
Effects of seamount characteristics in Central and Western Pacific Ocean on CPUEs of yellowfin tuna (Thunnus albacares) in longline and purse seine fisheries
-
摘要:
海山是海底重要的生物栖息地类型之一,是研究海洋生物多样性的热点区域。黄鳍金枪鱼 (Thunnus albacares) 广泛分布于中西太平洋,具有极高的生态和经济价值,然而,鲜有关于海山及其相关特征对黄鳍金枪鱼资源丰度和分布影响的研究。基于2010—2021年中西太平洋渔业委员会 (Western and Central Pacific Fisheries Commission, WCPFC) 汇总的延绳钓和围网渔业数据结合海山特征数据,采用广义加性模型 (Generalized additive model, GAM) 分析两种不同捕捞方式的黄鳍金枪鱼单位捕捞努力量渔获量 (Catch per unit effort, CPUE) 与海山相关特征之间的关系。结果表明,中西太平洋两种渔业方式的黄鳍金枪鱼渔获量主要来源于海山区域,海山特征对两种渔业黄鳍金枪鱼的CPUE均产生了极显著性影响 (P<0.001)。在延绳钓渔业中,较高的CPUE出现在山顶深度、粗糙度、底面积和海山密度较小、坡度较缓的区域;而在围网渔业中,较高的CPUE则出现在粗糙度较小、山顶深度较大、底面积较大、较陡峭且密集的海山区域。研究探讨了中西太平洋海山特征对黄鳍金枪鱼不同群体的影响机制,为今后进一步探索黄鳍金枪鱼种群分布和资源丰度变化与海洋环境的关系提供了参考与新思路。
Abstract:Seamounts are one of the important habitat types on the seafloor and a hotspot for marine biodiversity. Yellowfin tuna (Thunnus albacares) is widely distributed in the Western and Central Pacific Ocean (WCPO) with high ecological and economic value. However, there are few studies on the mechanisms by which seamounts and their associated features affect the abundance and distribution of yellowfin tuna resources. In this study, we used longline and purse seine fishery data summarized by the Western and Central Pacific Fisheries Commission (WCPFC) from 2010–2021, in addition with seamount characteristics data, to analyze the impacts of two different types of seamounts on the abundance and distribution of yellowfin tuna resources using a generalized additive model (GAM). GAM was utilized to examine the connection between catch per unit effort (CPUE) and seamounts characteristics of yellowfin tuna in two different fishing methods. The results show that in the WCPO, the yellowfin tuna catches in the two fisheries mainly originated from seamount areas, and seamount characteristics had a highly significant effect on CPUEs of yellowfin tuna in both fisheries (P<0.001). In the longline fishery, higher CPUE occurred in seamount areas with less peak depth, roughness, base area, and seamount density as well as gentler slopes, whereas higher CPUE occurred in the purse seine fishery in seamount areas with less roughness, greater peak depth, greater base area, as well as steeper and denser slopes. In summary, we explore the mechanism of the influence of WCPO seamount characteristics on different populations of yellowfin tuna, which provides new ideas and references for further exploring the relationship between the distribution of yellowfin tuna populations and changes in resource abundance with the marine environment in the future.
-
Keywords:
- Thunnus albacares /
- Seamount /
- Longline /
- Purse seine /
- CPUE /
- Generalized Additive Model (GAM)
-
海山是从海底隆起且至少高达1 000 m的地貌,全球约有4万座海山,几乎覆盖了整个海底[1]。海山作为远洋地区的生态“热点”,其独特的地形特征维持着周围海域特殊的水动力环境,有助于多种海洋生物群落的建立[2],是海洋生物重要的栖息地之一[1],在海洋生态系统中扮演着重要角色[3]。近年来,有关海山及其附近海域生物多样性的研究成为热点[4],大量研究发现,山顶深度较浅[5-7]、坡度较缓[7-10]或高度较高[11-13]的海山区域能聚集更多种类的海洋生物,具有更加复杂的生物多样性。中西太平洋 (Western and Central Pacific Ocean, WCPO) 海底地形复杂,海山数量约占全球海山数量的一半[14],是全球各大洋中最为典型的海山海域,也是全球金枪鱼渔业的主要产区,探索WCPO海山特征与金枪鱼渔业资源种群之间的关系,对海洋生态保护及实现渔业的可持续发展至关重要。
黄鳍金枪鱼 (Thunnus albacares) 是高度洄游的大洋性掠食性鱼类,主要捕食大洋中上层具有垂直洄游习性的物种,作为高营养级捕食者,其在维系海洋生态系统稳定和食物网平衡方面起到重要作用[15];此外,黄鳍金枪鱼还具有很高的经济价值,是WCPO商业性金枪鱼渔业和休闲渔业的主要目标种类之一[16]。黄鳍金枪鱼游动速度快、活动能力强、洄游距离长,其对食物的需求量大、对外界环境较为敏感,因此其种群分布和资源丰度更易受海洋物理和化学因素影响[17-22]。作为WCPO金枪鱼渔业的第二大目标种类,过去十年中,黄鳍金枪鱼资源的养护和可持续性发展受到中西太平洋渔业委员会 (Western and Central Pacific Fisheries Commission, WCPFC) 的高度重视[23],尽管有研究发现WCPO的海山及其周围海域聚集了大量的黄鳍金枪鱼群体,形成了资源丰度较高的区域[24-26];然而,鲜有研究关注海山及其特征对黄鳍金枪鱼资源丰度和种群分布的影响方式和机制。
本研究利用WCPFC汇总的延绳钓和围网渔业数据结合Yesson等[1]的海山数据,分析两种不同捕捞方法下黄鳍金枪鱼单位捕捞努力量渔获量 (Catch per unite effort, CPUE) 与海山特征之间的关系,探索山顶深度、高度、粗糙度、坡度、底面积和密度等海山特征对不同群体黄鳍金枪鱼资源丰度的影响方式和机制,以期为进一步开展WCPO黄鳍金枪鱼资源养护和可持续性开发提供科学依据。
