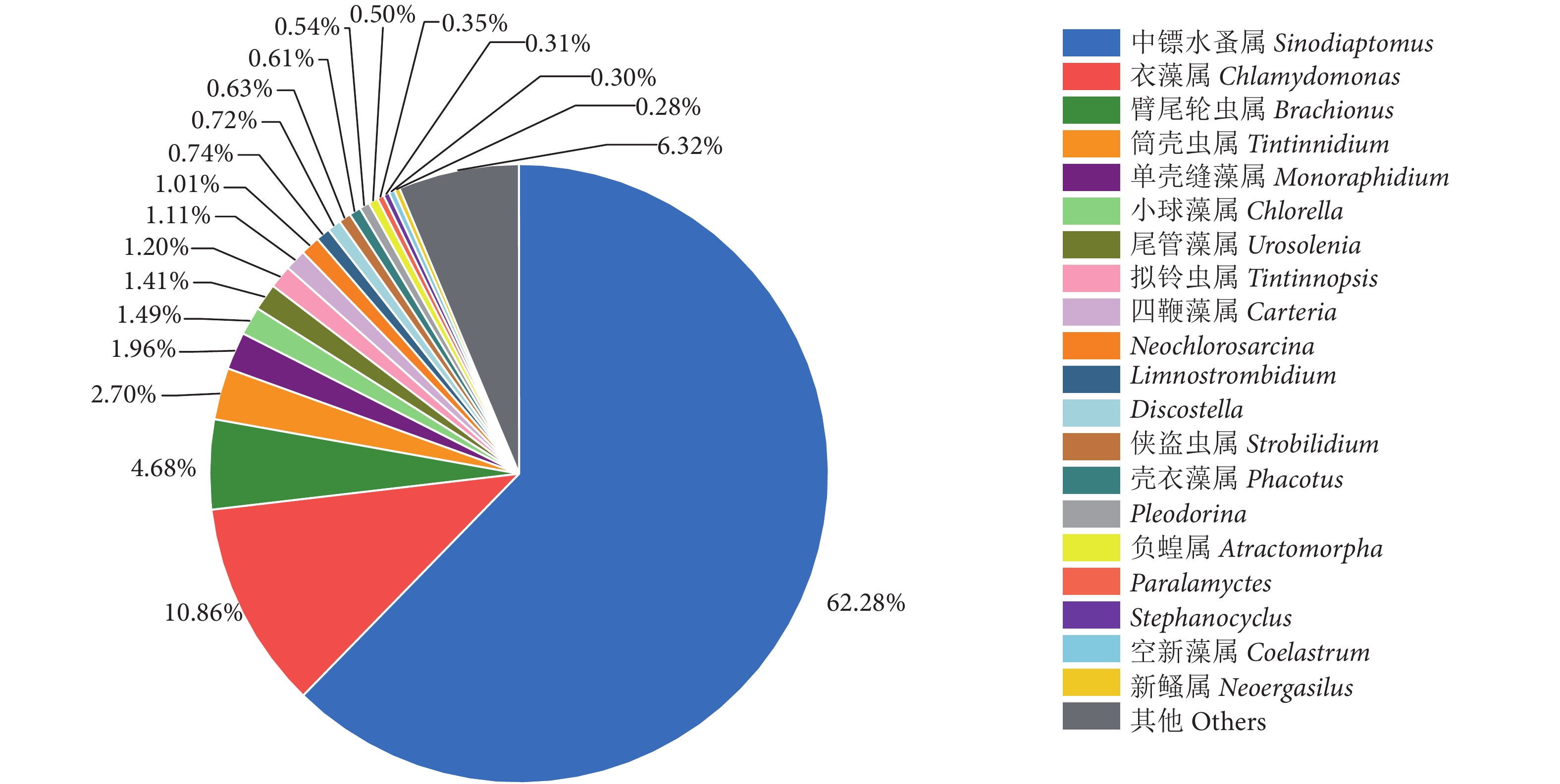
河道水样属水平相对丰度见图5。中镖水蚤属的相对丰度最高 (62.28%);其次是衣藻属(Chlamydomonas, 10.86%);臂尾轮虫属、筒壳虫属 (Tintinnidium) 和单壳缝藻属 (Monoraphidium) 占比也较高,分别为4.68%、2.70%和1.96%。
3. 讨论
3.1 河道斑鳠幼鱼的食性特征
鱼类食性是鱼类生物学和生态学研究的重要内容,鱼类食物谱的广度和组成是影响鱼类生态功能的重要因素[22]。一般来说,鱼类幼鱼孵化后不久就开口摄食,开始是觅食小型的浮游生物、有机碎屑等;随着鱼的生长,逐渐开始觅食大型的浮游生物、轮虫、无节幼体、小型枝角类等水生生物;待鱼体生长到一定阶段,开始分化出浮游生物食性、底栖生物食性、草食性、肉食性和杂食性等[23-24]。
本研究中,斑鳠幼鱼 [(13.97±0.62) cm] 的饵料生物包括甲壳类、轮虫、藻类、鱼类、多足类、原生动物、水生植物、线虫、扁形动物、腹毛动物、昆虫类和环节动物。其中,甲壳类、轮虫和藻类是斑鳠幼鱼主要摄食的饵料,其次是鱼类、多足类、原生动物和水生植物。以往研究通过胃含物形态学鉴定发现,斑鳠幼鱼 (128~169 mm) 以水生昆虫和环节动物为主要食物,藻类、浮游动物、植物碎片、有机碎屑和鱼类居次要地位[20]。基于高通量测序技术的食物组成研究结果与已有的传统形态学食性分析结果基本一致。高通量测序鉴定出了109种饵料生物,其中原生动物、轮虫、线虫在使用传统胃含物形态学方法时未能鉴定,本研究通过DNA宏条形码技术,获得了更多的斑鳠幼鱼胃含物饵料生物信息,进一步完善了斑鳠幼鱼的食物谱。本研究中,斑鳠基本是1龄幼鱼,此阶段是食性逐步分化的阶段,食性与成鱼有所差异。鱼类在此阶段仅作为斑鳠的次要食物,而在斑鳠成鱼 (400~1 070 mm) 中则为仅次于甲壳类的主要食物[20]。此外,斑鳠幼鱼的主要食物中的轮虫和藻类在成鱼中已变为水生昆虫、环节动物等。斑鳠幼鱼与其他幼鱼的食性也有所不同,斑点叉尾鮰幼鱼 (20~26 mm) 主要摄食摇蚊幼虫和枝角类[6],花鲈 (Lateolabrax japonicas) 幼鱼 (17~80 mm) 以桡足类、摇蚊幼虫以及糠虾类为食[25],与斑鳠同为鲿科的黄颡鱼 (Pelteobagrus fulvidraco) 幼鱼主要摄食枝角类,其次是轮虫[26]。斑鳠幼鱼除了摄食藻类外,还摄食甲壳类、轮虫、鱼类、多足类和原生动物等。较广的饵料来源和较少的主食种类,表明斑鳠幼鱼的饵料保障程度较高,饵料基础稳定,也表明贵州省罗甸县斑鳠仿生态保种场有较好的生态环境,能够为斑鳠幼鱼提供较广泛的饵料资源和栖息环境。
在斑鳠幼鱼胃含物中,从属水平看,中镖水蚤属占食物序列相对丰度最高,其次是臂尾轮虫属、鲃属和Paralamyctes,表明在罗甸保种场的河道中,斑鳠幼鱼的天然饵料主要来自桡足亚纲的中镖水蚤属,其次为轮虫纲臂尾轮科的臂尾轮虫属及鲤科鲃属鱼类等。
3.2 斑鳠幼鱼摄食选择性
幼鱼的摄食受饵料的易得性与物种的喜好性两方面的影响[26]。在本研究中,斑鳠幼鱼胃含物中丰度最高的类群来自河道水体中丰度最高的类群 (节肢动物门),胃含物中丰度较高的轮虫动物门、绿藻门也源于水体中丰度较高的生物类群。斑鳠选择它们作为食物,说明斑鳠幼鱼的食性选择受食物易得性的影响。从属水平来看,斑鳠幼鱼胃含物中相对丰度较高的中镖水蚤属、臂尾轮虫属也源自水体中这两属相对丰度较高的生物。斑鳠幼鱼选择资源量丰富的食物意味着捕食容易,即在觅食过程以最小的能量投入获取最大的收益,这些特征体现了斑鳠幼鱼摄食的生态适应性。以往对淀山湖光泽黄颡鱼 (P. nitidus)[27]和大亚湾黑棘鲷 (Acanthopagrus schlegeli)[28]的摄食研究结果也支持这一论点。
