
Biological characteristics of Ctenopharyngodon idellus in middle and lower reaches of Pearl River
-
摘要: 根据2019—2020年珠江中下游草鱼 (Ctenopharyngodon idellus) 种群调查数据,对其群体结构和生长现状进行了分析。结果显示,草鱼种群个体体长介于103~665 mm,平均体长为 (347.89±96.73) mm;体质量介于21.4~6 650.0 g,平均体质量为 (986.87±842.96) g。草鱼种群年龄结构为0+~5+龄,优势年龄组为1+、2+和3+龄,占样本总数的92.8%。草鱼体长 (L) 和体质量 (W) 呈显著幂函数关系,为匀速生长类型。Von Bertalanffy生长方程参数:L∞=1187 mm,k=0.128,t0=−0.282,W∞=31 344.51 g,生长特征指数φ=5.26,生长拐点年龄为8.29。总死亡系数Z=0.58,自然死亡系数M=0.29,捕捞死亡系数F=0.29,开发率E=0.5。草鱼雌雄个体数的比例为1.07∶1,性腺成熟度以Ⅱ期为主,相对繁殖力为46.28 粒·g−1。与历史资料相比发现,草鱼生长性能下降,种群呈现小型化,繁殖力下降,为促进珠江中下游草鱼资源的恢复,建议降低捕捞强度,开捕体长应大于791 mm,体质量应大于9 279.11 g。Abstract: In order to find out the current status of the population structure and growth characteristics of grass carp (Ctenopharyngodon idellus) in the middle and lower reaches of the Pearl River, we have investigated the grass carp population from 2019 to 2020. The results show that the body length of grass carp was 103−665 mm [average: (347.89±96.73) mm]. The body mass was 21.4−6 650.0 g [average: (986.87±842.96) g]. The population age was of 0+−5+, and the dominant age was 1+−3+, accounting for 92.8% of the total number of samples. The body length had a significant power function relation with body mass for uniform motion pattern. The growth could be described by Von Bertalanffy equation with the growth parameters: L∞=1187 mm, k=0.128, t0=−0.282, W∞=31 344.51 g. The index of length growth performance (φ) of grass carp was 5.26, and the growth inflection age was 8.29. The total mortality rate, natural mortality rate, fishing mortality rate and exploitation rate were 0.58, 0.29, 0.29 and 0.5, respectively. The sex ratio (Females/Males) was 1.07: 1. The sexual maturation was mainly at Stage II. Compared with the previous data, the growth performance and fecundity of grass carp all showed a decline tendency; the population structure tended to be miniaturized. In order to promote the recovery of grass carp resources in the middle and lower reaches of the Pearl River, it is recommended to reduce the fishing intensity. Besides, the catch body length should be greater than 791 mm and the mass should be greater than 9 279.11 g.
-
珠江是我国南方最大的河流,渔业资源十分丰富,其适宜的气候和水文条件为四大家鱼提供了重要的自然栖息地[1-2],19世纪70年代到80年代初期珠江水系四大家鱼年均渔获量达12 961.5 t[3]。然而近几十年来,珠江流域由于水坝及航道工程建设、环境污染和过度捕捞等因素影响,鱼类洄游通道受阻、产卵场功能丧失,渔业资源呈急剧衰退趋势[4-7]。为了遏制这一趋势,相关渔业管理部门实施了增殖放流、禁渔和打击非法捕捞等资源养护措施,草鱼 (Ctenopharyngodon idellus) 资源量有所恢复。
草鱼生长迅速且分布范围广,是我国重要的经济鱼类,也是珠江主要的渔业对象之一,其种群特征及资源状况一直备受关注。20世纪80年代,珠江水产研究所联合有关单位对珠江水系渔业资源进行了系统、全面地调查,对草鱼生长、繁殖、摄食等生物学特征进行了研究[8]。一些学者对国内各水系草鱼自然种群的生长特征做了比较研究[9-10]。近年来,珠江草鱼自然种群研究主要集中在空间分布和早期资源方面[2,7,11],现状条件下有关珠江草鱼自然种群结构和生长特征方面的研究还未见报道。鱼类年龄与生长研究是开展渔业资源评估、科学管理和可持续利用的基础[12],草鱼种群结构研究现状的缺失制约了其保护进程。因此本研究于2019—2020年对珠江中下游草鱼种群的渔业生物学特征进行了初步分析,以期为珠江草鱼资源恢复和养护提供参考依据。
1. 材料与方法
1.1 样本采集
2019年7月和2020年1、7、8月在珠江中下游的桂平、藤县、梧州、封开、德庆、肇庆、九江和斗门站位 (图1) 开展草鱼资源调查,草鱼样本由各站位当地渔民捕获,捕捞网具为流刺网、抛网和张网,共采集到样本236尾。将采集到的样本进行编号,在新鲜状态下测量体长 (L) 和体质量 (W),并进行解剖观察性腺发育情况。体长精确到1 mm,体质量精确到0.1 g。优势体长和体质量范围依照正态分布的特征,取样本范围“平均数±标准差”为优势体长和体质量的分布。在鱼体的背鳍起点下方至侧线鳞上方的中间区域取5~8枚形状完整、轮纹清晰的鳞片,装入鳞片袋并做好标记,用于年龄鉴定。根据各年龄组样本量所占频率,取前3位为优势年龄组。
1.2 年龄鉴定
将鳞片置于1%氢氧化钾 (KOH) 溶液中浸泡12 h,洗去表面黏液,清水冲洗后用吸水纸除去水分。鳞片用载玻片装片,使用奥林巴斯 (SZX16) 体视镜观察年轮特征并进行拍照。通过多次鉴定确定年龄并测量鳞径 (R) 和轮径 (r),测量位置固定在鳞片的前、侧区交界处的鳞肩部位,草鱼鳞片形态和年轮特征见图2。鳞片还未形成第1个年轮的,用0+表示;已形成第1个年轮的,用1+表示;依此类推,如2+,3+,······。
![]() 图 2 草鱼鳞片形态和年轮特征数字表示年轮,F表示鳞焦,箭头指示为幼轮。Figure 2. Scale and annuli characteristics of C. idellusThe numbers represent the annuli; F represents the focus; the arrow indicates the fry check.