1. 材料与方法
1.1 数据来源
黄鳍金枪鱼渔业数据来自2010—2021年WCPFC公共领域的延绳钓和围网渔业渔获量和努力量汇总数据 (https://www.wcpfc.int/public-domain/),数据空间范围为105°E—135°W, 60°N—85°S,分辨率为5°×5°,时间分辨率为月。
海山数据来自PANGAEA全球海山数据列表中WCPO海域的海山列表 (https://doi.ppangae.de/10.1594/PANGAEA.757562/),数据包括海山的经纬度、山顶深度、高度和底面积等海山特征,空间分辨率为0.008 3°。海山的坡度和粗糙度数据来自GEBCO的全球测深数据 (https://www.gebco.net/data_and_products/gridgri_bathymetry_data/)。
ONI数据源于美国国家海洋与大气局气候预报中心提供的同期对应月份的 Nio3.4区 (120°W—170°W, 5°N—5°S) 的月平均海表温度距平 (SSTa, https://www.cpc.ncep.noaa.gov/pproduct/precip/CWlink/MJO/enso.shtml/)。
1.2 数据处理与分析
1.2.1 渔业数据标准化
CPUE通常被用作衡量渔业资源丰度的指标[27]。本研究依据渔获量和努力量进行数据标准化处理,以经纬度5°×5°单元网格内的月平均CPUE (XCPUE) 作为中西太平洋黄鳍金枪鱼资源的丰度指标 (延绳钓渔业:尾·10−3钩;围网渔业:t·网−1),即:
$$ {X}_{\mathrm{C}\mathrm{P}\mathrm{U}\mathrm{E},y,m,i}=\frac{\sum {C}_{y,m,i}}{\sum {E}_{y,m,i}} $$ (1) 式中:$\sum {C}_{y,m,i} $ 为单元网格内相应年份的月度总渔获量;$ \sum {E}_{y,m,i} $ 为单元网格内相应年份的月度总捕捞努力量;y为年份;m为月份;i为网格单元。
1.2.2 GAM模型的构建
模型构建之前,利用方差膨胀因子 (Variance inflation factor, VIF) 对解释变量进行多重共线性检验,一般认为VIF<5,即不存在多重共线性问题[28]。
广义加性模型 (Generalized Additive Model, GAM) 是一种对自变量和因变量之间的关系没有任何严格函数形式限制的基于加性模型思想的广义线性模型[28]。GAM模型使用非参数的平滑函数拟合变量之间的关系,能很好地拟合变量间的非线性关系,被广泛应用于生态渔业回归分析和预测建模领域[29]。为探索海山及其特征对黄鳍金枪鱼资源丰度的影响,以黄鳍金枪鱼CPUE为响应变量,以海山的主要特征因子、时间和空间因子为解释变量,考虑到全球气候变化特别是厄尔尼诺-南方涛动 (El Niño-Southern Oscillation, ENSO) 现象引起的海洋环境因子变化会对黄鳍金枪鱼群体分布和资源丰度产生影响[30-31],将海洋厄尔尼诺指数 (Oceanic El Niño Index, ONI) 作为解释变量,以期更好地反映海山特征与黄鳍金枪鱼CPUE之间的关系。利用Gaussian分布和以自然对数为连接函数的GAM模型对黄鳍金枪鱼的CPUE与海山特征的关系进行拟合。GAM模型的一般形式如下:
$$ X_{\mathrm{CPUE}}=\gamma+{\sum}_i^n f_i\left(x_i\right)+\varepsilon $$ (2) 式中:XCPUE为黄鳍金枪鱼的CPUE;γ为模型的截距;ε为随机误差项;xi表示解释变量;fi(xi)为各解释变量 xi的任意单变量样条平滑函数。
根据赤池信息准则 (Akaike Information Criterion, AIC),在AIC值最小的解释变量预测函数基础上,依次添加解释变量得到AIC值最小的预测模型,一般认为AIC值越小,R2越接近1,偏差解释率越高,模型拟合效果越好[28]。
模型的构建在R (V4.2.0)中进行,其中GAM构建由mgcv包实现[28],VIF检验由car包实现。显著性水平α设为0.01。
2. 结果
2.1 黄鳍金枪鱼CPUE的分布
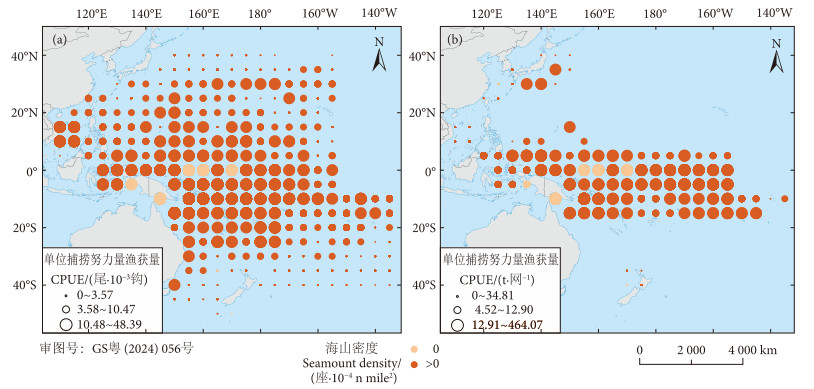
2010—2021年中西太平洋黄鳍金枪鱼资源分布如图1所示。由图可见,WCPO延绳钓渔业黄鳍金枪鱼CPUE主要分布在30°S—30°N区域 (图1-a),围网渔业黄鳍金枪鱼CPUE主要分布在20°S—10°N区域 (图1-b),且在海山区域具有较高的CPUE。通过计算2010—2021年海山区域渔获量在中西太平洋总体中的比例发现,2010—2021年延绳钓渔业共捕捞黄鳍金枪鱼约8.3×105 t,海山区域渔获量占98%;围网渔业共捕捞黄鳍金枪鱼约3.64×105 t,海山区域渔获量占89%。
![]() 图 1 2010—2021年WCPO延绳钓 (a) 和围网 (b) 黄鳍金枪鱼月平均CPUE的分布Fig. 1 Distribution of monthly average CPUEs of yellowfin tuna of longline (a) and purse seine (b) fisheries in WCPO from 2010 to 2021
图 1 2010—2021年WCPO延绳钓 (a) 和围网 (b) 黄鳍金枪鱼月平均CPUE的分布Fig. 1 Distribution of monthly average CPUEs of yellowfin tuna of longline (a) and purse seine (b) fisheries in WCPO from 2010 to 20212.2 GAM模型的筛选和拟合
初始变量的多重共线性检验结果见表1,VIF值均小于5,即初始变量的多重共线性程度较小。因此将年 (Y)、月(M)、经度 (Lon)、纬度 (Lat)、ONI、海山密度 (DS,座·10−4 n mile2)、山顶深度 (PD, m)、高度 (E, m)、底面积 (BA, km2)、坡度 (SP) 和粗糙度 (R) 作为解释变量构建GAM模型。
表 1 不同解释变量的多重共线性检验Table 1 Multicollinearity test for different explanatory variables变量 Variable VIF (Y) VIF (M) VIF (Lon) VIF (Lat) VIF (ONI) VIF (PD) 延绳钓 Longline 1.024 1.014 1.499 1.717 1.027 1.421 围网 Purse seine 1.011 1.008 3.37 2.111 1.014 1.812 变量 Variable VIF (E) VIF (BA) VIF (SP) VIF (R) VIF (DS) 延绳钓 Longline 1.852 1.58 3.517 2.983 1.097 围网 Purse seine 2.207 1.763 4.812 4.002 1.143 通过GAM模型拟合中西太平洋黄鳍金枪鱼CPUE与各解释变量的关系,使用AIC准则进行最优模型筛选,拟合结果见表2。其中,延绳钓渔业最优模型的偏差解释率为52.20%,决定系数R2为0.520;围网渔业最优模型的偏差解释率为51.00%,决定系数R2为0.505,最优模型表达式为:
表 2 最优GAM模型的筛选及拟合Table 2 Selection and fitting of optimal GAM解释变量
Explanatory variable延绳钓 Longline 围网 Purse seine 赤池信息准则
AIC偏差解释率
Deviance explained/%决定系数
R2赤池信息准则
AIC偏差解释率
Deviance explained/%决定系数
