
Study on relationship between Thunnus albacares catches and climatic factors based on CNN-BiLSTM model
-
摘要:
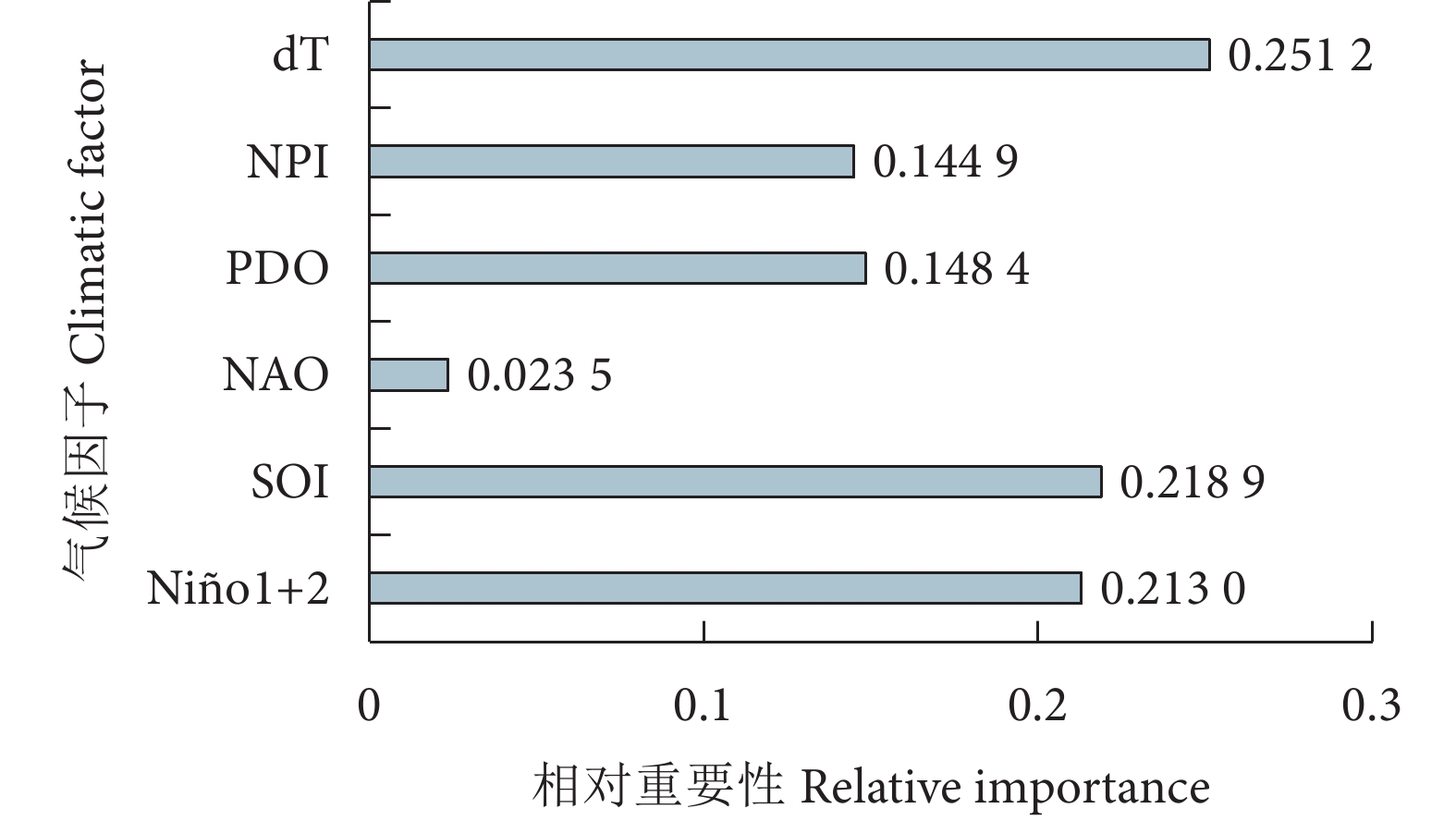
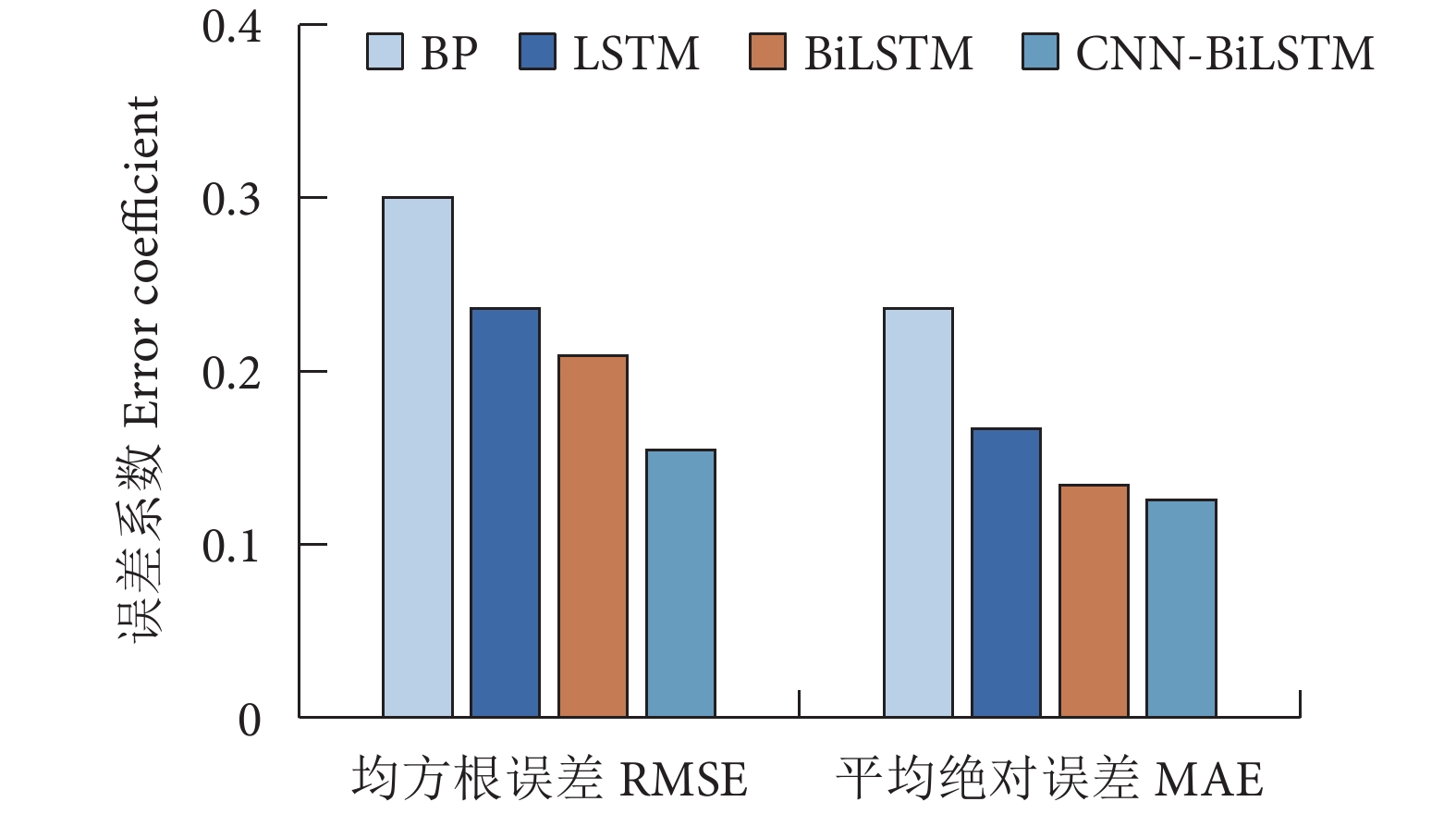
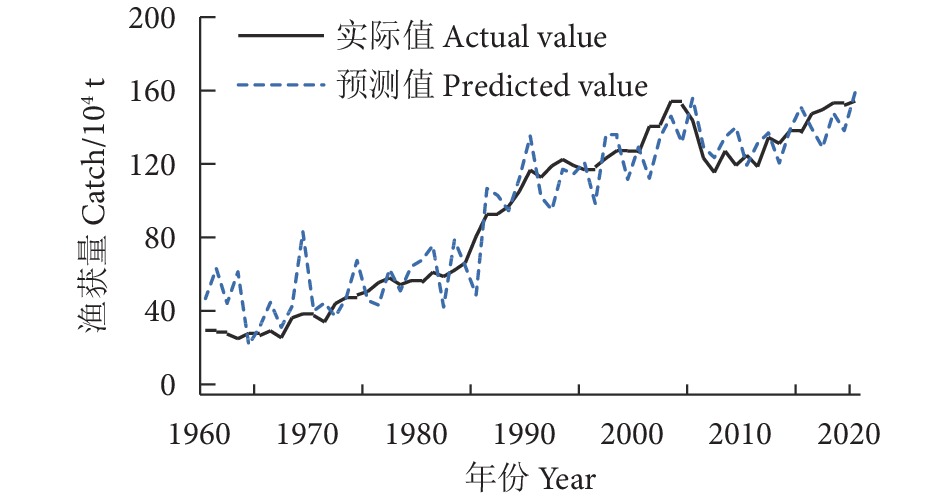
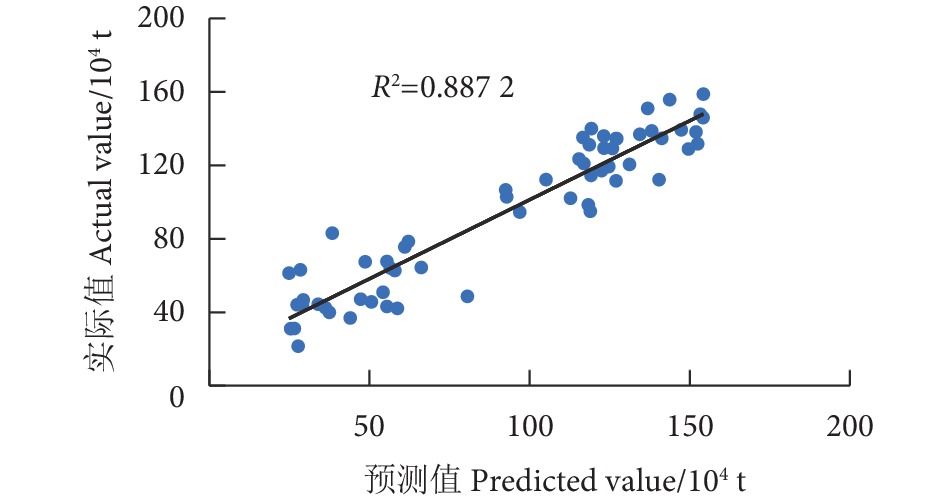
为探究气候因子对黄鳍金枪鱼渔获量的影响,根据1960—2021年的南方涛动指数 (SOI)、太平洋年代际涛动 (PDO)、北大西洋涛动 (NAO)、北太平洋指数 (NPI)、全球海气温度异常指标 (dT) 以及厄尔尼诺相关指标 (Niño1+2、Niño3、Niño4以及Niño3.4) 等9种气候因子数据和全球黄鳍金枪鱼渔获量数据,采用相关性分析、BP神经网络、长短期记忆网络 (LSTM) 模型、双向长短期记忆网络 (BiLSTM) 模型和卷积神经网络结合双向长短期记忆网络 (CNN-BiLSTM) 模型对低频气候因子与黄鳍金枪鱼渔获量的关系进行了研究。结果表明,气候变化表征因子对黄鳍金枪鱼渔获量的重要性依次为dT>SOI>Niño1+2>PDO>NPI>NAO,其对应的最佳滞后年限分别为0、11、6、5、15、0年。CNN-BiLSTM模型的预测效果最优,其后依次为BiLSTM模型、LSTM模型、BP神经网络模型。最优预测模型显示预测值与实际值的拟合优度为0.887,平均绝对误差为0.125,均方根误差为0.154,预测值与实际值变化趋势基本一致,模型拟合效果良好。