图 2 草鱼鳞片形态和年轮特征数字表示年轮,F表示鳞焦,箭头指示为幼轮。Figure 2. Scale and annuli characteristics of C. idellusThe numbers represent the annuli; F represents the focus; the arrow indicates the fry check.1.3 生长特征分析
体长-体质量关系采用Keys公式进行拟合:
$$ {W=a}{{L}}^{{b}} $$ (1) 式中:a为常数;b为生长指数。将b与3进行差异显著性检验,差异不显著时,鱼类匀速生长;差异显著时,为异速生长[13-14]。
体长-鳞径关系采用Rasa Lee方程进行拟合[12, 14]:
$$ {L=a}+{bR} $$ (2) 式中:a为体长与鳞径关系直线在y轴的截距;b为直线斜率,并对样本体长进行推算。
鱼类的生长指数计算参考文献[13],绝对增长率 (或增重率):
$ {{L}}_{{2}}-{{L}}_{{1}} $ 或$ {{W}}_{{2}}-{{W}}_{{1}} $ ;相对增长率:$ ({{L}}_{{2}}- $ $ {{L}}_{{1}}){/}{{L}}_{{1}} $ 或$ \left({{W}}_{{2}}-{{W}}_{{1}}\right){/}{{W}}_{{1}} $ ;瞬时增长率:$ {\ln}{{L}}_{{2}}- {\ln}{{L}}_{{1}}\; $ 或$ {\ln}{{W}}_{{2}}- {\ln}{{W}}_{{1}} $ ;生长指标:$ {{C}}_{{{\rm{e}}}}{=}\left({{{\rm{lg}}}}{{L}}_{{2}}-{{{\rm{lg}}}}{{L}}_{{1}}\right)/ {0.434}\;{3} ({{t}}_{{2}}- $ $ {{t}}_{{1}} ){{L}}_{{1}} $ ,式中:L2、L1和W2、W1分别为t2、t1时对应的体长和体质量。利用Von Bertalanffy生长方程描述草鱼的生长规律[13]:
$${L_t} = {L_\infty }\left[ {1 - {{\rm{e}}^{ - k\left( {t - {t_0}} \right)}}} \right],\;{W_t} = {W_\infty }{\left[ {1 - {{\rm{e}}^{ - k\left( {t - {t_0}} \right)}}} \right]^b}$$ (3) 式中:Lt、Wt为t龄时的体长和体质量;L∞、W∞为渐近体长和渐近体质量,k为生长系数,t0为理论生长起点年龄,b为生长指数。
生长拐点年龄计算公式为:
$ {{t}}_{\rm{i}}{=}{\ln}{b}{/}{k}{+}{{t}}_{{0}} $ ;生长特征指数计算公式[15]为:$\varphi = {{\rm{lg}}}k + 2{{\rm{lg}} }{L_\infty }$ 。对Von Bertalanffy生长方程求导变换,得到体长和体质量的生长速度 ($ {{\rm{d}}L}/{{\rm{d}}t} $ ,$ {{\rm{d}}W}/{{\rm{d}}t} $ )及加速度 ($ {{{\rm{d}}}}^{{2}}{L}/{{\rm{d}}}{{t}}^{{2}} $ ,$ {{{\rm{d}}}}^{{2}}{W}/{{\rm{d}}}{{t}}^{{2}} $ )方程。根据Beverton-Holt模型[16],用平均体长估算总死亡系数:
$ {Z}{=}{k}\left({{l}}_{\infty}-{l}\right){/}\left({l}-{{l}}{{\text{'}}}\right) $ ,${{l}}{{\text{'}}}\ $ 为起捕年龄渔获平均体长,l为渔获平均体长。Pauly认为鱼类的自然死亡不仅与其渐进体长有关,还与温度有关,因此他根据175种鱼类资源群体的资料得出了Pauly经验公式[17-18]:$ {\ln}{M}{=}-{0.015}\;{2}-{0.279}{\ln}{{L}}_{\infty}{+0.654}\;{3}{\ln}{k}+ $ $ {0.463}{\ln}{T} $ ,用于估算草鱼自然死亡系数M,其中年平均水温T取23.48 ℃,为实测水温数据。捕捞死亡系数$ F= Z- M $ ,开发率$ E=F/Z $ 。1.4 性比和性腺发育
统计草鱼样本的雌性和雄性个体数,性腺成熟度按照6级目测等级法判断,根据雌雄个体性腺发育情况,性腺发育达到Ⅳ期及以上为性成熟个体[12]。性腺成熟系数 (Gonadosomatic index, GSI) 计算公式为:
