
Length-based assessment of Nemipterus bathybius in northern South China Sea
-
摘要:
深水金线鱼 (Nemipterus bathybius) 是南海北部重要的底层经济鱼类,但近年来其资源呈现过度开发的态势。利用2014—2019年在南海北部底拖网调查中采集的3 059尾深水金线鱼生物学数据,使用基于长度的贝叶斯生物量评估 (Length-based Bayesian biomass estimation method, LBB) 和基于长度的繁殖潜力比(Length-based spawning potential ratio, LBSPR) 2种数据缺乏条件下的资源评估模型,对其资源状况进行了评估,为其种群科学管理和可持续利用提供技术支撑。结果表明,2014—2019年南海北部深水金线鱼的渐近体长 (Linf)、相对自然死亡率 (M/K) 和50%性成熟体长 (L50) 分别为23.7 cm、2.33和11.76 cm。LBB模型评估结果显示,其种群资源量水平 (B/BMSY)、50%渔获长度与最适可捕长度的比值 (Lc/Lc_opt) 分别为0.89、0.85,表明深水金线鱼处于轻度过度开发状态和生长型过度捕捞状态。LBSPR模型评估结果显示繁殖潜力比 (SPR) 为0.19,说明深水金线鱼正处于过度捕捞状态。通过先验参数的不确定性分析,发现LBB和LBSPR对参数Linf的设置极为敏感,对参数M/K的设置比较敏感,因此在使用LBB和LBSPR模型进行评估时应谨慎设置以上2种参数。
Abstract:Yellowbelly threadfin bream (Nemipterus bathybius), an economically important demersal fish species in the northern South China Sea, has been overexploited in recent years. To provide technical support for the scientific management and sustainable utilization of N. bathybius stocks, based on the biological data of 3 059 individuals of N. bathybius collected during bottom trawl surveys in the northern South China Sea from 2014 to 2019, we assessed the stock status by using two assessment models [Length-based Bayesian biomass (LBB) and length-based spawning potential ratio (LBSPR)] under data-poor conditions. Results show that the asymptotic fork length (Linf), relative natural mortality (M/K), and 50% sexually mature body length (L50) of N. bathybius in the northern South China Sea from 2014 to 2019 were 23.7 cm, 2.33 and 11.76 cm, respectively. The LBB assessment results show that the relative biomass level (B/BMSY) and ratio of length of 50% of the individuals captured by the gear to optimal length-at-first-capture (Lc/Lc_opt) were 0.89 and 0.85, respectively, indicating that N. bathybius was experiencing moderate overfishing and growth overfishing. The LBSPR results show a spawning potential ratio (SPR) of 0.19, indicating that N. bathybius was experiencing overfishing. Uncertainty analysis of the priori parameters reveals that LBB and LBSPR results were extremely sensitive to the setting of Linf and sensitive to the setting of M/K. Therefore, the above two parameters should be set with caution when using LBB and LBSPR.
-
Keywords:
- Nemipterus bathybius /
- Stock assessment /
- Data-poor /
- Uncertainty /
- Biomass /
- Spawning potential ratio
-
四指马鲅 (Elentheronema tetradactylum) 隶属于辐鳍鱼纲、鲈形目、马鲅亚目、马鲅科、四指马鲅属,俗称午鱼、马友,是一种暖温、广盐性鱼类,主要分布于太平洋、印度洋西部以及澳大利亚西海岸,在中国东海、南海均有分布[1-2]。四指马鲅具有肉质鲜美、生长速度快、营养价值高等特点[3],是我国重要的海洋经济养殖鱼类[4]。近年来,随着四指马鲅人工繁育技术的突破,已在广东、广西、福建、海南、浙江等沿海地区推广养殖,现已发展成为水产养殖的新品种[5]。国内对于四指马鲅的研究主要集中在组织结构[6-7]、环境胁迫[5,8]、营养成分[9]及养殖技术[10]等方面。目前四指马鲅的养殖存在苗种培育阶段死亡率较高的问题,严重制约了其养殖生产规模的扩大。
肠道作为鱼类消化吸收的主要器官[11],其结构与功能对鱼类的生长、发育至关重要。幼鱼的肠道发育情况直接关系到鱼类对营养物质的吸收和利用效率[12],肠道内存在的免疫细胞可以帮助幼鱼抵抗病原体入侵[13],肠黏膜所形成的特殊构造如皱襞高度、绒毛高度及隐窝深度等是评估肠道发育及其消化吸收功能的重要组织形态学指标[14]。此外,肠道绒毛上的杯状细胞也是反映肠道消化能力的重要指标[15]。幼鱼在生长过程中,其肠道结构逐步完善,饵料来源也趋于多样化,明显影响到肠道菌群的变化[16]。幼鱼摄食不同的饵料也会对肠道结构发育和肠道菌群组成产生影响[17]。此外,健康的肠道菌群可促进鱼类生长和减少肠道炎症,肠道菌群与宿主之间存在着密切的相互作用关系,它们不仅参与了营养物质代谢和能量转化[18],还可通过激活免疫细胞调节宿主的免疫系统[19],增强疾病抵抗能力[20]。以往研究表明,多种鱼类的肠道菌群结构在生长阶段处于动态平衡状态,3和6月龄花鲈 (Lateolabrax japonicus) 肠道微生物中的鲸杆菌属 (Cetobacterium) 相对丰度较高,9月龄的梭状芽孢杆菌属 (Clostridium) 相对丰度较高[13];大口黑鲈 (Micropterus salmoides) 肠道微生物中,气单胞菌属 (Aeromonas) 在第5天占据优势,而第25天假单胞菌属 (Pseudomonas)、乳球菌属 (Lactococcus)和不动杆菌属 (Acinetobacter) 相对丰度增加;第45天假单胞菌属和芽孢杆菌属(Bacillus)相对丰度较高,第180天成鱼肠道中检测出以邻单胞菌属 (Plssiomonas) 为代表的新优势菌群[21]。因此,研究四指马鲅不同生长发育阶段肠道结构形态及肠道菌群组成,可掌握其肠道对外源饵料的适应性及肠道菌群的变化规律,对其幼鱼期间的饵料投喂策略和病害防治具有重要意义。
本研究选用四指马鲅幼鱼进行养殖生长实验,结合肠道组织苏木素-伊红 (HE)染色切片观察、肠道菌群检测,比较分析了四指马鲅幼鱼在不同阶段的生长性能、肠道组织发育和结构形态变化以及肠道菌群结构;旨在揭示四指马鲅在幼鱼阶段的摄食生长与肠道发育、菌群变化之间的内在联系,为制定适宜的饵料投喂制度、病害防控等日常管理提供理论支撑。
1. 材料与方法
1.1 实验材料与管理
在广东省中山市四指马鲅养殖基地开展实验。2023年5月,采用同一批次的鱼苗6 000尾开展实验,平均全长为 (1.26±0.07) cm,平均体质量为 (1.5±0.1)×10−3 g。在室内育苗池 (4 m×5 m×1.2 m) 中暂养7 d,水体盐度为10‰,pH为7.6~8.0,溶解氧 (DO) 质量浓度>5 mg·L−1。暂养期间,投喂鳗鱼粉 (质量分数:蛋白质≥45%,脂肪≥10%),逐渐降低盐度至3‰。暂养结束后,转移到池塘内的3口网箱 (长3 m×宽4 m×高1.7 m) 中,养殖密度为100尾·m−3,池塘水体盐度为3‰~5‰,pH为7.5~8.1,养殖周期共12周。实验开始时,鱼为37日龄,在37~44日龄投喂鳗鱼粉,44~58日龄投喂粒径为0.6 mm的饲料,58~72日龄和72~86日龄均投喂粒径为1 mm的饲料,86~114日龄投喂粒径为2 mm的饲料,饲料粗蛋白质量分数为48%,粗脂肪质量分数为5%,粗灰分质量分数为6%。每天饱食投喂2次,日投喂量为体质量的5%。
1.2 肠道组织学分析
分别在37、44、58、72、86和114日龄采样,从每个网箱中随机挑选5尾鱼 (采样前停食1 d),记录全长、体长、体质量,计算特定生长率 (SGR)、平均日增质量 (ADG) 和肥满度 (CF);取3尾鱼中肠1 cm置于4% (w)多聚甲醛溶液固定,经过乙醇脱水、二甲苯透明、石蜡包埋、切片、HE染色和中性树脂封片,采用光学显微镜 (ZEISS Axio ScopeA1型)进行观察和记录图像。使用Image J在每个切片选取5根肠道绒毛,测量绒毛长度、宽度、肌层厚度,计算肠道结构变化情况。
1.3 肠道微生物16S rDNA基因高通量测序与分析
取3尾鱼,用75% (φ) 乙醇消毒体表,用无菌镊子取肠道组织,置于2 mL冻存管内并于 −80 ℃保存,以测定肠道菌群组成。采用HiPure Stool DNA mini Kit试剂盒 (Magen,中国)提取肠道内容物中微生物的总基因组DNA。DNA浓度使用NanoDrop 2000检测。使用正向引物338F (5'-ACTCCTACGAGGCAGCA-3') 和反向引物806R (5'-GGACTACHVGGGTWTCTAAT-3'),对16S rDNA基因的可变V3—V4区域进行扩增:94 ℃ 5 min;94 ℃ 30 s,53 ℃ 30 s,72 ℃ 30 s,共29个循环;72 ℃ 8 min。扩增后的PCR产物通过AMPure XP Beads进行纯化,并经过建库检测,将符合条件的样本文库使用Illumina NovaSeq平台对测序样本进行双端测序。
使用QIIME2 (V 2020.2)软件[22]对原始数据进行切除Barcods和引物、质量过滤、去噪和去嵌合体,将相似性100%的序列归为扩增子测序变体 (Amplicon sequence variants, ASV),得到包含细菌特征及相对丰度的表格和细菌代表性序列文件。使用Silva123数据库对代表性序列进行注释确定物种分类信息。通过QIIME命令summarize_taxa_through_plots.py生成各分类水平的细菌群落组成信息。通过QIIME命令alpha_diversity.py和collate_alpha.py 计算细菌α-多样性 (香农多样性指数Shannon和物种丰富度指数Richness)。通过QIIME命令beta_ diversity_through_plots.py计算细菌群落的β-多样性,利用基于Bray-Curtis距离的主坐标分析 (Principal coordinates analysis, PCoA) 表征不同组间肠道菌群结构的差异。使用线性判别分析LEfSe (Linear discriminant analysis effect Size)[23]确定不同日龄幼鱼肠道细菌组成的显著性差异,阈值对数评分为4.0。使用SPSS 22.0软件对肠道菌群属水平与肠道组织结构进行Pearson相关性分析,采用Origin 2024软件作图。