-
关键词:
- 黄鳍金枪鱼 /
- 渔获量 /
- 气候因子 /
- CNN-BiLSTM模型 /
- 相关性分析
Abstract:To explore the impact of climatic factors on Thunnus albacares catches, we studied its relationship with low-frequency climatic factors by using correlation analysis, BP neural network, LSTM model, BiLSTM model and CNN-BiLSTM model based on the data of nine climate factors, including Southern Oscillation Index (SOI), Pacific Decadal Oscillation (PDO), North Atlantic Oscillation (NAO), North Pacific Index (NPI), global sea-air temperature anomaly index (dT), El Niño-related indexes (Niño1+2, Niño3, Niño4, Niño3.4) from 1960 to 2021, as well as global T. albacares catches data. The results show that the importance of climate change characterization factors on T. albacares catches followed a descending order of dT>SOI>Niño1+2>PDO>NPI>NAO, whose corresponding optimal lag periods were 0, 11, 6, 5, 15 and 0 years, respectively. CNN-BiLSTM model had the highest prediction accuracy, followed by BiLSTM, LSTM and BP. The goodness of fit between the predicted and actual values of CNN-BiLSTM model was 0.887, with a mean absolute error of 0.125 and a root mean square error of 0.154. The trend of predicted values and actual values was basically consistent, indicating a good model fitting effect.
