
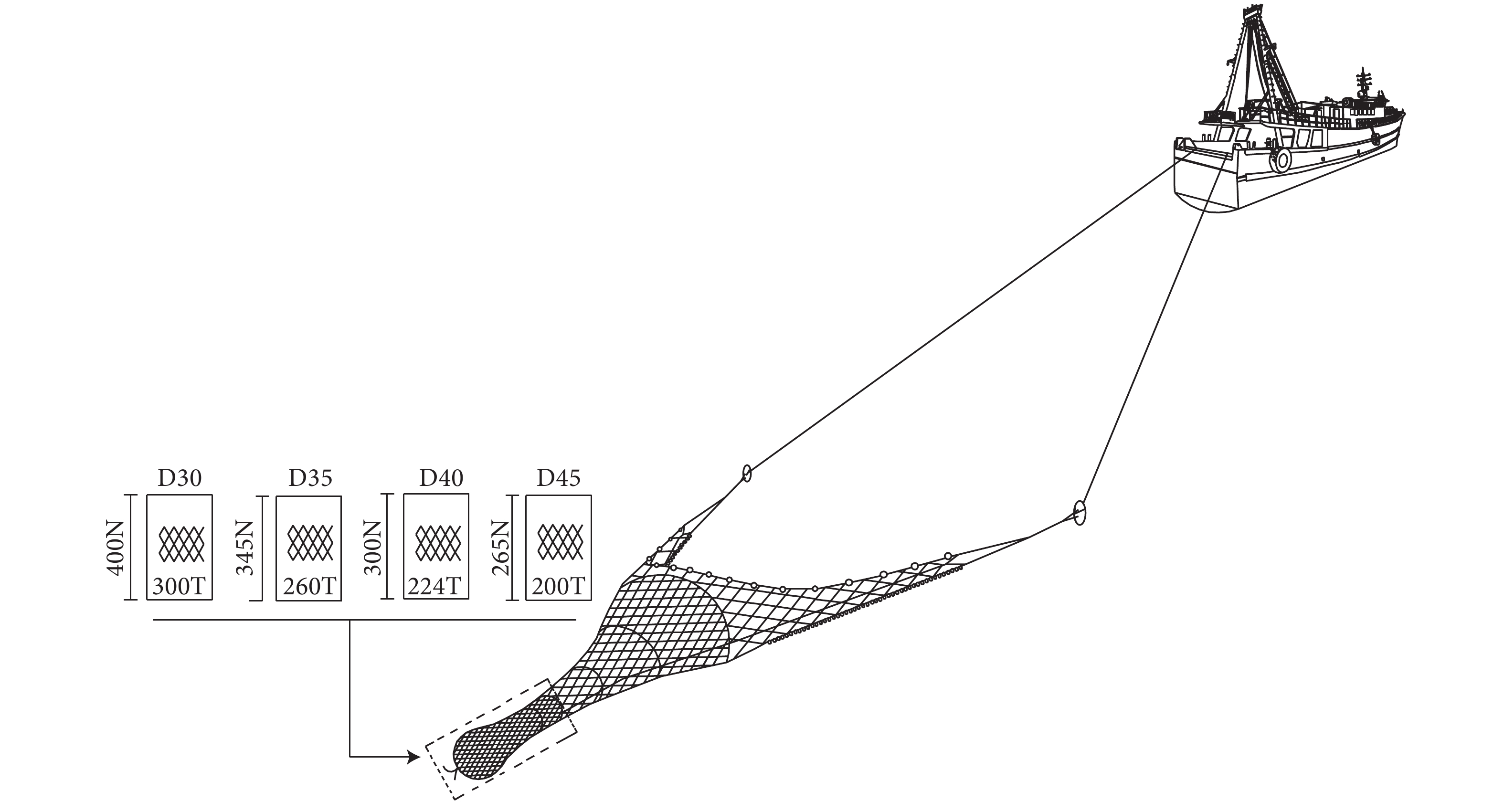
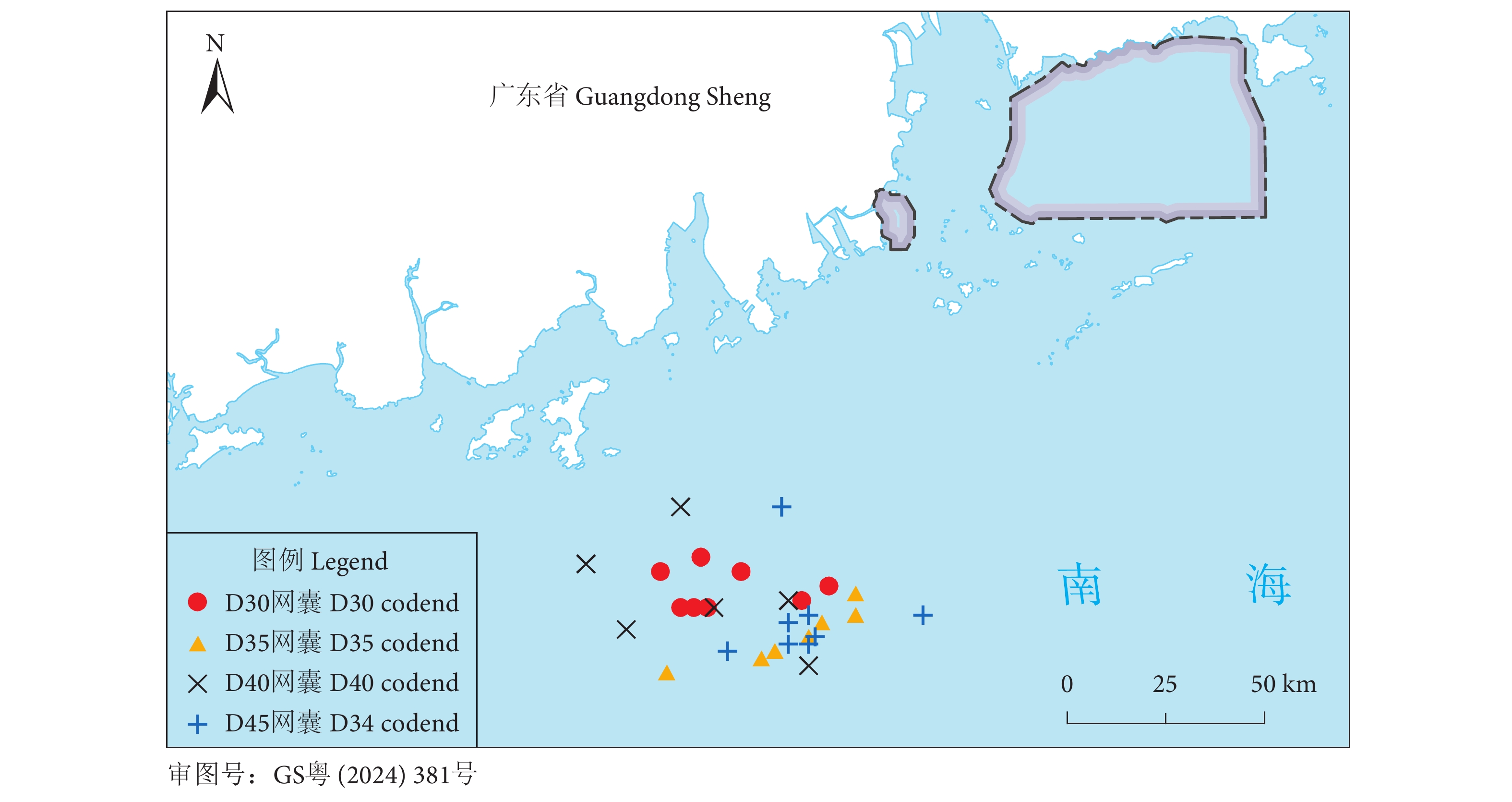
Size selectivity of codend-mesh size in single boat bottom otter trawls for Nemipterus virgatus in South China Sea
-
摘要:
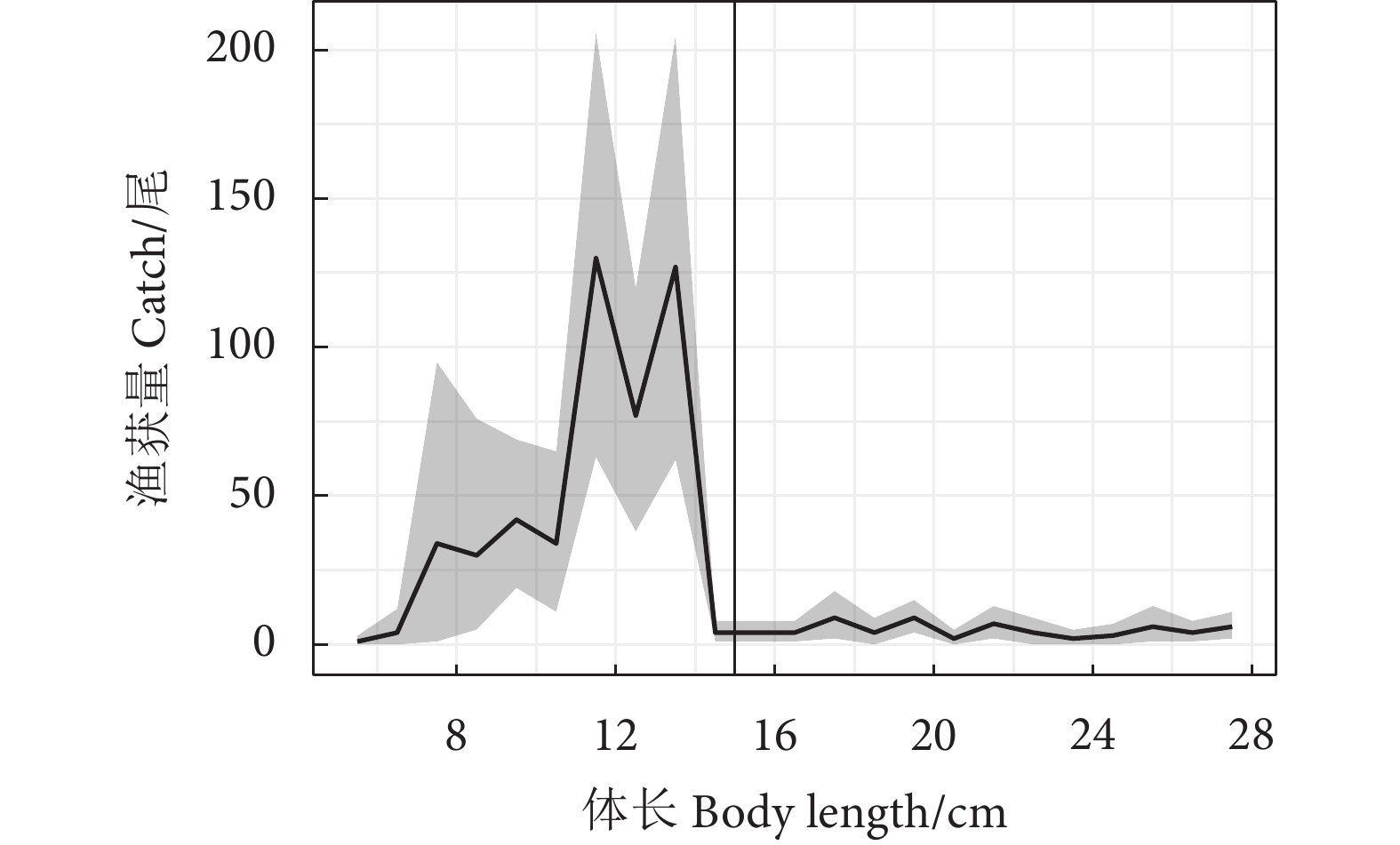
金线鱼 (Nemipterus virgatus) 是南海区拖网渔业的重要经济种类。为提高拖网对金线鱼的选择性,在南海区使用套网法对网目尺寸分别为30、35、40和45 mm的4组网囊开展了试验。使用双重拔靴法拟合网囊对金线鱼的选择性参数和选择性曲线,比较网囊间的选择性差异,并估算网囊的捕捞方式指数。结果显示,随着网目尺寸的增大,50%选择体长 (L50) 增大,选择性曲线的置信区间变窄,网囊对金线鱼幼鱼的渔获比例减小。试验网囊的L50均小于金线鱼的开捕规格 (15.0 cm),网囊对金线鱼的抛弃率大于67%。研究表明,增大网目尺寸能够优化网囊的选择性,但选择性仍需进一步提升,才能更好地促进金线鱼拖网渔业的可持续发展。
Abstract:Threadfin bream (Nemipterus virgatus) is an important species in the trawl fishery of the South China Sea. In order to improve the size selectivity and exploitation pattern of single boat bottom otter trawls for N. virgatus, we tested four experimental codends with mesh sizes of 30, 35, 40 and 45 mm by applying the covered codend method in the South China Sea. We analyzed the catch data with double-bootstrapping technique to estimate the selectivity parameters, selectivity curves, delta selectivity curves and exploitation pattern indicators. The results demonstrate that with increasing mesh sizes in codends, 50% retention length (L50) increased, confidence intervals of the selectivity curves became sharper, and retention fraction of undersized individuals decreased. However, when considering the minimum conservation reference size of the target species (15.0 cm), the L50 values of codends tested were less than 15.0 cm, and their discarded percentages were all over 67%. Though the study shows that increasing mesh sizes will improve size selectivity, it should be further improved to achieve a sustainable trawl fishery for threadfin bream.
