
Isolation, identification and antimicrobial susceptibility test of Edwardsiella tarda in Bahaba taipingensis
-
摘要:
2023 年入夏以来,东莞市黄唇鱼 (Bahaba taipingensis) 自然保护区救护基地出现黄唇鱼发病和死亡病例,症状主要为食欲下降,体表发红、充血,眼球突出,角膜白浊溃烂,吻部溃烂;解剖后发现腹腔有大量腹水,肝肿大呈褐色。为确定其病因,针对性制定防治措施,采集死亡病鱼的心脏、肝脏、脾脏、肾脏、肠、鳃丝等组织样品。从病死鱼体内分离到一株优势菌株,命名为DG230920,通过对菌株的形态特征、生理生化特性和16S rRNA基因序列综合分析,鉴定该菌为迟缓爱德华氏菌 (Edwardsiella tarda)。病理组织学检测结果显示,心脏、肝脏、脾脏、肾脏和鳃均有明显的炎症反应,肠出血、黏膜上皮细胞脱落,脾淋巴细胞稀疏、内皮细胞变性坏死,肾小管变性坏死,鳃见鳃丝脱落、出血。对分离菌株进行药敏分析,结果为对环丙沙星、左氧氟沙星高度敏感,最低抑菌质量浓度仅≤0.25 mg·mL−1;其次是厄他培南,最低抑菌质量浓度≤0.5 mg·mL−1。结果表明,黄唇鱼发病原因可能是迟缓爱德华氏菌感染,分离菌株遗传序列稳定,对临床使用的抗菌药物均为敏感。研究提示,应加强黄唇鱼迟缓爱德华氏菌感染的防治,选用敏感药物科学治疗。
Abstract:Since the summer of 2023, there have been cases of outbreak and death in the Bahaba taipingensis Natural Conservation Area in Dongguan City. The symptoms mainly include loss of appetite, reddening and congestion of body surface, protruding eyeballs, opaque and ulcerated corneas, as well as ulceration in snout. Dissection revealed a large amount of ascites in abdominal cavity and enlarged liver with brown color. To determine the cause of the disease, and make prevention and control measures, we collected tissue samples such as heart, liver, spleen, kidney, intestine and gills from the dead fish. A dominant strain was isolated from and named as DG230920. Through a comprehensive analysis of its morphological characteristics, physiological and biochemical properties, in addition to the 16S rRNA gene sequence analysis, the bacteria were identified as Edwardsiella tarda. Pathological histological examination results show obvious inflammatory reactions in heart, liver, spleen, kidney and gills. Hemorrhage and desquamation of epithelial cells were observed in intestine; sparse lymphocytes and necrosis of endothelial cells were found in the spleen; tubular degeneration and necrosis were observed in the kidney. Gills showed shedding and hemorrhage of gill filaments. Drug sensitivity analysis of the isolated strain revealed high sensitivity to ciprofloxacin and levofloxacin, with a minimum inhibitory concentration of only ≤0.25 mg·mL−1. The next most effective drug was imipenem, with a minimum inhibitory concentration of ≤0.5 mg·mL−1. The results indicate that the cause of B. taipingensis disease might be an infection with E. tarda, and the isolated strain exhibited stable genetic sequences and sensitivity to clinically used antibiotics. The study suggests that prevention and control measures should be strengthened against E. tarda infection in B. taipingensis, and sensitive drugs should be used for treatment.
-
大伙房水库地处辽河支流浑河中上游,辽宁省抚顺市境内,位于41°47′N~41°57′N,124°05′E~124°22′E,为带状河谷型水库。东西长约35 km,最大水深37 m,最大蓄水面积114 km2,总库容21.87×109 m3。水库渔获鱼类11种,主要经济鱼类5种,为鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、鲤(Cyprinus carpio)、鲫(Carassius auratus)、草鱼(Ctenopharyngodon idellus)。鲢、鳙为该水库主要增殖品种,约占总量的85%。每年10月投放鱼苗40 t左右,其中鳙58%、鲢27%,规格为35~100 g·尾-1。作为中国9大重点饮用水源保护地,合理的鱼类放养捕捞既可以优化水质、维持水库生态稳定,又能创造较高的经济价值。对库区鱼类资源不同季节现存量以及分布状况的把握,可为制定增殖放养和捕捞计划提供科学依据。
由于淡水水域多采用人工增殖放养和定置网箔大规格体长鱼类选择性捕捞,传统的渔业资源调查评估方法较难实施。近年来随着渔业声学测量技术在中国推广,渔业资源声学评估方法在淡水的应用逐渐增多且收到较好效果。包括武汉东湖、长江葛洲坝、新疆塞里木湖、青海湖、辽宁清河水库、三峡水库和楠溪江等不同种类鱼类的声学资源调查和空间分布[1-7]。中国虽然在2007年编制了针对海洋调查的《渔业声学资源调查与评估》规范[8],但是由于湖泊江河相对海洋水域范围狭窄且鱼类组成各异,目前已成熟的海洋声学评估技术尚不能简单复制于淡水调查,需要针对调查水域实际情况开展研究,获得最佳的调查方法。国外淡水渔业声学研究较少,鱼种与中国差异较大,可参考的结果也较少[9-13]。
因此该研究选择春、夏、秋(冬季冰层较厚不能进行声学调查)季节,在2012年7月19~21日、9月14~16日和2013年5月12~14日分别对水库进行鱼类资源声学调查,对鱼类的回波特征、单体目标大小和分布、资源现存量和分布及主要水文参数进行分析,考察鱼类资源分布随季节和水文参数变化的规律,为确定北方水库合理的声学调查方法提供科学依据。
1. 材料与方法
1.1 调查航线和站位的设定
大伙房水库为T型交叉带状河谷型水库,综合考虑水库形状、水深分布、采样点及网箱位置等因素,声学调查采用“之”字形航线对库区(上游有网箱的区域除外)进行断面声学回波采样。水文参数采样按上、中、下游设置共计8个采样站位,分别为位于上游支流交汇处的A1、A2,位于中游断面点B1、B2、B3,以及位于两主流交汇处C1、南侧C2及北侧大坝附近的C3,图 1为7月的调查航迹线和站位分布图,其余2次调查航线与7月航线基本相同。
![]() 图 1 大伙房水库7月声学调查航线及水文调查站位分布图A、B、C分别为库区上、中、下游Fig. 1 Acoustic survey trace and hydrographic investigation stations in Dahuofang Reservoir in July, 2012The A, B and C areas are upper, middle and lower reaches, respectively.