水体中脊索动物门的相对丰度较低 (第15位),鲤科鲃属鱼类的相对丰度更低,位于第20位之后,但在斑鳠幼鱼的胃含物中,这两个类群生物的相对丰度较高,脊索动物门在胃含物中位于第4位,鲤科鲃属鱼类位于第3位。表明斑鳠幼鱼的摄食受该物种自身喜好性的影响。张波等[29]对许氏平鲉 (Sebastes schlegelii) 的食性研究表明,摄食选择性并不完全是根据环境中饵料的丰度来决定的,其也可以摄食在环境中丰度较低的饵料种类。Greenstreet等[30]发现牙鳕 (Merlangus merlangius) 和黑线鳕 (Melanogrammus aeglefinus) 的食物组成与捕食者自身的捕食喜好相关。从摄取食物的能量收支来看,生物从食物中获得的能量大于捕食所消耗的能量,才能够获得能量。从食物中获得的能量越是大于捕食所消耗的能量,越能最大限度地获得能量,从而利于幼鱼生存。水体环境中存在的脊索动物门物种及鲃属鱼类,其蛋白质、脂质含量远高于其他类群,因此摄食它们可以有效地获得更多的能量。郭斌等[31]对海州湾黄鲫 (Setipinna taty) 幼鱼的食性研究表明,捕食成本随着食物的增大而下降。刘雪飞等[32]对黄山殷溪河光唇鱼 (Acrossocheilus fasciatus) 食性的测定结果也支持本研究能量收支效益的观点。
综上,斑鳠幼鱼摄食受饵料的易得性与物种的喜好性两方面的影响。从摄取饵料过程中的能量收支效益来看,饵料的易得性体现了斑鳠幼鱼以较少的能量消耗来获取食物;摄食的喜好性,一方面具有该物种的遗传特性,另一方面体现了斑鳠幼鱼通过捕食高蛋白、高脂肪的饵料来获取更多的能量,使其得以更好地存活及生长。
-
![]()
图 2 斑鳠幼鱼胃含物门水平相对丰度
Figure 2. Relative abundance of stomach contents of M. guttatus juvenile fish on phylum level
![]()
图 3 斑鳠幼鱼胃含物属水平相对丰度
Figure 3. Relative abundance of stomach contents of M. guttatus juvenile fish on genus level
表 1 有效序列数及OTU数统计
Table 1 Statistics of number of effective sequences and OTU
样品
Sample原始序列数
Number of original sequences有效序列数
Number of valid sequences有效序列平均长度
Average length of effective sequence/bp最短序列长度
Minimum sequence length/bp最长序列长度
Longest sequence length/bpOTU总数
Total number of OTU斑鳠幼鱼胃含物
Stomach contents of juvenile M. guttatus46 116 37 169 438 310 474 239 河道水环境样品