-
Keywords:
- Thunnus albacares /
- Catch /
- Climatic factors /
- CNN-BiLSTM model /
- Correlation analysis
-
-
![]()
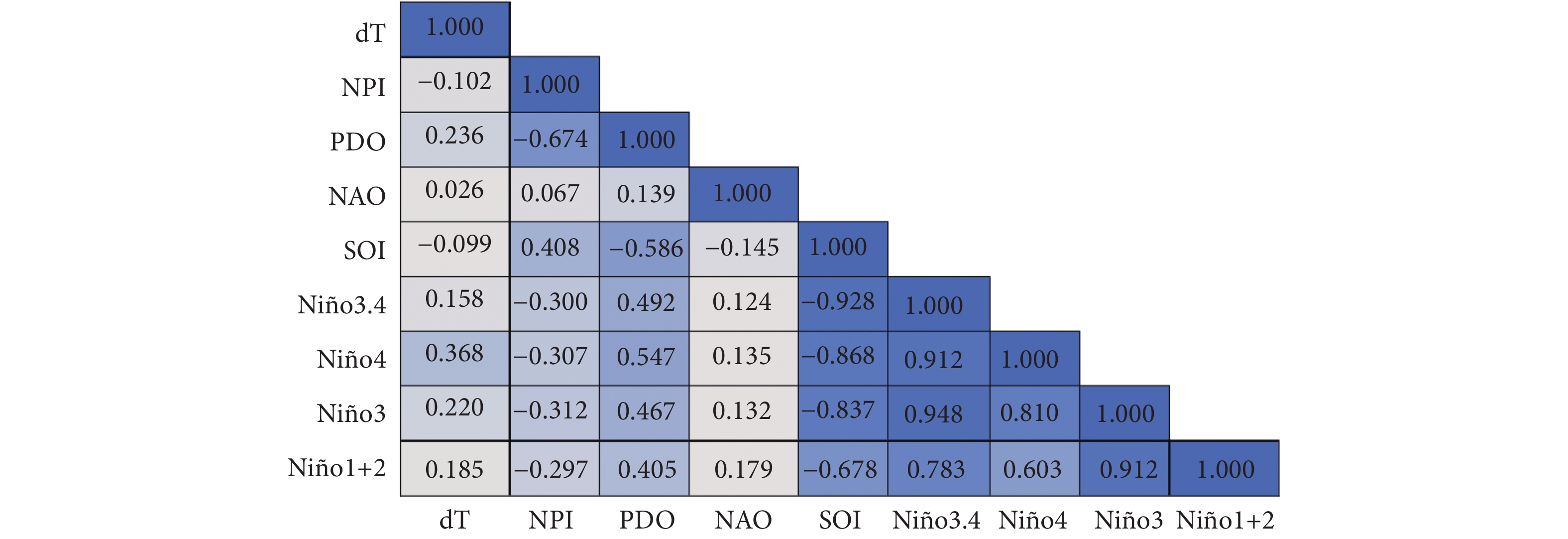
图 1 Spearman秩相关系数表
注:dT. 全球海气温度异常指标;NPI. 北太平洋指数;PDO. 太平洋年代际涛动;NAO. 北大西洋涛动;SOI. 南方涛动指数;Niño1+2, Niño3, Niño4, Niño3.4. 厄尔尼诺相关指标。表1、图2同此。
Figure 1. Spearman rank correlation coefficient of characterization factors
Note: dT. Global sea-air temperature anomaly index; NPI. North Pacific Index; PDO. Pacific Decadal Oscillation; NAO. North Atlantic Oscillation; SOI. Southern Oscillation Index; Niño1+2, Niño3, Niño4, Niño3.4. El Niño-related indexes. The same case in Table 1 and Fig. 2.