图 1 大伙房水库7月声学调查航线及水文调查站位分布图A、B、C分别为库区上、中、下游Fig. 1 Acoustic survey trace and hydrographic investigation stations in Dahuofang Reservoir in July, 2012The A, B and C areas are upper, middle and lower reaches, respectively.1.2 调查方法和数据后处理
使用多参数水质分析仪(美国YSI,6100型)在设置站位对水温、pH、溶解氧(DO)等参量进行垂直剖面测量,叶绿素a使用荧光法测量。声学调查使用小艇(长6 m,宽2.5 m)走航测量,平均航速6 kn。分裂波束科学鱼探仪(挪威Simrad,EY60型,70 kHz)的波束半功率指向全角为7°、脉冲宽度0.256 ms、发射功率300 W、脉冲收发间隔0.1 s。换能器放置于导流罩内,使用钢管悬挂固定于船弦右侧,吃水1 m。调查前用水质分析仪测量水温、盐度,计算声速和吸收系数,使用直径38.1 mm钨钢合金标准球对科学鱼探仪进行校正。使用GPS导航仪(美国Garmin,60CS型)测量位置信息,数据通过RS232接口与科学鱼探仪数据进行同步显示和保存。使用专用声学数据后处理软件Echoview 4.0(澳大利亚,Myriax)进行单体目标统计和资源评估。
1.3 鱼类资源声学评估方法
库区鱼类资源声学评估主要参考《渔业资源声学调查与评估》规范[8]中回波积分法进行。由于大伙房历年捕捞渔获物中鲢、鳙种类的比例占绝对优势(2011年鳙为52%,鲢为22%),且库区在6月~11月期间主要以定置网箔方式进行活鱼捕捞,无法提供对应声学回波的网具采样,因此现阶段暂按鳙的声学参数对整体资源量进行评估。
鳙的姿态平均目标强度(target strength,TS)与体长的关系式,使用基尔霍夫近似射线(KRM)模型计算的姿态平均TS结果[14-15]:
$$ T S_{\mathrm{a}}=21.1 \log L_{\mathrm{cm}}-67.1 $$ (1) 其中Lcm为鱼标准体长(cm)。分裂波束科学渔探仪调查时,TSa可以直接对单体回波进行检测后现场测量。
资源质量可以利用体长与体质量的拟合关系式进行换算,该研究使用鲢鳙的关系式为[14]:
$$ W=0.0167 L^{2.9806} $$ (2) 其中W表示质量(g);L为标准体长(cm)。最终在面积As(m2)内鱼类资源总质量为:
$$ W_T=A_S \sum\limits_{i=1}^k<n_i>W_i $$ (3) 其中下标i表示不同体长鱼类, < n>为单位水面的鱼类平均密度。在回波积分处理单元内,不同体长鱼类的密度为下式[15-17]:
$$ n_i=\frac{S_a}{<\mathsf{σ}_{b s}>} \frac{p_i<\mathsf{σ}_{b s}>_i}{^i\sum\limits_{i=1}^k p_i<\mathsf{σ}_{b s}>_i} $$ (4) 其中sa为回波积分值(单位水面的散射强度),p为通过单体检测并进行目标跟踪后,不同TS鱼类数量的频度值, < σbs>为姿态平均后向散射截面(与TS的线性值相等)。
在数据后处理过程中,根据库区鱼类回波分散分布以及库区狭长的地理特征,选择积分处理单元水平距离为100 m。使用Echoview软件数据处理按以下步骤进行:1)进行气泡和干涉信号等剔除;2)进行单体目标检测、跟踪和频度计算,获得不同TSa(< σbs>)频度p值;3)在积分处理单元内进行回波积分,获得积分值sa;4)利用公式(4)计算不同TS鱼类的密度(尾·m-2);5)根据公式(2)和(3)计算上、中、下游的鱼类资源体质量;6)统计库区内不同体长鱼类水平和垂直空间密度分布。
2. 结果
2.1 主要水文参数
不同季节各站位水温剖面见图 2。2013年5月和2012年7月有较明显的温跃层,5月上游6~8 m,中下游8~10 m,7月15~20 m,温跃层下的等温层水温为4~6 ℃。2012年9月15 m以深水温逐渐降低,无明显温跃层变化。表层水温5月、7月和9月别分为10~15 ℃,18~27 ℃和20~22 ℃,7月水温的下降梯度最大。
![]() 图 2 大伙房水库不同季节调查站位的水温剖面a. 2013年5月;b. 2012年7月;c. 2012年9月Fig. 2 Water temperature profile in survey stations of Dahuofang Reservoir in different seasonsa. May, 2013;b. July, 2012;c. September, 2012
图 2 大伙房水库不同季节调查站位的水温剖面a. 2013年5月;b. 2012年7月;c. 2012年9月Fig. 2 Water temperature profile in survey stations of Dahuofang Reservoir in different seasonsa. May, 2013;b. July, 2012;c. September, 2012不同季节ρ(DO)剖面见图 3。ρ(DO)随深度增加而逐渐减小。7月表层ρ(DO)明显高于5月和9月。7月5 m以浅水层ρ(DO)为10~14 mg·L-1,5 m以深水层ρ(DO)随深度下降逐渐降低。5月ρ(DO)随深度变化不大,从表层至底层均维持在3~5 mg·L-1。9月15 m以浅水层ρ(DO)维持在7~10 mg·L-1,15 m之后ρ(DO)随深度下降逐渐减少。5月和9月下游ρ(DO)要低于中上游,7月上游ρ(DO)最低。其他水文参数均值:5月pH为7.71、叶绿素a质量浓度为12.09 μg·L-1、透明度为1.96 m。7月pH为8.27、叶绿素a质量浓度为27.7 μg·L-1、透明度为1.50 m。9月pH为7.62、叶绿素a质量浓度为12.06 μg·L-1,透明度为2.25 m。
![]() 图 3 大伙房水库不同季节调查站位的溶解氧剖面图a. 2013年5月;b.2012年7月;c.2012年9月Fig. 3 Dissolved oxygen profile in survey stations of Dahuofang Reservoir in different seasonsa. May, 2013;b. July, 2012;c. September, 2012
图 3 大伙房水库不同季节调查站位的溶解氧剖面图a. 2013年5月;b.2012年7月;c.2012年9月Fig. 3 Dissolved oxygen profile in survey stations of Dahuofang Reservoir in different seasonsa. May, 2013;b. July, 2012;c. September, 2012大伙房水库水质呈弱碱性,pH夏季高于春秋季。受春季融冰等因素影响,5月ρ(DO)较低,7月夏季受高温、降水和径流影响,表层较高,9月蓄水后表层ρ(DO)减少。浮游植物夏季受降水径流和高日照影响呈峰值状态。7月受降水和径流影响透明度较低。
2.2 声学调查结果
2.2.1 鱼类声学回波及单体目标强度特征
不同季节鱼类回波特征见图 4~图 6。2012年7月鱼类回波密度较大,回波信号分层分布,主要集中于在0~5 m及5~15 m水层,15 m以下有分散的单体回波信号(图 4),偶尔有鱼群回波(图 4中c回波)。9月回波较7月分散,10 m以下水层有带状回波,中游区域偶尔出现鱼群,底层也有鱼类回波(图 5)。2013年5月整个水库单体回波信号较少,鱼群回波较多,主要分布在5~10 m水层,集中在中游(图 6)。
![]() 图 4 大伙房水库7月中游鱼类回波特征图a~b.气泡;c.鱼群Fig. 4 Characteristics of fish echo in middle reaches of Dahuofang Reservoir in Julya~b. bubble; c. fish stock
图 4 大伙房水库7月中游鱼类回波特征图a~b.气泡;c.鱼群Fig. 4 Characteristics of fish echo in middle reaches of Dahuofang Reservoir in Julya~b. bubble; c. fish stock![]() 图 5 大伙房水库9月中游鱼类单体回波特征a.