-
Keywords:
- Trawl /
- Codend mesh /
- Size selectivity /
- Nemipterus virgatus
-
由于鱼类染色体相较于其他脊椎动物具有较强的“可塑性”,所以鱼类最易进行杂交[1]。远缘杂交是一种应用广泛且效果较显著的育种方法,在杂交种的优质利用、诱导雌核发育以及抗逆性能的选育[2-4]等方面发挥着重要作用。虽然现在各种新育种手段不断应用于鱼类遗传育种,但是杂交育种仍是应用广泛且效果较为明显的一种育种方法。通过杂交育种可以使不同品种之间优良性状结合,研究显示很多杂交种均表现出了明显的杂种优势[5-12]。一般来说,分布地区距离越远、外部形态差异越大、基因型纯化程度越高的种间杂交,出现杂交优势的可能性越高。
短须裂腹鱼(Schizothorax wangchiachii)属鲤形目、鲤科、裂腹鱼亚科、裂腹鱼属,为长江上游特有鱼类,是金沙江及其支流、雅砻江和乌江的主要经济鱼类之一[13],其肉质细嫩、味道鲜美、营养丰富、深受青睐。近十多年来,野生短须裂腹鱼因过度捕捞、环境污染、水电站开发等,资源量越来越少。刘跃天等[14]于2006年首次报道短须裂腹鱼人工繁殖成功,获鱼苗7 508尾。随后由于其经济效益尚且产卵率和孵化成功率较高[15-17],人工繁殖数量越来越多,但在养殖过程中发现其易受小瓜虫、水霉病等侵袭,从而带来严重的经济损失。鲈鲤(Percocypris pingipingi)属鲤形目、鲤科、鲃亚科、鲈鲤属,主要分布于黔、川、滇长江上游及其支流,以及广西右江、珠江水系上游[18],是我国特有的珍稀野生鱼类,也是四川省和重庆市的重点保护动物[13]。鲈鲤的抗病能力较强,但在人工繁殖过程中产卵率和孵化成功率较低。
关于短须裂腹鱼与鲈鲤的胚胎及胚后发育均有报道[19-24],但目前尚未见有关短须裂腹鱼和鲈鲤杂交后代胚胎及仔稚鱼发育的研究报道。本研究利用隶属不同亚科的鲈鲤与短须裂腹鱼杂交,系统地观察了杂交F1代的胚胎及仔稚鱼发育,记录其发育特征及生长指数,旨在为短须裂腹鱼和鲈鲤的杂交育种以及今后优良性状养殖品种的选育提供基础资料。
1. 材料与方法
1.1 材料与培育条件
实验于2017年3月在凉山科华水生态工程有限公司进行,实验用短须裂腹鱼、鲈鲤亲鱼均为3~4龄,体质量为1 500~2 000 g,短须裂腹鱼(♀)×鲈鲤(♂) (DL)母本短须裂腹鱼的怀卵量约19 000粒,鲈鲤(♀)×短须裂腹鱼(♂) (LD)母本鲈鲤的怀卵量约12 000粒。
实验鱼养殖于约25 m2的鱼塘中,水温(14.5±1.0) ℃,pH 8.10~8.52,分别对性成熟的短须裂腹鱼和鲈鲤雌雄个体注射人绒毛膜促性腺激素(HCG)和促黄体激素释放激素类似物(LHRH-A2)人工催产,48~72 h后取成熟亲鱼轻压腹部,顺产获得成熟卵和精子,采用干法授精进行杂交。
分别将短须裂腹鱼卵和鲈鲤的精液,以及短须裂腹鱼的精液和鲈鲤的卵进行干法授精,待受精卵充分吸水膨胀后,在直径74 cm、高28 cm的孵化盒中流水孵化培育,密度为2 000~3 000粒·m–2,流量为60~70 mL·s–1,温度(14.5±1.0) ℃,pH 8.10~8.81,出膜后的仔鱼饲养于淡水鱼类资源与生殖发育教育部重点实验室室内循环系统,定期换水并排污。早期仔鱼以丰年虫为开口饵料,后混合投喂鱼苗专用微粒配合饲料S1,饵料和饲料分别购自山东爱家宠物水族用品有限公司和山东升索渔用饲料研究中心。
1.2 实验方法
利用体式显微镜(Nikon 6000)对DL和LD的胚胎及仔稚鱼发育过程进行观察拍照。参考国内外学者的研究[19-28],胚胎发育过程中每次随机取30~60粒胚胎,前12 h内每10 min取样1次,后每0.5~1 h取样1次,取样至出膜为止,胚胎发育的时间以60%以上的胚胎明显进入该发育阶段为准,观察期间水温为(14.5±1.0) ℃;仔稚鱼阶段每次随机取10尾,前20 d取样每天1次,之后间隔5 d取样1次,取样至鳞被覆盖完毕,用稀释的乙醇麻醉,同样采用体式显微镜进行观察、拍照并记录,然后测定其生长指数。
1.3 发育积温算法
以有效积温描述整个发育过程[29],各阶段积温的计算方法为:胚胎发育各阶段的积温(h·℃)=该阶段胚胎发育时间(h)×水温(℃)。
1.4 数据处理
使用Excel 2017对实验数据进行处理,利用Photoshop CS5对图像进行编辑。
2. 结果
2.1 短须裂腹鱼(♀) ×鲈鲤(♂)胚胎及仔稚鱼发育
2.1.1 胚胎发育
短须裂腹鱼的成熟卵为浅黄色、沉性、微黏性。在平均水温(14.5±1.0) ℃条件下,胚胎发育历时144.33 h,受精率为95.33%,孵化率为71.12%,参考短须裂腹鱼与鲈鲤的研究结果[19-24],将其胚胎发育过程分为6个阶段,共28个时期(表 1)。
表 1 短须裂腹鱼与鲈鲤杂交F1 代胚胎发育时序表Table 1 Embryonic development schedule of hybrid of S. wangchiachii and P. pingipingi胚胎发育时期
embryonic developmental stage主要特征
developmental characteristics短须裂腹鱼 (♀) ×鲈鲤 (♂)
S. wangchiachii (♀) ×
P. pingipingi (♂) (DL)鲈鲤 (♀) ×短须裂腹鱼 (♂)
P. pingipingi (♀) ×
S. wangchiachii (♂) (LD)时间
time积温/h·℃
accumulated temperature时间
time积温/h·℃
accumulated temperature受精卵阶段
fertilized egg period受精卵期 受精卵呈球形,卵间隙小 0 0 0 0 吸水膨胀期 受精卵吸水膨胀,卵间隙增大,卵膜呈透明状 45 min 10.88 1 h 1 min 14.79 胚盘隆起期 原生质向动物极集中并隆起形成胚盘 3 h 35 min 51.77 3 h 7 min 54.81 卵裂阶段
cleavage period2细胞期 胚盘表面凹陷形成分裂沟,将胚盘分裂为大小相似的2个分裂球 4 h 50 min 69.89 4 h 11 min 60.76 4细胞期 分裂球再次分裂,形成大小相似的4个细胞 5 h 44 min 83.09 4 h 47 min 69.31 8细胞期 分裂球呈几何级数增加,形成大小相似的8个细胞 6 h 30 min 94.25 5 h 44 min 83.09 16细胞期 继续分裂,形成16个细胞 7 h 53 min 114.41 6 h 39 min 96.43 桑葚胚期 卵裂速度加快,分裂球越来越小,细胞界限开始模糊,形成多细胞胚体,呈桑葚状 16 h 50 min 244.18 9 h 51 min 142.83 囊胚阶段
blastula period囊胚早期 细胞团高高隆起,形似帽状,细胞界限模糊不清 19 h 47 min 286.81 16 h 43 min 242.44 囊胚中期 细胞层向下扩散,囊胚高度下降,呈小丘状,胚层边缘逐渐平滑 21 h 1 min 304.79 18 h 40 min 270.72 囊胚晚期 囊胚继续下降,与卵黄囊形成一个近球形 26 h 23 min 382.51 20 h 9 min 292.18 原肠胚阶段
gastrula period原肠早期 胚层下包卵黄囊约1/2 42 h 45 min 620.02 38 h 34 min 559.27 原肠中期 胚盾出现,胚层下包卵黄囊约2/3 49 h 53 min 723.41 42 h 28 min 615.67 原肠晚期 胚层下包卵黄囊约3/4 53 h 3 min 769.23 51 h 15 min 743.13 器官形成阶段