![]()
图 2 气候变化表征因子相对重要性
Figure 2. Relative importance of climate change characterization factors
![]()
图 4 黄鳍金枪鱼渔获量实际值与预测值对比
Figure 4. Comparison of actual and predicted catches of T. albacares
![]()
图 5 黄鳍金枪鱼渔获量实际值与预测值线性关系
Figure 5. Linear relationship between actual and predicted catches of T. albacares
表 1 气候表征因子与黄鳍金枪鱼互相关关系
Table 1 Cross-correlation between climate change characterization factors and T. albacares
滞后年限
Lagging yearNiño1+2 SOI NAO PDO NPI dT 0 0.051 −0.078 −0.172 0.25 −0.052 0.892 1 0.065 −0.087 0.03 0.291 −0.101 0.845 2 0.08 −0.085 0.016 0.312 −0.12 0.794 3 0.117 −0.081 0.032 0.301 −0.129 0.743 4 0.135 −0.079 0.015 0.298 −0.145 0.69 5 0.147 −0.087 0.005 0.314 −0.179 0.638 6 0.159 −0.104 −0.006 0.299 −0.177 0.582 7 0.115 −0.101 −0.024 0.266 −0.189 0.52 8 0.063 −0.091 −0.028 0.226 −0.142 0.457 9 0.054 −0.109 −0.033 0.23 −0.17 0.41 10 0.065 −0.112 −0.011 0.248 −0.191 0.371 11 0.07 −0.164 −0.008 0.274 −0.189 0.335 12 0.062 −0.161 0.037 0.261 −0.186 0.281 13 0.035 −0.128 0.038 0.246 −0.19 0.244 14 0.042 −0.143 0.047 0.263 −0.193 0.209 15 0.068 −0.126 0.048 0.278 −0.203 0.164 注:加粗字体表示气候变化表征因子的最佳互相关系数。 Note: Bold fonts represent the optimal cross-correlations coefficients of climate change characterization factors.  下载: 导出CSV
下载: 导出CSV
-
[1] 石肖飞, 王啸, 王佚兮, 等. 热带中西太平洋海域黄鳍金枪鱼的摄食生物学特性[J]. 南方水产科学, 2022, 18(1): 43-51. [2] 郭爱, 陈新军. ENSO与中西太平洋金枪鱼围网资源丰度及其渔场变动的关系[J]. 海洋渔业, 2005(4): 338-342. [3] 林泓羽, 汪金涛, 陈新军, 等. 中西太平洋暖池时空变动及其对鲣分布的影响[J]. 南方水产科学, 2023, 19(3): 173-180. doi: 10.12131/20220235 [4] 周为峰, 陈亮亮, 崔雪森, 等. 异常气候下温跃层及时空因子对中西太平洋黄鳍金枪鱼渔场分布的影响[J]. 中国农业科技导报, 2021, 23(10): 192-201. [5] 王啸, 刘文俊, 张健. 基于ARIMA的海洋尼诺指数对中西太平洋黄鳍金枪鱼年际CPUE的影响[J]. 南方水产科学, 2023, 19(4): 10-20. doi: 10.12131/20230007 [6] 沈建华, 陈雪冬, 崔雪森. 中西太平洋金枪鱼围网鲣鱼渔获量时空分布分析[J]. 海洋渔业, 2006(1): 13-19. [7] LU H J, LEE K T, LIN H L, et al. Spatio-temporal distribution of yellowfin tuna Thunnus albacares and bigeye tuna Thunnus obesus in the Tropical Pacific Ocean in relation to large-scale temperature fluctuation during ENSO episodes[J]. Fish Sci, 2001, 67(6): 1046-1052. doi: 10.1046/j.1444-2906.2001.00360.x
[8] 李鹏, 许柳雄, 周成, 等. 中西太平洋金枪鱼围网自由鱼群渔场重心变动及其与南方涛动指数的关系[J]. 南方水产科学, 2020, 16(2): 70-76. [9] KLYASHTORIN L B. Long-term climate change and main commercial fish production in the Atlantic and Pacific[J]. Fish Res, 1998, 37(1/2/3): 115-125.