图 5 大伙房水库9月中游鱼类单体回波特征a. Fig. 5 Characteristics of single fish echo in middle reaches in Dahuofang Reservoir in Septembera. H.leucisculus; b. H.molitrix and A.nobilis; c. fish stock; d. C.carpio or C.idellus
Fig. 5 Characteristics of single fish echo in middle reaches in Dahuofang Reservoir in Septembera. H.leucisculus; b. H.molitrix and A.nobilis; c. fish stock; d. C.carpio or C.idellus![]() 图 6 大伙房水库5月中游鱼类回波特征(a和b为鱼群)Fig. 6 Characteristics of fish echo in Dahuofang Reservoir in May (a and b indicate small fish stocks)
图 6 大伙房水库5月中游鱼类回波特征(a和b为鱼群)Fig. 6 Characteristics of fish echo in Dahuofang Reservoir in May (a and b indicate small fish stocks)目标识别主要通过单体目标检测、不同鱼类的捕捞方式和渔获物水层分布的经验来判断。0~5 m层的TS较小(-70~-58 dB)、结合
不同季节单体目标检测结果见图 7。检测的数量,7月数量最多、9月次之、5月受鱼类集群的影响检测数量最小。7月调查TS峰值主要集中在-66~ -62 dB区域;9月调查TS主要集中分布在-64 dB附近;5月TS存在-54 dB、-48 dB和-42 dB 3个峰值。库区投放鲢、鳙鱼苗规格大于10 cm,根据公式(1)小于-46 dB的单体目标为非鲢、鳙的其他小型鱼类等。因此,在资源评估中该部分不计入资源量内。根据不同季节目标强度的频度能够反映出浮游动物数量夏季最多、秋季数量略为降低但规格增大、春季大幅度减少的特征。7月表层1~4 m为浮游动物的散射层。5~15 m水层浮游动物和鱼类混在一起。
![]() 图 7 大伙房水库不同季节目标强度分布图Fig. 7 Frequency distribution of fish target strength in Dahuofang Reservoir in different seasons
图 7 大伙房水库不同季节目标强度分布图Fig. 7 Frequency distribution of fish target strength in Dahuofang Reservoir in different seasons2.2.2 鱼类资源密度和资源量
不同季节不同体长规格鱼类所对应的平均密度见图 8。2012年7月上游鱼类密度较高,9月体长大于20 cm规格鱼类下游密度高。2013年5月上、中、下游鱼类密度差异不大。因此,5月鱼类在库区分布比较均匀;7月主要集中在较浅的上、中游,下游分布较少;9月主要在较深的下游分布,上、中游较少。资源量调查结果见图 9。7月、9月和5月分别在4 634 t,1 522 t和1 012 t左右。7月由于上游调查时间进入夜间、船上灯光的诱集效果以及浮游动物和小气泡等影响,造成评估结果过大。5月由于鱼类以小型集群形式存在,单体检测较少且航线短,资源量评估偏小。9月水文环境垂直变动小,鱼类呈单体分散状态,较为适合单体目标检测和声学评估。调查结果与利用历年各龄鱼渔获尾数和最大年龄和最后一年的资源尾数的POPO(1972)方法推算的资源量结果1 513 t基本一致[16]。
![]() 图 8 大伙房水库不同季节上(a)、中(b)、下(c)游不同体长的鱼类密度Fig. 8 Fish density for different fish lengths in upper (a), middle (b) and lower (c) reaches of Dahuofang Reservoir in different seasons
图 8 大伙房水库不同季节上(a)、中(b)、下(c)游不同体长的鱼类密度Fig. 8 Fish density for different fish lengths in upper (a), middle (b) and lower (c) reaches of Dahuofang Reservoir in different seasons![]() 图 9 不同季节大伙房水库不同体长的鱼类资源量Fig. 9 Abundance for different fish lengths in Dahuofang Reservoir in different seasons
图 9 不同季节大伙房水库不同体长的鱼类资源量Fig. 9 Abundance for different fish lengths in Dahuofang Reservoir in different seasons![]() 图 10 不同季节不同体长鱼类垂直分布a. 2013年5月;b. 2012年7月;c. 2012年9月Fig. 10 Vertical distribution of fishery resources for different body lengths in different seasonsa. May, 2013;b. July, 2012;c. September, 2012