organogenesis period神经胚期 胚体可见,神经板雏形出现 55 h 19 min 802.14 − − 肌节出现期 胚体前段微隆起,中部出现肌节,胚孔仍可见 58 h 50 min 853.04 − − 眼囊期 眼囊出现,呈长梭形 61 h 15 min 888.13 − − 耳囊期 胚体眼囊后两侧出现一对卵圆形耳囊 68 h 37 min 997.99 − − 胚孔封闭期 胚层将卵黄栓完全包裹,胚孔封闭,胚体明显且超过卵黄囊一半 72 h 15 min 1 047.63 − − 肌肉效应期 胚体开始扭动,作缓慢的颤动式收缩 75 h 23 min 1 093.16 − − 尾芽期 尾端略突出,游离于卵黄,尾芽出现 79 h 8 min 1 147.39 − − 晶体形成期 眼囊中出现圆形、透明晶体 96 h 4 min 1 393.02 − − 耳石期 耳囊增大,其内可见2个小点,为耳石 101 h 16 min 1 468.27 − − 围心腔期 头部与卵黄连接处出现围心腔 109 h 22 min 1 586.01 − − 心脏原基期 围心腔内可见呈短管状的心脏原基 116 h 16 min 1 685.05 − − 心跳期 心脏开始有节律地搏动,频率52~
58 次·min–1121 h 22 min 1 614.72 − − 出膜阶段
incubation period出膜前期 胚体运动加剧,运动幅度加大 139 h 59 min 2 029.71 − − 出膜期 胚体尾部击破卵膜,尾部伸出膜外,胚体脱离卵膜 144 h 20 min 2 092.79 − − 1) 受精卵阶段。刚受精时,受精卵呈黄色球形,卵径约3.24 mm (图1-1);然后开始吸水膨胀,卵周隙增大,受精后45 min,受精卵吸水膨胀到最大,直径约为4.05 mm,卵膜呈透明状(图1-2)。受精后3 h 35 min,原生质不断向动物极集中并隆起形成类似盘状结构,即为胚盘,此时进入胚盘期(图1-3)。
![]() 图 1 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 胚胎发育1. 受精卵期;2. 吸水膨胀;3. 胚盘隆起期;4. 2 细胞期;5. 4 细胞期;6. 8 细胞期;7. 16 细胞期;8. 桑葚胚期;9. 囊胚早期;10. 囊胚中期;11. 囊胚晚期;12. 原肠早期;13. 原肠中期;14. 原肠晚期;15. 神经胚期;16. 肌节出现期;17. 眼囊期;18. 耳囊期;19. 胚孔封闭期;20. 肌肉效应期;21. 尾芽期;22. 晶体形成期;23. 耳石期;24. 围心腔期;25. 心脏原基期;26. 心跳期;27. 出膜前期;28. 出膜期;标尺=1 mmFig. 1 Embryonic development of hybrid S. wangchiachii (♀)×P. pingiping (♂)1. fertilized egg; 2. swelling stage; 3. blastoderm stage; 4. 2-cell stage; 5. 4-cell stage; 6. 8-cell stage; 7. 16-cell stage; 8. multi-cell stage; 9. early-blastula stage; 10. mid-blastula stage, 11. late-blastula stage; 12. early-gastrula stage; 13. mid-gastrula stage; 14. late-gastrula stage; 15. neurula stage; 16. muscle burl stage; 17. eye vesicle stage; 18. otocyst stage; 19. blastopore closing stage; 20. muscular contraction stage; 21. tailbud formed stage; 22. eye lens formed stage; 23. otolithes stage; 24. pericardiac coelom stage; 25. rudiment of heart stage; 26. heart-beating stage; 27. pre-hatching stage; 28. hatching stage; bar=1 mm
图 1 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 胚胎发育1. 受精卵期;2. 吸水膨胀;3. 胚盘隆起期;4. 2 细胞期;5. 4 细胞期;6. 8 细胞期;7. 16 细胞期;8. 桑葚胚期;9. 囊胚早期;10. 囊胚中期;11. 囊胚晚期;12. 原肠早期;13. 原肠中期;14. 原肠晚期;15. 神经胚期;16. 肌节出现期;17. 眼囊期;18. 耳囊期;19. 胚孔封闭期;20. 肌肉效应期;21. 尾芽期;22. 晶体形成期;23. 耳石期;24. 围心腔期;25. 心脏原基期;26. 心跳期;27. 出膜前期;28. 出膜期;标尺=1 mmFig. 1 Embryonic development of hybrid S. wangchiachii (♀)×P. pingiping (♂)1. fertilized egg; 2. swelling stage; 3. blastoderm stage; 4. 2-cell stage; 5. 4-cell stage; 6. 8-cell stage; 7. 16-cell stage; 8. multi-cell stage; 9. early-blastula stage; 10. mid-blastula stage, 11. late-blastula stage; 12. early-gastrula stage; 13. mid-gastrula stage; 14. late-gastrula stage; 15. neurula stage; 16. muscle burl stage; 17. eye vesicle stage; 18. otocyst stage; 19. blastopore closing stage; 20. muscular contraction stage; 21. tailbud formed stage; 22. eye lens formed stage; 23. otolithes stage; 24. pericardiac coelom stage; 25. rudiment of heart stage; 26. heart-beating stage; 27. pre-hatching stage; 28. hatching stage; bar=1 mm2) 卵裂阶段。受精后4 h 50 min,胚盘向两边拉伸,其表面凹陷形成分裂沟,随着发育不断加深,分裂沟将胚盘一分为二,形成大小相似的2个细胞,即为2细胞期(图1-4);受精后5 h 44 min分裂球再次分裂,分裂沟与第一次分裂沟垂直,形成大小相似的4个细胞,进入4细胞期(图1-5);分裂球继续呈几何级数增加,受精后6 h 30 min形成大小相似的8个细胞,即为8细胞期(图1-6);受精后7 h 53 min分裂形成16个细胞,即为16细胞期(图1-7);随后分裂速度加快,依次形成32个细胞、64个细胞,随着细胞数目增多,细胞界限开始模糊,受精后16 h 50 min形成多细胞胚体,呈桑葚状,即为桑葚胚期(图1-8)。