1.4 统计与分析
四指马鲅幼鱼的生长性能指标包括特定生长率 (SGR)、平均日增质量 (ADG) 和肥满度 (CF),计算公式如下:
$$ \mathrm{SGR=(ln} {W} _{ {t} } \mathrm{−ln} {W} _{ \mathrm{0}} \mathrm{)/} {t} \mathrm{\times 100\% } $$ (1) $$ \mathrm{ADG=(}{W}_{{t}}\mathrm{−}{W}_{\mathrm{0}}\mathrm{)/}{t} $$ (2) $$ \mathrm{CF=} {W} _{ {t} } \mathrm{/} {L} ^{ \mathrm{3}} $$ (3) 式中:W0为初始体质量 (g);Wt为终末体质量 (g);t为试验周期 (d);L为平均体长 (cm)。
实验数据均以“平均值±标准误 ($\bar x $±$ s_{\bar x}$, n=5)”表示,通过SPSS 20.0软件进行单因素方差分析 (One-way ANOVA),通过Tukey's 检验方法进行多重比较。p<0.05表示具有显著性差异。
2. 结果
2.1 四指马鲅生长性能测定
实验期间,不同日龄的四指马鲅在特定生长率、平均日增质量、肥满度上均存在显著性差异 (p<0.05,表1)。其中,44日龄的特定生长率显著高于其他日龄 (p<0.05),37日龄的显著高于58和72日龄,其余各日龄之间差异不显著;86日龄的平均日增质量显著高于其他日龄 (p<0.05),其余日龄差异不显著;58~114日龄的肥满度随幼鱼生长而逐渐升高,且72、86和114日龄的肥满度显著高于58日龄 (p<0.05)。37与44日龄的肥满度显著高于其他日龄 (p<0.05),原因可能是其消化道未发育完全,存在卵黄囊,导致体型较为肥满。
表 1 不同日龄四指马鲅生长性能Table 1. Growth performance of E. tetradactylum at different days of age指标
Indicator组别 Group A B C D E F 体长 Body length/cm 2.37±0.25 3.47±0.21 6.20±.032 7.00±0.30 10.08±0.36 13.20±1.19 体质量 Body mass/g 0.24±0.02 0.71±0.07 1.60±0.34 3.46±0.65 8.62±1.21 44.72±11.37 特定生长率SGR/(%·d−1) 8.58±0.23b 15.36±1.73a 5.72±1.45c 5.38±1.62c 6.43±1.16bc 6.52±2.02bc 平均日增质量ADG/(g·d−1) 0.01±0.00a 0.07±0.01a 0.06±0.02a 0.13±0.05a 0.52±0.29b 1.15±0.22a 肥满度CF/(g·cm−3) 2.88±0.95a 2.47±0.32a 0.85±0.03c 1.44±0.17b 1.67±0.09b 1.77±0.42b 注:同行中不同小写字母间存在显著性差异 (p<0.05)。A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。 Note: Values with different lowercase letters within the same row are significantly different (p<0.05). A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively. 2.2 四指马鲅肠道组织结构变化
随着养殖时间的增加,四指马鲅肠道绒毛的长度、宽度和肌层厚度不断增加,且各日龄间存在显著性差异 (p<0.05),绒毛数量和杯状细胞数量也有所增加 (表2,图1)。绒毛长度由91.69 μm增长到312.78 μm,绒毛宽度由19.39 μm增长到42.76 μm,肌层厚度由12.31 μm增加到56.72 μm。在44日龄时肠道壁结构界限模糊,杯状细胞数量稀少,每个绒毛上分布1~4个,主要分布于肠道绒毛前端或近前端;58日龄时,肠道组织结构清晰,杯状细胞增多,每个绒毛上分布5~10个。72、86和114日龄时,肠道组织结构清晰,黏膜层、黏膜下层、肌层和浆膜结构明显,杯状细胞数量显著增多,每个绒毛分布11~34个,且均匀分布在绒毛的前、中、后部。
表 2 不同日龄四指马鲅肠道组织结构变化Table 2. Intestinal tissue structure of E. tetradactylum at different days of age指标
Indicator组别 Group B C D E F 绒毛长度 Villus length/μm 91.69±8.42d 124.33±22.00c 168.11±42.35b 189.60±25.91b 312.78±45.92a 绒毛宽度 Villus width/μm 19.39±4.39c 20.14±3.29c 28.40±4.33b 38.28±8.84a 42.76±10.51a 肌层厚度 Muscular thickness/μm 12.31±1.57d 16.57±6.22bc 20.03±5.86c 31.58±5.07b 56.72±15.13a 注:同行中不同小写字母间存在显著性差异 (p<0.05)。B、C、D、E、F分别表示44、58、72、86、114日龄的幼鱼。 Note: Values with different lowercase letters within the same row are significantly different (p<0.05). B, C, D, E and F represent the E. tetradactylum juveniles of 44, 58, 72, 86 and 114 days of age, respectively. ![]() 图 1 不同日龄四指马鲅肠道组织结构变化Figure 1. Intestinal tissue structure of E. tetradactylum at different days of age
图 1 不同日龄四指马鲅肠道组织结构变化Figure 1. Intestinal tissue structure of E. tetradactylum at different days of age2.3 肠道菌群结构变化
2.3.1 Alpha多样性指数
本研究共获得原始序列数2 534 257条,经质控和过滤后共得到有效序列2 388 796条,序列长度为236~249 bp,平均长度243 bp。按照97%的序列相似性聚类为7 543个OTUs,属于39个细菌门类、73个纲、128个目、235个科和520个属。不同日龄四指马鲅肠道内共有OTUs为228个,占总数的2.67%,37、44、58、72和86日龄的特有OTUs分别为1 567、1 330、1 354、1 073、1 005和986个 (图2)。随着日龄的增加,肠道内特有OTUs总数逐渐降低。不同日龄四指马鲅鱼苗的肠道菌群Alpha多样性情况见表3。不同日龄的Chao 1和ACE指数差异均不显著,在37日龄具有最大值。从Shannon指数来看,37日龄群落多样性最高,显著高于86日龄 (p<0.05)。不同日龄间的Simpson指数无显著性差异 (p>0.05)。
![]() 图 2 不同日龄四指马鲅肠道内细菌韦恩图分析注:每个椭圆代表一个分组,不同椭圆交集部分为不同分组间的共有物种。其中A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 2. VENN analysis of intestinal microbiota in E. tetradactylum at different days of ageNote: Each ellipse represents a member group, and the intersection of different ellipses represents the common species between different groups. A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.表 3 不同日龄四指马鲅鱼肠道菌群Alpha多样性统计Table 3. Alpha diversity of intestinal microbiota in E. tetradactylum at different days of age
图 2 不同日龄四指马鲅肠道内细菌韦恩图分析注:每个椭圆代表一个分组,不同椭圆交集部分为不同分组间的共有物种。其中A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 2. VENN analysis of intestinal microbiota in E. tetradactylum at different days of ageNote: Each ellipse represents a member group, and the intersection of different ellipses represents the common species between different groups. A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.表 3 不同日龄四指马鲅鱼肠道菌群Alpha多样性统计Table 3. Alpha diversity of intestinal microbiota in E. tetradactylum at different days of age指标
Indicator组别 Group A B C D E F Chao 1 指数 Chao 1 index 1 019.70±247.37 919.92±24.98 795.38±167.93 818.00±32.87 739.96±231.98 805.94±80.17 ACE 指数 ACE index 986.88±211.51 938.39±29.54 810.60±163.07 817.51±20.64 774.20±258.03 866.18±93.10 香农指数Shannon index 4.65±0.01a 4.21±0.08ab 4.72±0.61a 3.86±0.47ab 3.16±1.58b 4.52±0.37ab 辛普森指数Simpson index 0.97±0.01 0.95±0.01 0.96±0.02 0.93±0.04 0.94±0.02 0.97±0.02 注:同行中不同小写字母间存在显著性差异 (p<0.05)。A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。 Note: Values with different lowercase letters within the same row are significantly different (p<0.05). A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively. 2.3.2 基于门水平肠道菌群分析
在门水平上,四指马鲅肠道细菌种类及丰度从高到低 (取丰度前5类细菌)依次为变形菌门、放线菌门、厚壁菌门、梭杆菌门和蓝细菌门 (图3)。在幼鱼生长过程中,变形菌门为绝对优势菌群,其相对丰度在86日龄阶段有所下降,随后又明显回升。在多数采样阶段,放线菌门和厚壁菌门也是主要的细菌门类,变形菌门、放线菌门和厚壁菌门占肠道细菌总丰度的75%以上。在86日龄时,四指马鲅肠道内的梭杆菌门和蓝细菌门明显增加,伴随着变形菌门相对丰度的显著降低 (p<0.05)。
![]() 图 3 不同日龄四指马鲅肠道内细菌门类相对丰度注:A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 3. Relative abundances of microbes in E. tetradactylum at different days of age on phylum levelNote: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.
