
Optimization of process and mechanism of extracting astaxanthin from shrimp shells with deep eutectic solvent
-
摘要:
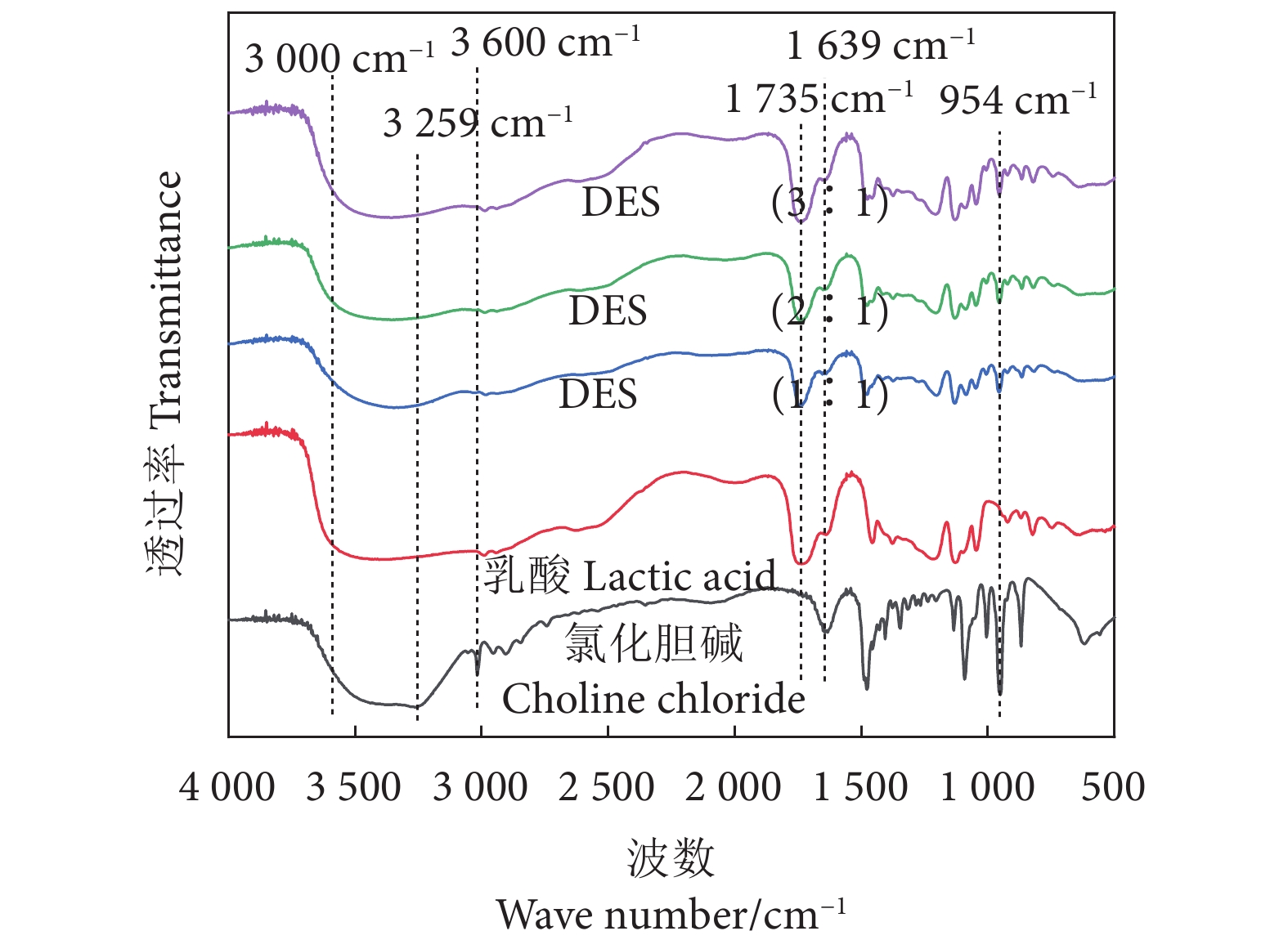
虾壳中虾青素的高效安全提取技术是促进虾类加工副产物变废为宝的途径之一。利用乳酸-氯化胆碱构成的低共熔溶剂提取虾壳中的虾青素,在单因素实验基础上,采用响应面实验优化提取工艺;通过低共熔溶剂的物化性质分析探究其对虾青素的提取机理。优化后的工艺条件为乳酸-氯化胆碱摩尔比为2.5∶1,含水量 (w) 为9.5%,超声时间为30.5 min,实际得率为24.41 μg·g−1。傅里叶变换红外光谱分析表明低共熔溶剂合成时官能团并未发生改变,二者融合产生的氢键可能是促进虾青素高效提取的主要原因,黏度会在一定程度上影响产品的提取过程。
Abstract:Development of efficient and safe extraction technology of astaxanthin from shrimp shells is crucial for transforming waste into valuable resources. In this study, astaxanthin in shrimp shells was extracted with deep eutectic solvent composed of lactic acid-choline chloride. A single-factor test was carried out to explore the influence of extraction conditions on the extraction rate, and a response surface test was adopted to optimize the extraction process. The mechanism of astaxanthin extraction with deep eutectic solvent was explored by physical and chemical analysis. The optimized process conditions were: molar ratio of 2.5∶1, water content of 9.5%, ultrasonic time of 30.5 min, and actual yield of 24.41 μg·g−1. The Fourier transform infrared spectroscopy shows that there was no change in functional groups during deep eutectic solvent synthesis, but hydrogen bond might be the reason for effective extraction of astaxanthin, and viscosity would affect the extraction process of product.