${\rm{GSI}} = (G_{\rm{W}}/E_{\rm{W}}) \times 100{\text{%}}$ ,式中:GW为性腺质量;EW为去内脏后的体质量。绝对繁殖力 (Absolute fecundity, FA)和相对繁殖力 (Relative fecundity, FR)计算公式:$F_{\rm{A}} = n \times \left( G_{{\rm{W}}} /w\right)$ ;$ F_{{\rm{R}}}=F_{{\rm{A}}}/{W} $ 。式中:n代表计数的卵粒数;GW代表卵巢质量;w代表卵巢子样质量;W代表体质量[12,19]。数据的整理、分析使用Excel 2010和R 4.0.0软件完成。
2. 结果
2.1 体长、体质量与年龄分布
草鱼个体体长、体质量频率分布情况如图3所示,其体长介于103~665 mm,平均体长为 (347.89±96.73) mm,优势体长为251~445 mm,占样本总数的68.22%。体质量介于21.4~6 650.0 g,平均体质量为 (986.87±842.96) g,优势体质量为144~1 830 g,占样本总数的79.24%。年龄鉴定结果见表1,样本的年龄组成为0+~5+龄,优势年龄组为1+~3+龄,占样本总数的92.8%,高龄鱼较少,平均体长分别为 (266.80±45.67)、(350.98±40.50)和 (446.83±41.97) mm;平均体质量分别为 (384.99±183.12)、(848.34±258.26) 和 (1 728.19±541.45) g。
![]() 图 3 珠江中下游草鱼体长、体质量频率分布Figure 3. Frequency distribution of C. idellus body length and body mass in middle and lower reaches of Pearl River表 1 珠江中下游草鱼种群年龄结构Table 1. Age structure of C. idellus populations in middle and lower reaches of Pearl River
图 3 珠江中下游草鱼体长、体质量频率分布Figure 3. Frequency distribution of C. idellus body length and body mass in middle and lower reaches of Pearl River表 1 珠江中下游草鱼种群年龄结构Table 1. Age structure of C. idellus populations in middle and lower reaches of Pearl River年龄
Age平均体长
Average body length/mm范围
Range/mm平均体质量
Average body mass/g范围
Range/g数量百分比
Quantity number/%0+ 161.22±30.74 103~214 81.63±36.10 21.4~149.7 3.8 1+ 266.80±45.67 172~360 384.99±183.12 92.8~810.0 32.2 2+ 350.98±40.50 262~440 848.34±258.26 310.2~1 396.0 35.6 3+ 446.83±41.97 362~541 1 728.19±541.45 829.3~3 361.6 25.0 4+ 536.00±41.14 505~600 2 996.98±718.81 2 317.2~4 200.0 2.1 5+ 616.33±50.05 565~665 4 900.17±1 543.82 3 730.4~6 650.0 1.3 2.2 体长与体质量的关系
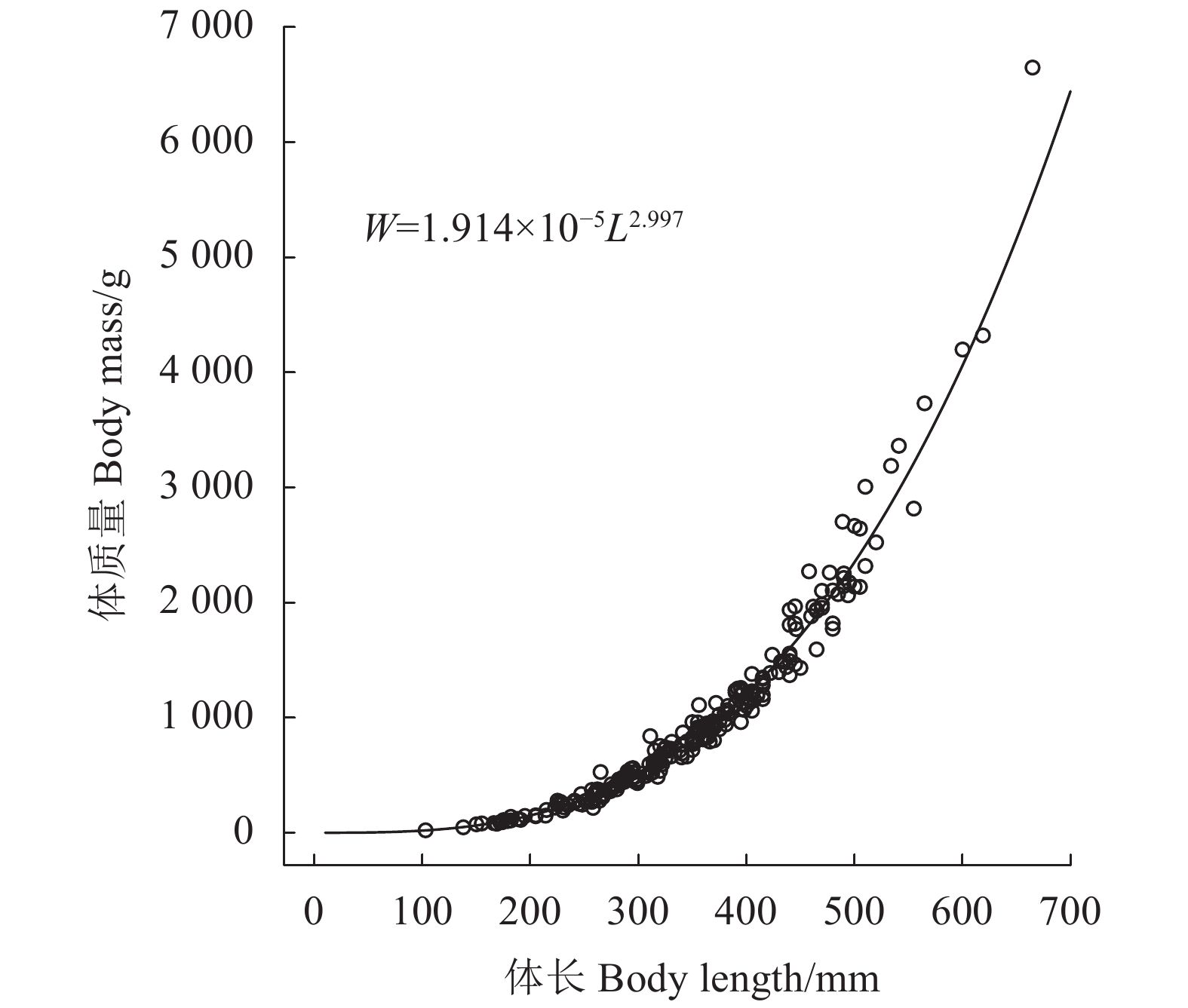
珠江中下游草鱼体长和体质量关系拟合结果为
$W = 1.914\;4 \times {10^{ - 5}}{L^{2.997}}$ (N=236, R2=0.99, P<0.01) (图4)。差异显著性分析结果显示,b值与3之间无显著差异 (P>0.01),表明珠江中下游草鱼为匀速生长类型。![]() 图 4 珠江中下游草鱼体长与体质量的关系Figure 4. Relationship between body length and mass of C. idellus in middle and lower reaches of Pearl River
图 4 珠江中下游草鱼体长与体质量的关系Figure 4. Relationship between body length and mass of C. idellus in middle and lower reaches of Pearl River2.3 体长与鳞径的关系及生长推算
珠江中下游草鱼体长与鳞径(R)呈线性相关:
$ {L=} $ $ {42.540}\;{9}{R}-{0.407}\;{4} $ (N=263, R2=0.92)。根据Rasa Lee方程计算出1~5龄草鱼的推算体长分别为180.35、300.37、404.21、505.86和581.31 mm,再由体长-体质量关系,计算出对应的推算体质量分别为110.56、510.02、1 241.72、2 432.34 和3 689.45 g。根据各龄草鱼推算体长、体质量计算草鱼的生长指数 (表2)。表 2 珠江中下游草鱼体长、体质量生长指数Table 2. Back calculated standard length of C. idellus groups in middle and lower reaches of Pearl River年龄