图 3 不同日龄四指马鲅肠道内细菌门类相对丰度注:A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 3. Relative abundances of microbes in E. tetradactylum at different days of age on phylum levelNote: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.2.3.3 基于属水平的肠道菌群分析
在属水平上,四指马鲅肠道细菌种类及丰度按从高到低 (取丰度前5类细菌) 顺序为伯克氏菌属 (Burkholderia)、鲸杆菌属 (Cetobacterium)、申氏杆菌属 (Shinella)、芽殖杆菌属 (Gemmobacter) 和硝酸盐还原污物球菌属 (Defluviimonas) (图4)。其中,37日龄的优势属为芽殖杆菌属、副球菌属 (Paracocccus) 和红细菌属 (Rhodobacter);44日龄为硝酸盐还原污物球菌属、芽殖杆菌属和申氏杆菌属;58日龄为伯克氏菌属、弧菌属 (Vibrio) 和罗尔斯通氏菌属 (Ralstonia);72日龄为申氏杆菌属、博斯氏菌属 (Bosea) 和红细菌属;86日龄为鲸杆菌属、副球菌属和伯克氏菌属;114日龄为伯克氏菌属、罗尔斯通氏菌属和弧菌属。其中,硝酸盐还原污物球菌属、鲸杆菌属和伯克氏菌属在44、58、86和114日龄显著增加,而申氏杆菌、红细菌属和博斯氏菌属在72日龄显著增加;副球菌属和伯克氏菌属在58和72日龄显著减少,而红细菌属在58和86日龄显著减少。另外,弧菌属的相对丰度与罗尔斯通氏菌属丰度呈正相关,红细菌属的相对丰度与弧菌属呈负相关。
![]() 图 4 不同日龄四指马鲅肠道内细菌属水平相对丰度注:A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 4. Relative abundances of microbes in E. tradactylum at different days of age on genus levelNote: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.
图 4 不同日龄四指马鲅肠道内细菌属水平相对丰度注:A、B、C、D、E、F分别表示37、44、58、72、86、114日龄的幼鱼。Figure 4. Relative abundances of microbes in E. tradactylum at different days of age on genus levelNote: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.2.3.4 不同日龄四指马鲅肠道内菌群变化
采用LEfSe分析方法比较了不同日龄四指马鲅肠道中所有细菌的相对丰度差异性,在6个阶段 (A—F) 中有38个细菌分类群的相对丰度存在显著性差异 (图5)。在属水平上表现出显著差异的种类有16个:37日龄时的Nesiotabacter、Nautella、聚球藻属(Synechococcus)和杨氏菌属 (Yangia),44日龄时的硝酸盐还原污物球菌属、芽殖杆菌属和红球菌属的相对丰度明显增加,58日龄时的不动杆菌属和弧菌属,72日龄时的申氏菌属、红细菌属和剑菌属 (Ensifer)的相对丰度明显增加,86日龄时的鲸杆菌属和斯塔普氏菌属,14日龄时的伯克霍尔德氏菌属和罗尔斯通属的相对丰度明显增加。
![]() 图 5 LEfSe分析物种组成图 (LDA Score>4)注:A、B、C、D、E、F 分别表示37、44、58、72、86、114 日龄的幼鱼。Figure 5. LEfSe analysis of species compsition (LDA socre>4)Note: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.
图 5 LEfSe分析物种组成图 (LDA Score>4)注:A、B、C、D、E、F 分别表示37、44、58、72、86、114 日龄的幼鱼。Figure 5. LEfSe analysis of species compsition (LDA socre>4)Note: A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.在四指马鲅幼鱼肠道中变形菌门所包含的特征微生物范围最广,其次为蓝细菌门和梭杆菌门 (图6)。变形菌门中的α-变形菌纲、γ-变形菌纲和β-变形菌纲的丰度最高。进一步分析发现,α-变形菌纲中的红杆菌科在37日龄时显著高于其他阶段,但其下的属在所有日龄中均表现出显著性表现;莫拉菌科和弧菌科在58日龄时的丰度最高,根瘤菌科在72日龄时最高,梭杆菌科在86日龄时最高,伯克氏菌科在114日龄时最高。
![]() 图 6 物种分类学分支图注:Cladogram由内到外辐射的圆圈分别对应界、门、纲、目、科、属不同的分类层级,层级间的连线代表所属关系。每个圆圈节点代表1个物种,黄色节点代表分组间差异不显著,差异物种跟随组进行着色。有颜色的扇形区域标注了特征微生物的下属分类区间。A、B、C、D、E、F 分别表示37、44、58、72、86、114 日龄的幼鱼。Figure 6. Cladogram of speciesNote: Cladogram consists of circles radiating from the inside out, each corresponding to different levels of classification such as kingdom, phylum, class, order, family, and genus. The lines connecting the circles represent their relationships. Each circle node represents a species, with yellow nodes indicating insignificant differences between groups, while significantly different species are colored according to their group. Colored sector regions represent the subclassification intervals of characteristic microorganisms.A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.
图 6 物种分类学分支图注:Cladogram由内到外辐射的圆圈分别对应界、门、纲、目、科、属不同的分类层级,层级间的连线代表所属关系。每个圆圈节点代表1个物种,黄色节点代表分组间差异不显著,差异物种跟随组进行着色。有颜色的扇形区域标注了特征微生物的下属分类区间。A、B、C、D、E、F 分别表示37、44、58、72、86、114 日龄的幼鱼。Figure 6. Cladogram of speciesNote: Cladogram consists of circles radiating from the inside out, each corresponding to different levels of classification such as kingdom, phylum, class, order, family, and genus. The lines connecting the circles represent their relationships. Each circle node represents a species, with yellow nodes indicating insignificant differences between groups, while significantly different species are colored according to their group. Colored sector regions represent the subclassification intervals of characteristic microorganisms.A, B, C, D, E and F represent the E. tetradactylum juveniles of 37, 44, 58, 72, 86 and 114 days of age, respectively.2.3.5 肠道菌群和肠道组织结构相关性分析
对四指马鲅幼鱼肠道组织结构和肠道菌属的相对丰度进行相关性分析发现 (图7),伯克氏菌属、弧菌属和罗尔斯通氏菌属与绒毛长度、肌层厚度呈显著正相关性 (p<0.05),芽殖杆菌属和硝酸盐还原污物球菌属与绒毛长度、宽度和肌层厚度呈显著负相关性 (p<0.05)。
![]() 图 7 相关性分析热图注:色块的颜色表示相关系数,其中红色代表正相关,蓝色代表负相关;颜色深浅代表相关系数大小 (*. p<0.05)。Figure 7. Heat map of correlation analysisNote: The colors of the color blocks represent the correlation coefficients. Red represents positive correlation and blue represents negative correlation; the color shades represent the size of the correlation coefficients (*. p<0.05).