-
Keywords:
- Astaxanthin /
- Deep eutectic solvent /
- Lactic acid /
- Choline chloride
-
广东鲂 (Megalobrama terminalis) 是一种江河洄游鱼类,在我国南方水系中占有重要的渔业生产地位[1-2]。我国南方水系复杂多样,并经历了多次河流袭夺,而气候变化导致的海平面波动是形成其特有河网系统的重要因素之一[3]。由于广东鲂对淡水环境依赖性较强,因此,海平面波动产生的地理隔离,对广东鲂种群的遗传结构产生了一定影响。Chen等[3]指出3个广东鲂地理遗传种群分别为珠江、漠阳江和海南岛万泉河种群,并发现广东鲂种群在不同的淡水栖息地中表现出明显的适应性分化。刘凯等[4]研究表明,受地理分布、食物来源、遗传特征和栖息环境等因素的叠加影响,鱼类的不同地理种群在肌肉营养成分上能够产生相对稳定的变异。然而,不同广东鲂地理种群肌肉营养成分是否存在显著差异尚不清楚。目前,对广东鲂研究多在于早期资源、资源捕捞量、性腺发育、繁殖策略、消化生理等方面[1-3,5-8],针对不同广东鲂种群肌肉营养成分和能量密度的研究尚未见报道。由于人类活动的不断加强 (如水利水电工程、航道治理、水污染、过度捕捞等),珠江野生广东鲂种群数量持续下降[2,7-8],漠阳江和万泉河广东鲂种群则呈现规模小、片段化分布特征,在其他陆河河流如榕江、鉴江、韩江中已难以监测到野生样本。因此,本研究测定了万泉河、漠阳江、西江广东鲂种群肌肉营养成分和能量密度,探究不同地理广东鲂种群肌肉营养成分差异,以期充实鱼类营养学和能量生态学研究材料,也为不同广东鲂的野生地理种群的分类保护和合理利用提供科学依据。
1. 材料与方法
1.1 样本采集
2020年6—7月分别于海南省琼海市万泉河琼海段 (QH, 110°27"36'E, 19°12"36'N)、广东省阳江市漠阳江段 (YJ, 111°42"2'E, 22°48"7'N) 以及广东省肇庆市珠江干流 (西江) 肇庆江段 (ZQ, 112°24"35'E, 23°5"24'N) 采集到150尾广东鲂,各采样点50尾 (雌、雄各25尾) 。采用哈希水质分析仪测量取样点的水温、盐度、溶解氧 (Dissolved oxygen, DO) 和pH,并测量样品的体长和体质量。采样站位的环境信息和样本生物学信息见表1。采用液氮快速冷冻样本,于–20 ℃冷冻保存,随后带回实验室−80 ℃保存。采集背部中后段肌肉 (每尾在相同位置采集20 g肌肉) 用于检测肌肉成分。各项指标由广东省质量监督食品检验站进行检测,各实验组均设置3个重复,每个重复含10尾样本 (背部肌肉捣碎)。
表 1 3个广东鲂地理种群环境信息和样本生物学信息Table 1. Basic environmental information, biological information of three M. terminalis populations指标
Index采样点 Sampling site 琼海 QH 阳江 YJ 肇庆 ZQ 环境指标 Environmental index 水温 Water temperature/℃ 30.0±0.2 29.2±0.4 28.6±0.3 盐度 Salinity 0.03±0.01 0.01±0.02 0.01±0.01 酸碱度 pH 7.7±0.3 7.9±0.3 7.8±0.3 溶解氧质量浓度 DO/(mg∙L−1) 6.8±0.2 6.7±0.2 7.0±0.2 生物学指标 Biological index 体长 Body length ($\overline { X}\pm { \rm {SD}} $)/mm 233±17.6 253±20.7 271±27.3 体质量 Body mass ($\overline { X}\pm { \rm {SD}} $)/g 354±19.4 424±33.1 521±30.1 1.2 肌肉营养成分和能量密度测定方法
样本水分测定采用GB 5009.3—2016;粗蛋白测定采用GB 5009.5—2016;脂肪测定采用GB 5009.4—2016;灰分测定采用GB 5009.4—2016;氨基酸测定采用GB T5009.124—2016;脂肪酸测定采用GB 5009.168—2016。采用电感耦合等离子体质谱仪 (7700 Series) 依据GB 5009—2016测定样本中钾 (K)、钙 (Ca)、钠 (Na)、磷 (P)、镁 (Mg)、锌 (Zn)、铁 (Fe)、铜 (Cu)、锰 (Mn) 等矿质元素含量。根据联合国粮农组织/世界卫生组织 (FAO/WHO) 提出和1991年中国预防医学科学院营养与食品卫生研究所提出的氨基酸评分模式,计算氨基酸评分 (Amino acid score, AAS)、化学评分 (Chemical score, CS) 和必需氨基酸指数 (Essential amino acid index, EAAI) [9-10]。Phillipson微量能量仪 (Gentry Instruments Inc., Aiken, South Carolina, USA) 测定能量密度。
1.3 数据分析
采用单因素方差分析 (One-way ANOVA) 检验不同广东鲂地理种群肌肉营养成分差异显著性。如差异显著,则采用多重比较方法比较平均数之间的差异,显著性水平为0.05。数据分析采用SPSS 19.0统计软件进行。实验数据均用“平均值±标准差 (
$\overline X \pm {\rm{SD}} $ )”表示。采用R (3.1.14) 对3个广东鲂种群肌肉生化分析结果进行主成分分析 (Principal component analysis, PCA)。2. 结果
2.1 一般营养成分和能量密度分析
万泉河广东鲂肌肉水分质量分数显著高于西江种群,而粗蛋白质质量分数则显著低于西江种群 (P<0.05,表2)。粗脂肪和灰分质量分数在3个地理种群中均无显著性差异。西江种群肌肉能量密度显著高于万泉河和漠阳江种群 (P<0.05)。