Age推算体长
Calculated
body length/
mm年增长量
Annual growth/
mm相对生长率
Relative
growth
rate瞬时生长率
Instantaneous
growth rate生长指标
Index of
growth推算体质量
Calculated
body mass/g年增长量
Annual growth/g相对生长率
Relative
growth
rate瞬时生长率
Instantaneous
growth rate推算体长*
Calculated
body length/
mm1 180.35 — — — — 110.56 — — — 193.0 2 300.37 120.02 0.67 0.51 92.00 510.02 399.46 3.61 1.53 349.0 3 404.21 103.84 0.35 0.30 89.18 1 241.72 731.70 1.43 0.89 471.0 4 505.86 101.65 0.25 0.22 90.67 2 432.34 1 190.62 0.96 0.67 547.0 5 581.31 75.45 0.15 0.14 70.33 3 689.45 1 257.11 0.52 0.42 598.0 注:*. 20世纪80年代。 Note: *. 1980s. 2.4 生长方程
通过Von Bertalanffy生长方程拟合草鱼生长规律,得到方程参数L∞=1187 mm,W∞=31344.51 g,k=0.128,t0=−0.282,生长特征指数φ=5.26。将各参数代入公式中,分别得到体长、体质量生长方程:
$$\begin{split} &\qquad \qquad \quad {L_t} = 1\;187\left[ {1 - {{\rm{e}}^{ - 0.128\; \left( {t + 0.282} \right)}}} \right];\\ &\qquad \qquad{W_t} = 31\;344.51{\left[ {1 - {{\rm{e}}^{ - 0.128\; \left( {t + 0.282} \right)}}} \right]^{2.997}} \end{split}$$ (4) 体长、体质量生长曲线见图5,草鱼体长生长曲线不具拐点,生长速度随年龄的增加而降低,体长随着年龄的增加趋近于一个固定值;体质量生长曲线呈S形,具有拐点。
![]() 图 5 珠江中下游草鱼体长和体质量的生长方程曲线Figure 5. Growth curve of C. idellus body length and body mass in middle and lower reaches of Pearl River
图 5 珠江中下游草鱼体长和体质量的生长方程曲线Figure 5. Growth curve of C. idellus body length and body mass in middle and lower reaches of Pearl River将草鱼体长、体质量生长方程分别对年龄t进行一阶和二阶求导得到体长、体质量的生长速度和加速度方程:
$$ {\text{体长生长速度:}}{\rm{d}}L/{\rm{d}}t = 151.94{{\rm{e}}^{ - 0.128\;\left( {t + 0.282} \right)}} $$ (5) $$ \qquad {\text{体长生长加速度:}}{{\rm{d}}^2}L/{\rm{d}}{t^2} = - 19.45{{\rm{e}}^{ - 0.128\;\left( {t + 0.282} \right)}};$$ $$ \begin{split} &\qquad {\text{体质量生长速度:}}\\ &\quad {\rm{d}}W/{\rm{d}}t = 10\;204.26{{\rm{e}}^{ - 0.128\;\left( {t + 0.282} \right)}}{\left[ {1 - {{\rm{e}}^{ - 0.121 \; \left( {t + 0.282} \right)}}} \right]^{1.997}} \end{split} $$ (6) $$\begin{split} &\qquad {\text{体质量生长加速度:}}\\ &\quad{{\rm{d}}^2}W/{\rm{d}}{t^2} = 1\;539.11{{\rm{e}}^{ - 0.128 \; \left( {t + 0.282} \right)}}{\left[ {1 - {{\rm{e}}^{ - 0.128 \; \left( {t + 0.282} \right)}}} \right]^{0.997}}\\ &\qquad\qquad\qquad \left[ {2.997{{\rm{e}}^{ - 0.128 \; \left( {t + 0.282} \right)}} - 1} \right] \end{split}$$ (7) 根据上述方程拟合出生长速度与加速度曲线 (图6)。