图 7 相关性分析热图注:色块的颜色表示相关系数,其中红色代表正相关,蓝色代表负相关;颜色深浅代表相关系数大小 (*. p<0.05)。Figure 7. Heat map of correlation analysisNote: The colors of the color blocks represent the correlation coefficients. Red represents positive correlation and blue represents negative correlation; the color shades represent the size of the correlation coefficients (*. p<0.05).3. 讨论
已有研究表明稚鱼、幼鱼阶段的消化道发育尚不完善,消化能力较弱,进食频率过高或过低都可能造成肠道受损,进而导致免疫力下降甚至感染疾病[24]。随着稚鱼成长为成鱼,其肠道内的组织、结构和功能逐渐发育成熟[25]。一般情况下,海水鱼类的仔稚鱼期在30日龄内[26-28],已有研究发现四指马鲅幼鱼43日龄已具完整的黏膜褶皱,消化系统结构与功能的发育已能基本满足机体的生长代谢需要[7]。肠道绒毛数量越多,长度和宽度越大,肠道吸收面积越大,机体的消化吸收能力越强[29]。杯状细胞可分泌黏液和消化酶,黏液有保护黏膜和润滑作用,消化酶可提高消化能力[30],杯状细胞数量的增多,也反映了消化能力的增强[31]。卵形鲳鲹 (Trachinotus ovatus) 肠道杯状细胞最早于孵化后的第3天出现在前肠,且数量随着仔鱼的生长而增加[32]。在本研究中,通过肠道组织切片观察发现37日龄四指马鲅肠道结构基本成型,但是绒毛数量、杯状细胞仍较少,肠道壁与黏膜层界限模糊,表明37~44日龄的四指马鲅仍处于稚鱼期,可投喂少量粉状饲料,但投喂量不宜过多。在58~114日龄,四指马鲅肠道结构发育明显,与成鱼基本一致,肠道绒毛长度、宽度逐渐增加,肠道绒毛数量和杯状细胞数量明显增加,表明其消化能力不断增强,因此在58日龄后逐渐增加人工饵料的投喂量可能更符合四指马鲅幼鱼的消化机能发育规律。此外,通过肠道属水平细菌与肠道结构进行Pearson相关性分析发现,伯克氏菌属和罗尔斯通氏菌属的相对丰度随着绒毛长度和肌层厚度的增加而升高,芽殖杆菌属和硝酸盐还原污物球菌属的相对丰度随着绒毛长度、宽度和肌层厚度的增加而降低。已有研究表明,伯克氏菌属和罗尔斯通氏菌属广泛存在于动物肠道和水环境中[33],随着幼鱼的生长发育,肠道绒毛增长,细菌的相对丰度也会增加。以上结果表明,随着鱼类的不断生长,其肠道细菌多样性在属水平上具有较大差异,且肠道内优势菌属与肠道结构发育存在相关性。优势菌属动态变化与组织结构相互联系,使得水生动物能够更好地获得生长所需的营养。
鱼类不同生长发育阶段的肠道菌群处于动态变化状态。肠道菌群易受生物和非生物因素影响,如水环境理化指标[34]、发育阶段[13]、个体差异[35]、不同区域[36]和生活习性[37]等因素均会影响肠道的菌群组成。肠道菌群在自然选择下与宿主形成寄生或共生关系[38],主要参与消化分解、营养代谢并影响宿主免疫系统[39]。已有研究表明,海水鱼肠道内的细菌门类主要以变形菌门、厚壁菌门和拟杆菌门等为主[37]。