表 2 3个广东鲂地理种群肌肉中的一般营养成分和能量密度Table 2. Nutritional composition of muscle of three M. terminalis populations项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 水分质量分数 Moisture mass fraction/% 79.0±0.5a 78.2±0.9ab 77.0±0.6b 粗蛋白质质量分数 Crude protein mass fraction/% 18.3±0.5b 19.1±0.7ab 20.2±0.5a 粗脂肪质量分数 Crude lipid mass fraction/% 1.2±0.1 1.1±0.1 1.3±0.1 粗灰分质量分数 Ash mass fraction/% 1.1±0.1 1.2±0.1 1.2±0.1 能量密度 Energy density/(kJ·g−1) 3.1±0.1b 3.2±0.2b 3.6±0.2a 注:同行不同上标字母表示差异显著 (P<0.05),下表同此。 Note: Different superscript letters within the same row indicate significant difference (P<0.05). The same case in the following tables. 2.2 氨基酸组成分析与评价
3个广东鲂地理种群共检测出18种常见氨基酸 (表3)。西江种群肌肉中的总氨基酸含量 (Total amino acids, TAA) 最高,漠阳江种群次之,万泉河种群最低。在必需氨基酸 (Essential aamino acid, EAA) 中,西江种群的赖氨酸和亮氨酸含量显著高于万泉河种群 (P<0.05),漠阳江种群介于两者之间。呈味氨基酸中,西江种群肌肉中的天冬氨酸、谷氨酸、甘氨酸和丙氨酸含量均显著高于万泉河种群 (P<0.05)。漠阳江种群肌肉中必需氨基酸/总氨基酸 (EAA/TAA) 最高,而西江种群最低。呈味氨基酸/总氨基酸 (DAA/TAA) 在3个广东鲂地理种群肌肉中无明显差异。芳香氨基酸/支链氨基酸 (BCAA/AAA) 在万泉河种群肌肉中最高,漠阳江种群次之,西江种群最低。将3个广东鲂地理种群肌肉的EAAI进行标准模式 (FAO/WHO) 及全鸡蛋蛋白质模式2种评价 (表3),分别计算出各EAA的AAS、CS和EAAI (表4)。万泉河、漠阳江和西江种群肌肉中的第一限制性氨基酸为蛋氨酸+半胱氨酸,第二限制性氨基酸为缬氨酸,其余各EAA的AAS均高于1;各EAA的CS与AAS结果保持一致。3个广东鲂地理种群肌肉中的EAAI达80以上,说明其氨基酸组成十分均衡。其中西江种群肌肉EAAI最高 (85.05)。
表 3 3个广东鲂地理种群肌肉氨基酸组成Table 3. Comparison of amino acid composition of muscles of three M. terminalis populations项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 天冬氨酸* Asp 1.80±0.04b 1.82±0.07b 1.99±0.05a 苏氨酸# Thr 0.80±0.02 0.80±0.03 0.86±0.04 丝氨酸 Ser 0.72±0.02 0.73±0.03 0.80±0.04 谷氨酸* Glu 2.78±0.06b 2.68±0.10b 2.99±0.14a 脯氨酸 Pro 0.61±0.02 0.63±0.02 0.66±0.04 甘氨酸* Gly 0.83±0.04b 0.87±0.04ab 0.97±0.06a 丙氨酸* Ala 1.06±0.02b 1.09±0.03b 1.18±0.08a 缬氨酸#△ Val 0.85±0.02 0.87±0.03 0.92±0.04 蛋氨酸 Met 0.53±0.01 0.53±0.02 0.57±0.03 异亮氨酸#△ Ile 0.78±0.02 0.79±0.02 0.83±0.03 亮氨酸#△ Leu 1.41±0.03b 1.41±0.05ab 1.52±0.05a 半胱氨酸 Cys 0.11±0.07 0.19±0.03 0.43±0.15 酪氨酸◆ Tyr 0.62±0.02 0.61±0.02 0.66±0.02 苯丙氨酸# Phe 0.72±0.02 0.74±0.02 0.80±0.03 赖氨酸# Lys 1.73±0.05a 1.73±0.09ab 1.87±0.03b 组氨酸○ His 0.42±0.01 0.45±0.04 0.48±0.05 精氨酸○ Arg 1.07±0.04 1.07±0.04 1.17±0.07 色氨酸# Trp 0.27±0.04 0.35±0.03 0.37±0.05 氨基酸总量 TAA 17.11±0.58a 17.37±0.69a 19.07±0.57b 呈味氨基酸总量 DAA 6.47±0.06a 6.46±0.08a 7.13±0.07b 必需氨基酸/非必需氨基酸 EAA/NEAA/% 72.41 73.03 69.95 必需氨基酸/总氨基酸 EAA/TAA/% 38.34 38.51 37.60 半必需氨基酸/总氨基酸 SEAA/TAA/% 8.36 8.41 8.26 芳香氨基酸/支链氨基酸 BCAA/AAA/% 4.90 5.03 4.05 注:#. 必需氨基酸;○. 半必需氨基酸;*. 呈味氨基酸;△. 支链氨基酸;◆. 芳香氨基酸。 Note: #. Essential amino acid; ○. Semiessential amino acid; *. Delicious amino acid; △. Branched chain amino acid; ◆. Aromatic amino acid. 表 4 3个广东鲂地理种群肌肉氨基酸评价Table 4. Evaluation of essential amino acids composition of muscle of three M. terminalis populations评价模式
Evaluation method氨基酸
Amino acids广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 氨基酸评分 AAS 苏氨酸 Thr 1.09 1.05 1.07 缬氨酸 Val 0.94 0.92 0.92 色氨酸 Trp 1.53 1.91 1.91 异亮氨酸 Ile 1.06 1.03 1.03 亮氨酸 Leu 1.09 1.05 1.07 赖氨酸 Lys 1.73 1.66 1.70 苯丙氨酸 Phe+酪氨酸 Tyr 1.20 1.17 1.19 蛋氨酸 Met+半胱氨酸 Cys 0.82 0.78 0.80 化学评分 CS 苏氨酸 Thr 0.93 0.90 0.91 缬氨酸 Val 0.71 0.69 0.69 色氨酸 Trp 0.96 1.24 1.31 异亮氨酸 Ile 0.80 0.78 0.78 亮氨酸 Leu 0.90 0.86 0.88 赖氨酸 Lys 1.34 1.28 1.31 苯丙氨酸 Phe+酪氨酸 Tyr 0.81 0.78 0.80 蛋氨酸 Met+半胱氨酸 Cys 0.47 0.45 0.46 必需氨基酸指数 EAAI 83.31 83.37 85.05 2.3 脂肪酸组成分析