草鱼体长生长速度随着年龄的增加而降低,最后趋近于0;生长加速度始终小于0,但体长生长下降速度随年龄的增加而减小。草鱼体质量生长具有拐点,拐点年龄为8.29,对应的体长和体质量分别为790.78 mm和9279.11 g。拐点之前生长速度逐渐增加,生长加速度先增加后降低;拐点之后生长速度逐渐降低,生长加速度先降低后略微呈现上升趋势。
![]() 图 6 珠江中下游草鱼体长、体质量生长速度与加速度曲线Figure 6. Curves of body length, body mass growth rate and acceleration of C. idellus in middle and lower reaches of Pearl River
图 6 珠江中下游草鱼体长、体质量生长速度与加速度曲线Figure 6. Curves of body length, body mass growth rate and acceleration of C. idellus in middle and lower reaches of Pearl River2.5 死亡系数
根据B-H模型,利用平均体长估算总死亡系数Z=0.58。根据Pauly经验公式估算自然死亡系数M=0.29,捕捞死亡系数F=Z−M=0.29,开发率E=F/Z=0.5。
2.6 性比与性腺成熟度
本研究共对197尾草鱼进行性别鉴定,在可辨认性别的个体中,雌性72尾,雄性67尾,雌、雄个体数的比例为1.07∶1,接近1∶1。性腺成熟度分析发现,采集的草鱼样本性腺成熟度以Ⅱ期 (68.53%) 为主,其次是Ⅰ期 (29.44%)。Ⅳ期和Ⅴ期分别占比1.52%和0.51%,未见Ⅲ期和Ⅵ期个体。性腺成熟系数 (GSI) 分析结果显示Ⅰ期为0.06,Ⅱ期雌性0.33、雄性0.09,Ⅳ期雌性6.73、雄性0.38,Ⅴ期雌性12.6。本研究性成熟草鱼仅7月和8月在梧州和桂平采集到1尾雄性和3尾雌性,年龄分别为4+和5+。3尾性成熟雌性草鱼绝对繁殖力分别为287 057、113 635和107 300粒,相对繁殖力分别为54.00、31.82和53.03粒·g−1,卵径为1.65 mm。
3. 讨论
3.1 草鱼年龄组成
20世纪80年代调查结果显示,珠江草鱼种群体长介于156~702 mm,年龄结构为0+~5+龄,西江-珠江三角洲优势年龄组为0+~2+龄,个体数量占比78%;广西江段优势年龄组为1+~3+龄,占88%以上[8]。本研究在珠江中下游采集的草鱼体长介于103~665 mm,年龄结构为0+~5+龄,优势年龄组为1+~3+龄,个体数量占比为92.8%。与20世纪80年代相比,采集的草鱼样本年龄和体长范围变化不大;但种群各龄组对应的推算体长和体质量均有所减小 (表1),表明草鱼生长性能下降。与其他水域近年调查相比[20-21],珠江中下游草鱼年龄结构相对简单,未见5+龄以上个体,且低龄鱼个体占绝对优势,表明珠江草鱼处于过度开发状态,需要加大其资源保护力度。
3.2 草鱼生长性能
鱼类体长-体质量关系常用幂函数 (W=aLb)方程拟合,b通常为2.5~3.5[22]。b的变化与鱼类的生长和营养有关,其差异主要受环境 (栖息地、季节)和生物学 (摄食、性别、生长期、胃饱满度)因素影响[23]。本研究中,珠江中下游草鱼的b为2.997,与3差异不显著,表现为匀速生长类型;与20世纪80年代调查资料的b (2.84) 存在显著差异,这可能与其栖息环境变化有关,与80年代比较,珠江中下游水域在水体理化因子、饵料生物资源等方面都发生了巨大变化,这些变化会影响鱼类的生长。生长参数 (L∞、k和t0) 是用于评估鱼类资源开发状况的动态综合模型的基础数据,这些参数有助于比较不同物种或同一物种在不同时间和地点的生长情况[24]。研究表明L∞和k之间存在负相关关系,L∞越大,k就越小[25],生长特征指数φ综合了L∞和k的效应[26]。将本研究与其他研究结果进行比较可以看出 (表3),20世纪80年代的长江中上游水域φ最大,表明其水域环境最有利于草鱼的生长,20世纪80年代的珠江水域次之,黑龙江和博斯腾湖水域最低。鱼类的生长受水温、饵料资源、捕捞等生存环境状况的影响[21],黑龙江和博斯腾湖水体温度相较于其他流域较低,可能是限制草鱼生长的主要因素之一。珠江中下游草鱼生长特征指数低于20世纪80年代,说明目前草鱼的生存环境受到胁迫,生长性能有所下降,这与河流开发和过度捕捞有关。
表 3 不同水域或时期草鱼种群生长参数比较Table 3. Comparison of growth parameters of C. idellus populations in different waters or periods水域