在本研究中,四指马鲅幼鱼肠道内的优势菌门为变形菌门、放线菌门、厚壁菌门、梭杆菌门和蓝细菌门,与虹鳟 (Oncorhynchus mykiss)[40]、赤点石斑鱼 (Epinephelus akaara)[41]、军曹鱼 (Rachycentron canadum)[42]肠道内的优势菌门相近。研究表明,大口黑鲈肠道内的气单胞菌属、不动杆菌属、肠杆菌属分别在第5、第25和第45天成为优势菌属,在第180 天时增加了多个新的优势菌属,例如邻单胞菌属、大肠埃希菌属-志贺氏菌属和鲸杆菌属等;随着幼鱼生长和饲料转换,七彩神仙鱼 (Symphysodon haraldi) 肠道内的内优势菌属由鲸杆菌属、邻单胞菌属和未识别的紫单胞菌科转化为鲸杆菌属、未分类消化链球菌科和Breznakia;不同月龄间花鲈的肠道微生物群落结构也表现出显著性差异,3和6月龄以鲸杆菌属、邻单胞菌属和罗姆布茨菌属为主,9月龄以梭菌属为优势菌属。鲸杆菌属作为一种共生菌,普遍存在于多种鱼类肠道中,具有调节肠道菌群组成、改善肝脏健康的功能[43]。在集约化养殖水库中的6种健康鱼类肠道中均检测到鲸杆菌属,该菌在杂交条纹鲈 (Morone saxatilis)、欧洲鲈 (Dicentrarchus labrax)、美国红鱼 (Sciaenops ocellatus)、奥尼罗非鱼 (Oreochromis)、平头鲻 (Mugil cephalus) 中均属于优势菌属[44]。本研究中,鲸杆菌属为四指马鲅在86日龄时的主要优势菌属,而在其余阶段丰度较低,这可能与其消化能力增强有关,也可能与宿主的健康状况密切相关。有研究表明,鲸杆菌属是潜在的益生菌,可产生维生素B12和抗菌肽,促进营养吸收,提高机体抗病力[43,45]。此外,随着鱼类生长过程中肠道发育的逐渐完善,其肠道菌群变得更加复杂,不同时期的菌属具有较大差异,但菌群多样性却呈下降趋势[35,46];与本研究结果一致,即随着幼鱼生长,肠道菌群的Alpha多样性指数呈降低趋势,其肠道微生物在不同时期处于动态平衡状态,与大口黑鲈[21]、七彩神仙鱼[47]、花鲈[13]等肠道菌群的变化情况相近。
由于幼鱼的肠道发育尚未完善,饵料来源和转换较为频繁,易造成菌群组成不稳定[48]。有研究表明,不同开口饵料、转食过程及饲料种类均会对鱼类肠道菌群造成影响。本研究中,对37~44日龄的四指马鲅幼鱼投喂鳗鱼粉,其余日龄投喂同一品牌的营养成分一致但粒径大小不同的饲料。在幼鱼转食颗粒饲料后的58日龄,肠道菌群的多样性增加、丰富度降低,与大口黑鲈转食饲料[16]前后肠道菌群的Alpha多样性指数变化一致;而在转食颗粒饲料后,在门水平上,优势菌门未发生改变,说明变形菌门、厚壁菌门和放线菌门是维系四指马鲅肠道菌群稳态的主要门类;而在属水平上,弧菌属和罗尔斯通氏菌属明显增加,在58和114日龄时为优势菌属。罗尔斯通氏菌属和弧菌属中的大多数菌被认为是水产养殖中的机会致病菌[49],危害水产动物健康。本研究中,肠道罗尔斯通氏菌属和弧菌属的相对丰度在转食颗粒饲料后上升,在养殖中应加强对这一阶段的防控措施。添加益生菌、中草药制剂可以有效抑制弧菌属的生长和繁殖[50]。