R2年 Y 48 437 3.46 0.034 14 789 1.45 0.013 月 M 48 314 4.04 0.039 14 628 4.22 0.039 经度 Lon 45 574 15.10 0.15 12 616 31.10 0.308 纬度 Lat 33 835 49.70 0.496 10 963 47.50 0.472 海洋厄尔尼诺指数 ONI 33 766 49.90 0.498 10 940 47.80 0.475 山顶深度 PD/m 33 422 50.60 0.505 10872 48.50 0.481 高度 E/m 33 115 51.30 0.512 10 806 49.00 0.486 粗糙度 R 33 081 51.40 0.513 10 757 49.50 0.491 底面积 BA/km2 32 900 51.80 0.517 10 727 49.80 0.494 坡度 SP 32 805 52.00 0.519 10 674 50.30 0.498 海山密度 DS/(座·10−4 n mile2) 32 758 52.20 0.52 10 593 51.00 0.505 $$ \begin{array}{c} \ln \left(X_{\mathrm{CPUE}}+1\right) \sim f(Y)+s(M)+s(\mathrm{Lon})+s(\mathrm{Lat})+\\ s(\mathrm{ONI})+s(\mathrm{DS})+s(\mathrm{PD})+s(E)+s(\mathrm{BA})+s(\mathrm{SP})+s(R) \end{array} $$ (3) 式中:Y作为因子;其他变量为自然立方样条平滑 (Natural cube spline smoother) 函数s。
中西太平洋黄鳍金枪鱼CPUE与各影响因子间的关系如表3所示,在P<0.01条件下,对黄鳍金枪鱼延绳钓渔业和围网渔业CPUE影响显著的变量分别包括时空变量月、经度、纬度,气候变量ONI以及海山特征变量山顶深度、高度、底面积、坡度、粗糙度、海山密度。
表 3 延绳钓和围网渔业黄鳍金枪鱼CPUE的影响因子Table 3 Factors affecting CPUE of yellowfin tuna in longline and purse seine fisheries影响因子
Factor延绳钓 Longline 围网 Purse seine 自由度 DF F P 自由度 DF F P 月 M 6.95 38.57 <0.001*** 7.52 25.98 <0.001*** 经度 Lon 8.839 240 <0.001*** 8.689 34.14 <0.001*** 纬度 Lat 8.974 1079.86 <0.001*** 8.854 127.56 <0.001*** 海洋厄尔尼诺指数 ONI 7.46 10.04 <0.001*** 7.079 4.42 <0.001*** 山顶深度 PD/m 3.966 130.31 <0.001*** 3.823 17.55 <0.001*** 高度 E/m 3.765 20.94 <0.001*** 1.962 20.09 <0.001*** 粗糙度 R 3.5 20.81 <0.001*** 3.856 20.16 <0.001*** 底面积 BA/km2 3.481 35.39 <0.001*** 4.805 4.5 0.000 812*** 坡度 SP 3.985 26.73 <0.001*** 3.87 13.94 <0.001*** 海山密度 DS/(座·10−4n mile2) 3.104 15.34 <0.001*** 3.493 24.14 <0.001*** 注:***. 极显著性影响 (P<0.001)。 Note: ***. Very significant effect (P<0.001). 2.3 海山特征对CPUE的影响
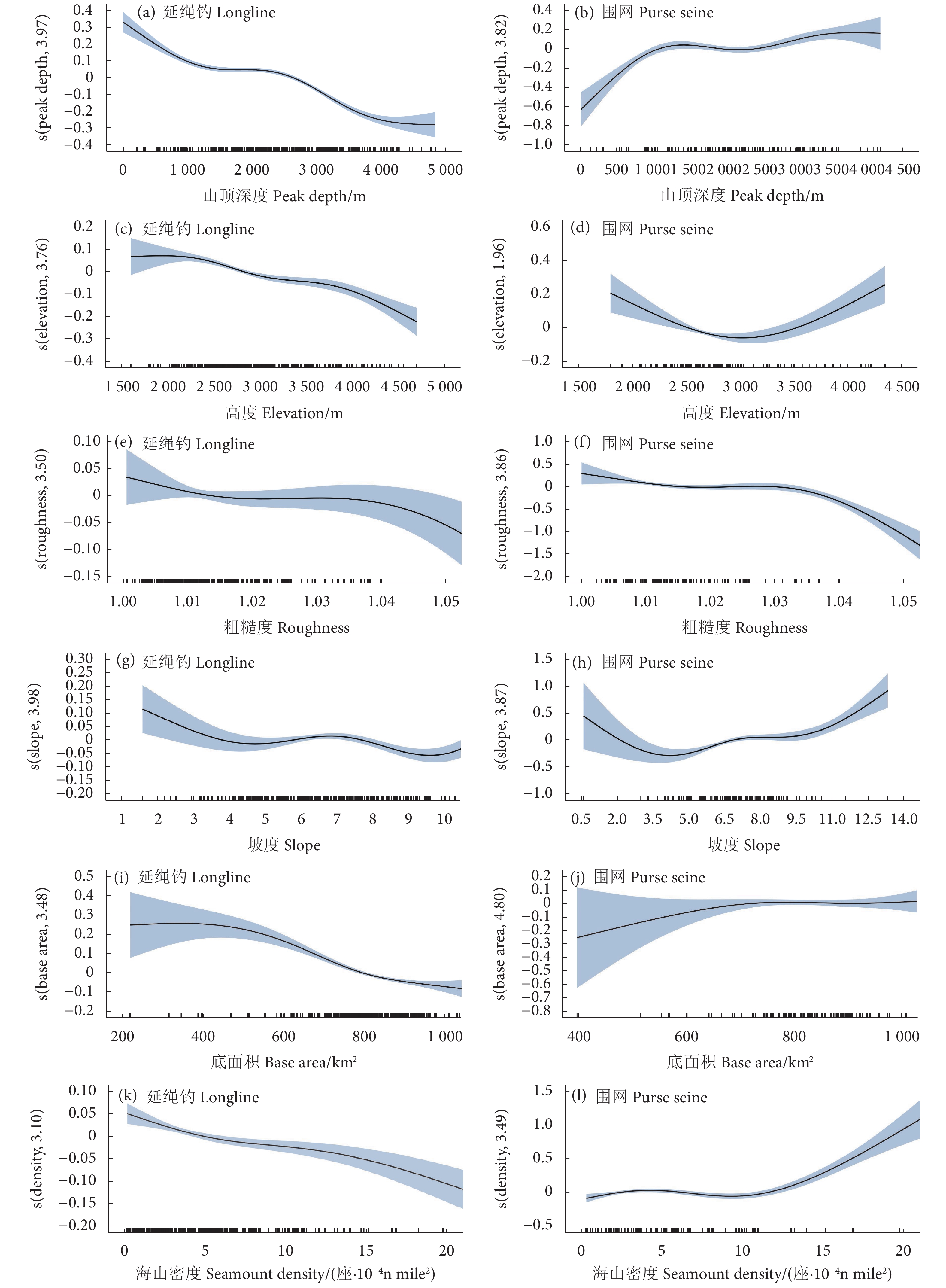
影响中西太平洋延绳钓渔业和围网渔业黄鳍金枪鱼CPUE的海山特征如图2所示,黄鳍金枪鱼CPUE与海山特征之间的关系均为极显著相关 (表2,P<0.001)。在延绳钓渔业中,黄鳍金枪鱼CPUE随着海山山顶深度 (图2-a)、高度 (图2-c)、粗糙度 (图2-e)、坡度 (图2-g)、底面积 (图2-i) 和密度 (图2-k) 的增加而减小。
![]() 图 2 海山特征对黄鳍金枪鱼CPUE的影响Fig. 2 Effects of seamount characteristics on CPUE of yellowfin tuna
图 2 海山特征对黄鳍金枪鱼CPUE的影响Fig. 2 Effects of seamount characteristics on CPUE of yellowfin tuna在围网渔业中,黄鳍金枪鱼CPUE随着山顶深度 (图2-b)、底面积 (图2-j) 和密度 (图2-l) 的增加而上升,海山高度 (图2-d) 和坡度 (图2-h) 对黄鳍金枪鱼CPUE的影响呈先减后增的趋势,海山的粗糙度和黄鳍金枪鱼CPUE整体为负相关 (图2-f)。
3. 讨论
3.1 海山特征对WCPO黄鳍金枪鱼栖息与分布的影响
与平坦的海底相比,海山及附近海域具有更高的生产力[32],其独特的地形特征和海洋模式是构成这些海域食物网结构和功能高度变化的主要原因[33]。在地球自转影响下,海流被海山阻碍后分裂,并在另一侧汇聚,在海山周围形成一个孤立的反气旋流和等密度穹窿,即泰勒柱[34]。泰勒柱会使水中的营养物质涌向海山顶部区域[3],这些营养物质可以促进浮游生物生长,是形成食物网的底层基础,促进了海山附近的消费者和顶级捕食者的扩散和聚集[25]。作为大洋中的顶级捕食者之一,黄鳍金枪鱼食性广泛,主要以高度聚集的小型鱼类、头足类和甲壳类等为食[18]。海山特征会影响黄鳍金枪鱼的资源丰度、资源分布及洄游,常作为黄鳍金枪鱼迁徙途中的导航路标、捕食中转站及产卵地[26]。本研究使用GAM模型对黄鳍金枪鱼的CPUE与海山特征之间的关系进行了深入分析,发现了一系列重要的关联性,在WCPO海域,海山的山顶深度、高度、坡度、粗糙度、底面积、密度对延绳钓渔业和围网渔业黄鳍金枪鱼CPUE均存在极显著影响。
海山生态系统的形成高度依赖于海山区域的营养物质浓度和初级生产力,泰勒柱可以将深层丰富的营养物质抬升到海山顶部[34],山顶深度是影响泰勒柱形成的关键因素[35],浅海山通常具有较高的生产力[6]。Morato等[25-26]认为,金枪鱼等远洋鱼类常因捕食而聚集在浅海山附近,本研究也发现在延绳钓渔业中,黄鳍金枪鱼较高的CPUE出现在海山山顶深度较浅的区域,印证了上述观点,海山顶部丰富的食物来源满足了黄鳍金枪鱼的摄食需求。然而,在围网渔业中,黄鳍金枪鱼的CPUE随着山顶深度的增加而上升,主要原因可能是其群体习性的差异:首先,延绳钓渔业捕获的黄鳍金枪鱼主要为分散存在的成年黄鳍金枪鱼个体,而围网渔业捕获的黄鳍金枪鱼主要为金枪鱼群体且较年幼[36],作为机会主义掠食者[17],黄鳍金枪鱼可能会放弃海山顶部丰富的食物资源以躲避天敌威胁;其次,围网作业覆盖范围较广,可能捕获了较深水域中正在追赶猎物的黄鳍金枪鱼群[19]。海山高度也是影响黄鳍金枪鱼CPUE的因素之一,在延绳钓渔业中,随着海山高度的增加,黄鳍金枪鱼CPUE呈下降趋势。这可能是因为过高的海山位于冷水团上方[37],不利于生物生长,从而限制了金枪鱼的食物资源。围网渔业中,海山高度对黄鳍金枪鱼CPUE的影响呈先减后增的趋势,CPUE主要集中在高度适中的海山区域,这也反映了黄鳍金枪鱼幼龄群体的生境偏好,这些海山可能为黄鳍金枪鱼提供了更复杂的栖息地以躲避捕食者的威胁[3];关于海山高度对黄鳍金枪鱼的影响,还有待进一步探究。
海山密度和底面积会影响海山区域的生物多样性,底面积和密度较大的海山具有更高的生物量[9,32]。本研究结果显示,延绳钓渔业黄鳍金枪鱼的CPUE随着海山密度和底面积的增加而减小,其原因可能是黄鳍金枪鱼个体间的竞争及行为调整等。首先,底面积较大、较密集的海山区域拥有更多的栖息地和食物来源[38],黄鳍金枪鱼被吸引而聚集在一个有限的区域时,不同个体间将不得不竞争相对有限的食物资源,甚至捕食同类,导致该区域的CPUE较低。其次,黄鳍金枪鱼是高度迁徙性鱼类,可以根据食物资源的可获得性来改变其迁徙路径和捕食行为[20],当其察觉到天敌在海山周围的密度增加并且食物更难获得时,一些个体可能会选择离开,以寻找更有利的捕食场所。而在围网渔业中,海山底面积和密度与黄鳍金枪鱼的CPUE呈正相关。由上述可知,围网捕获的黄鳍金枪鱼个体多为幼龄群体,游泳能力较差[39],易受天敌威胁;因此,海山密度、底面积较大的区域可能为黄鳍金枪鱼幼龄群体提供了更多的生存机会。
陡峭的海山上具有裸露的岩石,其表面生长着巨型藻类和珊瑚等生物,造就了复杂的海山栖息地环境[40];平缓的海山则覆盖了大量沉积物,这些高营养基质为海洋生物提供了生命之源[3]。粗糙度是陆地水文模拟和预测的重要因子之一,同时,也是衡量海底沉积物的重要物理性质[41]。本研究发现,无论延绳钓渔业还是围网渔业,海山的粗糙度与黄鳍金枪鱼的CPUE均呈负相关。如前文所述,在食物资源充足的较粗糙的海山区域,黄鳍金枪鱼个体之间的竞争以及面临的威胁也将增加,这可能是导致该结果的原因之一。海山坡度对黄鳍金枪鱼CPUE的影响在两种渔业中有所区别,在延绳钓渔业中,黄鳍金枪鱼的CPUE随海山坡度增加而减小,海山坡度较小的区域具有稳定的底质和适宜的水流条件,生物多样性较高,为黄鳍金枪鱼提供了丰富的食物来源[42]。围网渔业中,黄鳍金枪鱼CPUE整体上随着海山坡度的增加而上升,这可能表明,坡度较大的海山具有复杂的栖息地环境条件,捕捞难度增加,给黄鳍金枪鱼幼鱼提供了更多的逃逸机会[43]。