图 10 不同季节不同体长鱼类垂直分布a. 2013年5月;b. 2012年7月;c. 2012年9月Fig. 10 Vertical distribution of fishery resources for different body lengths in different seasonsa. May, 2013;b. July, 2012;c. September, 20122.2.3 渔业资源的垂直分布
通过对不同水层的单体目标回波检测,分析获得不同体长鱼类在不同季节的垂直分布特征,分布频度特征见图 10。5月鱼类主要集中在0~10 m水层;7月鱼类集中在5~15 m水层。体长小于50 cm的鱼类主要分布在0~15 m水层,大于50 cm的鱼类主要分布在5~15 m水层;9月鱼类主要分布于10~20 m水层,其中5~15 m水层较为集中,0~5 m水层鱼类分布密度较5月和7月少,而15 m以深水层鱼类密度较5月和7月大。
2.3 水文参数对鱼类垂直分布的影响
5月库区鱼类主要分布在0~10 m水层,即温跃层以浅水层,水温在6~12 ℃(图 2-a);7月主要集中在5~15 m的温跃层以浅水层,相比于表层较高的水温,鲢、鳙更适宜中层的水温16~21 ℃;9月主要分布在10~20 m附近水层,表层的分布少,9月水温垂直梯度变化不大(18~21 ℃)。与7月相比鱼类分布的水层范围进一步扩大(图 2-c)。不同季节的DO、pH及叶绿素a质量浓度等对鲢、鳙的分布影响不明显,受温度影响较大。
3. 讨论
该研究选取夏季、初秋和春末季节进行调查,对应库区融冰对流结束(5月)、放水(7月)和储水(9月)的水体变化过程。结合各季节主要水文参数剖面和声学调查的水平及垂直鱼类目标空间分布结果,可以获得鱼类在不同季节的行为变动规律[17-19]。该方法对研究北方冬季结冰型水库湖泊鱼类资源季节变动有较好的效果。
3.1 鱼类的垂直分布特征
对于鲢、鳙的垂直分布规律,传统的捕捞经验一般认为鲢、鳙鱼类主要分布于10 m附近的水层且较大个体分布在相对较深的水层。该研究结果显示鲢、鳙主要集中在10~15 m之间的水层,这与传统经验相吻合。而
3.2 声学调查季节的选择
秋季库区蓄水,上游径流水的影响相对夏季少,库区整体水文环境较稳定,水温随深度增加变化不是明显,声学调查结果显示,整个库区鱼类回波呈均匀分散分布,无集群现象。通过对比各季节库区水文和声学调查所得数据,再结合放苗量、渔获物抽样对2012年2~5龄资源量推算所得的结果(1 513 t,未公开),秋季进行声学调查结果1 522 t与渔获物抽样推算的资源量结果基本一致,因此大伙房水库进行声学调查的最适季节为秋季。
3.3 库区声学调查与资源量评估
根据库区狭长的地形特点,该研究声学调查采用“之”字型走航方法,其优点是可以简单避开库区内设置的定置网箔。由于调查范围相对海洋调查较小,航线的断面间隔也远小于一般海洋调查[8]。从调查结果看,夏秋季节库区内的鲢、鳙基本分布较为密集和连续,调查航线基本满足随机性和相邻航线的非相关性的声学采样要求,能够较好地反映鱼类在库区内的水平和垂直分布。由于使用较小的快艇调查,航行稳定性较差,航迹线与设计航线相比存在偏差。但是考虑到夏秋季节鱼类较均匀分布的现状,对资源量评估影响不大。
《渔业资源声学调查与评估》规范[8]要求配合网具的生物采样,用于识别鱼种和确定体长分布。由于该研究中无法进行拖网采样,参考定置网箔渔获数据进行鱼种分配。但网具的大规格鱼类选择性导致评估产生误差,今后可以使用无选择性的三重刺网进行采样,配合声学调查。大伙房鱼类种类较多,约11种,根据历年的生产,鳙占52%以上,因此资源评估模型按鳙的声学参数进行,虽有一定的偏差,但考虑到其他鱼类的体长范围较大规格鲢、鳙要小很多,对较大个体鱼类资源量评估影响较小。
淡水鱼类的TS测量相对较少[10-11],现在一般参考相近鱼类或者使用理论模型进行TS计算,影响了声学资源评估的精度[4-7],该文使用公式(1)进行其他鱼种和鲢、鳙幼鱼等的体长推算也尚不够严密,还需要在今后的工作中对不同鱼种和体长的TS进行精密测量。另外,一般水库会有植物淹没而产生的小气泡,对声学调查会有一定的影响,该研究通过连续垂直分布回波(图 4的a、b型回波)的观测进行消除[20-21]。对于气泡的自动识别还需要在将来研究。图 4和图 6中的斜线规律回波为干涉噪声,来自于小型船艇的部分材料共振,在数据后处理中消除。
声学测量技术的应用提高了淡水鱼类资源分布和资源量调查的效率,且鱼类回波数据可以长期保存,通过数据的累计和对比分析可以为揭示淡水鱼类的生态行为规律提供科学依据。
-
![]()
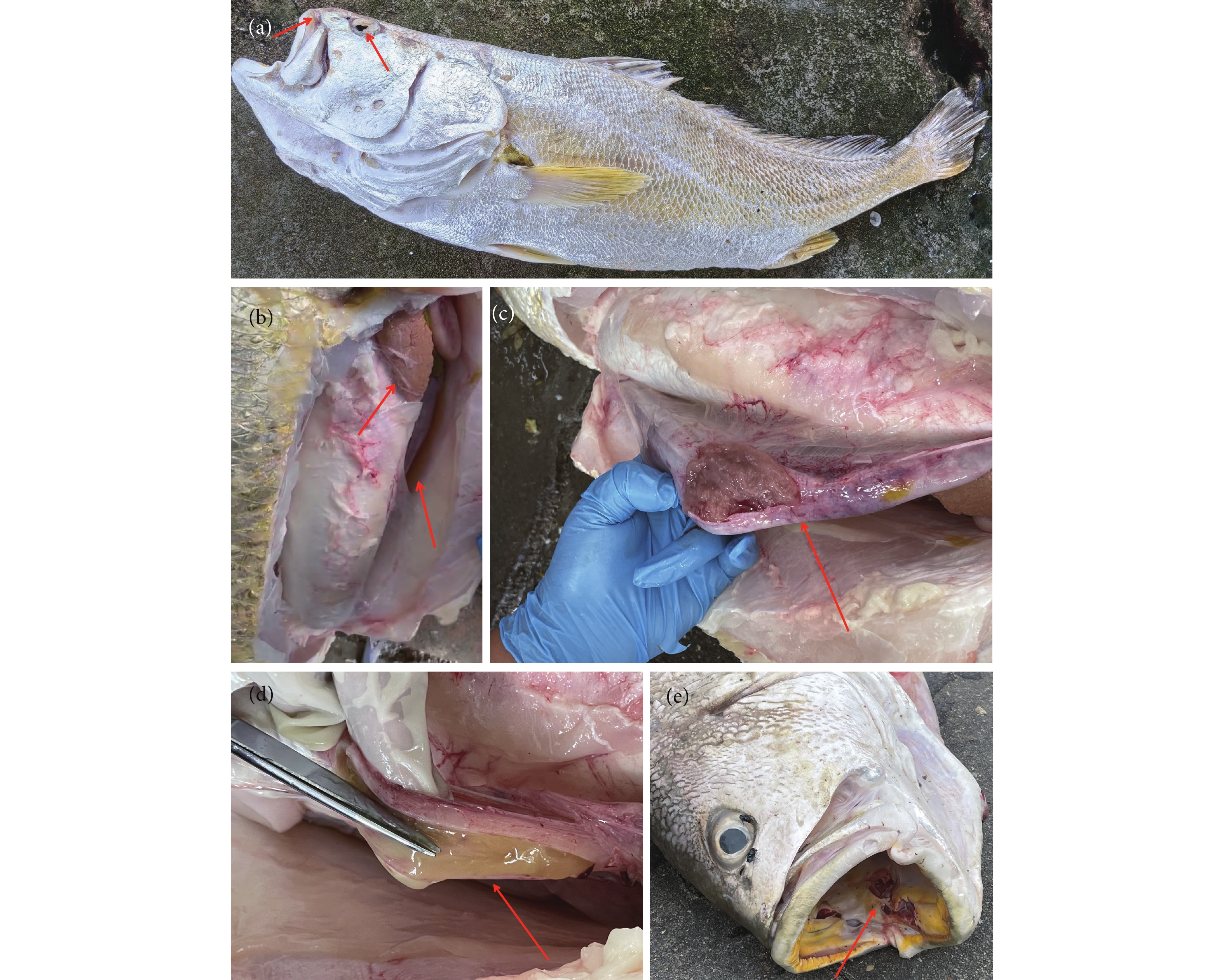
图 1 部分病死黄唇鱼的外部特征和解剖后内脏状态
注: a. 吻溃疡,眼球突出,角膜白浊溃烂;b. 肝肿大,肝脏表面有出血点,变脆呈褐色,腹腔有大量腹水;c. 肠黏膜出血、充血;d. 肠腔内有炎性渗出物;e. 口腔溃疡。
Figure 1. Symptoms of some infected B. taipingensis
Note: a. Ulcers on the snout, protruding eyeballs, corneal opacification and ulceration; b. Enlarged liver, hemorrhagic spots on the liver surface, brittle texture with brown color, and a large amount of ascites in the abdominal cavity; c. Hemorrhage and congestion of the intestinal mucosa; d. Inflammatory exudate present in the intestinal avity; e. Oral ulcer.