Sample of river channel water84 413 70 343 416 303 475 441  下载: 导出CSV
下载: 导出CSV
表 2 斑鳠幼鱼胃含物饵料生物种类
Table 2 Types of stomach content of M. guttatus juvenile
门
Phylum属水平种类数
Number of species on genus level绿藻门 Chlorophyta 31 纤毛虫门 Ciliophora 22 轮虫动物门 Rotifera 11 节肢动物门 Arthropoda 11 丝足虫门 Cercozoa 8 硅藻门 Bacillariophyta 7 线虫动物门 Nematoda 6 脊索动物门 Chordata 3 链型植物门 Streptophyta 3 Endomyxa 2 腹毛动物门 Gastrotricha 1 Haptista 1 Imbricatea 1 扁形动物门 Platyhelminthes 1 环节动物门 Annelida 1 总计Total 109
下载: 导出CSV
表 3 斑鳠幼鱼胃含物饵料生物表
Table 3 Biota of bait in stomach contents of M. guttatus juvenile
饵料类别 (数量百分比、种类)
Bait category
(Percentage, Category)门水平
Phylum level纲水平
Class level属水平
Genus level属水平相对丰度
Relative abundance on genus level/%甲壳类 Crustacean
(56.38%, 9)节肢动物门 甲壳纲 明镖水蚤属 Heliodiaptomus 0.01 中剑水蚤属 Mesocyclops 0.03 新鳋属 Neoergasilus 0.28 美丽猛水蚤属 Nitokra 0.10 中镖水蚤属 Sinodiaptomus 54.85 温剑水蚤属 Thermocyclops 0.21 日猛水蚤属 Tisbe 0.22 软甲纲 沼虾属 Macrobrachium 0.66 对虾属 Penaeus 0.02 轮虫 Rotifer
(12.37%, 11)轮虫动物门 真轮纲 Adineta 0.03 晶囊轮属 Asplanchna 1.04 臂尾轮虫属 Brachionus 10.19 Collotheca 0.01 狭甲轮虫属 Colurella 0.21 聚花轮虫属 Conochilus 0.13 三肢轮虫属 Filinia 0.59 巨腕轮属 Hexarthra 0.04 旋轮虫属 Philodina 0.04 多肢轮虫属 Polyarthra 0.02 异尾轮属 Trichocerca 0.07 藻类 Algae
(11.83%, 39)Haptista Centroplasthelida Choanocystis 0.01 硅藻门 硅藻纲 菱形藻属 Nitzschia 0.03 圆筛藻纲 Acanthoceras 0.02 直链藻属 Aulacoseira 0.01 碟星藻属 Discostella 0.32 Stephanocyclus 2.20 尾管藻属 Urosolenia 0.25 间藻纲 无管眼藻属 Extubocellulus 0.01 绿藻门 绿藻纲 负蝗属 Atractomorpha 0.06 荀球藻属 