3个广东鲂地理种群肌肉中共检测出23种常见脂肪酸 (表5),其中包括7 种饱和脂肪酸 (Saturated fatty acid, SFA) 7种单不饱和脂肪酸 (Monounsaturated fatty acid, MUFA) 和9种多不饱和脂肪酸 (Polyunsaturated fatty acids, PUFA)。SFA中C14:0、C16:0和C22:0在万泉河种群肌肉中的含量显著高于漠阳江和西江种群 (P<0.05)。万泉河种群肌肉中MUFA总量显著低于西江和漠阳江种群 (P<0.05)。其中,C16:1、C18:1 n-9t、C18:1 n-9c漠阳江种群肌肉中含量最高,而在万泉河种群肌肉中含量最低。C22:1 n-9、C24:1在漠阳江种群肌肉中含量显著低于西江和万泉河种群。西江种群肌肉中二十二碳六希酸 (DHA) 含量最高,显著高于万泉河和漠阳江种群 (P<0.05)。PUFA在万泉河种群肌肉中含量最高,西江种群次之,漠阳江种群最低。
表 5 3个广东鲂地理种群肌肉脂肪酸组成Table 5. Comparison of fatty acids of muscles of three M. terminalis populations% 项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 肉豆蔻酸 C14:0 5.36±0.85a 3.53±0.41b 3.40±0.32b 十五碳酸 C15:0 1.02±0.25 0.66±0.05 0.70±0.13 棕榈酸 C16:0 27.33±1.47 a 21.83±1.16 b 21.80±1.99 b 珠光脂酸 C17:0 2.03±0.35 0.99±0.18 1.13±0.33 硬脂酸 C18:0 11.34±1.79a 5.97±1.03b 8.14±1.65ab 花生酸 C20:0 0.29±0.05a 0.15±0.02b 0.21±0.07ab 花生酸 C22:0 0.30±0.08a 0.10±0.01c 0.17±0.01b ∑饱和脂肪酸 SFA 47.67±2.12a 33.23±2.85b 35.56±4.35b 肉豆蔻烯酸 C14:1 0.06±0.02 0.12±0.02 0.08±0.01 棕榈油酸 C16:1 6.21±1.22b 10.45±1.73a 8.10±0.52ab 顺-11-二十碳一烯酸 C20:1 2.32±0.36 1.85±0.08 2.06±0.15 顺-15-二十四碳一烯酸 C24:1 0.27±0.09a 0.06±0.01b 0.24±0.10a 反式油酸 C18:1 n-9t 0.21±0.05b 0.43±0.07a 0.35±0.07ab 油酸 C18:1 n-9c 16.73±3.87b 35.53±4.35a 31.80±3.65a 二十二碳一烯酸 C22:1 n-9 1.09±0.36a 0.11±0.03b 1.03±0.61a ∑单不饱和脂肪酸 MUFA 26.89±3.93a 48.54±5.20b 43.66±7.08b 亚油酸 C18:2 n-6c 4.30±0.64a 2.23±0.33b 1.59±0.64b α-亚麻酸 C18:3 n-3 5.54±0.86a 4.03±1.56ab 2.35±0.51b γ-亚麻酸 C18:3 n-6 0.14±0.02a 0.08±0.02ab 0.01±0.00b 顺,顺-11,14-二十碳二烯酸 C20:2 0.39±0.08 0.34±0.01 0.36±0.04 顺-11,14,17-二十碳三烯酸 C20:3 n-3 0.32±0.05 0.27±0.06 0.21±0.08 顺,顺,顺-8,11,14-二十碳三烯酸 C20:3 n-6 0.39±0.09a 0.22±0.03b 0.14±0.01c 花生四烯酸 C20:4 n-6 (ARA) 5.27±0.89a 2.94±0.42b 4.88±0.95a 二十碳五烯酸 C20:5 n-3 (EPA) 3.16±0.72 4.06±0.10 4.09±0.87 二十二碳六烯酸 C22:6 n-3 (DHA) 5.74±1.03b 3.95±0.13c 7.03±1.28a ∑多不饱和脂肪酸 PUFA 25.11±2.83a 18.04±2.47b 20.64±5.61ab 2.4 矿质元素组成分析
3个广东鲂地理种群肌肉中均含有丰富的矿质元素,其中K质量分数最高,Ca次之 (表6)。西江种群肌肉中K和Ca质量分数显著高于万泉河种群,而Na和Mg质量分数则显著低于万泉河种群 (P<0.05)。万泉河种群肌肉Zn质量分数显著高于漠阳江和西江种群,而Mn和Fe质量分数显著低于漠阳江和西江种群 (P<0.05)。
表 6 3个广东鲂地理种群肌肉矿质元素组成Table 6. Mineral element of muscle of three M. terminalis populationsmg∙kg−1 元素
Element广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 钾 K 3 340.05±105.36b 3 460.04±192.92ab 3 820.36±221.12a 钙 Ca 1 050.12±28.87 b 1 100.25±40.02 ab 1 200.11±34.64 a 钠 Na 487.34±58.96a 345.35±7.23b 385.57±30.66b 镁 Mg 298.65±3.79 303.05±2.65 332.31±5.51 磷 P 241.59±25.97 230.45±2.08 247.78±4.04 锌 Zn 6.47±0.15a 4.16±0.18b 3.82±0.17b 铁 Fe 3.06±0.13c 4.75±0.14b 6.43±0.18a 铜 Cu 0.12±0.00 0.12±0.01 0.13±0.01 锰 Mn 0.21±0.01c 0.38±0.02b 0.64±0.01a 2.5 3个广东鲂种群生化分析结果的主成分分析