Site生长指数
b拐点年龄
ti生长系数
k渐进体长
L∞/mm生长特征指数
φ珠江中下游 (本研究) Middle and lower reaches of the Pearl River (this study) 2.997 8.29 0.128 1187 5.26 珠江 (1981—1983)[8] Pearl River 2.84 4.75 0.19 1069 5.34 珠江 (1983—1987)[9] Pearl River 2.86 8.00 0.13 1367 5.39 长江湖口 (1983—1987)[9] Yangtze Hukou 2.73 5.70 0.19 1215 5.44 长江中上游 (1981—1984)[10] Upper and middle Yangtze River 2.75 3.23 0.33 959 5.48 长江中上游 (1996—1999)[27] Upper and middle Yangtze River 2.87 5.38 0.19 1180 5.42 长江上游江津 (2008—2010)[21] Jiangjin, upper Yangtze River 2.82 6.28 0.17 1029 5.25 梁子湖 (1997—1999)[28] Liangzi Lake 2.89 4.66 0.22 1065 5.39 黑龙江 (1983—1987)[9] Heilongjiang River 3.05 7.30 0.16 976 5.18 博斯腾湖 (2014)[20] Bositeng Lake 2.83 7.50 0.14 1038 3.16 3.3 草鱼的繁殖特征
鱼类性腺发育的主要影响因素包括自身激素水平和环境因子 (如水温、光照、降雨和水流、盐度、饵料以及产卵基质等)[29-30]。珠江草鱼性成熟年龄一般为3龄[8],长江原种草鱼雄性比雌性普遍早熟1年,雄性初次性成熟年龄为3龄,最大性成熟年龄为4龄;雌性初次性成熟年龄为4龄,最大性成熟年龄为5龄[31]。本研究采集的雄性性成熟年龄为4+龄,雌性性成熟年龄为4+龄和5+龄。此次调查草鱼相对繁殖力平均为46.28粒·g−1,与20世纪80年代的调查结果相比 (82.66粒·g−1),草鱼的繁殖能力衰退。草鱼是一种江河半洄游性鱼类,平时分散栖息于中下游及支流水体中育肥,繁殖季节集群到产卵场产卵[8]。20世纪80年代调查显示,西江最大的草鱼产卵场是广西桂平的东塔产卵场,广东境内未发现产卵场[8]。本调查只在桂平和梧州采集到性成熟草鱼,在封开以下未采集到性成熟个体,这与20世纪80年代的调查结果相一致[8]。草鱼繁殖季节一般集中在4月底至6月初,7月以后则显著减少[8],由于珠江禁渔期为3月1日至6月30日[32],所以未采集该时间段内的样本。采集的草鱼样本中性成熟个体仅占2%,也说明草鱼自然繁殖的补充群体资源不足。近年来,珠江流域广泛开展了增殖流放活动,对缓解草鱼资源衰退起到了积极作用。但仍需要开展有效保护措施,以提高性成熟草鱼个体在自然种群中的比例,增加自然繁殖的幼鱼补充群体。
3.4 珠江中下游草鱼种群保护
本研究中,珠江中下游草鱼开发率为0.5。鱼类种群的最适开发率为0.5,若开发率大于0.5,则该种群处于过度捕捞状态[18,33-34]。Wang等[35]认为,开发率为0.5的评价标准适用于温带鱼类,而热带、亚热带水域鱼类由于生命周期短,生长迅速,可以承受较大的开发率。珠江中下游属亚热带气候区,因此该水域草鱼种群可能尚未达到最适开发水平。然而,本研究中草鱼种群达到性成熟年龄及以上的个体占28.4%,4+龄以上个体仅占3.4%,说明目前草鱼种群仍面临较大捕捞压力,捕捞结构不合理,不利于草鱼种群的保护。本研究中的珠江中下游草鱼,生长年龄拐点为8.29龄,表明目前草鱼种群处于过度捕捞状态。而历史调查资料中草鱼的生长拐点为4.75龄,对应体长和体质量为713 mm和6661 g。这种变化可能与其种质退化及饵料生物匮乏有关,草鱼是江河洄游性、植食性鱼类,珠江水系梯级开发程度大,水坝建设阻碍了其上下游种群基因交流,导致其种质退化;另外,由于航道整治、无序挖沙、河岸硬化等人类活动破坏水生植物,致使其食物资源匮乏,导致草鱼生长速度下降。鱼类的生长拐点年龄可作为渔业生产中开捕规格的参考依据[13,36]。因此,建议珠江中下游草鱼适宜捕捞年龄为8龄以上,有利于改善草鱼种群的年龄结构和增加种群中性成熟个体的数量。综上所述,在珠江中下游实施禁渔期和增殖放流的同时,还应规范渔具渔法,降低捕捞强度,确保低龄鱼和补充群体的数量,以促进珠江中下游草鱼资源的恢复。
-
![]()
图 2 草鱼鳞片形态和年轮特征
数字表示年轮,F表示鳞焦,箭头指示为幼轮。
Figure 2. Scale and annuli characteristics of C. idellus
The numbers represent the annuli; F represents the focus; the arrow indicates the fry check.