水环境与鱼类的健康息息相关,良好的水环境不仅能为其提供适宜的生存环境,还能减少疾病发生。与成鱼相比,幼鱼肠道微生物更易受水环境影响[35]。伯克氏菌属[51]、申氏菌属[52]、红细菌属[53]和博斯氏菌属[54]均属于反硝化菌,广泛存在于水环境和土壤中,可促进养殖环境的氮循环,在牙鲆 (Paralichthys olivaceus) 幼鱼肠道中亦有存在[55]。本研究中,幼鱼肠道中的伯克氏菌属、申氏菌属、红细菌属和博斯氏菌属丰度在44和72日龄时出现显著变化,且红细菌属的相对丰度较高,弧菌的相对丰度较低,二者可能存在一定程度的相互抑制作用。已有研究表明,光合细菌可以抑制弧菌[56],光合细菌与弧菌有一定的拮抗作用,通过添加光合菌制剂可使弧菌数量最多下降23.5%[57]。本研究中,不同日龄的四指马鲅幼鱼肠道中均存在不同比例的弧菌属,但均未发生明显病害,可能是因为弧菌属于原籍菌群或具有条件致病性[58]。
4. 结论
58日龄的四指马鲅肠道结构发育较为完善,在此阶段适合增加人工颗粒饵料投喂。变形菌门、放线菌门和厚壁菌门是四指马鲅幼鱼肠道中的优势菌门,不同日龄间的优势菌属具有显著性差异,且肠道内优势菌属的相对丰度与肠道结构发育状况具有相关性。在四指马鲅的生长过程中,由鳗鱼粉转食颗粒饲料阶段,有害菌丰度增加,应格外注意饵料转换时期的病害防治。
-
![]()
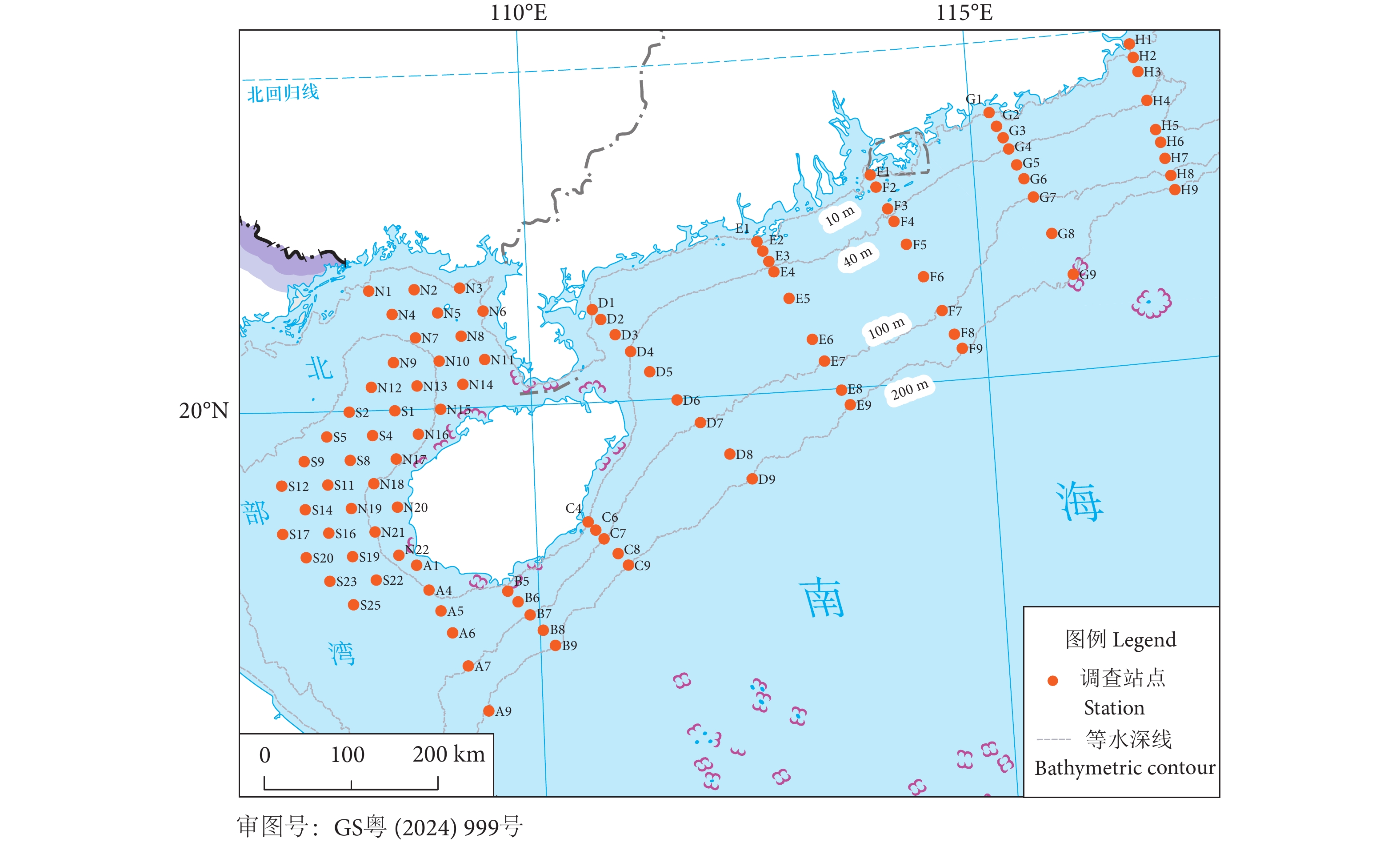
图 1 南海北部近海研究区域及站点分布
Figure 1. Study area and station distribution in offshore northern South China Sea
![]()
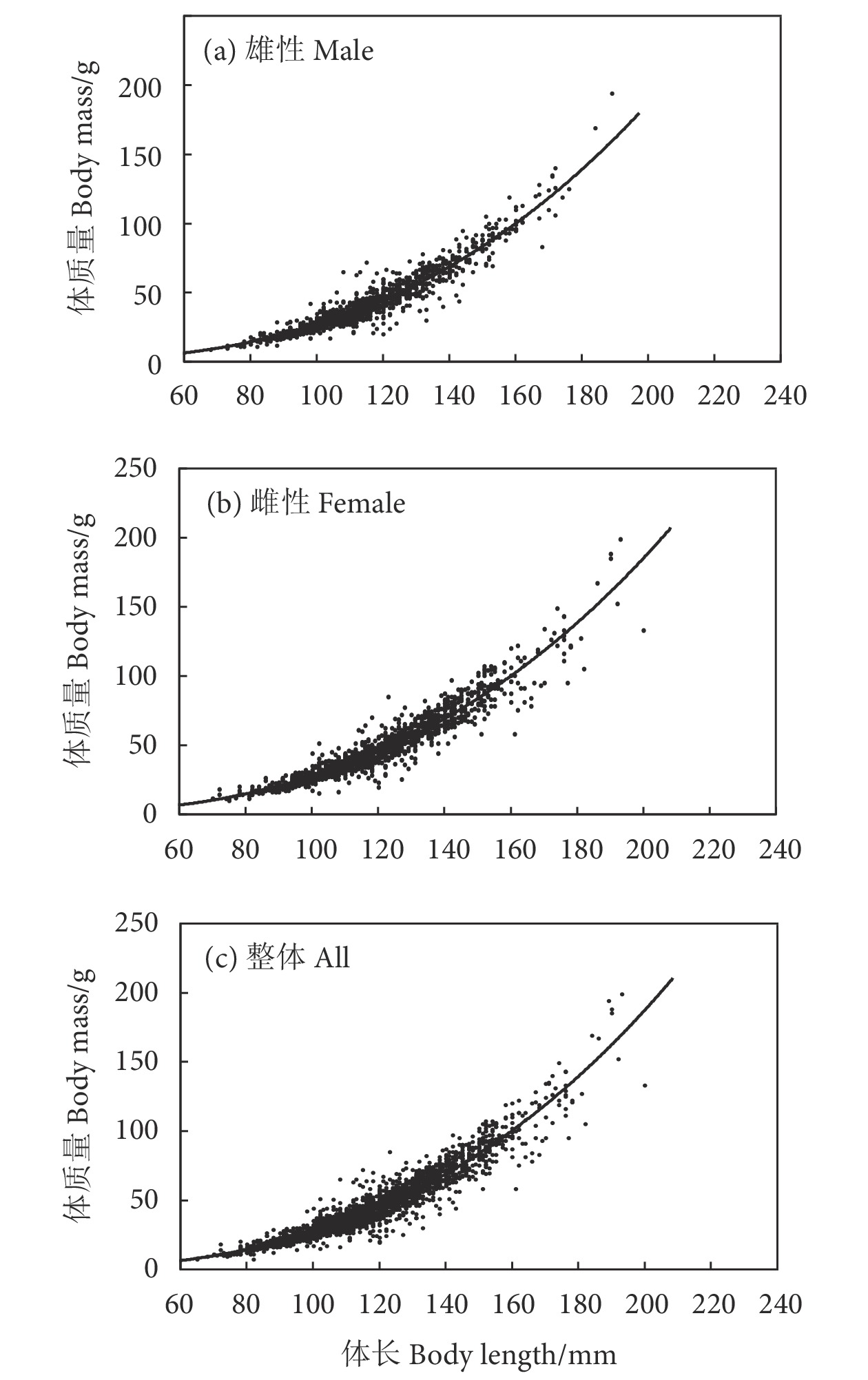
图 2 南海北部雄性、雌性、整体深水金线鱼体长-体质量关系
Figure 2. Length-body mass relationship among male, female and all N. bathybius in northern South China Sea
![]()
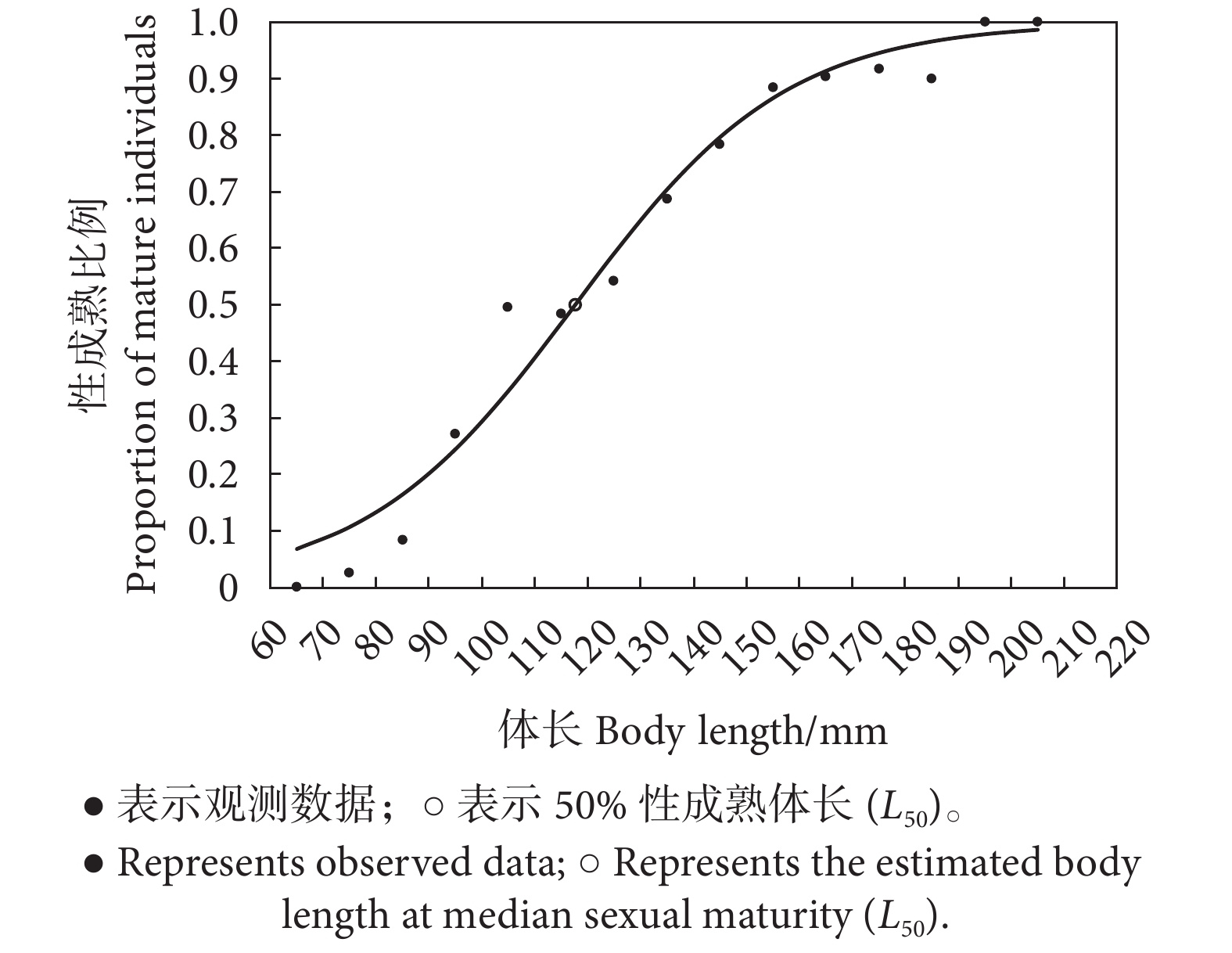
图 3 南海北部深水金线鱼性成熟比例与体长拟合的逻辑斯蒂曲线
Figure 3. Logistic model fitted for relationship between body length and percentage of mature N. bathybius in northern South China Sea
![]()
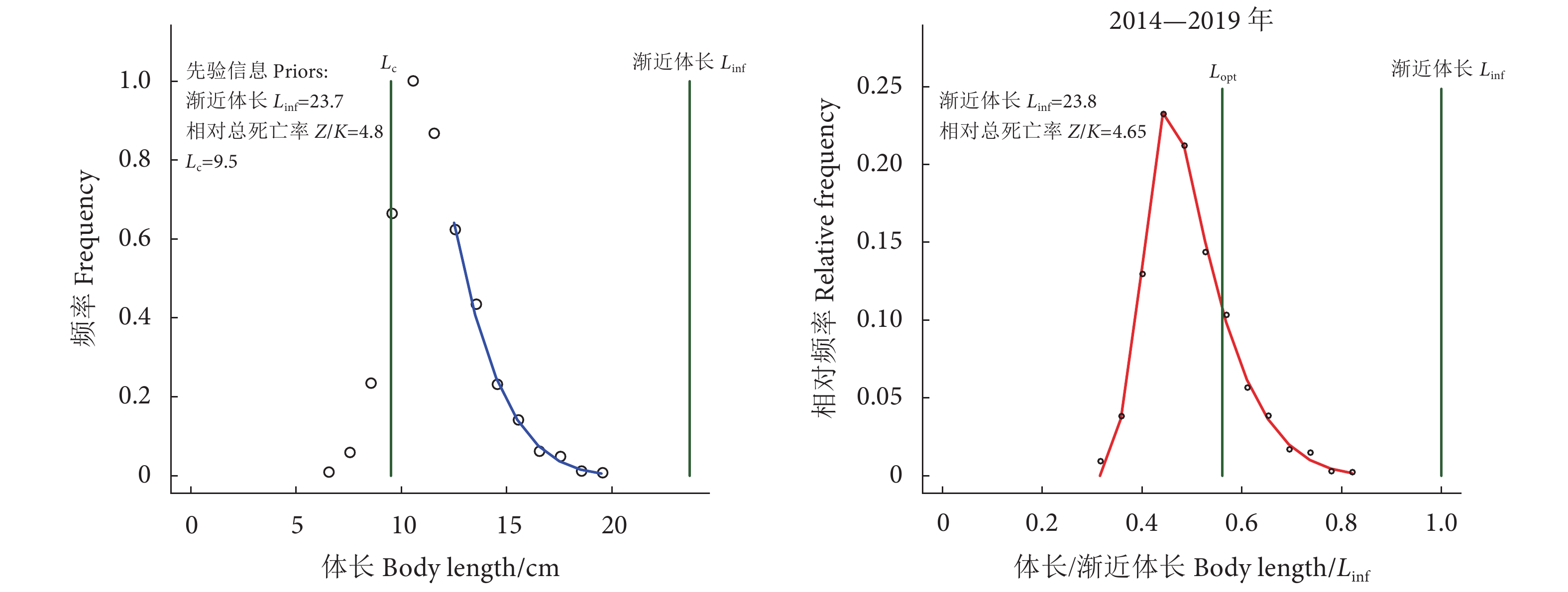
图 4 南海北部深水金线鱼基于LBB方法的分析结果
注:左图为模型对体长数据的拟合曲线,右图为LBB方法的预测;Lc是50%个体被渔具捕获的体长;Lopt为未开发种群获得最大生物量的捕捞体长。
Figure 4. Estimated values for N. bathybius in northern South China Sea by length-based Bayesian biomass assessment model
Note: The left figure shows the fits of the model to length data, and the right figure shows the predictions of the LBB analysis; Lc. Length of 50% of the individuals captured by the gear; Lopt. Fishing body length for obtaining maximum biomass in undeveloped populations.