Bracteacoccus 0.01 四鞭藻属 Carteria 0.22 衣藻属 Chlamydomonas 2.66 绿梭藻属 Chlorogonium 0.04 空星藻属 Coelastrum 0.27 Dangeardinia 0.04 迪森藻属 Deasonia 0.07 链带藻属 Desmodesmus 0.14 异形藻属 Dysmorphococcus 0.15 胶胞藻属 Gloeomonas 0.32 Gungnir 0.08 Hamakko 0.04 软壳藻属 Microglena 1.34 单针藻属 Monoraphidium 1.40 麦可属 Mychonastes 0.03 Neochlorosarcina 0.14 润衣藻属 Oogamochlamys 0.01 介壳藻属 Phacotus 0.11 杂球藻属 Pleodorina 0.17 Rotundella 0.01 四角藻属 Tetraedron 0.04 维衣藻属 Vitreochlamys 0.01 平藻纲 单鞭藻属 Pedinomonas 0.01 共球藻纲 小球藻属 Chlorella 0.56 索囊藻属 Choricystis 0.56 Jaagichlorella 0.03 Myrmecia 0.22 卵囊藻属 Oocystis 0.11 微绿藻属 Picochlorum 0.07 石莼纲 石莼属 Ulva 0.06 鱼类 Fish
(6.33%, 3)脊索动物门 鱼纲 鲃属 Barbus 6.16 Eutropiichthys 0.10 翠鳞鱼属 Herklotsichthys 0.07 多足类Myriapods
(5.64%, 1)节肢动物门 唇足纲 Paralamyctes 5.64 原生动物Protozoa
(4.00%, 33)Endomyxa norank_Endomyxa 浮霉菌属 Planctomyxa 0.01 Thalassomyxa 0.02 Imbricatea norank_Imbricatea 奇鞭藻属 Thaumatomastix 0.48 丝足虫门 norank_Cercozoa Cavernomonas 0.01 无吻虫属 Clautriavia 0.12 Massisteria 0.01 Neocercomonas 0.11 Paracercomonas 0.01 Tremula 0.06 Thecofilosea Trachyrhizium 0.04 Ventrifissura 1.02 纤毛虫门 Armophorea Brachonella 0.01 异毛纲 Linostomella 0.04 核残迹纲 Loxodes 0.03 叶口纲 Balantidion 0.02 串须茶属 Cyclotrichium 0.10 Lacrymaria 0.04 寡膜纲 草履虫属 Paramecium 0.08 伪钟虫属 Pseudovorticella 0.04 钟虫属 Vorticella 0.39 叶咽纲 吸管虫属 Acineta 0.03 壳吸管虫属 Paracineta 0.10 前口纲 Balanion 0.07 Hexasterias 0.03 裸口虫属 Holophrya 0.01 Levicoleps 0.03 尾毛虫属 Urotricha 0.04 旋毛纲 异列虫属 Anteholosticha 0.04 纤毛属 Hypotrichidium 0.04 Limnostrombidium 0.09 侠盗虫属 Strobilidium 0.07 筒壳虫属 Tintinnidium 0.59 拟铃虫属 Tintinnopsis 0.22 水生植物 Hydrophyte