![]()
图 3 珠江中下游草鱼体长、体质量频率分布
Figure 3. Frequency distribution of C. idellus body length and body mass in middle and lower reaches of Pearl River
![]()
图 4 珠江中下游草鱼体长与体质量的关系
Figure 4. Relationship between body length and mass of C. idellus in middle and lower reaches of Pearl River
![]()
图 5 珠江中下游草鱼体长和体质量的生长方程曲线
Figure 5. Growth curve of C. idellus body length and body mass in middle and lower reaches of Pearl River
![]()
图 6 珠江中下游草鱼体长、体质量生长速度与加速度曲线
Figure 6. Curves of body length, body mass growth rate and acceleration of C. idellus in middle and lower reaches of Pearl River
表 1 珠江中下游草鱼种群年龄结构
Table 1 Age structure of C. idellus populations in middle and lower reaches of Pearl River
年龄
Age平均体长
Average body length/mm范围
Range/mm平均体质量
Average body mass/g范围
Range/g数量百分比
Quantity number/%0+ 161.22±30.74 103~214 81.63±36.10 21.4~149.7 3.8 1+ 266.80±45.67 172~360 384.99±183.12 92.8~810.0 32.2 2+ 350.98±40.50 262~440 848.34±258.26 310.2~1 396.0 35.6 3+ 446.83±41.97 362~541 1 728.19±541.45 829.3~3 361.6 25.0 4+ 536.00±41.14 505~600 2 996.98±718.81 2 317.2~4 200.0 2.1 5+ 616.33±50.05 565~665 4 900.17±1 543.82 3 730.4~6 650.0 1.3  下载: 导出CSV
下载: 导出CSV
表 2 珠江中下游草鱼体长、体质量生长指数
Table 2 Back calculated standard length of C. idellus groups in middle and lower reaches of Pearl River
年龄
Age推算体长
Calculated
body length/
mm年增长量
Annual growth/
mm相对生长率
Relative
growth
rate瞬时生长率
Instantaneous
growth rate生长指标
Index of
growth推算体质量
Calculated
body mass/g年增长量
Annual growth/g相对生长率
Relative
growth
rate瞬时生长率
Instantaneous
growth rate推算体长*
Calculated
body length/
mm1 180.35 — — — — 110.56 — — — 193.0 2 300.37 120.02 0.67 0.51 92.00 510.02 399.46 3.61 1.53 349.0 3 404.21 103.84 0.35 0.30 89.18 1 241.72 731.70 1.43 0.89 471.0 4 505.86 101.65 0.25 0.22 90.67 2 432.34 1 190.62 0.96 0.67 547.0 5 581.31 75.45 0.15 0.14 70.33 3 689.45 1 257.11 0.52 0.42 598.0 注:*. 20世纪80年代。 Note: *. 1980s.
下载: 导出CSV
表 3 不同水域或时期草鱼种群生长参数比较
Table 3 Comparison of growth parameters of C. idellus populations in different waters or periods
水域
Site生长指数
b拐点年龄
ti生长系数
k渐进体长
L∞/mm生长特征指数
φ珠江中下游 (本研究) Middle and lower reaches of the Pearl River (this study) 2.997 8.29 0.128 1187 5.26 珠江 (1981—1983)[8] Pearl River 2.84 4.75 0.19 1069 5.34 珠江 (1983—1987)[9] Pearl River 2.86 8.00 0.13 1367 5.39 长江湖口 (1983—1987)[9] Yangtze Hukou 2.73 5.70 0.19 1215 5.44 长江中上游 (1981—1984)[10] Upper and middle Yangtze River 2.75 3.23 0.33 959 5.48 长江中上游 (1996—1999)[27] Upper and middle Yangtze River 2.87 5.38 0.19 1180 5.42 长江上游江津 (2008—2010)[21] Jiangjin, upper Yangtze River 2.82 6.28 0.17 1029 5.25 梁子湖 (1997—1999)[28] Liangzi Lake 2.89 4.66 0.22 1065 5.39 黑龙江 (1983—1987)[9] Heilongjiang River 3.05 7.30 0.16 976 5.18 博斯腾湖 (2014)[20] Bositeng Lake 2.83 7.50 0.14 1038 3.16
下载: 导出CSV
-
[1] 帅方敏, 李新辉, 黄艳飞, 等. 珠江水系四大家鱼资源现状及空间分布特征研究[J]. 水生生物学报, 2017, 41(6): 1336-1344. doi: 10.7541/2017.165 [2] 谭细畅, 李跃飞, 赖子尼, 等. 西江肇庆段鱼苗群落结构组成及其周年变化研究[J]. 水生态学杂志, 2010, 31(5): 27-31. [3] 李思发, 陆伟民, 周碧云, 等. 长江、珠江、黑龙江鲢、鳙、草鱼渔业资源状况[J]. 淡水渔业, 1990(6): 15-20. [4] CHEN Y, QU X, XIONG F, et al. Challenges to saving China's freshwater biodiversity: fishery exploitation and landscape pressures[J]. Ambio, 2020, 49(4): 926-938. doi: 10.1007/s13280-019-01246-2
[5] REID A J, CARLSON A K, CREED I F, et al. Emerging threats and persistent conservation challenges for freshwater biodiversity[J]. Biol Rev, 2019, 94(3): 849-873. doi: 10.1111/brv.12480
[6] ZHANG H, KANG M, SHEN L, et al. Rapid change in Yangtze fisheries and its implications for global freshwater ecosystem management[J]. Fish Fish, 2020, 21(3): 601-620. doi: 10.1111/faf.12449
[7] TAN X, LI X, LEK S, et al. Annual dynamics of the abundance of fish larvae and its relationship with hydrological variation in the Pearl River[J]. Environ Biol Fish, 2010, 88(3): 217-225. doi: 10.1007/s10641-010-9632-y