综合3个广东鲂种群生化分析结果,并进行PCA。西江种群分布距均万泉河和漠阳江种群较远,万泉河种群和漠阳江种群相对较近。PCA共提取了2个主成分,对变异的累积贡献率为80.25%。其中主成分1的贡献率为50.75%,主成分2的为29.50% (图1)。
![]() 图 1 3个广东鲂种群肌肉生化成分主成分分析散点图Figure 1. Scattering diagram of principal components for biochemical analysis parameters of muscle of three M. terminalis populations
图 1 3个广东鲂种群肌肉生化成分主成分分析散点图Figure 1. Scattering diagram of principal components for biochemical analysis parameters of muscle of three M. terminalis populations3. 讨论
鱼类肌肉中蛋白质和脂肪含量是评价其营养价值的重要指标[11]。3 个广东鲂地理种群肌肉的粗蛋白质量分数 (18.3%~20.2%) 高于团头鲂 (Megalobrama amblycephala)、鲤 (Cyprinus carpio)、鲢 (Hypophthalmichthys molitrix)、鳙 (H. nobilis) 和草鱼 (Ctenopharyngodon idella),与翘嘴鲌 (Culter alburnus) 接近[12-13]。3个广东鲂地理种群肌肉粗脂肪质量分数 (1.1%~1.3%) 较团头鲂、翘嘴鲌、鲢、鳙、斑鳜 (Siniperca scherzeri) 等低[12-14],与常见的海水鱼类如牙鲆 (Paralichthys olivaceus)[15]、黄斑篮子鱼 (Siganus oramin)[16]和日本鳗鲡 (Anguilla japonica)[17]类似,表现出典型的低脂肪、高蛋白的特点。本研究发现,3个广东鲂种群肌肉生化PCA结果显示西江种群分布距万泉河和漠阳江种群较远,可能是由于栖息地环境因子以及饵料生物种类存在明显差异。本研究还发现,西江种群能量密度显著高于漠阳江和万泉河种群。能量密度被认为是衡量鱼体能量储备水平的重要指标,能直接反映鱼类发育状况以及对外界环境因子的适应性[18]。鱼类生殖洄游是主动的、定期定向的高耗能运动,且鱼体自身能量储备有限,因此鱼类洄游须尽可能地调节自身身体结构、能量储备和代谢能力以适应生殖洄游的需要[19-20]。3个广东鲂种群生殖洄游距离存在明显差异,可能是导致种群间肌肉中能源物质的积累程度不同的主要原因之一。有研究发现鱼类肌肉能量累积和消耗与其洄游能力密切相关[21-22]。
鱼类肌肉中蛋白质的营养价值由各种EAA含量和组成比例决定[23-24]。本研究显示,在3个广东鲂种群肌肉中谷氨酸含量均最高,谷氨酸作为一种重要呈味氨基酸,具有促进脑发育、治疗神经系统疾病等作用[25]。3个广东鲂种群肌肉中谷氨酸含量均高于团头鲂与翘嘴鲌[12]。西江种群肌肉中谷氨酸含量显著高于漠阳江和万泉河种群,表明西江种群肌肉较万泉河和漠阳江种群风味更佳。3个广东鲂种群肌肉中赖氨酸含量均较高,其中,西江种群肌肉中赖氨酸含量最高。赖氨酸是人体EAA之一,不仅具有提高蛋白质利用率和促进人体生长发育的作用,还可以增强免疫力、改善神经系统、预防骨质疏松[24,26]。在FAO/WHO提出的人体均衡蛋白需求理想模式中,EAA/NEAA>60%的蛋白质质量较好[27],3个野生广东鲂种群肌肉均属于良好的蛋白源。西江种群肌肉中的EAAI最高 (85.05),说明其肌肉中EAA组成最为平衡,蛋白质营养价值最高。肌肉中的脂肪酸含量是影响肌肉风味的重要因素之一[28]。本研究发现,西江和漠阳江种群肌肉均表现出MUFA的高占比。有研究表明,MUFA在调节人体脂质代谢方面具有重要的生理作用[29]。PUFA中DHA与EPA含量是评价鱼类营养成分的关键指标[30]。西江种群肌肉中的DHA含量显著高于漠阳江和万泉河种群,表明西江种群肌肉的脂肪质量较高。
矿物质元素是构成人体组织的重要成分,参与人体内多种物质的代谢和生理活动[31]。3个广东鲂种群肌肉中Na、K、Ca等常规矿质元素以及Fe、Zn、Cu、Mn等微量元素均有检出。K、Fe、Zn等矿质元素含量低于异齿裂腹鱼 (Schizothoraxo connori)[32]。Ca含量显著高于褐点石斑鱼 (Epinephelus fuscoguttatus) 和青石斑鱼 (E. awoara) 等多种海鱼[31]。西江种群肌肉中Fe含量显著高于漠阳江和万泉河种群,而Zn含量则显著低于万泉河种群。Fe具有造血功能和促进人体生长的作用等,Zn可以促进儿童智力的正常发育[33]。3个广东鲂地理种群肌肉多种微量元素含量差异显著,这可能是由于栖息水环境的差异所致。万泉河种群相对西江种群,其主要栖息水域为河口,盐度相对较高,易受潮汐影响。有研究发现淡水环境中Fe含量均显著高于海水,Zn含量明显低于海水环境[34-35]。因此,栖息地环境差异导致了Zn在万泉河种群肌肉中富集度更高,Fe和Mn则在西江种群肌肉中富集度更高。
-
![]()
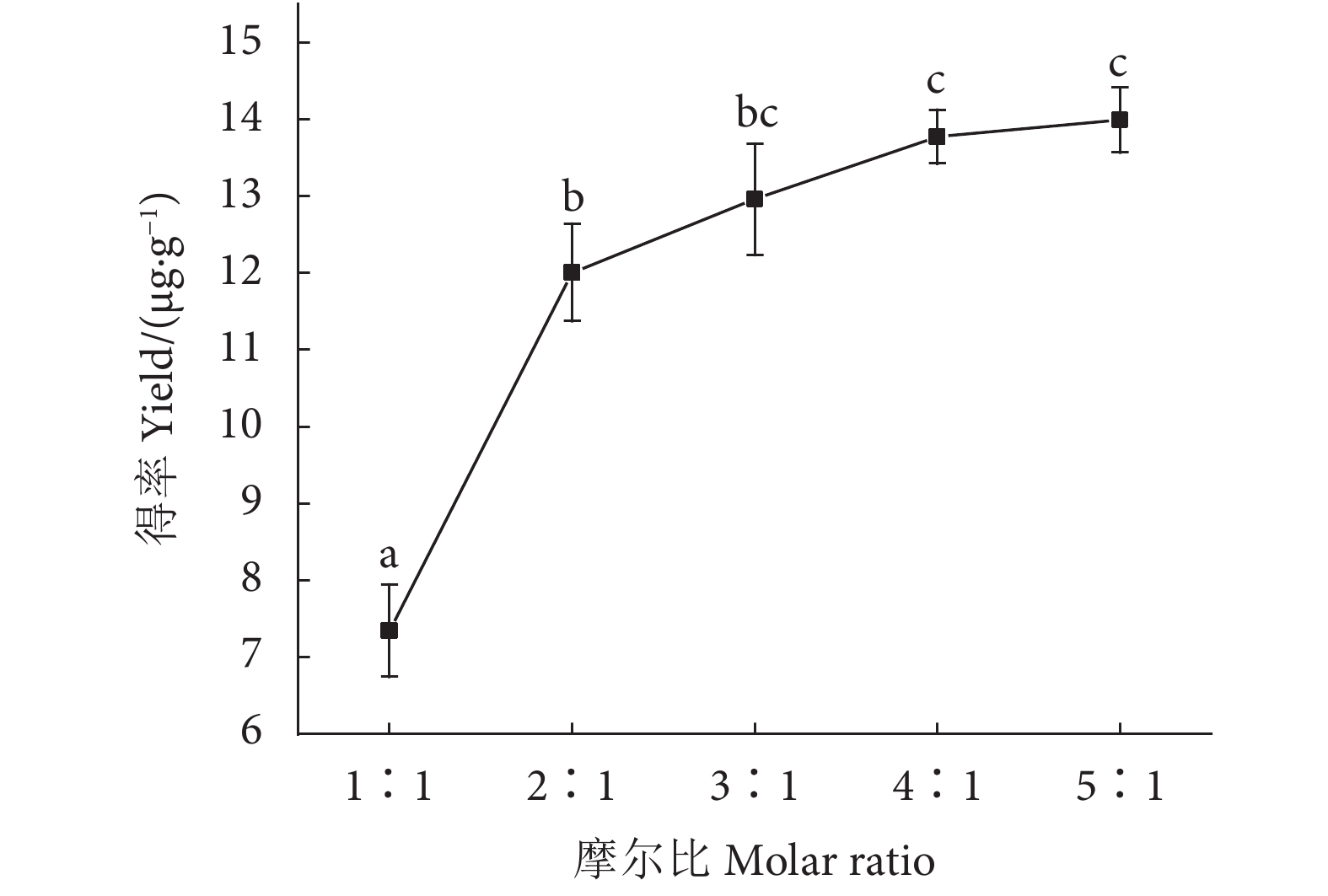
图 3 不同摩尔比的DES对虾青素得率的影响
注:不同字母表示差异显著 (P<0.05)。
Figure 3. Effect of DES with different molar ratio on astaxanthin extraction rate
Note: Different letters represent significant differences (P<0.05).
![]()
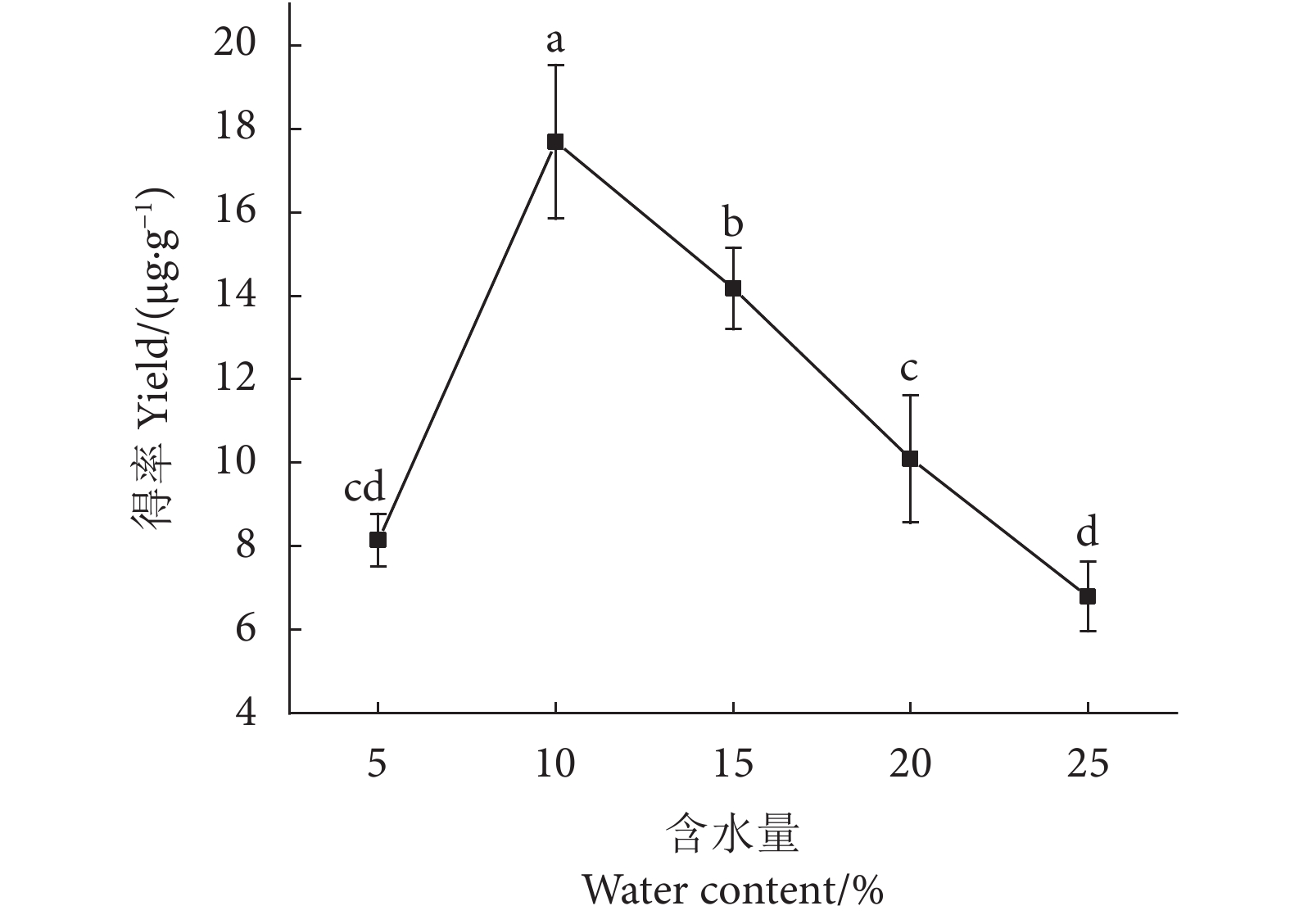
图 4 不同含水量的DES对虾青素得率的影响
注:不同字母表示差异显著(P<0.05)
Figure 4. Effect of DES with different water content on astaxanthin extraction rate
Note: Different letters represent significant differences (P<0.05).