![]()
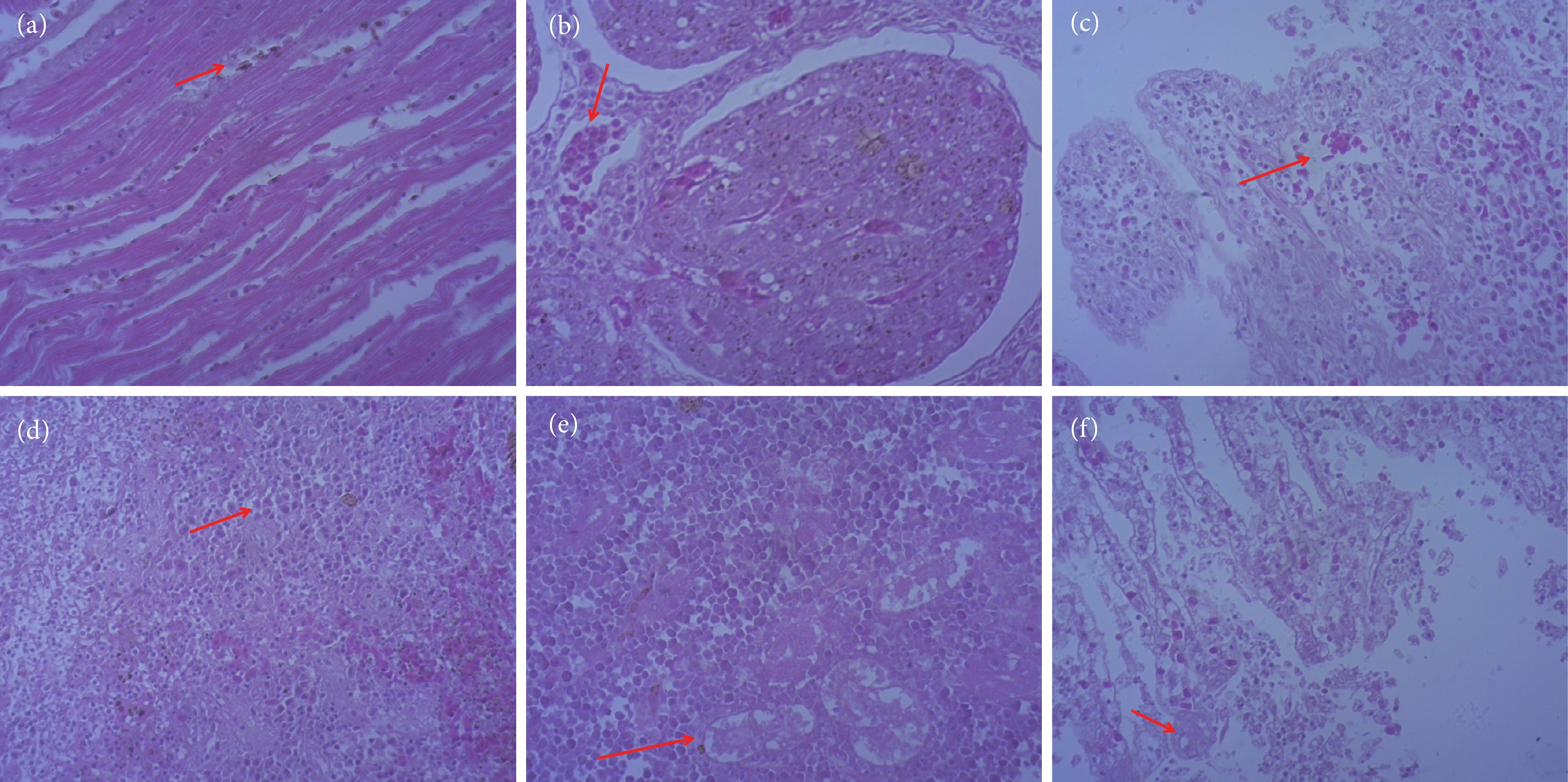
图 2 病死黄唇鱼心脏、肝脏、肠、脾、肾、鳃丝组织病理切片 (HE, 200×)
注:a. 心肌呈现淋巴细胞浸润性心肌炎;b. 肝细胞团间见淋巴细胞浸润性肝炎;c. 肠见出血、黏膜上皮细胞脱落和炎性细胞浸润;d. 淋巴细胞稀疏,内皮细胞变性坏死;e. 肾小管变性坏死,炎性细胞浸润;f. 鳃见鳃丝的脱落、出血、炎性细胞浸润。
Figure 2. Pathological sections of heart, liver, intestine, spleen, kidney and gill filaments of dead B. taipingensis (HE, 200×)
Note: a. Lymphocyte infiltration between myocardial cells; b. Lymphocyte infiltration between liver cell clusters; c. Hemorrhage, mucosal epithelial cell shedding and inflammatory cell infiltration in the intestine; d. Sparse lymphocytes, endothelial cell degeneration and necrosis; e. Renal tubular degeneration and necrosis, inflammatory cell infiltration; f. Shedding of gill filaments, hemorrhage, and inflammatory cell infiltration in the gills.
![]()
图 3 菌株DG230920的菌落形态、显微形态
注:a. TSA 培养基上的菌落形态;b. 血琼脂平板培养48 h后的菌落形态;c. 革兰氏染色的镜检图片 (1 000×)。
Figure 3. Morphology and microphotograph of DG230920 on TSA plate
Note: a. Colony morphology of DG230920 on TSA plate; b. Colony morphology on blood nutrient agar after 48 h; c. Gram's stain microphotograph of DG230920 (1 000×).
![]()
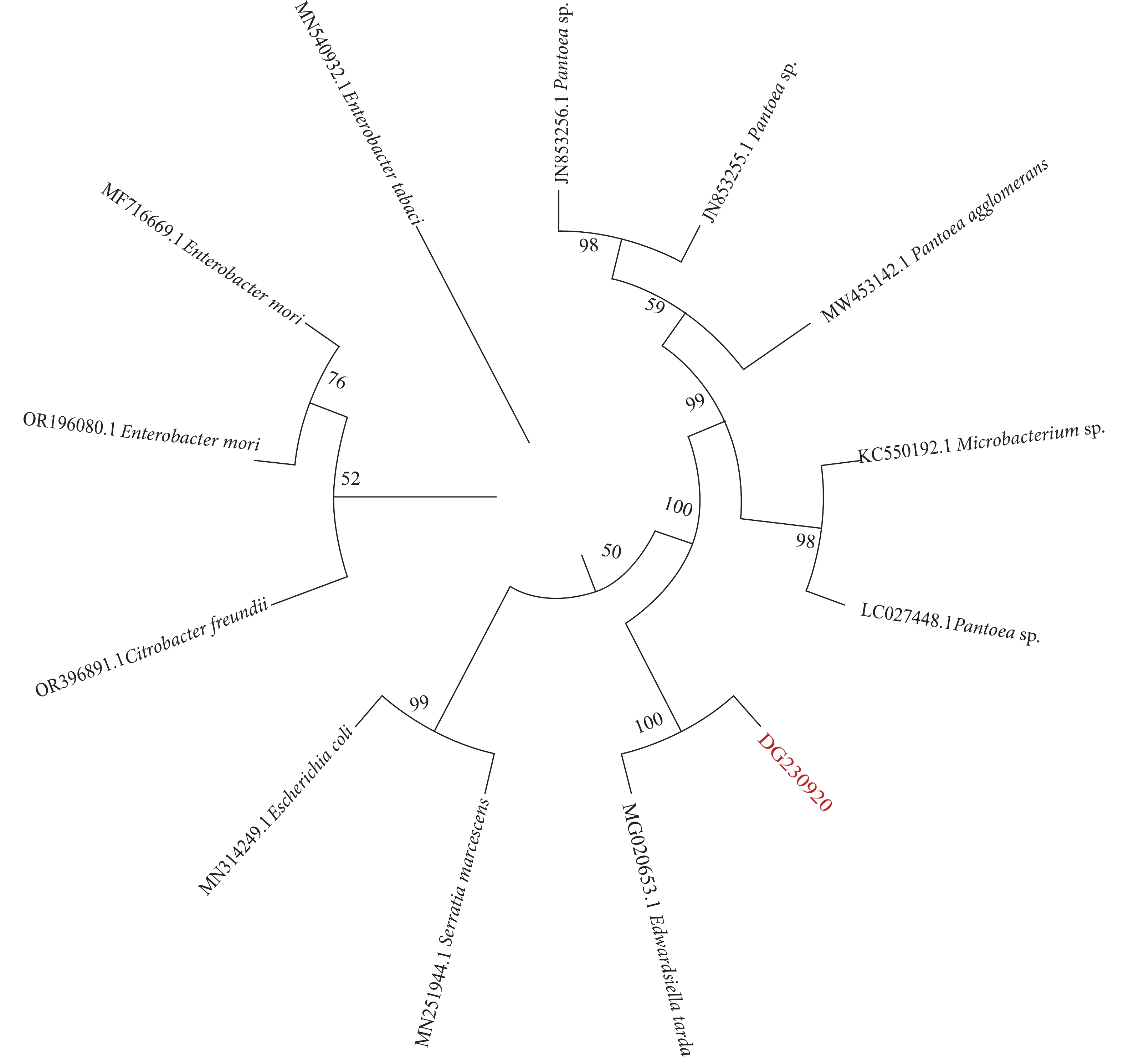
图 5 分离菌株DG230920基于16S rRNA序列的系统进化树
Figure 5. Phylogenetic tree based on 16S ribosomal RNA sequences
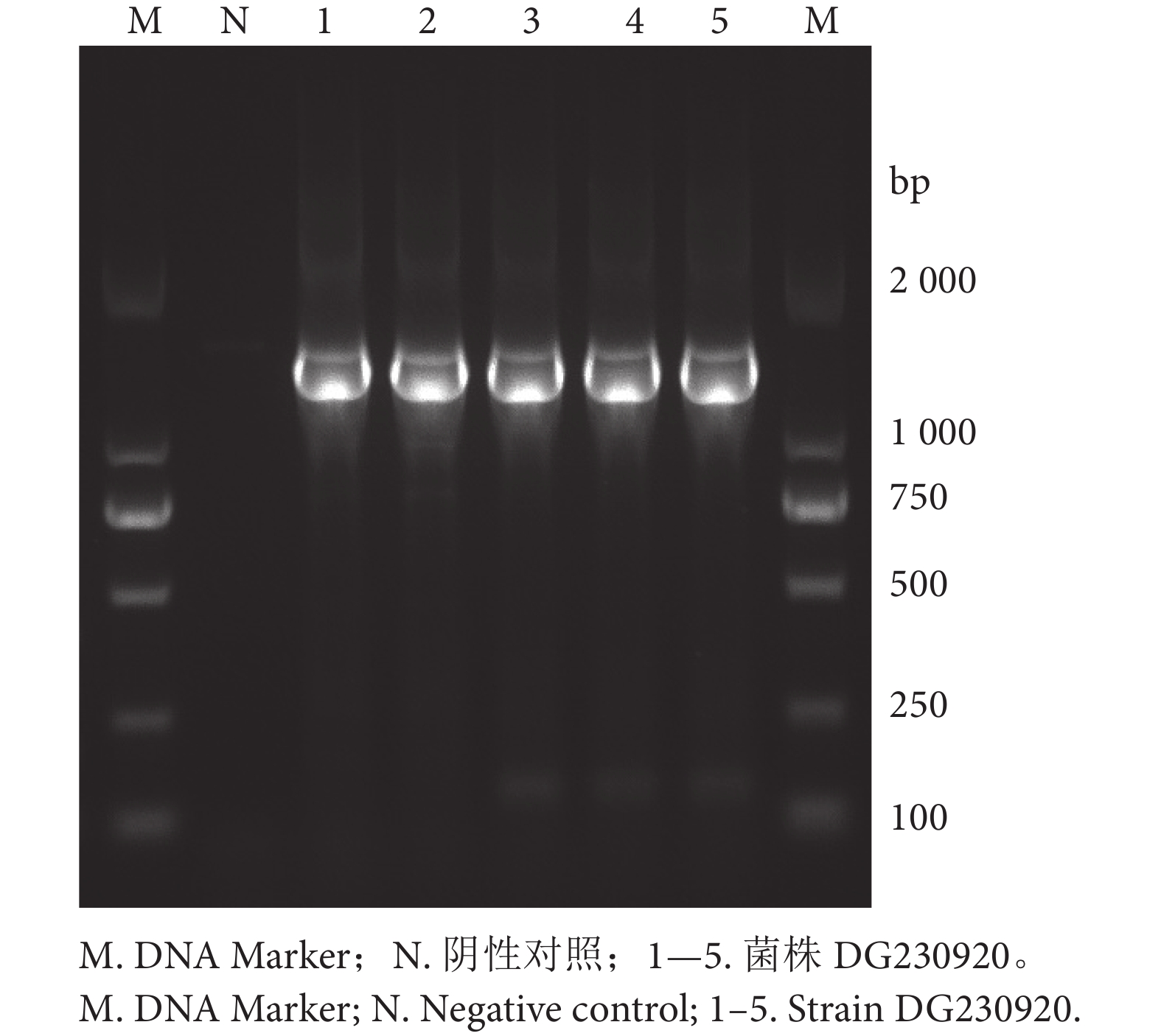
表 1 16S rRNA引物、反应体系及反应程序
Table 1 Primers of 16S rRNA, reaction and protocol