(2.90%, 3)链型植物门 木兰纲 Camellia 0.07 大豆属 Glycine 2.77 双星藻纲 新月藻属 Closterium 0.06 线虫Nematode
(0.27%, 6)线虫动物门 色矛纲 滑刃线虫属 Aphelenchoides 0.01 Chromadorina 0.01 异小杆属线虫 Heterorhabditis 0.10 单宫线虫属 Monhystera 0.01 Pellioditis 0.01 Enoplea Viscosia 0.13 扁形动物Platyhelminth
(0.15%, 1)扁形动物门 被杆体纲 Phaenocora 0.15 腹毛动物Gastrotricha
(0.07%, 1)腹毛动物门 norank_Gastrotricha 鼬虫属 Chaetonotus 0.07 昆虫类Hexapeopoda
(0.03%, 1)节肢动物门 昆虫纲 大白蚁属 Macrotermes 0.03 环节动物 Annelid
(0.03%, 1)环节动物门 环带纲 瓢体虫属 Aeolosoma 0.03
下载: 导出CSV
表 4 水样生物种类
Table 4 Biological species of water sample
门
Phylum属水平种类数
Number of species on genus level绿藻门 Chlorophyta 52 纤毛虫门 Ciliophora 45 硅藻门 Bacillariophyta 15 轮虫动物门 Rotifera 14 节肢动物门 Arthropoda 13 线虫动物门 Nematoda 10 丝足虫门 Cercozoa 9 Endomyxa 6 Tubulinea 6 Haptista 5 腹毛动物门 Gastrotricha 4 链型植物门 Streptophyta 3 环节动物门 Annelida 2 顶复动物亚门 Apicomplexa 2 脊索动物门 Chordata 2 Imbricatea 2 扁形动物门 Platyhelminthes 2 Microsporidia 1 总计 Total 193
下载: 导出CSV
-
[1] RIEMANN L, ALFREDSSON H, HANSEN M M, et al. Qualitative assessment of the diet of European eel larvae in the Sargasso Sea resolved by DNA barcoding[J]. Biol Lett, 2010, 6: 819-822. doi: 10.1098/rsbl.2010.0411
[2] ROUL S K, GANGAA U, ABDUSSAMAD E M, et al. Diet composition and feeding habits of flat needlefish Ablennes hians (Valenciennes, 1846) (Beloniformes: Belonidae) in the Southeastern Arabian Sea[J]. Indian J Geo Mar Sci, 2023, 52(1): 24-35.