[8] 陆奎贤. 珠江水系渔业资源[M]. 广州: 广东科技出版社, 1990: 136-139. [9] 李思发, 吴力钊, 王强, 等. 长江、珠江、黑龙江鲢、鳙、草鱼种质资源研究[M]. 上海: 上海科学技术出版社, 1990: 1-228. [10] 田见龙, 王东. 长江野生草鱼性状和生长的研究[J]. 河南师范大学学报 (自然科学版), 1989(2): 60-65. [11] 李新辉, 李跃飞, 张迎秋, 等. 珠江肇庆段漂流性鱼卵、仔鱼监测日志 (2006)[M]. 北京: 科学出版社, 2020: 7-102. [12] 陈新军, 刘必林. 渔业资源生物学[M]. 北京: 科学出版社, 2017: 32-45. [13] 殷名称. 鱼类生态学[M]. 北京: 中国农业出版社, 2003: 51-55. [14] FROESE R, TSIKLIRAS A C, STERGIOU K I. Editorial note on weight-length relations of fishes[J]. Acta Ichthyol Piscat, 2011, 41(4): 261-263. doi: 10.3750/AIP2011.41.4.01
[15] ABU EL-NASR T M A. Age and growth of the fish, Gerres filamentosus (Cuvier, 1829) from Hurghada Red Sea, Egypt[J]. Egypt J Aquat Res, 2017, 43(3): 219-227. doi: 10.1016/j.ejar.2017.07.003
[16] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 2005: 124-130. [17] PAULY D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks[J]. ICES J Mar Sci, 1980, 39(2): 175-192. doi: 10.1093/icesjms/39.2.175
[18] 朱书礼, 李新辉, 李跃飞, 等. 西江广东肇庆段赤眼鳟的年龄鉴定及生长研究[J]. 南方水产科学, 2013, 9(2): 27-31. doi: 10.3969/j.issn.2095-0780.2013.02.005 [19] 陈得仿, 王腾, 刘永, 等. 大亚湾黑鲷繁殖特征的生物学研究[J]. 南方水产科学, 2019, 15(5): 41-47. doi: 10.12131/20190051 [20] 陈朋, 马燕武, 谢春刚, 等. 博斯腾湖草鱼生长特征的研究[J]. 淡水渔业, 2016, 46(4): 38-43. doi: 10.3969/j.issn.1000-6907.2016.04.006 [21] 熊飞, 刘红艳, 段辛斌, 等. 长江上游草鱼种群结构与生长特征[J]. 湖南师范大学自然科学学报, 2014, 37(4): 16-22. [22] FROESE R. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations[J]. J Appl Ichthyol, 2006, 22(4): 241-253. doi: 10.1111/j.1439-0426.2006.00805.x
[23] HANIF M A, ISLAM M A, SIDDIK M A B, et al. Length-weight relationships of three estuarine fish species from Bangladesh[J]. J Appl Ichthyol, 2018, 34(4): 1065-1067. doi: 10.1111/jai.13707
[24] MEHANNA S F, OSMAN A G M, FARRAG M M S, et al. Age and growth of three common species of goatfish exploited by artisanal fishery in Hurghada fishing area, Egypt[J]. J Appl Ichthyol, 2018, 34(4): 917-921. doi: 10.1111/jai.13590
[25] GUBIANI E A, GOMES L C, AGOSTINHO A A. Estimates of population parameters and consumption/biomass ratio for fishes in reservoirs, Paraná State, Brazil[J]. Neotrop Ichthyol, 2012, 10: 177-188. doi: 10.1590/S1679-62252012000100017
[26] PAULY D, MUNRO J L. Once more on the comparison of growth in fish and invertebrates[J]. Fishbyte, 1984, 2(1): 21-23.
[27] 陈大庆, 刘绍平, 段辛斌, 等. 长江中上游主要经济鱼类的渔业生物学特征[J]. 水生生物学报, 2002, 26(6): 618-622. doi: 10.3321/j.issn:1000-3207.2002.06.007 [28] 崔奕波, 李钟杰. 长江流域湖泊的渔业资源与环境保护[M]. 北京: 科学出版社, 2005: 193-228. [29] 徐钢春, 鲍明明, 杜富宽, 等. 鱼类性腺发育及产卵类型研究进展[J]. 长江大学学报 (自然科学版), 2017, 14(6): 43-48. [30] 温海深, 林浩然. 环境因子对硬骨鱼类性腺发育成熟及其排卵和产卵的调控[J]. 应用生态学报, 2001, 12(1): 151-155. doi: 10.3321/j.issn:1001-9332.2001.01.035 [31] 李思发, 周碧云, 吕国庆, 等. 长江鲢、鳙、草鱼和青鱼原种亲鱼标准与检测的研究[J]. 水产学报, 1997, 21(2): 143-151. [32] 孔一颖, 粤海渔. 珠江禁渔期增加两个月[J]. 海洋与渔业, 2017(3): 17. doi: 10.3969/j.issn.1672-4046.2017.03.002 [33] PATTERSON K. Fisheries for small pelagic species: an empirical approach to management targets[J]. Rev Fish Biol Fish, 1992, 2(4): 321-338. doi: 10.1007/BF00043521
[34] GRAY C A, BARNES L M, ROBBINS W D, et al. Length- and age-based demographics of exploited populations of stout whiting, Sillago robusta Stead, 1908[J]. J Appl Ichthyol, 2017(6): 1073-1082.
[35] WANG X H, QIU Y S, DU FY, et al. Population parameters and dynamic pool models of commercial fishes in the Beibu Gulf, northern South China Sea[J]. Chin J Oceanol Limn, 2012, 30(1): 105-117.
[36] 王茂元. 闽江中游鲢的年龄与生长[J]. 中国农学通报, 2020, 36(11): 124-129. -
期刊类型引用(1)
1. 张翠萍,袁丽媚,吴妤莘,叶志权,陈晓莹,赖星星,李强,舒琥. 珠江口花鰶的生物学特征研究. 水生生物学报. 2023(04): 637-647 .  百度学术
百度学术
其他类型引用(4)



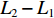
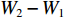
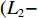




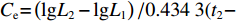

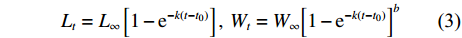


















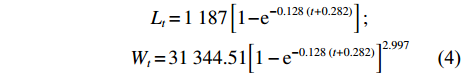

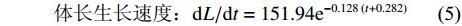
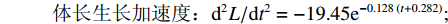




计量
- 文章访问数: 1130
- HTML全文浏览量: 339
- PDF下载量: 70
- 被引次数: 5
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