![]()
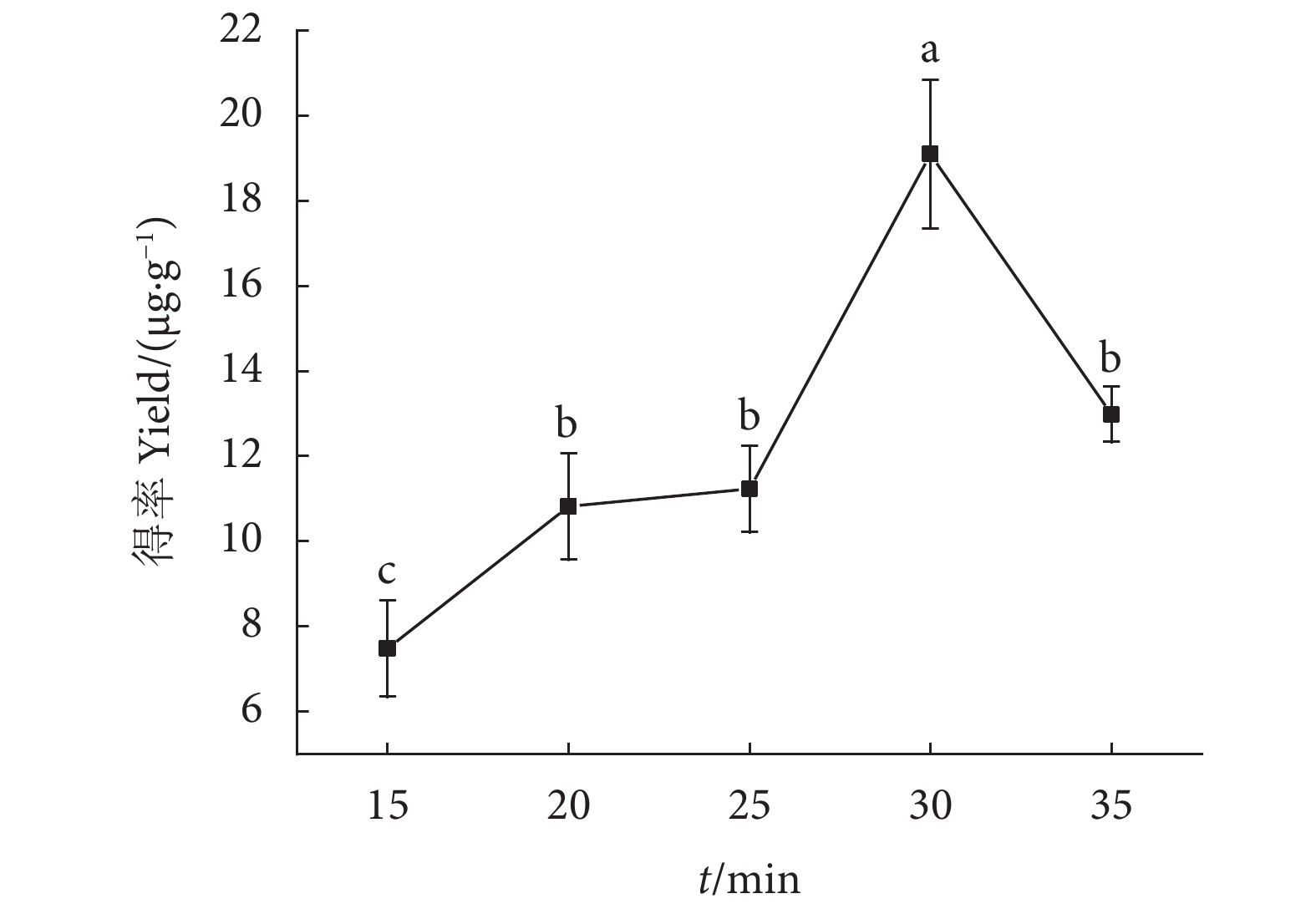
图 5 不同超声时间对虾青素得率的影响
注:不同字母表示差异显著(P<0.05)。
Figure 5. Effect of different ultrasonic time on astaxanthin extraction rate
Note: Different letters represent significant differences(P<0.05).