[10] SOYKAN C U, EGUCHI T, KOHIN S, et al. Prediction of fishing effort distributions using boosted regression trees[J]. Ecol Appl, 2014, 24(1): 71-83. doi: 10.1890/12-0826.1
[11] CHEUNG W W L, LAN V, SARMIENTO J, et al. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change[J]. Glob Change Biol, 2010(16): 24-35.
[12] 王少琴, 许柳雄, 朱国平, 等. 中西太平洋金枪鱼围网的黄鳍金枪鱼CPUE时空分布及其与环境因子的关系[J]. 大连海洋大学学报, 2014, 29(3): 303-308. [13] 唐峰华, 崔雪森, 杨胜龙, 等. 海洋环境对中西太平洋金枪鱼围网渔场影响的GIS时空分析[J]. 南方水产科学, 2014, 10(2): 18-26. [14] 肖启华, 黄硕琳. 气候变化对东南太平洋智利竹䇲鱼渔获量的影响[J]. 中国水产科学, 2021, 28(8): 1020-1029. [15] 周永江, 姚宜斌, 熊永良, 等. 基于Spearman秩相关系数的PWV与PM2.5相关性研究[J]. 大地测量与地球动力学, 2020, 40(3): 236-241. [16] 肖启华. 气候变化背景下东南太平洋智利竹䇲鱼资源评估模型研究[D]. 上海: 上海海洋大学, 2021: 16-42. [17] 肖启华, 黄硕琳. 气候变化对海洋渔业资源的影响[J]. 水产学报, 2016, 40(7): 1089-1098. [18] 徐策, 张力, 余静, 等. 气候变化对中国近海捕捞渔业的影响: 以太平洋年代际涛动为例[J]. 资源科学, 2022, 44(2): 386-400. [19] 宋利明, 任士雨, 张敏, 等. 基于集成学习的大西洋热带水域大眼金枪鱼渔情预报[J]. 水产学报, 2023, 47(4): 64-76. [20] 刘佳翰, 陈克绪, 马建, 等. 基于卷积神经网络和随机森林的三相电压暂降分类[J]. 电力系统保护与控制, 2019, 47(20): 112-118. [21] 许回. 不同空间分辨率对LSTM渔场预报精度的影响及在最佳空间分辨率下其预报结果与QRM结果的比较[D]. 上海: 上海海洋大学, 2023: 18-21. [22] 周凯, 焦龄霄, 胡志坚, 等. 基于注意力机制的CNN-BiLSTM操作票自动校核方法[J]. 武汉大学学报(工学版), 2023, 56(9): 1114-1123. [23] 陈纬楠, 胡志坚, 岳菁鹏, 等. 基于长短期记忆网络和LightGBM组合模型的短期负荷预测[J]. 电力系统自动化, 2021, 45(4): 91-97. [24] 范宇雪, 王江文, 梅桂明, 等. 基于BI-LSTM的小样本滚动轴承故障诊断方法研究[J]. 噪声与振动控制, 2020, 40(4): 103-108. [25] 郭力, 郑良瑞, 冯浪. 基于相关性分析与CNN-BiLSTM神经网络的PSZ陶瓷磨削表面粗糙度智能预测[J]. 南京航空航天大学学报, 2023, 55(3): 401-409. [26] 袁红春, 张永, 张天蛟. 基于EMD-BiLSTM的太平洋大眼金枪鱼渔场预报模型研究[J]. 渔业现代化, 2021, 48(1): 87-96. [27] LIN D M, CHEN X J, CHEN Y, et al. Ovarian development in Argentinean shortfin squid Illexargentinus: group-synchrony for corroboration of intermittent spawning strategy[J]. Hydrobiologia, 2017, 795(1): 327-339. doi: 10.1007/s10750-017-3154-y
[28] RODHOUSE P G. Managing and forecasting squid fisheries in variable environments[J]. Fish Res, 2001, 54(1): 3-8. doi: 10.1016/S0165-7836(01)00370-8
[29] 余为, 陈新军, 易倩. 不同气候模态下西北太平洋柔鱼渔场环境特征分析[J]. 水产学报, 2017, 41(4): 525-534. [30] 陈丙见, 冯志萍, 余为. 厄尔尼诺和拉尼娜发生期太平洋褶柔鱼秋生群资源丰度的响应研究[J]. 中国水产科学, 2022, 29(11): 1636-1646. [31] 周甦芳. 厄尔尼诺-南方涛动现象对中西太平洋鲣鱼围网渔场的影响[J]. 中国水产科学, 2005, 12(6): 73-78. [32] 官文江, 朱江峰, 高峰. 印度洋长鳍金枪鱼资源评估的影响因素分析[J]. 中国水产科学, 2018, 25(5): 1102-1114. [33] 李娜, 陆化杰, 陈新军. 基于不同BP神经网络的西南大西洋阿根廷滑柔鱼渔场预报模型比较[J]. 