[3] HAMBRIGHT K D. Experimental analysis of prey selection by largemouth bass: role of predator mouth width and prey body depth[J]. Trans Am Fish Soc, 1991, 120(4): 500-508. doi: 10.1577/1548-8659(1991)120<0500:EAOPSB>2.3.CO;2
[4] FLORIN A, LAVADOS G. Feeding habits of juvenile flatfish in relation to habitat characteristics in the Baltic Sea[J]. Estuar Coastal Shelf Sci, 2010, 86(4): 607-612. doi: 10.1016/j.ecss.2009.11.028
[5] NAKAYA M, ABE T. Feeding habits of juvenile slime flounder Microstomus achne in the coastal area of southern Hokkaido[J]. Ichthyol Res, 2011, 58(4): 377-381. doi: 10.1007/s10228-011-0231-5
[6] YAMAZAKI K, KANOU K, ARAYAMA K. Nocturnal activity and feeding of juvenile channel catfish, Ictalurus punctatus, around offshore breakwaters in Lake Kasumigaura, Japan[J]. Ichthyol Res, 2019, 66: 166-171. doi: 10.1007/s10228-018-0653-4
[7] 龚玉艳, 陈作志, 张俊, 等. 南海北部陆坡海域秋季金鼻眶灯鱼的摄食习性[J]. 南方水产科学, 2015, 11(5): 90-99. doi: 10.3969/j.issn.2095-0780.2015.05.011 [8] 李晓雪, 刘伟, 唐富江, 等. 松花江哈尔滨段三种鮈亚科鱼类的食性与营养级分析[J]. 水产学杂志, 2013, 26(3): 29-33. doi: 10.3969/j.issn.1005-3832.2013.03.006 [9] 苟妮娜, 王开锋, 边坤. 秦巴山区多鳞白甲鱼食性的初步研究[J]. 西北农业学报, 2020, 29(8): 1141-1147. doi: 10.7606/j.issn.1004-1389.2020.08.003 [10] 梁吉, 熊中豪, 马代强, 等. 硬刺松潘裸鲤消化道结构及食性的初步研究[J]. 水产科学, 2024, 43(3): 437-444. [11] SATO T, NAKAMURA K, NISHIMOTO A, et al. Feeding ecology of juvenile Pacific bluefin tuna Thunnus orientalis in the Sea of Japan[J]. Mar Freshw Res, 2021, 73(3): 377-387. doi: 10.1071/MF21200
[12] 严丽君, 王普, 施启龙, 等. 动物食性分析在生态学中的应用研究进展: 基于DNA宏条形码技术[J]. 生态学报, 2023, 43(8): 3007-3019. [13] WHITE N E, GUZIK M T, AUSTIN A D, et al. Detection of the rare Australian endemic blind cave eel (Ophisternon candidum) with environmental DNA: implications for threatened species management in subterranean environments[J]. Hydrobiologia, 2020, 847(15): 3201-3211. doi: 10.1007/s10750-020-04304-z
[14] 吴鹏, 王腾, 刘永, 等. 高通量测序分析西沙晋卿岛两种草食性鱼类对大型海藻的摄食差异[J]. 生态学杂志, 2023, 42(6): 1509-1516. [15] AlBAINA A, AGUIRRE M, ABAD D, et al. 18S rRNA V9 metabarcoding for diet characterization: a critical evaluation with two sympatric zooplanktivorous fish species[J]. Nat Ecol Evol, 2016, 6(6): 1809-1824. doi: 10.1002/ece3.1986