![]()
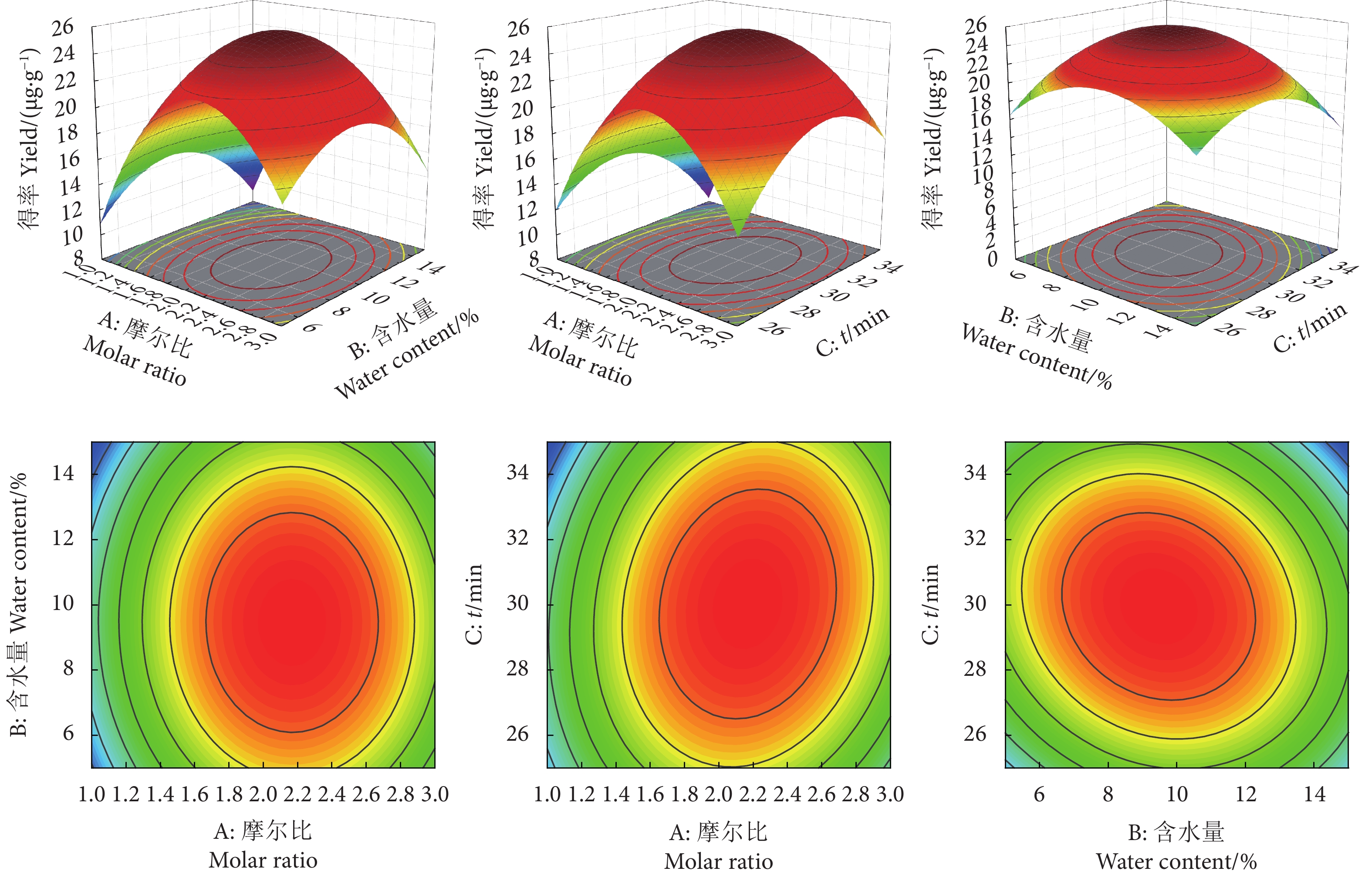
图 6 各因素交互作用对虾青素提取得率的影响
Figure 6. Effects of interaction of various factors on astaxanthin extraction rate
![]()
图 7 DES、乳酸和氯化胆碱傅里叶变换红外光谱图
Figure 7. FTIR spectra of DES, lactic acid and choline chloride
表 1 不同摩尔比及含水量的DES理化性质
Table 1 Physical and chemical properties of DES with different molar ratio and water content
DES 乳酸-氯化胆碱摩尔比
Molar ratio of lactic aicd-choline chloride含水量
Water content/%黏度
Viscosity/(mPa·s)熔融状态
Molten stateDES-1 1∶1 5 180.47±1.11g 澄清透明 DES-2 1∶1 10 75.33±0.40d 澄清透明 DES-3 1∶1 15 40.37±1.42b 澄清透明 DES-4 2∶1 5 122.23±1.32f 澄清透明 DES-5 2∶1 10 49.37±0.70c 澄清透明 DES-6 2∶1 15 30.47±1.98a 澄清透明 DES-7 3∶1 5 115.73±1.63e 澄清透明 DES-8 3∶1 10 48.13±0.80c 澄清透明 DES-9 3∶1 15 30.87±1.18a 澄清透明 注:不同字母表示差异显著 (P<0.05)。 Note: Different letters represent significant differences (P<0.05).  下载: 导出CSV
下载: 导出CSV
表 2 Box-Behnken实验设计及结果
Table 2 Design and results of Box-Behnken experiment
实验号
No.A:摩尔比
Molar ratioB:含水量
Water content/%C:时间
t/minY:得率
Yield/(μg·g−1)1 3∶1 10 25 13.54 2 2∶1 15 35 14.25 3 3∶1 10 35 16.55 4 2∶1 10 30 26.14 5 2∶1 10 30 25.89 6 2∶1 5 25 16.29 7 1∶1 10 35 9.02 8 1∶1 5 30 10.62 9 2∶1 5 35 18.07 10 1∶1 10 25 12.32 11 3∶1 5 30 17.40 12 3∶1 15 30 14.69 13 2∶1 10 30 24.67 14 2∶1 15 25 17.46 15 2∶1 10 30 26.72 16 2∶1 10 30 24.30 17 1 15 30 8.08
下载: 导出CSV
表 3 Box-Behnken实验回归模型方差分析
Table 3 AVOVA of Box-Behnken experiment
方差来源
Source平方和
Sum of squares自由度
Degree of freedom均方
Mean squareF P 显著性
Sig.模型 Model 591.36 9 65.71 58.46 <0.000 1 极显著 A 61.27 1 61.27 54.51 0.000 2 极显著 B 7.80 1 7.80 6.94 0.033 7 显著 C 0.369 8 1 0.369 8 0.3290 0.584 2 不显著 AB 0.007 2 1 0.007 2 0.0064 0.938 3 不显著 AC 9.95 1 9.95 8.86 0.020 6 显著 BC 6.23 1 6.23 5.54 0.050 8 不显著 A² 286.80 1 286.80 255.17 <0.000 1 极显著 B² 88.83 1 88.83 79.03 <0.000 1 极显著 C² 82.75 1 82.75 73.62 <0.000 1 极显著 残差 Residual 7.87 7 1.12 失拟项 Lack of fit 3.70 3 1.23 1.18 0.421 5 不显著 纯误差 Pure error 4.17 4 1.04 总和 Total 599.23 16
下载: 导出CSV
-
[1] CHANG M X, XIONG F. Astaxanthin and its effects in inflammatory responses and inflammation-associated diseases: recent advances and future directions[J]. Molecules, 2020, 25(22): 5342. doi: 10.3390/molecules25225342
[2] HAMA S, UENISHI S, YAMADA A, et al. Scavenging of hydroxyl radicals in aqueous solution by astaxanthin encapsulated in liposomes[J]. Biol Pharm Bull, 2012, 35(12): 2238-2242. doi: 10.1248/bpb.b12-00715
[3] 相欢, 刘宇航, 李来好, 等. 虾青素运载体系的研究进展[J]. 现代食品科技, 2023, 39(9): 364-370. [4] 闫娇, 高坤鹏, 余可欣, 等. 脂肪酶Sv-lip5的异源表达及其在虾青素酯水解中的应用[J]. 南方水产科学, 2022, 18(2): 31-38. doi: 10.12131/20210293 [5] 苏家齐, 祝华萍, 朱长波, 等. 盐度和钠离子/钾离子对凡纳滨对虾幼虾存活与组织结构的影响[J]. 南方水产科学, 2021, 17(5): 45-53. [6] 高芳芳, 王迪, 陈胜军, 等. 罗氏沼虾氨基脲分布特征及贮藏变化研究[J]. 南方水产科学, 2023, 19(1): 147-154. doi: 10.12131/20220138 [7] 农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2021中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2021: 24. [8] 全国水产技术推广总站, 中国水产学会. 中国小龙虾产业发展报告(2021)[J]. 中国水产, 2021(7): 27-33. [9] 侯会绒, 孙兆远, 贡汉坤. 超声波提取克氏原螯虾壳中虾青素[J]. 食品与发酵工业, 2015, 41(9): 209-214. [10] ZHAO T, YAN X J, SUN L J, et al. Research progress on extraction, biological activities and delivery systems of natural astaxanthin[J]. Trends Food Sci Technol, 2019, 91: 354-361. doi: 10.1016/j.jpgs.2019.07.014
[11] SHARAYEI P, AZARPAZHOOH E, ZOMORODI S, et al. Optimization of ultrasonic-assisted extraction of astaxanthin from green tiger (Penaeus semisulcatus) shrimp shell[J]. Ultrason Sonochem, 2021, 76: 105666. doi: 10.1016/j.ultsonch.2021.105666