3.2 海山对黄鳍金枪鱼资源管理的影响及海山栖息地的保护
WCPO的黄鳍金枪鱼广泛分布于30°N—20°S海域[17]。黄鳍金枪鱼通常停留在表层与混合层之间,或分布在温跃层顶部 (18~31 ℃) [44]。部分研究表明,为了追赶猎物或躲避捕食者,它们会迅速下潜至1 600 m或在200~400 m水层内并往返跳潜[45];因此,难以在海洋中建立黄鳍金枪鱼的静态保护区[46]。此外,从渔业执法和管理的角度来看,在整个海洋范围内建立黄鳍金枪鱼保护区可能不切实际[47],但可以通过有针对性地建立局部海洋保护区来实现黄鳍金枪鱼等洄游类物种的保护,即保护黄鳍金枪鱼所在的关键区域,如产卵场、觅食聚集地等[48];因此,识别这些关键区域对海洋保护区的设计至关重要。本研究结果可为黄鳍金枪鱼的渔业管理提供一些依据,在延绳钓渔业中,可以限制在山顶深度较小或坡度较平缓的海山区域的捕捞活动,因为这些海山可能是黄鳍金枪鱼的觅食地[32];而围网渔业中,则应适当关注海山底面积和密度较大或坡度较陡的海山区域的捕捞活动,因为这些区域不仅给黄鳍金枪鱼提供了庇护地[43],还可能是其产卵场[25]。为了黄鳍金枪鱼资源的可持续性利用,可以考虑根据不同的海山特征来调整捕捞活动或根据海山特征进行分组帮助确定黄鳍金枪鱼渔场养护的优先顺序[5],并制定相关的渔业管理政策[49-51]。
大型远洋鱼类,特别是金枪鱼,倾向于聚集在上升流区[52],靠近海洋大陆斜坡、岛屿或海山[53],海山在维持海洋生物多样性和海洋生态平衡方面起着至关重要的作用[54],为海洋生物提供了复杂的栖息地[8,25,55]。黄鳍金枪鱼是海山生态系统的一部分,其资源丰度和分布受到海山特征的影响[3]。因此,保护和管理海山生态系统对于维持黄鳍金枪鱼的可持续性捕获至关重要[33],海山海域不仅为金枪鱼提供了丰富的饵料,同时也是其重要的产卵场[56-58];基于渔业CPUE所反映的海山及其特征对群体的影响可能更多地体现了群体集群和觅食行为对海山的响应,而海山对产卵场的影响机制,仍需进一步探索。
本研究揭示了WCPO海山特征与黄鳍金枪鱼CPUE之间的关系,研究结果为黄鳍金枪鱼与海山生态系统之间复杂相互关系提供了部分见解,为黄鳍金枪鱼资源的可持续性开发与利用以及海山生态系统的保护提供了一定的理论参考。为了进一步了解大洋洄游类物种群体与海山之间的关系,未来研究中,在构建模型时可以考虑海洋锋面、涡流、洄游路线、产卵场的研究等因素,以建立更全面的模型来解释洄游类物种群体的变化。
-
![]()
图 1 2010—2021年WCPO延绳钓 (a) 和围网 (b) 黄鳍金枪鱼月平均CPUE的分布
Figure 1. Distribution of monthly average CPUEs of yellowfin tuna of longline (a) and purse seine (b) fisheries in WCPO from 2010 to 2021
![]()
图 2 海山特征对黄鳍金枪鱼CPUE的影响
Figure 2. Effects of seamount characteristics on CPUE of yellowfin tuna
![]()
Figure 1. Distribution of monthly average CPUEs of yellowfin tuna in longline (a) and purse seine (b) fisheries in the WCPO from 2010 to 2021
表 1 不同解释变量的多重共线性检验
Table 1 Multicollinearity test for different explanatory variables
变量 Variable VIF (Y) VIF (M) VIF (Lon) VIF (Lat) VIF (ONI) VIF (PD) 延绳钓 Longline 1.024 1.014 1.499 1.717 1.027 1.421 围网 Purse seine 1.011 1.008 3.37 2.111 1.014 1.812 变量 Variable VIF (E) VIF (BA) VIF (SP) VIF (R) VIF (DS) 延绳钓 Longline 1.852 1.58 3.517 2.983 1.097 围网 Purse seine 2.207 1.763 4.812 4.002 1.143  下载: 导出CSV
下载: 导出CSV
表 2 最优GAM模型的筛选及拟合
Table 2 Selection and fitting of optimal GAM
解释变量
Explanatory variable延绳钓 Longline 围网 Purse seine 赤池信息准则
AIC偏差解释率
Deviance explained/%决定系数
R2赤池信息准则
AIC偏差解释率
Deviance explained/%决定系数
R2年 Y 48 437 3.46 0.034 14 789 1.45 0.013 月 M 48 314 4.04 0.039 14 628 4.22 0.039 经度 Lon 45 574 15.10 0.15 12 616 31.10 0.308 纬度 Lat 33 835 49.70 0.496 10 963 47.50 0.472 海洋厄尔尼诺指数 ONI 33 766 49.90 0.498 10 940 47.80 0.475 山顶深度 PD/m 33 422 50.60 0.505 10872 48.50 0.481 高度 E/m 33 115 51.30 0.512 10 806 49.00 0.486 粗糙度 R 33 081 51.40 0.513 10 757 49.50 0.491 底面积 BA/km2 32 900 51.80 0.517 10 727 49.80 0.494 坡度 SP 32 805 52.00 0.519 10 674 50.30 0.498 海山密度 DS/(座·10−4 n mile2) 32 758 52.20 0.52 10 593 51.00 0.505
下载: 导出CSV
表 3 延绳钓和围网渔业黄鳍金枪鱼CPUE的影响因子
Table 3 Factors affecting CPUE of yellowfin tuna in longline and purse seine fisheries
影响因子
Factor延绳钓 Longline 围网 Purse seine 自由度 DF F P 自由度 DF F P 月 M 6.95 38.57 <0.001*** 7.52 25.98 <0.001*** 经度 Lon 8.839 240 <0.001*** 8.689 34.14 <0.001*** 纬度 Lat 8.974 1079.86 <0.001*** 8.854 127.56 <0.001*** 海洋厄尔尼诺指数 ONI 7.46 10.04 <0.001*** 7.079 4.42 <0.001*** 山顶深度 PD/m 3.966 130.31 <0.001*** 3.823 17.55 <0.001*** 高度 E/m 3.765 20.94 <0.001*** 1.962 20.09 <0.001*** 粗糙度 R 3.5 20.81 <0.001*** 3.856 20.16 <0.001*** 底面积 BA/km2 3.481 35.39 <0.001*** 4.805 4.5 0.000 812*** 坡度 SP 3.985 26.73 <0.001*** 3.87 13.94 <0.001*** 海山密度 DS/(座·10−4n mile2) 3.104 15.34 <0.001*** 3.493 24.14 <0.001*** 注:***. 极显著性影响 (P<0.001)。 Note: ***. Very significant effect (P<0.001).
下载: 导出CSV
Table 3 Factors affecting the CPUE of yellowfin tuna in longline and purse seine fisheries
 下载: 导出CSV
下载: 导出CSV
-
[1] YESSON C, LETESSIER T, NIMMO-SMITH A, et al. Improved bathymetry leads to >