3) 囊胚阶段。细胞继续分裂,细胞层数和数量不断增加,受精后19 h 47 min细胞团高高隆起,形似帽状,细胞界限模糊不清,即为囊胚早期(图1-9);隆起的细胞层向下扩散,逐渐变低,并开始下包,囊胚高度下降,受精后21 h 1 min呈小丘状,胚层边缘逐渐平滑,即为囊胚中期(图1-10);囊胚继续下包,受精后26 h 23 min囊胚细胞紧贴卵黄,与卵黄一起近似球形,进入囊胚晚期(图1-11)。
4) 原肠胚阶段。随着细胞的进一步分裂,动物极细胞数目越来越多并逐渐向植物极方向延伸,受精后42 h 45 min胚层下包卵黄囊约1/2,即为原肠胚早期(图1-12);受精后49 h 53 min胚盾出现,胚层下包卵黄囊约2/3,为原肠胚中期(图1-13);受精后53 h 3 min胚层下包卵黄囊3/4,进入原肠胚晚期(图1-14)。
5) 器官形成阶段。受精后55 h 19 min胚层即将包完卵黄,胚体可见,神经板雏形出现,此时为神经胚期(图1-15);受精后58 h 50 min进入肌节形成期,胚体前段微微隆起,中部出现约8节肌节(图1-16),此时胚孔仍可见;受精后61 h 15 min头部两侧分化出2个眼囊,呈长梭形,即为眼囊期(图1-17);受精后68 h 37 min肌节进一步增多,约20节,胚体眼囊后两侧出现一对卵圆形耳囊,比眼囊小,进入耳囊期(图1-18);受精后72 h 15 min胚层将卵黄栓完全包裹,胚孔封闭,胚体长超过卵黄囊一半,此时为胚孔封闭期(图1-19),在胚孔封闭期观察到轻微的卵黄运动(图 2)。受精后75 h 23 min胚体开始扭动,频率和幅度都较小,约15次·min–1,作缓慢的颤动式收缩,进入肌肉效应期(图1-20);受精后79 h 8 min尾端略突出,游离于卵黄,尾芽出现,即为尾芽期(图1-21),此时胚体运动加剧,约40次·min–1;受精后96 h 4 min眼囊中出现圆形、透明晶体,进入晶体形成期(图1-22);受精后101 h 16 min耳囊增大,其内可见2个小点,为耳石,胚体发育至耳石期(图1-23);受精后109 h 22 min头部与卵黄连接处出现空的围心腔,即为围心腔期(图1-24);受精后116 h 16 min围心腔增大,其内可见呈短管状的心脏原基,即为心脏原基期(图1-25);受精后121 h 22 min心脏进入心跳期,开始有节律地搏动,频率为52~58次·min–1 (图1-26)。
![]() 图 2 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 代卵黄运动过程Fig. 2 Yolk movement process of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)
图 2 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 代卵黄运动过程Fig. 2 Yolk movement process of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)6) 出膜阶段。受精后139 h 59 min胚体运动剧烈,运动幅度大,胚体即将出膜,此时为出膜前期(图1-27);受精后144 h 20 min胚体尾部将卵膜击破,尾部随即伸出卵膜外,伴随着不停摆动,胚体逐渐脱离卵膜,即为出膜期,初孵仔鱼长度约为(10.85±0.21) mm (图1-28)。
2.1.2 短须裂腹鱼(♀) ×鲈鲤(♂)仔稚鱼发育
1) 仔稚鱼发育特征。短须裂腹鱼(♀) ×鲈鲤(♂)杂交的F1代仔稚鱼发育良好(图3),参照国内外的划分方法将其发育时期划分为3个阶段:卵黄囊期仔鱼(刚出膜仔鱼至卵黄囊消失时),历时17 d;晚期仔鱼(从卵黄囊消失至鳍条发育完整),历时45 d;稚鱼期(从鳍条发育完成至鳞被覆盖完毕),历时105 d (表 2)。
![]() 图 3 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 代仔稚鱼发育1. 1 d 仔鱼;2. 2 d 仔鱼;3. 3 d 仔鱼;4. 4 d 仔鱼;5. 5 d 仔鱼;6. 6 d 仔鱼;7. 7 d 仔鱼;8. 8 d 仔鱼;9. 9 d 仔鱼;10. 10 d 仔鱼;11. 11 d 仔鱼;12. 12 d 仔鱼;13. 13 d 仔鱼;14. 14 d 仔鱼;15. 15 d 仔鱼;16. 16 d 仔鱼;17. 17 d 仔鱼;18. 45 d 仔鱼;19. 105 d 稚鱼;标尺=1 mmFig. 3 Larvae and juveniles development of hybrid S. wangchiachii (♀)×P. pingipingi (♂)1. 1 d larvae; 2. 2 d larvae; 3. 3 d larvae; 4. 4 d larvae; 5. 5 d larvae; 6. 6 d larvae; 7. 7 d larvae; 8. 8 d larvae; 9. 9 d larvae; 10. 10 d larvae; 11.11 d larvae; 12. 12 d larvae; 13. 13 d larvae; 14. 14 d larvae; 15. 15 d larvae; 16. 16 d larvae; 17. 17 d larvae; 18. 45 d larvae; 19. 105 d juveniles; bar=1 mm表 2 短须裂腹鱼 (♀) ×鲈鲤 (♂) 仔稚鱼发育特征表Table 2 Larval and juvenile development schedule of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)
图 3 短须裂腹鱼 (♀) ×鲈鲤 (♂) 的F1 代仔稚鱼发育1. 1 d 仔鱼;2. 2 d 仔鱼;3. 3 d 仔鱼;4. 4 d 仔鱼;5. 5 d 仔鱼;6. 6 d 仔鱼;7. 7 d 仔鱼;8. 8 d 仔鱼;9. 9 d 仔鱼;10. 10 d 仔鱼;11. 11 d 仔鱼;12. 12 d 仔鱼;13. 13 d 仔鱼;14. 14 d 仔鱼;15. 15 d 仔鱼;16. 16 d 仔鱼;17. 17 d 仔鱼;18. 45 d 仔鱼;19. 105 d 稚鱼;标尺=1 mmFig. 3 Larvae and juveniles development of hybrid S. wangchiachii (♀)×P. pingipingi (♂)1. 1 d larvae; 2. 2 d larvae; 3. 3 d larvae; 4. 4 d larvae; 5. 5 d larvae; 6. 6 d larvae; 7. 7 d larvae; 8. 8 d larvae; 9. 9 d larvae; 10. 10 d larvae; 11.11 d larvae; 12. 12 d larvae; 13. 13 d larvae; 14. 14 d larvae; 15. 15 d larvae; 16. 16 d larvae; 17. 17 d larvae; 18. 45 d larvae; 19. 105 d juveniles; bar=1 mm表 2 短须裂腹鱼 (♀) ×鲈鲤 (♂) 仔稚鱼发育特征表Table 2 Larval and juvenile development schedule of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)时间
time特征
characteristic全长/mm
total length体长/mm
body length图序