[12] 周佳悦, 候艳丽, 王凡予, 等. 超声辅助低共熔溶剂提取红松树皮原花青素及动力学研究[J]. 食品工业科技, 2023, 44(14): 229-236. [13] LUCIANA I, VANESSA B, WANDERSON D, et al. Deep eutectic solvents for the production and application of new materials[J]. Appl Mater Today, 2018, 10: 30-50. doi: 10.1016/j.apmt.2017.11.005
[14] 张艺欣, 邬旭然, 何若菡, 等. 天然低共熔溶剂萃取酚类物质研究进展[J]. 食品与机械, 2022, 38(11): 212-216, 240. [15] CHEN Z, RAGAUSKAS A, WAN C X. Lignin extraction and upgrading using deep eutectic solvents[J]. Ind Crops Prod, 2020, 147: 112241. doi: 10.1016/j.indcrop.2020.112241
[16] BI W T, TIAN M L, ROW K H, et al. Evaluation of alcohol-based deep eutectic solvent in extraction and determination of flavonoids with response surface methodology optimization[J]. J Chromatogr A, 2013, 1285: 22-30. doi: 10.1016/j.chroma.2013.02.041
[17] ALAM M, MUHAMMAD G, KHAN M, et al. Choline chloride-based deep eutectic solvents as green extractants for the isolation of phenolic compounds from biomass[J]. J Clean Prod, 2021, 309: 127445. doi: 10.1016/j.jclepro.2021.127445
[18] ZHANG H, TANG B K, ROW K H. A green deep eutectic solvent-based ultrasound-assisted method to extract astaxanthin from shrimp byproducts[J]. Anal Lett, 2014, 47: 742-749. doi: 10.1080/00032719.2013.855783
[19] SANTOS C, PADILHA C, DAMASCENO K, et al. Astaxanthin recovery from shrimp residue by solvent ethanol extraction using choline chloride: glycerol deep eutectic solvent as adjuvant[J]. J Braz Chem Soc, 2021, 32(5): 1030-1039.
[20] 吴晓青, 刘永静, 孙燕丽. 低共熔溶剂提取橄榄多酚及其抗氧化活性评价[J]. 食品工业, 2023, 44(6): 80-86. [21] STACHOWIAK B, SZULC P. Astaxanthin for the food industry[J]. Molecules, 2021, 26(9): 2666. doi: 10.3390/molecules26092666
[22] 李海飞, 杨毅, 亓雨芮, 等. 超声波辅助酸性天然低共熔溶剂提取黑果腺肋花楸花青素及其稳定性和抗氧化活性分析[J]. 食品工业科技, 2023, 44(8): 259-269. [23] 杨塬兴. 低共熔溶剂分离蔗渣组分及其利用研究[D]. 广州: 华南理工大学, 2022: 27-28. [24] 梁珍珍. 基于氯化胆碱/二元醇的低共熔溶剂制备、理化性质及提取燕麦蛋白研究[D]. 西安: 陕西科技大学, 2022: 19-20. [25] 刘戈妹. 低共熔溶剂对玉米淀粉结构及理化性质影响的研究[D]. 西安: 陕西科技大学, 2023: 23-24. [26] LING J, CHAN Y S, NANDONG J, et al. Formulation of choline chloride/ascorbic acid natural deep eutectic solvent: characterization, solubilization capacity and antioxidant property[J]. LWT, 2020, 133: 110096. doi: 10.1016/j.lwt.2020.110096
[27] 张雁东, 吴晓静, 李子轩, 等. 温度扰动下氯化胆碱/丙三醇溶液的二维红外光谱研究[J]. 光谱学与光谱分析, 2023, 43(10): 3047-3051. [28] 张言, 尹静梅, 崔颖娜. 低共熔法在萃取酚类化合物中的应用进展[J]. 石油化工, 2022, 51(1): 92-99. doi: 10.3969/j.issn.1000-8144.2022.01.014 [29] FLORINDO C, BRANCO L C, MARRUCHO I M. Quest for green-solvent design: from hydrophilic to hydrophobic (deep) eutectic solvents[J]. Chemsuschem, 2019, 12(8): 1549-1559. doi: 10.1002/cssc.201900147
[30] LIU Y T, CHEN YA, XING Y J, et al. Synthesis and characterization of novel ternary deep eutectic solvents[J]. Chin Chem Lett, 2014, 25: 104-106. doi: 10.1016/j.cclet.2013.09.004
[31] 李苗, 陈必清, 何敏. 氯化胆碱-尿素低共熔离子液体的粘度和电导率[J]. 湖北大学学报(自然科学版), 2018, 40(1): 96-102. [32] 张荣清, 刘云云, 陈海峰, 等. 低共熔溶剂体系提取中药渣中肉桂醛[J]. 化学工程, 2023, 51(4): 22-27. [33] LEE Y R, ROW K H. Comparison of ionic liquids and deep eutectic solvents as additives for the ultrasonic extraction of astaxanthin from marine plants[J]. J Ind Eng Chem, 2016, 39: 87-92.
[34] ZOU T B, JIA Q, LI H W, et al. Response surface methodology for ultrasound-assisted extraction of astaxanthin from Haematococcus pluvialis[J]. Mar Drugs, 2013, 5: 1644-1655.
-
期刊类型引用(1)
1. 庄平,赵峰,罗刚,张涛,石小涛,冯广朋,王思凯. 水生生物资源增殖放流的发展历程与问题思考. 水生生物学报. 2025(01): 43-54 .  百度学术
百度学术
其他类型引用(0)




计量
- 文章访问数: 550
- HTML全文浏览量: 36
- PDF下载量: 33
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