![]()
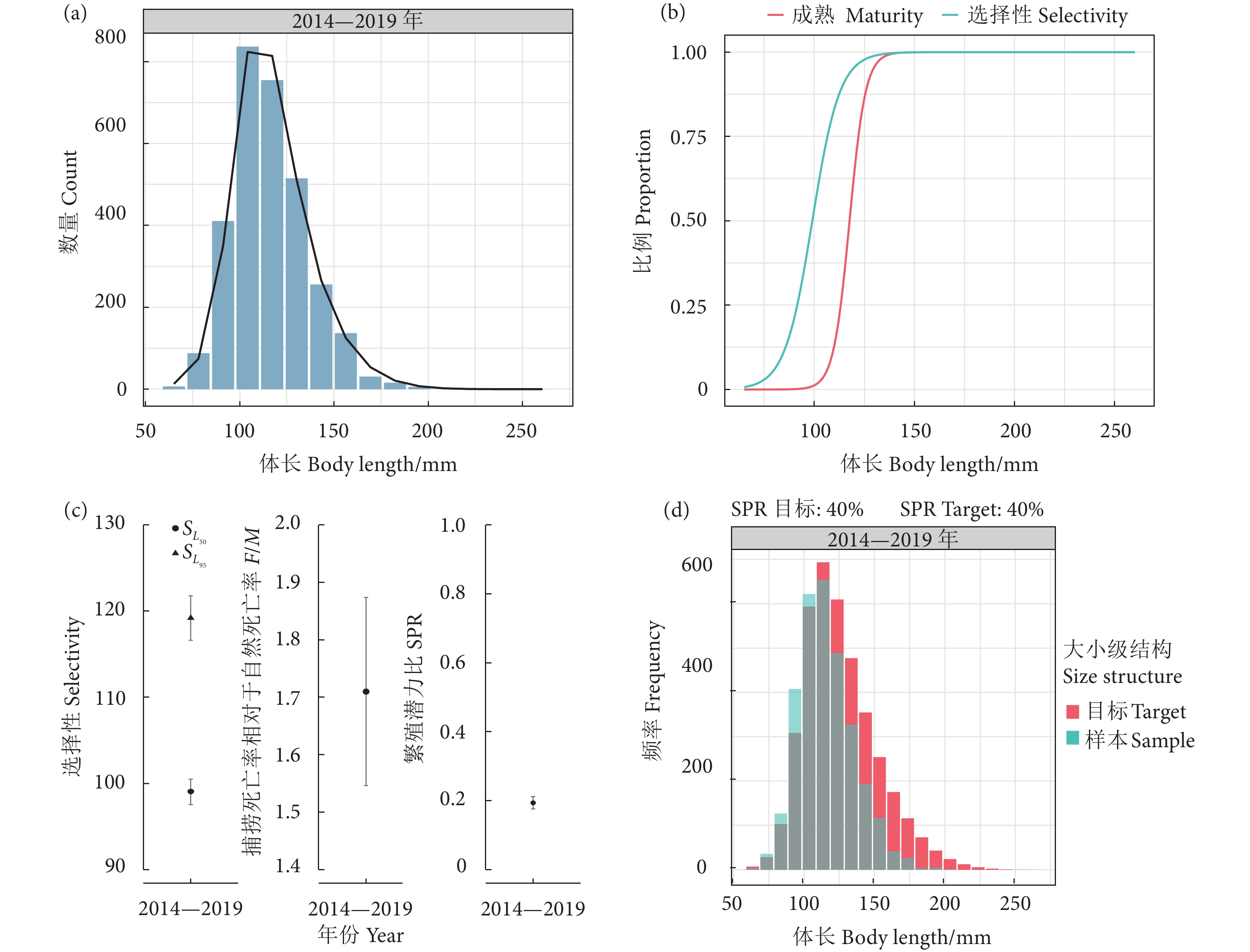
图 5 南海北部深水金线鱼基于LBSPR方法的分析结果
注:a. 长度-频率分布(柱状图),预测的种群大小组成(折线图);b. LBSPR模型拟合的性成熟和选择性曲线,其中L50=11.76 cm和L95=12.94 cm时;c. 选择性参数($S_{L_{50}}$和 $S_{L_{95}} $)、捕捞死亡率与自然死亡率的比值(F/M)和繁殖潜力比的估计值;d. 在目标SPR=0.4时,观测到的长度-频率数据与预期的大小组成。
Figure 5. Length-based spawning potential ratio for N. bathybius in northern South China Sea
Note: a. Length-frequency distribution (Pillars), and predicted fished size composition (Line chart); b. Maturity and selectivity curves from the fitted LBSPR model when L50 was 11.76 cm and L95 was 12.94 cm; c. Distribution of mean selectivity parameters ($S_{L_{50}} $ and $S_{L_{95}} $), fishing mortality to natural mortality (F/M), and spawning potential ratio; d. Observed length-frequency data against an expected size composition at a target SPR of 0.4.
表 1 南海北部深水金线鱼的样品信息
Table 1 Sample information of N. bathybius in northern South China Sea
年份
Year月份
Month样本数
Sampling
number体长 Body length/mm 范围
Range均值±标准差
Mean±Standard
deviation2014 7 293 93~193 121.2±16.3 10 225 77~184 119.9±21.6 2015 1 120 73~172 121.8±18.7 4 330 75~189 111.4±19.2 2016 7 304 84~200 121.8±19.4 10 78 74~178 122.1±23.8 2017 1 71 72~176 109.3±18.7 4 197 72~158 115.0±15.3 2018 3 220 75~176 113.8±19.0 8 87 78~182 126.0±24.2 2019 3 524 73~176 106.6±16.8 9 610 65~192 117.3±19.0  下载: 导出CSV
下载: 导出CSV
表 2 LBB和LBSPR方法所需数据目录
Table 2 Data for LBB and LBSPR methods
参数
Parameter估算值
Estimated value基于长度的繁殖潜力比模型
Length-based spawning potential ratio
(LBSPR) assessment model基于长度的贝叶斯估计模型
Length-based Bayesian biomass
(LBB) assessment model长度-频率数据
Length–frequency data/ √ √ 渐近体长 (Linf)
Asymptotic length/cm23.7 cm √ 〇 生长系数 (K)
Growth coefficient0.326 〇 自然死亡率 (M)
Natural mortality0.762 〇 相对自然死亡率M/K 2.33 √ 〇 50%性成熟体长 (L50)
Body length at 50% sexual maturity/cm11.76 √ 〇 95%性成熟体长 (L95)
Body length at 95% sexual maturity/cm12.94 √ 渐近体长的变异系数
Coefficient of variation of Linf0.1 √ 长度-体质量关系参数 (a)
Length-body mass relationship parameter6.854 0×10−5 〇 长度-体质量关系参数 (b)
Length-body mass relationship parameter2.797 4 〇 注:“√” 代表必需数据;“〇”代表可选数据。 Note: "√" represents required data; "〇" represents optional data.
下载: 导出CSV
表 3 LBB和LBSPR在不同先验参数下的评估结果
Table 3 Estimated results of LBB and LBSPR parameters with different priors
组号
Group No.先验参数
Prior parameterLBSPR结果
Results of LBSPRLBB结果
Results of LBBLinf/mm L50/mm M/K F/M SPR B/BMSY F/M Lc/Lc_opt 1 237 117.6 2.33 1.71 0.19 0.89 1.00 0.85 2 232 1.73 0.20 0.99 0.90 0.87 3 234 1.76 0.20 0.94 0.97 0.86 4 236 1.71 0.19 0.90 1.00 0.85 5 238 1.84 0.18 0.87 1.10 0.84 6 240 1.73 0.19 0.84 1.10 0.83 7 242 1.94 0.17 0.81 1.10 0.82 8 237 112.6 2.33 1.71 0.21 0.89 1.00 0.85 9 114.6 1.71 0.21 0.89 1.00 0.85 10 116.6 1.71 0.20 0.89 1.00 0.85 11 118.6 1.71 0.19 0.89 1.00 0.85 12 120.6 1.71 0.18 0.89 1.00 0.85 13 122.6 1.71 0.17 0.89 1.00 0.85 14 237 117.6 2.28 1.77 0.19 0.84 1.10 0.83 15 2.30 1.75 0.19 0.87 1.00 0.84 16 2.32 1.73 0.19 0.87 1.00 0.84 17 2.34 1.70 0.20 0.89 1.00 0.84 18 2.36 1.68 0.20 0.92 1.00 0.86 19 2.38 1.66 0.20 0.92 0.99 0.86
下载: 导出CSV
表 4 不同时期深水金线鱼种群参数
Table 4 Population parameters of N. bathybius in different periods
参数
Parameter年份 Year 1992 1997 2009 2017 渐近体长 Asymptotic length (Linf)/mm 242 220 235 237 生长系数Growth coefficient (K) 0.39 0.45 0.48 0.326 50%性成熟体长Body length at 50% sexual maturity (L50)/mm 128.6 116.5 110.5 117.6
下载: 导出CSV
-
[1] 陈再超, 刘继兴. 南海经济鱼类[M]. 广州: 广东科学与技术出版社, 1982: 184-188. [2] 宁平. 中国金线鱼科鱼类分类、系统发育及动物地理学研究 [D]. 青岛: 中国科学院海洋研究所, 2012: 25-28. [3] 王雪辉, 邱永松, 杜飞雁. 南海北部深水金线鱼生物学及最适开捕体长[J]. 应用生态学报, 2005(12): 2428-2434. doi: 10.3321/j.issn:1001-9332.2005.12.041 [4] 陈作志, 孔啸兰, 徐姗楠, 等. 北部湾深水金线鱼种群参数的动态变化[J]. 水产学报, 2012, 36(4): 584-591. [5] 崔奕波. 鱼类生物能量学的理论与方法[J]. 水生生物学报, 1989, 13(4): 15. doi: 10.3321/j.issn:1000-3207.1989.04.003 [6] 李忠炉, 卢伙胜, 甘喜萍, 等. 北部湾口海域深水金线鱼生长和死亡研究[J]. 水产科学, 2009, 28(10): 556-562. doi: 10.3969/j.issn.1003-1111.2009.10.002 [7] 陈作志, 林昭进, 邱永松. 基于AHP的南海海域渔业资源可持续利用评价[J]. 自然资源学报, 2010, 25(2): 249-257. doi: 10.11849/zrzyxb.2010.02.009 [8] 张魁, 廖宝超, 许友伟, 等. 基于渔业统计数据的南海区渔业资源可捕量评估[J]. 海洋学报, 2017, 39(8): 25-33. [9] ZHANG K, CAI Y C, LIAO B C, et al. Population dynamics of threadfin porgy Evynnis cardinalis, an endangered species on the IUCN red list in the Beibu Gulf, South China Sea[J]. J Fish Biol, 2020, 97(2): 479-489. doi: 10.1111/jfb.14398
[10] ZHANG K, GUO J Z, XU Y W, et al. Long-term variations in fish community structure under multiple stressors in a semi-closed marine ecosystem in the South China Sea[J]. Sci Total Environ, 2020, 745: 140892. doi: 10.1016/j.scitotenv.2020.140892
[11] ZHANG K, LI J J, HOU G, et al. Length-based assessment of fish stocks in a data-poor, jointly exploited (China and Vietnam) fishing ground, northern South China Sea[J]. Front Mar Sci, 2021, 8: 718052. doi: 10.3389/fmars.2021.718052
[12] 耿喆, 朱江峰, 夏萌, 等. 数据缺乏条件下的渔业资源评估方法研究进展[J]. 海洋湖沼通报, 2018, 164(5): 130-137. [13] 刘维达, 林昭进, 江艳娥, 等. 南海北部陆架区底层渔业资源的空间分布特征[J]. 热带海洋学报, 2011, 30(5): 95-103. doi: 10.3969/j.issn.1009-5470.2011.05.013 [14] MILDENBERGER T K, TAYLOR M H, WOLFF M. TropFishR: an R package for fisheries analysis with length-frequency data[J]. Methods Ecol Evol, 2017, 8(11): 1520-1527. doi: 10.1111/2041-210X.12791
[15] PAULY D. Some simple methods for the assessment of tropical fish stocks. FAO Fisheries Technical Paper No. 234[M]. Rome: FAO, 1983: 50-57.