No. of picture第1天 1st day 内源性营养,卵黄囊占鱼体比值较大,前段膨大呈球状,大小约1.96 mm×1.76 mm,头部与球状卵黄囊夹角约为30°,后端呈棒状,仔鱼活动较少,多卧于水底 11.36±0.26 8.47±0.31 图3-1 第2天 2nd day 前卵黄囊与头部夹角约45°,心脏搏动微弱,心率约55 次·min–1,心脏附近可见红色血液流动,耳石明显 11.49±0.27 8.53±0.24 图3-2 第3天 3rd day 前卵黄囊与头部夹角约为90°,眼球出现黑色素 12.71±0.13 9.02±0.12 图3-3 第4天 4th day 眼球黑色素明显增多,卵黄囊体积进一步缩小 12.98±0.29 9.68±0.23 图3-4 第5天 5th day 卵黄囊上出现一条明显的血管,胸鳍原基出现,上下颌开始张合,约8 次·min–1 13.37±0.57 9.78±0.48 图3-5 第6天 6th day 鳃可见明显红色血液流动 14.12±0.42 10.11±0.37 图3-6 第7天 7th day 背部出现少量呈星芒状的黑色素,全身血液颜色加深,心脏搏动有力,心率约75 次·min–1 14.41±0.36 10.28±0.44 图3-7 第8天 8th day 胸鳍形成,背鳍原基出现,上下颌张合明显,约35 次·min–1 14.79±0.59 10.49±0.61 图3-8 第9天 9th day 上下颌张合剧烈,约97 次·min–1,心率85 次·min–1 15.04±0.64 10.52±0.41 图3-9 第10天 10th day 胸鳍充分伸展,可摆动,头顶和背部黑色素增多,卵黄囊上也出现黑斑,心跳更剧烈,约105 次·min–1 15.09±0.68 10.54±0.52 图3-10 第11天 11th day 尾部出现少量黑色素,尾鳍下叶出现4~8个鳍条 15.18±0.32 10.58±0.31 图3-11 第12天 12th day 尾部黑色素增多,游泳迅速,鳔充气 15.33±0.48 10.61±0.36 图3-12 第13天 13th day 背鳍原基隆起呈三角形,卵黄囊吸收约1/2 15.45±0.66 10.66±0.57 图3-13 第14天 14th day 尾鳍鳍条分支增多,约8~14个 15.85±0.61 10.82±0.48 图3-14 第15天 15th day 卵黄囊吸收约2/3 15.95±0.72 11.07±0.57 图3-15 第16天 16th day 背鳍出现2~4个鳍条 16.02±0.57 11.13±0.46 图3-16 第17天 17th day 卵黄消耗完,尾鳍鳍膜边缘开始内凹,肠道有明显排遗,进入后期仔鱼 16.11±0.43 11.19±0.36 图3-17 第45天 45th day 胸鳍、尾鳍、背鳍、臀鳍、腹鳍鳍条基本发育完整,躯干呈半透明状,进入稚鱼期 25.84±0.82 19.46±0.62 图3-18 第105天 105th day 鳞被已覆盖完毕,背部呈青黑色,腹部灰白色,已基本具备成鱼的身体特征,进入幼鱼期 56.74±0.53 45.87±0.67 图3-19 2) 仔稚鱼生长情况。初孵仔鱼全长为(10.85±0.21) mm,体长为(8.23±0.18) mm,出膜第17天仔鱼全长为(16.11±0.43) mm,体长为(11.19±0.36) mm,出膜第105天幼鱼全长为(56.74±0.53 mm),体长为(45.87±0.67) mm,总体呈明显增长趋势(图 4)。
![]() 图 4 短须裂腹鱼 (♀) ×鲈鲤 (♂) 仔稚鱼生长指数图Fig. 4 Growth index of larvae and juveniles of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)
图 4 短须裂腹鱼 (♀) ×鲈鲤 (♂) 仔稚鱼生长指数图Fig. 4 Growth index of larvae and juveniles of hybrid of S. wangchiachii (♀) and P. pingipingi (♂)2.2 鲈鲤(♀) ×短须裂腹鱼(♂)胚胎发育
鲈鲤的成熟卵为金黄色、沉性、微黏性。在平均水温(14.5±1.0) ℃条件下,前期发育较正常,发育至原肠期所需时间为51 h 15 min,所需积温为743.13 h·℃ (表 1)。
2.2.1 囊胚期之前
刚受精时,受精卵呈球形,卵径约为2.32 mm,然后开始吸水膨胀,受精后1 h 1 min,受精卵吸水膨胀到最大,直径约为3.26 mm,此后发育过程除时间长短外均与正交一致。
2.2.2 囊胚期之后
囊胚期开始死亡个体增多,原肠期末有85.83%的胚胎死亡,原因多为下包过程中卵黄膜破裂,卵黄物质溢出导致胚胎死亡,另外在囊胚期和原肠期卵黄内出现异常团块(图 5)。经过原肠期的胚胎,个体之间发育时间相差较大,最后仅12尾出膜,且出膜后的仔鱼均畸形,多为围心腔肿大、体轴弯曲和棒状卵黄囊细短或无(图 5),出膜3 d后畸形个体全部死亡。
![]() 图 5 鲈鲤 (♀) ×短须裂腹鱼 (♂) F1 代死亡胚胎和仔鱼a. 卵黄外出现不明团块;b. 卵黄膜破裂,卵黄物质溢出;c. 围心腔肿大仔鱼;d. 棒状卵黄囊细短仔鱼;e. 体轴弯曲仔鱼;标尺=1 mmFig. 5 Dead embryos and larvae of hybrid of P. pingipingi (♀) and S. wangchiachii (♂)a. an unidentified mass appears outside the yolk; b. membrane of yolk rupture; yolk is leaking; c. pericardial swelling in the larvae; d. the clavicularyolk is short; e. the body axis of fish bends; bar=1 mm
图 5 鲈鲤 (♀) ×短须裂腹鱼 (♂) F1 代死亡胚胎和仔鱼a. 卵黄外出现不明团块;b. 卵黄膜破裂,卵黄物质溢出;c. 围心腔肿大仔鱼;d. 棒状卵黄囊细短仔鱼;e. 体轴弯曲仔鱼;标尺=1 mmFig. 5 Dead embryos and larvae of hybrid of P. pingipingi (♀) and S. wangchiachii (♂)a. an unidentified mass appears outside the yolk; b. membrane of yolk rupture; yolk is leaking; c. pericardial swelling in the larvae; d. the clavicularyolk is short; e. the body axis of fish bends; bar=1 mm3. 讨论
3.1 短须裂腹鱼(♀) ×鲈鲤(♂)的F1胚胎发育特征
温度是鱼类胚胎发育过程中的重要因素。左鹏翔等[22]在(19±1) ℃条件下观察的鲈鲤胚胎发育时间为101 h 40 min,赖见生等[23]在(18.0±0.5) ℃时观察的鲈鲤胚胎发育时间为126 h 28 min,王永明等[24]在(15.0±0.7) ℃时观察鲈鲤胚胎发育时间为164 h;左鹏翔等[20]在(14±1) ℃条件下观察的短须裂腹鱼的胚胎发育时间为254 h 40 min,刘阳等[19]在12.7~14.0 ℃时观察到短须裂腹鱼胚胎发育时间为192 h。由此可见,适当的温度范围内,两亲本胚胎发育时间受发育温度的影响,且温度越高,发育时间越短。本研究在水温(14.5±1.0) ℃条件下,短须裂腹鱼(♀) ×鲈鲤(♂)的F1代胚胎发育时间为144.33 h。发育温度相近时,杂交F1代的胚胎发育速度快于短须裂腹鱼与鲈鲤。推测是由于杂种优势导致其胚胎发育速度增快,或不同地区水质不同以及孵化条件的差异等导致胚胎发育时间的不同。