4000 new seamount predictions in the global ocean–but beware of phantom seamounts[J]. UCL Press, 2021, 4: 1-9.[2] YESSON C, CLARK M R, TAYLOR M L, et al. The global distribution of seamounts based on 30 arc seconds bathymetry data[J]. Deep-Sea Res I, 2011, 58(4): 442-453. doi: 10.1016/j.dsr.2011.02.004
[3] ROGERS A D. The biology of seamounts: 25 years on[J]. Adv Mar Biol, 2018, 79: 137-224.
[4] BO M, COPPARI M, BETTI F, et al. Unveiling the deep biodiversity of the Janua Seamount (Ligurian Sea): first Mediterranean sighting of the rare Atlantic bamboo coral Chelidonisis aurantiaca Studer, 1890[J]. Deep-Sea Res I, 2020, 156: 103186. doi: 10.1016/j.dsr.2019.103186
[5] PITCHER T J, MORATO T, HART P J B, et al. Seamounts: ecology, fisheries & conservation[M]. Oxford: Blackwell Pub, 2007: 85-100, 442-475.
[6] GONZLEZ-IRUSTA J M, DE LA TORRIENTE A, PUNZN A, et al. Living at the top. Connectivity limitations and summit depth drive fish diversity patterns in an isolated seamount[J]. Mar Ecol Prog Ser, 2021, 670: 121-137. doi: 10.3354/meps13766
[7] BRIDGES A E H, BARNES D K A, BELL J B, et al. Depth and latitudinal gradients of diversity in seamount benthic communities[J]. J Biogeogr, 2022, 49(5): 904-915. doi: 10.1111/jbi.14355
[8] MCCLAIN C R, LUNDSTEN L. Assemblage structure is related to slope and depth on a deep offshore Pacific seamount chain[J]. Mar Ecol, 2015, 36(2): 210-220. doi: 10.1111/maec.12136
[9] HANN C H, SMITH T D, AND TORRES L G. A sperm whale's perspective: the importance of seasonality and seamount depth[J]. Mar Mammal Sci, 2016, 32(4): 1470-1481. doi: 10.1111/mms.12320
[10] ROWDEN A A, SCHLACHER T A, WILLIAMS A, et al. A test of the seamount oasis hypothesis: seamounts support higher epibenthic megafaunal biomass than adjacent slopes[J]. Mar Ecol, 2010, 31(1): 95-106.
[11] CAMPANELLA F, COLLINS M A, YOUNG E F, et al. First insight of meso-and bentho-pelagic fish dynamics around remote seamounts in the South Atlantic Ocean[J]. Front Mar Sci, 2021, 8: 693.
[12] VASSALLO P, PAOLI C, ALESSI J, et al. Seamounts as hot-spots of large pelagic aggregations[J]. Mediterr Mar Sci, 2018, 19(3): 444-458. doi: 10.12681/mms.15546
[13] KVILE K , TARANTO G H, PITCHER T J, et al. A global assessment of seamount ecosystems knowledge using an ecosystem evaluation framework[J]. Biol Conserv, 2014, 173: 108-120. doi: 10.1016/j.biocon.2013.10.002
[14] DEARY A L, MORET-FERGUSON S, ENGELS M, et al. Influence of central pacific oceanographic conditions on the potential vertical habitat of four tropical tuna species1[J]. Pac Sci, 2015, 69(4): 461-475. doi: 10.2984/69.4.3
[15] DUFFY L M, KUHNERT P M, PETHYBRIDGE H R, et al. Global trophic ecology of yellowfin, bigeye, and albacore tunas: understanding predation on micronekton communities at ocean-basin scales[J]. Deep-Sea Res II, 2017, 140: 55-73. doi: 10.1016/j.dsr2.2017.03.003
[16] DORTEL E, PECQUERIE L, CHASSOT E. A Dynamic Energy Budget simulation approach to investigate the eco-physiological factors behind the two-stanza growth of yellowfin tuna (Thunnus albacares)[J]. Ecol Model, 2020, 437: 109297. doi: 10.1016/j.ecolmodel.2020.109297
[17] PECORARO C, ZUDAIRE I, BODIN N, et al. Putting all the pieces together: integrating current knowledge of the biology, ecology, fisheries status, stock structure and management of yellowfin tuna (Thunnus albacares)[J]. Rev Fish Biol Fish, 2016, 27(4): 811-841.
[18] 石肖飞, 王啸, 王佚兮, 等. 热带中西太平洋海域黄鳍金枪鱼的摄食生物学特性[J]. 南方水产科学, 2022, 18(1): 43-51. doi: 10.12131/20210140 [19] SCHAEFER K M, FULLER D W, AND BLOCK B A. Movements, behavior, and habitat utilization of yellowfin tuna (Thunnus albacares) in the Pacific Ocean off Baja California, Mexico, determined from archival tag data analyses, including unscented Kalman filtering[J]. Fish Res, 2011, 112(1/2): 22-37.