[16] 陈晓雷, 李敏, 陈作志, 等. 基于宏条形码技术的南海亮眶灯鱼食性初步分析[J]. 南方水产科学, 2022, 18(3): 22-29. doi: 10.12131/20210199 [17] 李玉龙, 陈百灵, 鲍相渤, 等. 基于DNA宏条形码技术的沙氏下鱵幼鱼食性分析[J]. 中国水产科学, 2023, 30(4): 393-405. doi: 10.12264/JFSC2023-0003 [18] 杨家坚. 斑鳠鱼种人工培育技术研究[J]. 内陆水产, 2000(11): 5-7. [19] 陈涛, 于丹. 野生斑鳠幼鱼与成鱼的肌肉营养成分测定[J]. 贵州农业科学, 2015, 43(11): 120-123. doi: 10.3969/j.issn.1001-3601.2015.11.031 [20] 胡隐昌, 陈焜慈, 李恒颂, 等. 珠江斑鳠的食性[J]. 水产学报, 2003, 27(4): 301-306. [21] 刘闯. 三种星虫动物的肠道内含物粒径组成和食物来源研究[D]. 海口: 海南大学, 2023: 24-25. [22] CLEMENTS K D, GERMAN D P, PICHE J, et al. Integrating ecological roles and trophic diversification on coral reefs: multiple lines of evidence identify parrotfishes as micro-phages[J]. Biol J Linn Soc, 2017, 120(4): 729-751.
[23] 黄权, 孙兆和. 鸭绿江上游花羔红点鲑的年龄、生长、食性和繁殖的研究[J]. 水产学报, 1998, 22(1): 2-9. [24] 李培伦, 王继隆, 唐富江, 等. 马苏大麻哈鱼幼鱼消化系统形态学和组织学观察[J]. 水产学杂志, 2021, 34(4): 43-48. doi: 10.3969/j.issn.1005-3832.2021.04.007 [25] FUJI T, KASAI A, SUZUKI K W, et al. Freshwater migration and feeding habits of juvenile temperate seabass Lateolabrax japonicus in the stratified Yura River estuary, the Sea of Japan[J]. Fish Sci, 2010, 76(4): 643-652. doi: 10.1007/s12562-010-0258-y
[26] 方巍, 樊启学, 杨瑞斌, 等. 黄颡鱼早期发育阶段摄食研究[J]. 水生态学杂志, 2010, 31(5): 32-37. [27] 刘其根, 吴杰洋, 颜克涛, 等. 淀山湖光泽黄颡鱼食性研究[J]. 水产学报, 2015, 39(6): 859-866. [28] 郭家彤, 王腾, 陈得仿, 等. 大亚湾黑棘鲷的摄食习性[J]. 中国水产科学, 2021, 28(8): 1041-1050. doi: 10.12264/JFSC2020-0576 [29] 张波, 李忠义, 金显仕. 许氏平鲉的食物组成及其食物选择性[J]. 中国水产科学, 2014, 21(1): 134-141. [30] GREENSTREET S P R, MCMILLAN J A, ARMSTRONG E. Seasonal variation in the importance of pelagic fish in the diet of piscivorous fish in the Moray Firth, NE Scotland: a response to variation in prey abundance[J]. ICES J Mar Sci, 1998, 55(1): 121-133. doi: 10.1006/jmsc.1997.0258
[31] 郭斌, 张波, 戴芳群, 等. 海州湾黄鲫幼鱼的食性及其随叉长的变化[J]. 水产学报, 2010, 34(6): 921-927. [32] 刘雪飞, 汪晗, 余银姣, 等. 黄山殷溪河光唇鱼食性特征的初步研究[J]. 淡水渔业, 2016, 46(5): 25-29. -
期刊类型引用(1)
1. 卢玉典,莫飞龙,刘康,韦玲静,张盛,甘宝江,刘羚,杨宾兰. 3个不同群体斑鳠的肌肉营养成分比较. 饲料研究. 2024(24): 118-122 .  百度学术
百度学术
其他类型引用(0)


计量
- 文章访问数: 277
- HTML全文浏览量: 26
- PDF下载量: 76
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