在黄河裸裂尻鱼(Schizopygopsis pylzovi)[30]、胡子鲇(Clarias fuscus)[31]的胚胎发育过程中均有强烈的卵黄运动,短须裂腹鱼(♀) ×鲈鲤(♂)的F1代在胚孔封闭期观察到轻微的卵黄运动。在短须裂腹鱼[19-21]和鲈鲤[22-24]的胚胎发育及裂腹鱼亚科其他鱼类,如尖裸鲤(Oxygymnocypris stewartii)[32]、小裂腹鱼(S. parvus)[33]、细鳞裂腹鱼(S. chongi)[34]等中均未见有卵黄运动的报道。刘文生等[31]认为卵黄运动有助于细胞在卵黄囊上排列以及对以后细胞分化的调整,推测杂交F1代胚胎发育速度快于亲本可能与卵黄运动有关,但卵黄运动具体形成原因及其对胚胎发育的影响机制有待进一步研究。
短须裂腹鱼(♀) ×鲈鲤(♂)的F1代胚胎发育过程与亲本基本一致,但也有其自身特点,尤其是器官发育阶段,鲈鲤和短须裂腹鱼均是在胚孔封闭后开始出现肌节、眼囊和耳囊,而其杂交F1代是出现肌节和眼囊、耳囊后胚孔才封闭,可能由于不同鱼类的胚胎发育具有其独特性[35],所以具体发育过程不尽相同。
3.2 短须裂腹鱼(♀) ×鲈鲤(♂)的F1仔稚鱼发育特征
短须裂腹鱼(♀) ×鲈鲤(♂)的F1代卵黄期仔鱼前期主要由卵黄囊提供营养物质,后期以丰年虫投喂,生长加快,其外形与双亲基本一致。仔鱼从孵化到卵黄囊吸收完毕历时17 d,其发育情况与生长指数均接近短须裂腹鱼[19-21]。目前未见短须裂腹鱼稚鱼发育的相关报道,杂交F1代仔稚鱼历时105 d发育完成,生长优势不明显,发育速度与生长速度均慢于鲈鲤[23]。推测杂交F1代后期仔鱼和稚鱼的生长速度更接近短须裂腹鱼,相较于亲本,短须裂腹鱼(♀) ×鲈鲤(♂)的F1代仔稚鱼生长发育的差异还有待进一步的实验验证。
短须裂腹鱼和鲈鲤属于不同亚科之间的远缘杂交,具有一定的遗传多样性。目前,对短须裂腹鱼和鲈鲤杂交育种的研究有限,杂交F1代在对环境因子的耐受性、抗病力等方面是否优于亲本,还有待实验验证。本实验的研究结果将为以后的进一步研究提供理论基础。
-
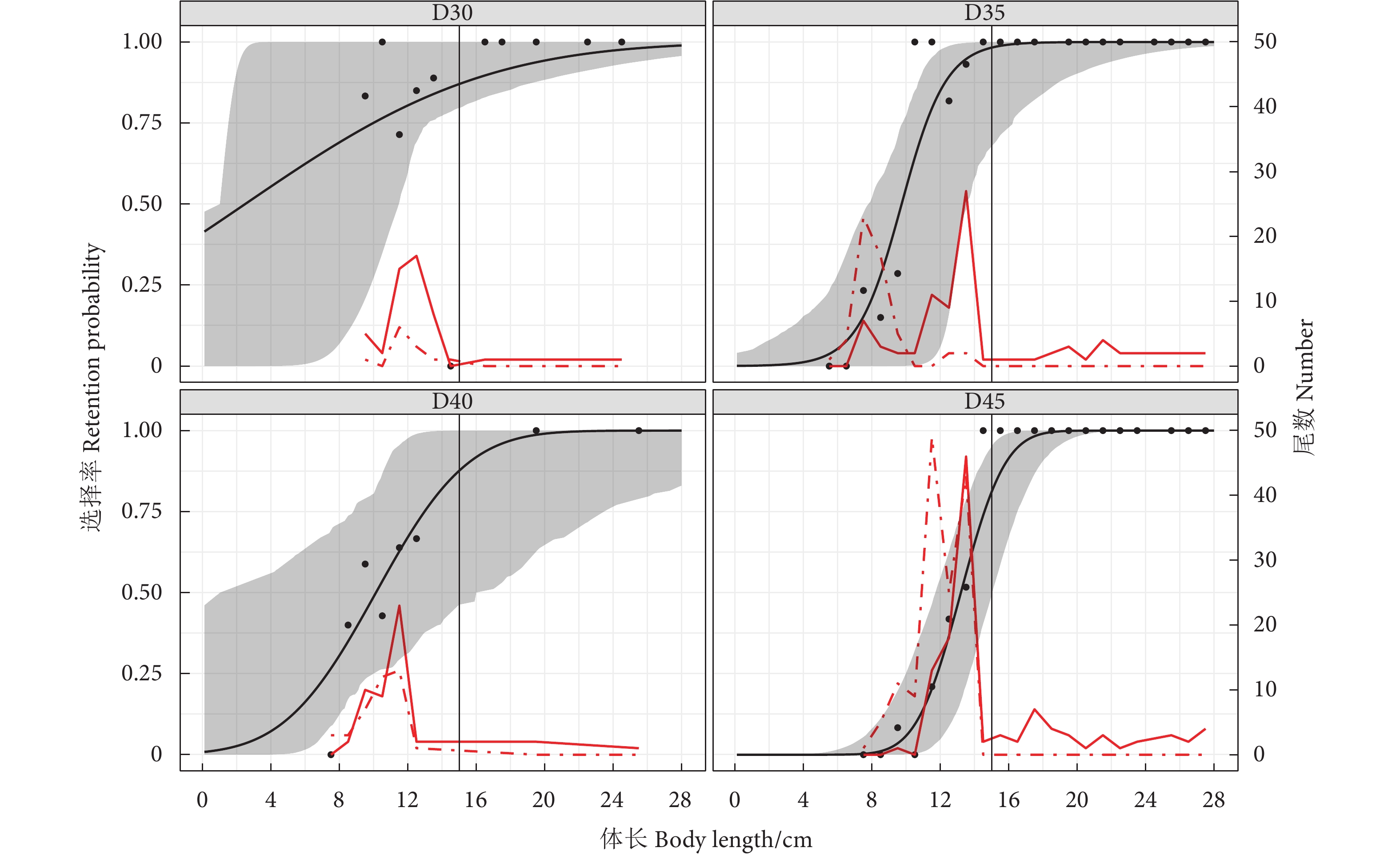
![]()
图 4 试验网囊对金线鱼的选择性曲线
Figure 4. Selectivity curves of experimental codends for N. virgatus
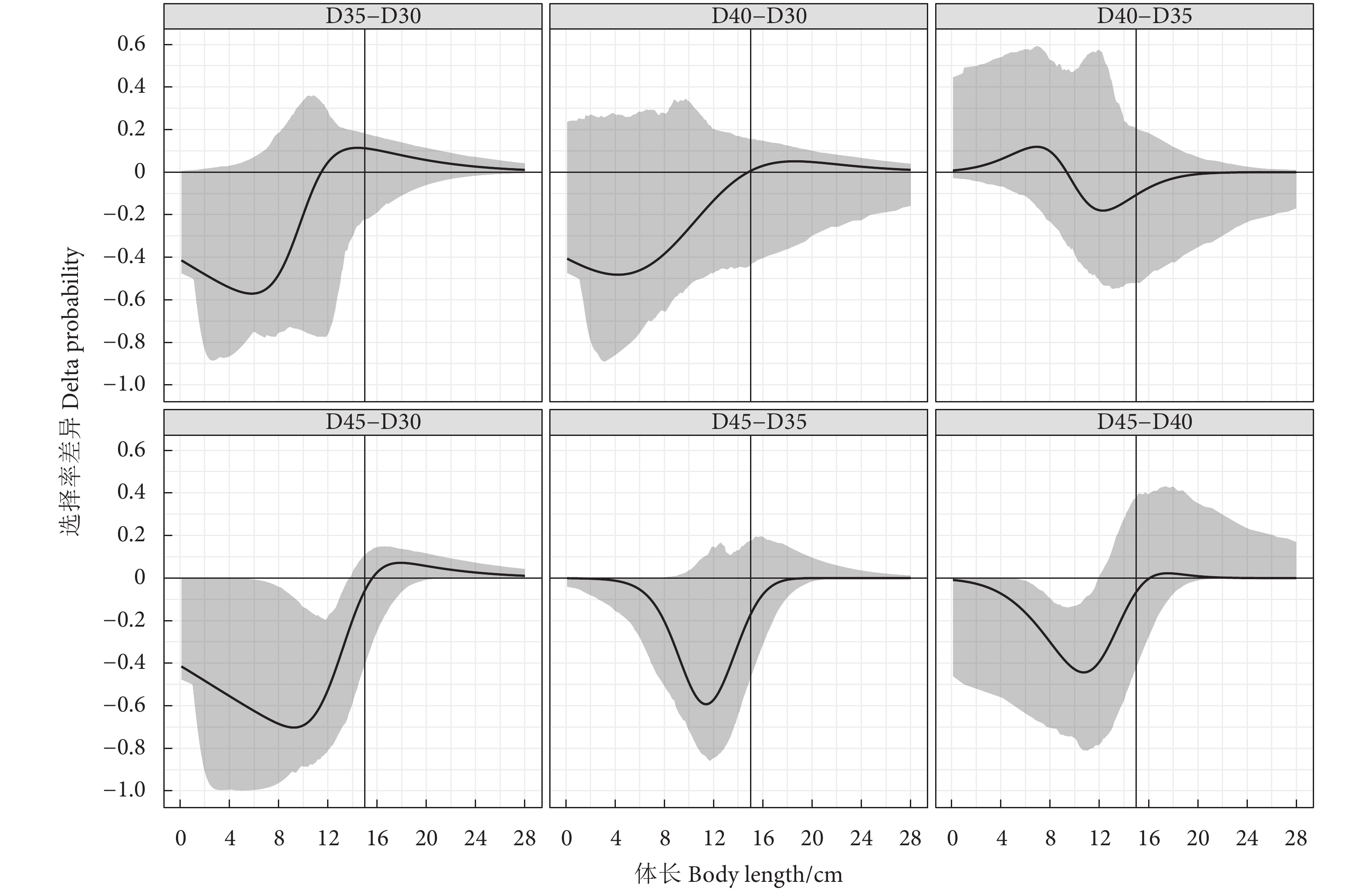
![]()
图 5 试验网囊对金线鱼的选择性差异曲线
Figure 5. Delta selectivity curves of experimental codends for N. virgatus
表 1 模型拟合的AIC值
Table 1 Akaike's information criterion (AIC) values of each model
网囊
Codend模型 Model Logit Probit Gompertz Richards D30 64.81 64.75 64.84 66.38 D35 99.26 99.70 100.55 101.23 D40 117.91 117.82 117.83 119.83 D45 262.93 262.49 265.52 263.35 注:最佳模型AIC值加粗。 Note: AIC values of the optimal models are in bold.  下载: 导出CSV
下载: 导出CSV
表 2 试验网囊的选择性参数及拟合度
Table 2 Selective parameters and fit statistics obtained from tested codends
网囊
Codend参数 Parameter 模型
Model50%选择体长
L50/cm选择范围
SR/cmP值
P-value残差
Deviance自由度
Degree of freedomD30 Probit 2.48 (1.00~11.47) 14.97 (1.00~20.84) 0.634 5 7.02 9 D35 Logit 9.73 (7.79~13.54) 2.87 (1.00~8.18) 0.868 3 12.39 19 D40 Probit 10.13 (1.00~16.00) 5.68 (2.04~27.44) 0.614 6 4.46 6 D45 Probit 13.20 (11.70~15.03) 2.75 (1.61~3.86) 0.995 4 6.18 18
下载: 导出CSV
表 3 试验网囊对金线鱼的捕捞方式指数
Table 3 Exploitation pattern indicators of experimental codends for N. virgatus
网囊
CodendInP−/% InP+/% Rdn/% D30 78.70
(53.10~100.00)94.56
(88.83~100.00)86.26
(77.38~90.54)D35 72.73
(18.96~87.57)99.83
(90.51~100.00)84.61
(58.82~89.02)D40 61.53
(31.29~89.03)97.99
(68.31~100.00)82.57
(74.58~89.04)D45 27.68
(7.88~50.13)98.57
(93.16~99.87)67.94
(38.70~80.43)
下载: 导出CSV
-
[1] 陈国宝, 梁沛文. 南海海洋鱼类原色图谱[M]. 北京: 科学出版社, 2016: 227. [2] 农业部渔业局. 2023中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2023: 39. [3] 农业部渔业局. 2012中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2012: 48. [4] 袁蔚文. 北部湾底层渔业资源的数量变动和种类更替[J]. 中国水产科学, 1995, 2(2): 57-65. [5] 王雪辉, 邱永松, 杜飞雁. 南海北部金线鱼生长、死亡和最适开捕体长研究[J]. 中国海洋大学学报, 2004, 34(2): 224-230. [6] 杨吝. 南海区海洋渔具渔法[M]. 广州: 广东科技术出版社, 2002: 85-86. [7] 杨吝, 张旭丰, 孙典荣, 等. 广东省拖网网囊调查[J]. 湛江海洋大学学报, 2001, 21(4): 22-26. [8] 张鹏, 杨吝, 张旭丰, 等. 刺网网目尺寸对南海区金线鱼选择性研究[J]. 南方水产, 2009, 1(2): 61-66. [9] 张鹏, 杨吝, 张旭丰, 等. 南海区金线鱼刺网网目选择性[J]. 中国水产科学, 2010, 17(5): 1085-1093. [10] 杨炳忠, 张鹏, 谭永光, 等. 基于SELECT模型的南海金线鱼刺网选择性研究[J]. 南方水产科学, 2016, 12(5): 8-15. [11] 杨吝, 谭永光, 张旭丰. 南海底拖网方、菱目网囊选择性研究[J]. 湛江海洋大学学报, 2002, 22(3): 19-25. [12] 中华人民共和国农业农村部. 关于实施海洋捕捞准用渔具最小网目尺寸制度的通告[EB/OL]. (2013-12-20)[2023-08-15]. http://www.moa.gov.cn/nybgb/2013/dseq/201805/t20180510_6141829.htm. [13] WILEMAN D A, FERRO R S T, FONTEYNE R, et al. Manual of methods of measuring the selectivity of towed fishing gear[R]. Copenhagen, Denmark: International Council for the Exploration of the Sea ICES Coop Res Rep, 1996(215): 1-51.