[16] PAULY D, MUNRO J L. Once more on the comparison of growth in fish and invertebrates[J]. Fishbyte, 1984, 2(1): 1-21.
[17] PAULY D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks[J]. ICES J Mar Sci, 1980, 39(2): 175-192. doi: 10.1093/icesjms/39.2.175
[18] THEN A Y, HOENIG J M, HALL N G, et al. Evaluating the predictive performance of empirical estimators of natural mortality rate using information on over 200 fish species[J]. ICES J Mar Sci, 2015, 72(1): 82-92. doi: 10.1093/icesjms/fsu136
[19] YU Y, ZHANG H R, JIN J B, et al. Trends of sea surface temperature and sea surface temperature fronts in the South China Sea during 2003–2017[J]. Acta Oceanol Sin, 2019, 38(4): 106-115. doi: 10.1007/s13131-019-1416-4
[20] VILA Y, SILVA L, TORRES M A, et al. Fishery, distribution pattern and biological aspects of the common European squid Loligo vulgaris in the Gulf of Cadiz[J]. Fish Res, 2010, 106(2): 222-228. doi: 10.1016/j.fishres.2010.06.007
[21] PRINCE J, VICTOR S, KLOULCHAD V, et al. Length based SPR assessment of eleven Indo-Pacific coral reef fish populations in Palau[J]. Fish Res, 2015, 171: 42-58. doi: 10.1016/j.fishres.2015.06.008
[22] PONS M, COPE J M, KELL L T. Comparing performance of catch-based and length-based stock assessment methods in data-limited fisheries[J]. Can J Fish Aquat Sci, 2020, 77(6): 1026-1037. doi: 10.1139/cjfas-2019-0276
[23] ZHANG K, ZHANG J, ZHANG P, et al. This is what we know: assessing the stock status of the data-poor skipjack tuna (Katsuwonus pelamis) fishery in the South China Sea[J]. Front Mar Sci, 2023, 10: 1095411. doi: 10.3389/fmars.2023.1095411
[24] PALOMARES M L D, FROESE R, DERRICK B, et al. A preliminary global assessment of the status of exploited marine fish and invertebrate populations[R]. Vancouver: The University of British Columbia, 2018: 14-32.
[25] FROESE R, WINKER H, CORO G, et al. A new approach for estimating stock status from length frequency data[J]. ICES J Mar Sci, 2019, 76(1): 350-351. doi: 10.1093/icesjms/fsy139
[26] HORDYK A, ONO K, VALENCIA S, et al. A novel length-based empirical estimation method of spawning potential ratio (SPR), and tests of its performance, for small-scale, data-poor fisheries[J]. ICES J Mar Sci, 2014, 72(1): 217-231.
[27] WALTERS C J , MARTELL S J D. Fisheries ecology and management[M]. Princeton: Princeton University Press, 2004: 155-157.
[28] ALAM M S, LIU Q, SCHNEIDER P, et al. Length-based stock assessment for the data-poor Bombay duck fishery from the Northern Bay of Bengal Coast, Bangladesh[J]. J Mar Sci Eng, 2022, 10(2): 213. doi: 10.3390/jmse10020213
[29] RICHARD K, OUSMANE S, FENG W, et al. Length-based assessment methods for the conservation of a pelagic shark, Carcharhinus falciformis from the Tropical Pacific Ocean[J]. Fish Basel, 2022, 7(4): 184.
[30] HORDYK A, ONO K, SAINSBURY K, et al. Some explorations of the life history ratios to describe length composition, spawning-per-recruit, and the spawning potential ratio[J]. ICES J Mar Sci, 2014, 72(1): 204-216.
[31] DOWLING N A, DICHMONT C M, HADDON M, et al. Empirical harvest strategies for data-poor fisheries: a review of the literature[J]. Fish Res, 2015, 171: 141-153. doi: 10.1016/j.fishres.2014.11.005
[32] 夏恒睿, 王琨, 董秀强, 等. 基于体长-繁殖潜力比方法的海州湾小黄鱼资源状态评估[J]. 中国海洋大学学报(自然科学版), 2022, 52(12): 25-32. [33] GULLAND J A. Fish stock assessment: a manual of basic method[M]. New York: Wiley, 1983: 87-125.
[34] XU Y W, ZHANG P, PANHWAR S K, et al. The initial assessment of an important pelagic fish, Mackerel scad, in the South China Sea using data-poor length-based methods[J]. Mar Coast Fish, 2023, 15(5): 10258. doi: 10.1002/mcf2.10258
[35] LIAO B C, XU Y W, SUN M S, et al. Performance comparison of three data-poor methods with various types of data on assessing southern Atlantic albacore fishery[J]. Front Mar Sci, 2022, 9: 825461. doi: 10.3389/fmars.2022.825461
[36] CONOVER D O, MUNCH S B. Sustaining fisheries yields over evolutionary time scales[J]. Science, 2002, 297(5578): 94-106. doi: 10.1126/science.1074085
[37] 耿平, 张魁, 陈作志, 等. 北部湾蓝圆鲹生物学特征及开发状态的年际变化[J]. 南方水产科学, 2018, 14(6): 1-9. doi: 10.12131/20180106 [38] 史登福, 张魁, 蔡研聪, 等. 南海北部带鱼群体结构及生长、死亡和性成熟参数估计[J]. 南方水产科学, 2020, 16(5): 51-59. doi: 10.12131/20200055 [39] HOMMIK K, FITZGERALD C J, KELLY F, et al. Dome-shaped selectivity in LB-SPR: length-based assessment of data-limited inland fish stocks sampled with gillnets[J]. Fish Res, 2020, 229: 105574. doi: 10.1016/j.fishres.2020.105574
[40] 王雪辉, 杜飞雁, 邱永松. 南海北部主要经济鱼类体长与体重关系[J]. 台湾海峡, 2006(2): 262-266. [41] 王雪辉, 邱永松, 杜飞雁, 等. 基于长度贝叶斯生物量法估算北部湾二长棘鲷种群参数[J]. 水产学报, 2020, 44(10): 1654-1662. [42] ZHANG K, ZHANG J, SHI D F, et al. Assessment of coral reef fish stocks from the Nansha Islands, South China Sea, using length-based Bayesian biomass estimation[J]. Front Mar Sci, 2021, 7: 610707. doi: 10.3389/fmars.2020.610707







计量
- 文章访问数: 126
- HTML全文浏览量: 71
- PDF下载量: 64
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