[20] WRIGHT S R, RIGHTON D, NAULAERTS J, et al. Yellowfin tuna behavioural ecology and catchability in the South Atlantic: the right place at the right time (and depth)[J]. Front Mar Sci, 2021, 8: 664593. doi: 10.3389/fmars.2021.664593
[21] DAGORN L, HOLLAND K N, HALLIER J P, et al. Deep diving behavior observed in yellowfin tuna (Thunnus albacares)[J]. Aquat Living Resour, 2006, 19(1): 85-88. doi: 10.1051/alr:2006008
[22] CLARK M R, ROWDEN A A, SCHLACHER T, et al. The ecology of seamounts: structure, function, and human impacts[J]. Annu Rev Mar Sci, 2010, 2: 253-278. doi: 10.1146/annurev-marine-120308-081109
[23] PILLING G, SCOTT F, AND HAMPTON S. Minimum target reference points for WCPO yellowfin and bigeye tuna consistent with alternative LRP risk levels, and multispecies implications[R/OL]. [2030-09-10] Pohnpei: Technical Report WCPFC-SC15-2019/MI-WP-01, 2019. https://www.wcpfc.int/doc/wcpfc16-2019-15-update-sc15-mi-wp-01/minimum-target-reference-points-wcpo-yellowfin-and-bigeye.pdf
[24] ZHAO R J, ZHAO F, FENG L, et al. A deep seamount effect enhanced the vertical connectivity of the planktonic community across 1,000 m above summit[J]. J Geophys Res-Oceans, 2023, 128(3): e2022JC018898. doi: 10.1029/2022JC018898
[25] MORATO T, HOYLE S D, ALLAIN V, et al. Seamounts are hotspots of pelagic biodiversity in the open ocean[J]. P Natl Acad Sci USA, 2010, 107(21): 9707-9711. doi: 10.1073/pnas.0910290107
[26] MORATO T, VARKEY D A, DAMASO C, et al. Evidence of a seamount effect on aggregating visitors[J]. Mar Ecol Prog Ser, 2008, 357(4): 23-32.
[27] MAUNDER M N, STARR P J. Fitting fisheries models to standardised CPUE abundance indices[J]. Fish Res, 2003, 63(1): 43-50. doi: 10.1016/S0165-7836(03)00002-X
[28] PEDERSEN E J, MILLER D L, SIMPSON G L, et al. Hierarchical generalized additive models in ecology: an introduction with mgcv[J]. PeerJ, 2019, 7: e6876. doi: 10.7717/peerj.6876
[29] MARN-ENRQUEZ E, RAMREZ-PREZ J S, RUIZ-DOMNGUEZ M, et al. Effect of marine climate and baitfish availability on the tuna baitboat fishery CPUE off northwestern Mexico[J]. Ocean Coast Manag, 2023, 232: 106418. doi: 10.1016/j.ocecoaman.2022.106418
[30] VAIHOLA S, KININMONTH S. Climate change potential impacts on the tuna fisheries in the exclusive economic zones of Tonga[J]. Diversity, 2023, 15(7): 844. doi: 10.3390/d15070844
[31] LEHODEY P, BERTRAND A, HOBDAY A J, et al. ENSO impact on marine fisheries and ecosystems[M]. Washington: Wiley-American Geophysical Union, 2020: 429-451.
[32] BO M, COPPARI M, BETTI F, et al. The high biodiversity and vulnerability of two Mediterranean bathyal seamounts support the need for creating offshore protected areas[J]. Aquat Conserv, 2021, 31(3): 543-566. doi: 10.1002/aqc.3456
[33] HANAFI-PORTIER M, SAMADI S, CORBARI L, et al. Multiscale spatial patterns and environmental drivers of seamount and island slope megafaunal assemblages along the Mozambique channel[J]. Deep-Sea Res I, 2024, 203: 104198. doi: 10.1016/j.dsr.2023.104198
[34] PITCHER T J, MORATO T, HART P J B, et al. Seamounts: ecology, fisheries & conservation[M]. Oxford: Blackwell Pub, 2007: 238-239.
[35] CLARK M R. Deep sea seamount fisheries: a review of global status and future prospects[J]. Lat Am J Aquat Res, 2009, 37(3): 501-512. doi: 10.3856/vol37-issue3-fulltex-16
[36] ITANO D. A summary of operational, technical and fishery information on WCPO purse seine fisheries operating on floating objects[R/OL]. [2023-09-10]Honolulu: Scientific Committee 3rd Regular Session of Western and Central Pacific Fisheries Commission, 2007. https://www.wcpfc.int/system/files/SC3_FT-IP-4_Itano%20FO_complete.pdf
[37] PITCHER T J, MORATO T, HART P J B, et al. seamounts: ecology, fisheries & conservation[M]. Oxford: Blackwell Pub, 2007: 85-100.
[38] PRECIADO I, CARTES J E, PUNZN A, et al. Food web functioning of the benthopelagic community in a deep-sea seamount based on diet and stable isotope analyses[J]. Deep-Sea Res I, 2017, 137: 56-68.
[39] BRILL R W, LUTCAVAGE M E. Understanding environmental influences on movements and depth distributions of tunas and billfishes can significantly improve population assessments[C]//American Fisheries Society Symposium. Phoenix: American Fisheries Society, 2001: 179-198.
[40] AUSCAVITCH S R, DEERE M C, KELLER A G, et al. Oceanographic drivers of deep-sea coral species distribution and community assembly on seamounts, islands, atolls, and reefs within the phoenix islands protected area[J]. Front Mar Sci, 2020, 7: 42. doi: 10.3389/fmars.2020.00042
[41] CLARK M R, BOWDEN D A. Seamount biodiversity: high variability both within and between seamounts in the Ross Sea region of Antarctica[J]. Hydrobiologia, 2015, 761(1): 161-180. doi: 10.1007/s10750-015-2327-9
[42] VASSALLO P, PAOLI C, ALIANI S, et al. Benthic diversity patterns and predictors: a study case with inferences for conservation[J]. Mar Pollut Bull, 2020, 150: 110748. doi: 10.1016/j.marpolbul.2019.110748
[43] LAN K W, SHIMADA T, LEE M A, et al. Using remote-sensing environmental and fishery data to map potential yellowfin tuna habitats in the tropical Pacific Ocean[J]. Remote Sens-Basel, 2017, 9(5): 444. doi: 10.3390/rs9050444
[44] 杨胜龙, 张忭忭, 靳少非, 等. 中西太平洋延绳钓黄鳍金枪鱼渔场时空分布与温跃层关系[J]. 海洋学报, 2015, 37(6): 78-87. doi: http://en.cnki.com.cn/Article_en/CJFDTOTAL-SEAC201506008.htm [45] HOOLIHAN J, WELLS R, LUO J, et al. Vertical and horizontal movements of yellowfin tuna in the Gulf of Mexico[J]. Mar Coast Fish, 2014, 6(1): 211-222. doi: 10.1080/19425120.2014.935900
[46] LASCELLES B, NOTARBARTOLO DI SCIARA G, AGARDY T, et al. Migratory marine species: their status, threats and conservation management needs[J]. Aquat Conserv, 2014, 24(S2): 111-127. doi: 10.1002/aqc.2512
[47] DUERI S, MAURY O. Modelling the effect of marine protected areas on the population of skipjack tuna in the Indian Ocean[J]. Aquat Living Resour, 2013, 26(2): 171-178.
[48] GRSS A. Modelling the impacts of marine protected areas for mobile exploited fish populations and their fisheries: what we recently learnt and where we should be going[J]. Aquat Living Resour, 2015, 27(3/4): 107-133.
[49] CLARK M R, WATLING L, ROWDEN A A, et al. A global seamount classification to aid the scientific design of marine protected area networks[J]. Ocean Coast Manag, 2011, 54(1): 19-36. doi: 10.1016/j.ocecoaman.2010.10.006
[50] HOWELL K L. A benthic classification system to aid in the implementation of marine protected area networks in the deep/high seas of the NE Atlantic[J]. Biol Conserv, 2010, 143(5): 1041-1056.