[14] FRYER R J. A model of between-haul variation in selectivity[J]. ICES J Mar Sci, 1991, 48(3): 281-290. doi: 10.1093/icesjms/48.3.281
[15] MILLAR R B. Incorporation of between-haul variation using bootstrapping and nonparametric estimation of selection curves[J]. Fish Bull, 1993, 91(3): 564-572.
[16] HERRMANN B, SISTIAGA M, NIELSEN K N, et al. Understanding the size selectivity of redfish (Sebastes spp.) in North Atlantic trawl codends[J]. J Northw Atl Fish Sci, 2012, 44: 1-13. doi: 10.2960/J.v44.m680
[17] AKAIKE H. A new look at the statistical model identification[J]. IEEE Trans Automat Contr, 1974, 19(6): 716-722. doi: 10.1109/TAC.1974.1100705
[18] EFRON B. Bootstrap methods: another look at the jackknife[J]. Ann Statisti, 1979, 7: 1-26.
[19] HERRMANN B, KRAG L A, KRAFFT B A. Size selection of Antarctic krill (Euphausia superba) in a commercial codend and trawl body[J]. Fish Res, 2018, 207: 49-54. doi: 10.1016/j.fishres.2018.05.028
[20] YANG B Z, HERRMANN B, YAN L, et al. Size selectivity and catch efficiency of diamond-mesh codends in demersal trawl fishery for conger pike (Muraenesox cinereus) of the South China Sea[J]. Ocean Coast Manage, 2021, 211: 105777. doi: 10.1016/j.ocecoaman.2021.105777
[21] 杨炳忠, 万荣. 基于双重拔靴法拟合拖网网囊选择性的不确定性[J]. 中国水产科学, 2023, 30(9): 1166-1175. [22] 陈丕茂. 南海北部主要捕捞种类最适开捕规格研究[J]. 水产学报, 2004, 28(4): 393-400. [23] YANG B Z, HERRMANN B. Simple and effective: T90 codends improve size selectivity and catch efficiency compared with diamond-mesh codends for mantis shrimp (Oratosquilla oratoria) in demersal trawl fishery of the South China Sea[J]. Front Mar Sci, 2022, 9: 939269. doi: 10.3389/fmars.2022.939269
[24] HERRMANN B, SISTIAGA M, LARSEN R, et al. Effect of three different codend designs on the size selectivity of juvenile cod in the Barents Sea shrimp trawl fishery[J]. Fish Res, 2019, 219: 1-8.
[25] YANG B Z, HERRMANN B, WAN R. Effects of codend mesh-shape and size on the size selectivity of demersal trawl targeting banded scad (Alepes djedaba) in the Beibu Gulf, South China Sea[J]. J Mar Sci Eng, 2023, 11(7): 1390. doi: 10.3390/jmse11071390
[26] GRAHAM N, KYNOCH R J, FRYER R J. Square mesh panel in demersal trawls: further data relating haddock and whiting selectivity to panel position[J]. Fish Res, 2003, 62: 361-375. doi: 10.1016/S0165-7836(02)00279-5
[27] GRAHAM N, JONES E G, REID D G. Review of technological advances for the study of fish behavior in relation to demersal fishing trawls[J]. ICES J Mar Sci, 2004, 61: 1036-1043. doi: 10.1016/j.icesjms.2004.06.006
[28] BRCIC J, HERRMANN B, SALA A. Can a square-mesh panel inserted in front of the cod end improve size and species selectivity in Mediterranean trawl fisheries?[J]. Can J Fish Aquat Sci, 2018, 75: 704-713. doi: 10.1139/cjfas-2017-0123
-
期刊类型引用(3)
1. 左鹏翔,金方彭,王志飞,吴俊颉,赵静霞,赵利刚,王文玉,毕晓敏,沈智敏,李波,冷云. 细鳞裂腹鱼(♀)与鲈鲤(♂)杂交及生长对比试验初报. 中国农学通报. 2024(11): 159-164 .  百度学术
百度学术
2. 陈军平,沈方方,武慧慧,张佳鑫,于若梦,付永杰,谢国强,赵道全. 我国鱼类胚胎发育研究进展. 江苏农业科学. 2021(17): 45-52 . 百度学术
3. 梁孟,魏开金,朱祥云,马宝珊,徐滨,徐进. 饥饿和再投喂对短须裂腹鱼幼鱼体组分、消化酶活性及RNA/DNA的影响. 中国农学通报. 2020(23): 151-160 . 百度学术
其他类型引用(1)







计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
- 被引次数: 4
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