引物
Primer引物序列 (5'—3')
Primer sequence (5'–3')反应体系 Reaction mixture 反应程序
Protocol片段大小
Size/bp组分
Component用量
Volume/µL上游引物
Upstream primerAGAGTTTGATCCTGGCTCAG PCR Master Mix 12.5 94 ℃预变性2 min;94 ℃变性45 s,52 ℃退火1 min,72 ℃延伸2 min,35 次循环;72 ℃延伸7 min;4 ℃保温 约1 500 上游引物 1.0 下游引物
Downstream primerGGTTACCTTGTTACGACTT 下游引物 1.0 模板 2.0 ddH2O 8.5  下载: 导出CSV
下载: 导出CSV
表 2 DG230920菌株的生理生化鉴定结果
Table 2 Physiological and biochemical identification results of DG230920
生化项目
Biochemical
item结果
Result生化项目
Biochemical
item结果
Result生化项目
Biochemical
item结果
Result生化项目
Biochemical
item结果
Result生化项目
Biochemical
item结果
Result丙氨酸-苯丙氨酸-
脯氨酸芳胺酶(APPA)− 侧金盏花醇
(ADO)− L-吡咯烷酮芳胺酶
(PyrA)− L-阿拉伯醇
(IARL)− D-纤维二糖
(dCEL)− β-半乳糖苷酶
(BGAL)− Β-N-乙酰氨基葡糖苷酶(BNAG) + 谷氨酰芳胺酶 pNA
(AGLTp)− D-葡萄糖
(dGLU)+ Γ-谷氨酰转移酶(GGT) − 发酵/葡萄糖
(OFF)+ β-葡糖苷酶
(BGLU)− D-麦芽糖
(vdMAL)+ D-甘露醇
(dMAN)− D-甘露糖
(dMNE)+ β-木糖苷酶
(BXYL)− Β-丙氨酸芳胺酶 pNA(BAlap) − L-脯氨酸芳胺酶
(ProA)− 脂肪酶
(LIP)− 古老糖
(PLE)− 酪氨酸芳胺酶
(TyrA)− 尿素酶
(URE)− D-山梨醇
(dSOR)− 蔗糖
(SAC)− D-塔格糖
(dTAG)− D-海藻糖
(dTRE)− 柠檬酸盐(钠)
(CIA)+ 丙二酸盐
(MNT)− 5-酮基-D-葡萄糖酸盐(5KG) − M-乳酸盐碱化(N-ILATk) − α-葡糖苷酶
(AGLU)− 琥珀酸盐碱化
(SUCT)+ Β-N-乙酰氨基半乳糖苷酶 (NAGA) − Α-半乳糖苷酶
(AGAL)− 磷酸酶
(PHOS)+ 甘氨酸芳胺酶
(GlyA)− 鸟氨酸脱羧酶
(ODC)+ 赖氨酸脱羧 (LDC) + L-组氨酸同
(IHISa)− 香豆酸
(CMT)+ Β-葡萄糖醛酸酶
(BGUR)− O/129 耐药
(O129R)− 谷氨酸-甘氨酸-精氨酸芳胺酶(GGAA) − L-苹果酸盐同化
(IMLTa)− ELLMAN
(ELLM)+ L-乳酸盐同化
(ILATa)− 注:+. 阳性;−. 阴性。 Note: +. Positive; −. Negtive.