[51] FREIWALD A. Cold-water corals and ecosystems[M]. Heidelberg: Springer Science & Business Media, 2005: 259-276.
[52] RAMOS A, SANTIAGO J, SANGRA P, et al. An application of satellite-derived sea surface temperature data to the skipjack (Katsuwonus pelamis Linnaeus, 1758) and albacore tuna (Thunnus alalunga Bonaterre, 1788) fisheries in the north-east Atlantic[J]. Int J Remote Sens, 1996, 17(4): 749-759. doi: 10.1080/01431169608949042
[53] FIEDLER P C, BERNARD H J. Tuna aggregation and feeding near fronts observed in satellite imagery[J]. Cont Shelf Res, 1987, 7(8): 871-881. doi: 10.1016/0278-4343(87)90003-3
[54] CLARK M R, SCHLACHER T A, ROWDEN A A, et al. Science priorities for seamounts: research links to conservation and management[J]. PLoS One, 2012, 7(1): e29232. doi: 10.1371/journal.pone.0029232
[55] BARCKHAUSEN U, ROESER H A, von HUENE R. Magnetic signature of upper plate structures and subducting seamounts at the convergent margin off Costa Rica[J]. J Geophys Res-Sol Ea, 1998, 103(B4): 7079-7093. doi: 10.1029/98JB00163
[56] WRIGHT S R, RIGHTON D, NAULAERTS J, et al. Fidelity of yellowfin tuna to seamount and island foraging grounds in the central South Atlantic Ocean[J]. Deep-Sea Res I, 2021, 172: 103513. doi: 10.1016/j.dsr.2021.103513
[57] LAM C H, TAM C, KOBAYASHI D R, et al. Complex dispersal of adult yellowfin tuna from the main Hawaiian islands[J]. Front Mar Sci, 2020, 7: 138. doi: 10.3389/fmars.2020.00138
[58] MONDAL S, RAY A, LEE M A, et al. Projected changes in spawning ground distribution of mature albacore tuna in the Indian Ocean under various global climate change scenarios[J]. J Mar Sci Eng, 2023, 11(8): 1565. doi: 10.3390/jmse11081565
[1] YESSON C, LETESSIER T, NIMMO-SMITH A, et al. Improved bathymetry leads to > 4 000 new seamount predictions in the global ocean-but beware of phantom seamounts [J]. UCL Press, 2021, 4: 1–9. [2] YESSON C, CLARK M R, TAYLOR M L, et al. The global distribution of seamounts based on 30 arc seconds bathymetry data [J]. Deep-Sea Res Ⅰ, 2011, 58 (4): 442–453. [3] ROGERS A D. The biology of seamounts: 25 years on [J]. Adv Mar Biol, 2018, 79: 137–224. [4] BO M, COPPARI M, BETTI F, et al. Unveiling the deep biodiversity of the Janua Seamount (Ligurian Sea): first Mediterranean sighting of the rare Atlantic bamboo coral Chelidonisis aurantiaca Studer, 1890 [J]. Deep-Sea Res Ⅰ, 2020, 156: 103186. [5] PITCHER T J, MORATO T, HART P J B, et al. Seamounts: ecology, fisheries & conservation [M]. Oxford: Blackwell Pub, 2007: 85–100, 442–475. [6] GONZLEZ-IRUSTA J M, DE LA TORRIENTE A, PUNZN A, et al. Living at the top. Connectivity limitations and summit depth drive fish diversity patterns in an isolated seamount [J]. Mar Ecol Prog Ser, 2021, 670: 121–137. [7] BRIDGES A E H, BARNES D K A, BELL J B, et al. Depth and latitudinal gradients of diversity in seamount benthic communities [J]. J Biogeogr, 2022, 49(5): 904–915. [8] MCCLAIN C R, LUNDSTEN L. Assemblage structure is related to slope and depth on a deep offshore Pacific seamount chain [J]. Mar Ecol, 2015, 36(2): 210–220. [9] HANN C H, SMITH T D, TORRES L G. A sperm whale's perspective: the importance of seasonality and seamount depth [J]. Mar Mammal Sci, 2016, 32(4): 1470–1481. [10] ROWDEN A A, SCHLACHER T A, WILLIAMS A, et al. A test of the seamount oasis hypothesis: seamounts support higher epibenthic megafaunal biomass than adjacent slopes [J]. Mar Ecol, 2010, 31(1): 95–106. [11] CAMPANELLA F, COLLINS M A, YOUNG E F, et al. First insight of meso-and bentho-pelagic fish dynamics around remote seamounts in the South Atlantic Ocean [J]. Front Mar Sci, 2021, 8: 693. [12] VASSALLO P, PAOLI C, ALESSI J, et al. Seamounts as hot-spots of large pelagic aggregations [J]. Mediterr Mar Sci, 2018, 19(3): 444–458. [13] KVILE K, TARANTO G H, PITCHER T J, et al. A global assessment of seamount ecosystems knowledge using an ecosystem evaluation framework [J]. Biol Conserv, 2014, 173: 108–120. [14] DEARY A L, MORET-FERGUSON S, ENGELS M, et al. Influence of central Pacific oceanographic conditions on the potential vertical habitat of four tropical tuna species 1 [J]. Pac Sci, 2015, 69(4): 461–475. [15] DUFFY L M, KUHNERT P M, PETHYBRIDGE H R, et al. Global trophic ecology of yellowfin, bigeye, and albacore tunas: understanding predation on micronekton communities at ocean-basin scales [J]. Deep-Sea Res Ⅱ, 2017, 140: 55–73. [16] DORTEL E, PECQUERIE L, CHASSOT E. A Dynamic Energy Budget simulation approach to investigate the eco-physiological factors behind the two-stanza growth of yellowfin tuna (Thunnus albacares) [J]. Ecol Model, 2020, 437: 109297. [17] PECORARO C, ZUDAIRE I, BODIN N, et al. Putting all the pieces together: integrating current knowledge of the biology, ecology, fisheries status, stock structure and management of yellowfin tuna (Thunnus albacares) [J]. Rev Fish Biol Fish, 2016, 27(4): 811–841. [18] SHI X F, WANG X, WANG Y X, et al. Feeding biology of yellowfin tuna (Thunnus albacares) in tropical central and western Pacific Ocean [J]. South China Fish Sci, 2022, 18(1): 43–51 (in Chinese) [19] SCHAEFER K M, FULLER D W, BLOCK B A. Movements, behavior, and habitat utilization of yellowfin tuna (Thunnus albacares) in the Pacific Ocean off Baja California, Mexico, determined from archival tag data analyses, including unscented Kalman filtering [J]. Fish Res, 2011, 112(1/2): 22–37. [20] WRIGHT S R, RIGHTON D, NAULAERTS J, et al. Yellowfin tuna behavioural ecology and catchability in the South Atlantic: the right place at the right time (and depth) [J]. Front Mar Sci, 2021, 8: 664593. [21] DAGORN L, HOLLAND K N, HALLIER J P, et al. Deep diving behavior observed in yellowfin tuna (Thunnus albacares) [J]. Aquat Living Resour, 2006, 19(1): 85–88. [22] CLARK M R, ROWDEN A A, SCHLACHER T, et al. The ecology of seamounts: structure, function, and human impacts [J]. Annu Rev Mar Sci, 2010, 2: 253–278. [23] PILLING G, SCOTT F, HAMPTON S. Minimum target reference points for WCPO yellowfin and bigeye tuna consistent with alternative LRP risk levels, and multispecies implications [R/OL]. [2030-09-10] Pohnpei: Technical Report WCPFC-SC15-2019/MI-WP-01, 2019. https://www.wcpfc.int/doc/wcpfc16-2019-15-update-sc15-mi-wp-01/minimum-target-reference-pointswcpo-yellowfin-and-bigeye.pdf [24] ZHAO R J, ZHAO F, FENG L, et al. A deep seamount effect enhanced the vertical connectivity of the planktonic community across 1, 000 m above summit [J]. J Geophys Res-Oceans, 2023, 128(3): e2022JC018898. [25] MORATO T, HOYLE S D, ALLAIN V, et al. Seamounts are hotspots of pelagic biodiversity in the open ocean [J]. P Natl Acad Sci USA, 2010, 107(21): 9707–9711. [26] MORATO