广东海洋大学学报, 2017, 37(1): 65-71. [34] 丁鹏, 邹晓荣, 白思琦, 等. 东南太平洋智利竹䇲鱼渔场时空分析与资源丰度的预测[J]. 大连海洋大学学报, 2021, 36(4): 629-636. [35] 丁鹏, 邹晓荣, 冯超, 等. 东南太平洋智利竹䇲鱼的洄游路线[J]. 大连海洋大学学报, 2021, 36(6): 1027-1034. [36] 陈雪忠, 樊伟, 崔雪森, 等. 基于随机森林的印度洋长鳍金枪鱼渔场预报[J]. 海洋学报(中文版), 2013, 35(1): 158-164. [37] 房崇鑫, 盛震宇, 夏明, 等. 基于CNN-BiLSTM混合神经网络的雷达信号调制方式识别[J/OL]. 无线电工程: 1-10. [2023-09-28]. [38] 国显达, 那日萨, 崔少泽. 基于CNN-BiLSTM的消费者网络评论情感分析[J]. 系统工程理论与实践, 2020, 40(3): 653-663. [39] HAMPTON J, LEWIS A, WILLIAMS P. The western and central Pacific tuna fishery: overview and status of stocks [M]// New Caledonia: Secretariat of the Pacific Community, 1999: 8-12.
[40] HAMPTON J. Estimates of tag-reporting and tag-shedding rates in a large-scale tuna tagging experiment in the western tropical Pacific Ocean[J]. Oceanogr Lit Rev, 1997, 95(1): 68-79.
[41] BAKUN A, PARRISH R H. Comparative studies of coastal pelagic fish reproductive habitats: the Brazilian sardine (Sardinella aurita)[J]. ICES J Mar Sci, 1990, 46(3): 269-283. doi: 10.1093/icesjms/46.3.269
[42] DEARY A L, MORET-FERGUSON S, ENGELS M, et al. Influence of Central Pacific oceanographic conditions on the potential vertical habitat of four tropical tuna species[J]. Pac Sci, 2015, 69(4): 461-476. doi: 10.2984/69.4.3
[43] ERNESTO T O, ARTURO M M, ARMANDO T, et al. Variation in yellowfin tuna (Thunnus albacares) catches related to El Niño-Southern Oscillation events at the entrance to the Gulf of California[J]. Fish Bull, 2006, 104(2): 197-203.
[44] WILLIAMS P, RUAIA T. Overview of tuna fisheries in the western and central Pacific Ocean, including economic conditions–2019[C]//16th WCPFC Scientific Committee Meeting (SC16). Pohnpei State, Federated States of Micronesia: WCPFC, 2020: 7.
[45] SIRABELLA P, GIULIANI A, COLOSIMO A, DIPPNER J W. Breaking down the climate effects on cod recruitment by principal component analysis and canonical correlation[J]. Mar Ecol Prog Ser, 2001(216): 213-222.
[46] OTTERSEN G, STENSETH N C. Atlantic climate governs oceanographic and ecological variability in the Barents Sea[J]. Limnol Oceanogr, 2001, 46(7): 1774-1780. doi: 10.4319/lo.2001.46.7.1774
[47] WU L X, LIU Z Y, LIU Y, et al. Potential global climatic impacts of the North Pacific Ocean[J]. Geophys Res Lett, 2005, 32(24): L24710.


计量
- 文章访问数: 199
- HTML全文浏览量: 42
- PDF下载量: 53
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