下载: 导出CSV
表 3 菌株DG230920的药敏试验结果
Table 3 Drug sensitivity test results of DG230920
抗菌剂
Antibiotic最低抑菌浓度
MIC/(mg·mL−1)结果①
Result抗菌剂
Antibiotic最低抑菌浓度
MIC/(mg·mL−1)结果①
Result氨苄西林 Ampicillin ≤2 S 厄他培南 Ertapenemc ≤0.5 S 氨苄西林/舒巴坦 Ampicillin/Sulbactam ≤2 S 亚胺培南 Imipenemc ≤1 S 哌拉西林/他唑巴坦
Piperacillin/Tazobactamm≤4 S 阿米卡星 Amikcin ≤2 S 头孢唑林 Cefazolin ≤4 S 庆大霉素 Gentamicin ≤1 S 头孢替坦 Cefotetan ≤4 S 妥布霉素 Tobramycin ≤1 S 头孢他啶 Ceftazidime ≤1 S 环丙沙星 Ciprofloxacin ≤0.25 S 头孢曲松 Ceftriaoxone ≤1 S 左氧氟沙星 Levofloxacin ≤0.25 S 头孢吡肟 Cetepime ≤1 S 呋喃妥因 Nitrofurantoin ≤16 S 氨曲南 Aztreonam ≤1 S 复方新诺明
Trimethoprim/Sulfamethoxazole≤20 S 注:① 药物的敏感度 (S代表对药物敏感)。 Note: ① Drug sensitivity (S represents sensitivity).
下载: 导出CSV
-
[1] GU Y G, HUANG H H, LIANG Y, et al. Micro-CT and SEM investigation of sound absorption structure and chambers in the otoliths of giant panda fish species: Chinese bahaba (Bahaba taipingensis)[J]. Micron, 2022, 161: 1-7.
[2] 卢伟华, 叶普仁. 黄唇鱼Bahaba flavolabiata (Lin)资源调查报告[J]. 现代渔业信息, 2002, 17(5): 10-14. [3] 莫介化, 刘文瑜, 陆丙乾, 等. 池养条件下黄唇鱼幼鱼周年生长特性研究[J]. 广东农业科学, 2023, 50(3): 129-136. [4] 李希国, 卢伟华, 张汉霞, 等. 野生黄唇鱼救护技术研究[J]. 现代渔业信息, 2011, 26(4): 14-15, 35. [5] 黄健生, 叶永昌, 黄建强. 黄唇鱼生长规律的初步研究[J]. 科学养鱼, 2020(8): 71-72. doi: 10.3969/j.issn.1004-843X.2020.08.036 [6] 区又君, 廖锐, 李加儿, 等. 黄唇鱼矢耳石的生长特性及鱼体生长的关系[J]. 江苏农业科学, 2013, 41(6): 189-192. doi: 10.3969/j.issn.1002-1302.2013.06.070 [7] HE W, LU W H, LI X G, et al. Taxonomic status of Chinese bahaba (Bahaba taipingensis) and its phylogenetic relationship with other species in the family Sciaenidae[J]. Mitochondrial DNA, 2012, 23(2): 53-61. doi: 10.3109/19401736.2011.653797
[8] 赵彦花, 区又君, 温久福, 等. 基于微卫星标记的黄唇鱼遗传多样性研究[J]. 南方水产科学, 2019, 15(4): 127-132. doi: 10.12131/20180261 [9] ZHAO L L, GAO T X, LU W H. Complete mitochondrial DNA sequence of the endangered fish (Bahaba taipingensis): mitogenome characterization and phylogenetic implications[J]. ZooKeys, 2015, 546: 181-195. doi: 10.3897/zookeys.546.5964
[10] 区又君, 温久福, 李加儿, 等. 黄唇鱼(Bahaba taipingensis)的全基因组测序和基因组特征研究[J]. 基因组学与应用生物学, 2020, 39(2): 491-498. [11] LI H Q, SONG Z C, HUI J G, et al. Vocalization behavior of Chinese bahaba (Bahaba taipingensis) during the reproduction season[J]. Mar Sci Engin, 2023, 11(4): 712. doi: 10.3390/jmse11040712
[12] 张丝雨, 张琳玲, 黄洪辉, 等. 黄唇鱼声谱特征的初步分析[J]. 南方水产科学, 2018, 14(6): 34-42. doi: 10.12131/20180090 [13] 陆丙乾, 陆昌胜, 黄建强, 等. 黄唇鱼的生物学初步研究[J]. 湖北农业科学, 2021, 60(14): 100-103. [14] HOSHINA T. On a new bacterium, Paracolobactrum anguillimortiferum n. sp.[J]. Nippon Suisan Gakkaishi, 1962, 28(2): 162-164. doi: 10.2331/suisan.28.162
[15] 邹文政, 余建明, 蔡鸿娇, 等. 迟缓爱德华氏菌感染斑马鱼的模型构建及病理分析[J]. 微生物学报, 2023, 63(12): 4686-4697. [16] 徐赟霞, 王禹, 韩进刚, 等. 一株血鹦鹉迟缓爱德华氏菌的分离鉴定及药敏试验[J]. 河北渔业, 2021(6): 8-11. doi: 10.3969/j.issn.1004-6755.2021.06.003 [17] 靳仁娉. 迟缓爱德华氏菌唾液酸酶的克隆表达与功能研究[D]. 青岛: 中国海洋大学, 2012: 1. [18] 樊威, 贺扬, 黄小丽, 等. 一株斑点叉尾鮰源爱德华氏菌的分离鉴定及药敏试验[J]. 水产科学, 2018, 37(6): 806-811. [19] 吴勇亮, 苗鹏飞, 于辉, 等. 鳜鱼致病性迟缓爱德华氏菌的分离鉴定及药敏试验[J]. 南方农业学报, 2018, 49(4): 794-799. doi: 10.3969/j.issn.2095-1191.2018.04.27 [20] 程俊茗, 万明月, 周晋扬, 等. 鲫源迟缓爱德华氏菌的分离鉴定及其毒力基因的检测[J]. 微生物学通报, 2017, 44(10): 2380-2390. [21] KORANTENG E A, SHAHARIOR H, KOFI S P F, et al. Antibacterial activity of Mallotus japonicus (L.F.) Müller Argoviensis on growth of Aeromonas hydrophila, A. salmonicida, Edwardsiella tarda and Vibrio anguillarum[J].J Appl Microbiol, 2021, 132(1): 298-310.
[22] 宝小雪, 刘华, 余莹英, 等. 牛蛙源迟缓爱德华氏菌和弗氏柠檬酸杆菌的分离鉴定及药敏试验[J]. 黑龙江畜牧兽医, 2022(1): 122-125, 129, 139. [23] 孙敬, 柯浩, 于交平, 等. 龟源迟缓爱德华氏菌分离鉴定及灭活疫苗研制[J]. 四川农业大学学报, 2020, 38(3): 353-358. [24] 朱凝瑜, 曹飞飞, 郑晓叶, 等. 中华鳖 (Pelodiscus sinensis) 迟缓爱德华氏菌 (Edwardsiella tarda) 的分离鉴定与药物敏感性分析[J]. 中国渔业质量与标准, 2018, 8(4): 65-71. doi: 10.3969/j.issn.2095-1833.2018.04.008 [25] GRIMONT P A D, GRIMONT F, RICHARD C, et al. Edwardsiella hoshinae, a new species of Enterobacteriaceae[J]. Curr Microbiol, 1980, 4(6): 347-351.