T, VARKEY D A, DAMASO C, et al. Evidence of a seamount effect on aggregating visitors [J]. Mar Ecol Prog Ser, 2008, 357(4): 23–32. [27] MAUNDER M N, STARR P J. Fitting fisheries models to standardised CPUE abundance indices [J]. Fish Res, 2003, 63(1): 43–50. [28] PEDERSEN E J, MILLER D L, SIMPSON G L, et al. Hierarchical generalized additive models in ecology: an introduction with mgcv [J]. PeerJ, 2019, 7: e6876. [29] MARN-ENRQUEZ E, RAMREZ-PREZ J S, RUIZ-DOMNGUEZM, et al. Effect of marine climate and baitfish availability on the tuna baitboat fishery CPUE off northwestern Mexico [J]. Ocean Coast Manag, 2023, 232: 106418. [30] VAIHOLA S, KININMONTH S. Climate change potential impacts on the tuna fisheries in the exclusive economic zones of Tonga [J]. Diversity, 2023, 15(7): 844. [31] LEHODEY P, BERTRAND A, HOBDAY A J, et al. ENSO impact on marine fisheries and ecosystems [M]. Washington: Wiley American Geophysical Union, 2020: 429–451. [32] BO M, COPPARI M, BETTI F, et al. The high biodiversity and vulnerability of two Mediterranean bathyal seamounts support the need for creating offshore protected areas [J]. Aquat Conserv, 2021, 31(3): 543–566. [33] HANAFI-PORTIER M, SAMADI S, CORBARI L, et al. Multiscale spatial patterns and environmental drivers of seamount and island slope megafaunal assemblages along the Mozambique channel [J]. Deep-Sea Res I, 2024, 203: 104198. [34] PITCHER T J, MORATO T, HART P J B, et al. Seamounts: ecology, fisheries & conservation [M]. Oxford: Blackwell Pub, 2007: 238–239. [35] CLARK M R. Deep sea seamount fisheries: a review of global status and future prospects [J]. Lat Am J Aquat Res, 2009, 37(3): 501–512. [36] ITANO D. A summary of operational, technical and fishery information on WCPO purse seine fisheries operating on floating objects [R/OL]. [2023-09-10] Honolulu: Scientific Committee 3rd Regular Session of Western and Central Pacific Fisheries Commission, 2007. https://www.wcpfc.int/system/files/SC3_FT-IP-4_Itano%20FO_complete.pdf [37] PITCHER T J, MORATO T, HART P J B, et al. seamounts: ecology, fisheries & conservation [M]. Oxford: Blackwell Pub, 2007: 85–100. [38] PRECIADO I, CARTES J E, PUNZN A, et al. Food web functioning of the benthopelagic community in a deep-sea seamount based on diet and stable isotope analyses [J]. Deep-Sea Res I, 2017, 137: 56–68. [39] BRILL R W, LUTCAVAGE M E. Understanding environmental influences on movements and depth distributions of tunas and billfishes can significantly improve population assessments [C] // American Fisheries Society Symposium. Phoenix: American Fisheries Society, 2001: 179–198. [40] AUSCAVITCH S R, DEERE M C, KELLER A G, et al. Oceanographic drivers of deep-sea coral species distribution and community assembly on seamounts, islands, atolls, and reefs within the phoenix islands protected area [J]. Front Mar Sci, 2020, 7: 42. [41] CLARK M R, BOWDEN D A. Seamount biodiversity: high variability both within and between seamounts in the Ross Sea region of Antarctica [J]. Hydrobiologia, 2015, 761(1): 161–180. [42] VASSALLO P, PAOLI C, ALIANI S, et al. Benthic diversity patterns and predictors: a study case with inferences for conservation [J]. Mar Pollut Bull, 2020, 150: 110748. [43] LAN K W, SHIMADA T, LEE M A, et al. Using remote-sensing environmental and fishery data to map potential yellowfin tuna habitats in the tropical Pacific Ocean [J]. Remote Sens-Basel, 2017, 9(5): 444. [44] YANG S L, ZHANG B B, JIN S F, et al. Relationship between the temporal-spatial distribution of longline fishing grounds of yellowfin tuna (Thunnus albacares) and the thermocline characteristics in the Western and Central Pacific Ocean [J]. Acta Oceanologica Sinica, 2015, 37(6): 78–87 (in Chinese). [45] HOOLIHAN J, WELLS R, LUO J, et al. Vertical and horizontal movements of yellowfin tuna in the Gulf of Mexico [J]. Mar Coast Fish, 2014, 6(1): 211–222. [46] LASCELLES B, NOTARBARTOLO DI SCIARA G, AGARDY T, et al. Migratory marine species: their status, threats and conservation management needs [J]. Aquat Conserv, 2014, 24(S2): 111–127. [47] DUERI S, MAURY O. Modelling the effect of marine protected areas on the population of skipjack tuna in the Indian Ocean [J]. Aquat Living Resour, 2013, 26(2): 171–178. [48] GRSS A. Modelling the impacts of marine protected areas for mobile exploited fish populations and their fisheries: what we recently learnt and where we should be going [J]. Aquat Living Resour, 2015, 27(3/4): 107–133. [49] CLARK M R, WATLING L, ROWDEN A A, et al. A global seamount classification to aid the scientific design of marine protected area networks [J]. Ocean Coast Manag, 2011, 54(1): 19–36. [50] HOWELL K L. A benthic classification system to aid in the implementation of marine protected area networks in the deep/high seas of the NE Atlantic [J]. Biol Conserv, 2010, 143(5): 1041–1056. [51] FREIWALD A. Cold-water corals and ecosystems [M]. Heidelberg: Springer Science & Business Media, 2005: 259–276. [52] RAMOS A, SANTIAGO J, SANGRA P, et al. An application of satellite-derived sea surface temperature data to the skipjack (Katsuwonus pelamis Linnaeus, 1758) and albacore tuna (Thunnus alalunga Bonaterre, 1788) fisheries in the north-east Atlantic [J]. Int J Remote Sens, 1996, 17(4): 749–759. [53] FIEDLER P C, BERNARD H J. Tuna aggregation and feeding near fronts observed in satellite imagery [J]. Cont Shelf Res, 1987, 7(8): 871–881. [54] CLARK M R, SCHLACHER T A, ROWDEN A A, et al. Science priorities for seamounts: research links to conservation and management [J]. PLoS One, 2012, 7(1): e29232. [55] BARCKHAUSEN U, ROESER H A, von HUENE R. Magnetic signature of upper plate structures and subducting seamounts at the convergent margin off Costa Rica [J]. J Geophys Res-Sol Ea, 1998, 103(B4): 7079–7093. [56] WRIGHT S R, RIGHTON D, NAULAERTS J, et al. Fidelity of yellowfin tuna to seamount and island foraging grounds in the central South Atlantic Ocean [J]. Deep-Sea Res I, 2021, 172: 103513. [57] LAM C H, TAM C, KOBAYASHI D R, et al. Complex dispersal of adult yellowfin tuna from the main Hawaiian islands [J]. Front Mar Sci, 2020, 7: 138. [58] MONDAL S, RAY A, LEE M A, et al. Projected changes in spawning ground distribution of mature albacore tuna in the Indian Ocean under various global climate change scenarios [J]. J Mar Sci Eng, 2023, 11(8): 1565. -
期刊类型引用(2)
1. 李东旭,邹晓荣,周淑婷. 中太平洋黄鳍金枪鱼CPUE时空分布及其与环境因子的关系. 南方水产科学. 2024(04): 68-76 .  本站查看
本站查看
2. 程高,陈国宝,陈丕茂,佟飞,牛麓连,陈钰祥. 基于声学技术定点监测海洋牧场鱼类资源昼夜变化研究. 南方水产科学. 2024(05): 63-70 . 本站查看
其他类型引用(0)






计量
- 文章访问数: 246
- HTML全文浏览量: 56
- PDF下载量: 69
- 被引次数: 2
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