[26] 陈茂金, 黄海玲, 付党华, 等. 圈养虎迟缓爱德华氏菌的分离鉴定及药敏分析[J]. 野生动物学报, 2018, 39(1): 110-114. doi: 10.3969/j.issn.1000-0127.2018.01.016 [27] 张晓君, 陈翠珍, 房海, 等. 牙鲆迟钝爱德华氏菌吲哚阴性变异株感染症及其病原特性研究[J]. 中国人兽共患病杂志, 2004, 20(12): 1079-1083. [28] 朱壮春, 史相国, 张淑杰, 等. 牙鲆腹水病病原研究[J]. 水产科学, 2006, 25(7): 325-329. doi: 10.3969/j.issn.1003-1111.2006.07.001 [29] ZHANG X J, ZHAN W B, CHEN C Z, et al. Studies on the infection and pathogen of Edwardsiella tarda in flounder Paralichthys olivaceus[J]. Acta Hydrobiologica Sinica, 2005, 29(1): 31-37.
[30] 陈翠珍, 房海, 张晓君, 等. 牙鲆与大菱鲆病原迟钝爱德华氏菌生物学特性及系统发育学分析[J]. 高技术通讯, 2005, 15(10): 82-88. doi: 10.3321/j.issn:1002-0470.2005.10.017 [31] 李杰, 莫照兰, 茅云翔, 等. 两株养殖大菱鲆体表出血病原菌的分离鉴定[J]. 海洋科学, 2008, 32(10): 1-5. [32] XIAO J F, WANG Q Y, LIU Q, et al. Isolation and identification of fish pathogen Edwardsiella tarda from mariculture in China[J]. Aquac Res, 2008, 40(1): 13-17. doi: 10.1111/j.1365-2109.2008.02101.x
[33] 靳洪振, 王玲玲, 吴亚鑫, 等. 一株黄颡鱼源迟缓爱德华氏菌弱毒株的分离鉴定及特性分析[J]. 淡水渔业, 2023, 53(5): 22-31. doi: 10.3969/j.issn.1000-6907.2023.05.003 [34] 邓显文, 谢芝勋, 刘加波, 等. 罗非鱼迟缓爱德华氏菌的分离与鉴定[J]. 水生态学杂志, 2009, 2(1): 114-117. [35] AMANDI A, HIU S F, ROHOVEC J S, et al. Isolation and characterization of Edwardsiella tarda from fall chinook salmon (Oncorhynchus tshawytscha)[J]. Appl Environ Microbiol, 1982, 43(6): 1380-1384. doi: 10.1128/aem.43.6.1380-1384.1982
[36] 彭钟琴, 黄璐. 大口黑鲈源迟缓爱德华氏菌的分离鉴定及药敏试验[J]. 智慧农业导刊, 2021(19): 26-29. [37] 吴静, 王庚申, 柳敏海, 等. 美洲黑石斑迟缓爱德华氏菌分离、鉴定及致病性研究[J]. 水生生物学报, 2020, 44(1): 143-152. doi: 10.7541/2020.017 [38] UCKO M, COLOMI A, DUBYTSKA L, et al. Edwardsiella piscicida-like pathogen in cultured grouper[J]. Dis Aquat Organ, 2016, 121(2): 141-148. doi: 10.3354/dao03051
[39] BUJÁN N, TORANZO A E, MAGARIÑOS B. Edwardsiella piscicida: a significant bacterial pathogen of cultured fish[J]. Dis Aquat Organ, 2018, 131(1): 59-71. doi: 10.3354/dao03281
[40] CASTRO N, TORANZO A E, DEVESA S, et al. First description of Edwardsiella tarda in Senegalese sole, Solea senegalensis (Kaup)[J]. J Fish Dis, 2012, 35(1): 79-82. doi: 10.1111/j.1365-2761.2011.01325.x
[41] SHETTY M, MAITI B, VENUGOPAL M N, et al. First isolation and characterization of Edwardsiella tarda from diseased striped catfish, Pangasianodon hypophthalmus (Sauvage)[J]. J Fish Dis, 2014, 37(3): 265-271. doi: 10.1111/jfd.12039
[42] 诸葛石养, 苏爱荣, 黄彦, 等. 金黄色葡萄球菌β溶血素研究进展[J]. 右江医学, 2021, 49(6): 473-476. doi: 10.3969/j.issn.1003-1383.2021.06.014 [43] 周永林. 齐墩果酸抑制β-内酰胺酶和细菌性溶血素活性作用及其机制研究[D]. 长春: 吉林大学, 2021: 45. [44] JAUNEIKAITE E, FERGUSON T, MOSAVIE M, et al. Staphylococcus aureus colonization and acquisition of skin and soft tissue infection among Royal Marines recruits: a prospective cohort study[J]. Clin Microbiol Infect, 2020, 26(3): 381.e1-381.e6. doi: 10.1016/j.cmi.2019.07.014
[45] GONZALEZ C D, LEDO C, CELA E, et al. The good side of inflammation: Staphylococcus aureus proteins SpA and Sbi contribute to proper abscess formation and wound healing during skin and soft tissue infections[J]. Biochim Biophys Acta Mol Basis Dis, 2019, 1865(10): 2657-2670. doi: 10.1016/j.bbadis.2019.07.004
[46] DOS S, ISTVAN P, QUIRINO B F, et al. Functional metagenomics asa tool for identification of new antibiotic resistance genes from natural environments[J]. Microb Ecol, 2017, 73: 479-491. doi: 10.1007/s00248-016-0866-x
[47] LEUNG K Y, WANG Q, ZHENG X C, et al. Versatile lifestyles of Edwardsiella: free-living, pathogen, and core bacterium of the aquatic resistome[J]. Virulence, 2022, 13(1): 5-18. doi: 10.1080/21505594.2021.2006890
[48] 祝镔. 黄颡鱼迟缓爱德华氏菌的分离鉴定与药物敏感实验[J]. 广西农学报, 2022, 37(6): 35-40. doi: 10.3969/j.issn.1003-4374.2022.06.008 [49] 邵建春. 黄鳝源迟缓爱德华氏菌的鉴定、分型及全基因组测序[D]. 武汉: 华中农业大学, 2016: 27. [50] 叶旭红, 林先贵, 王一明. 养殖澳洲宝石鱼迟缓爱德华氏菌的分离鉴定及致病基因的检测[J]. 淡水渔业, 2010, 40(1): 50-54. doi: 10.3969/j.issn.1000-6907.2010.01.009 -
期刊类型引用(5)
1. 高铭悦,楚馨,刘思玉,姚嘉赟,蔺凌云,沈锦玉,潘晓艺. 大口黑鲈蛙虹彩病毒传代致弱株免疫效果评价. 水生生物学报. 2025(06): 23-33 .  百度学术
百度学术
2. 黄仁善,钟纯怡,张帝,强俊,陶易凡,李岩,董亚伦,路思琪,徐跑. 不同大口黑鲈群体生长性能、肌肉营养成分、肠道形态结构及菌群组成的比较研究. 动物营养学报. 2024(02): 1158-1172 . 百度学术
3. 李硕,陈静妮,赵立宁,黄春萍,黄锦炉,王贵平,仲颖. 抗大口黑鲈蛙虹彩病毒卵黄抗体的制备及其间接ELISA检测方法的建立. 南方水产科学. 2024(02): 129-139 . 本站查看
4. 傅建军,龚雅婷,朱文彬,强俊,张林兵,王兰梅,罗明坤,董在杰. 大口黑鲈微卫星多重PCR体系及3个群体的遗传分析. 水生生物学报. 2023(09): 1514-1522 . 百度学术
5. 张涛,周剑光,张林,甘金华,何力. 大口黑鲈形态特征、染色体核型及眼晶状体乳酸脱氢酶的电泳分析. 西北农业学报. 2023(10): 1503-1511 . 百度学术
其他类型引用(4)













计量
- 文章访问数: 187
- HTML全文浏览量: 34
- PDF下载量: 46
- 被引次数: 9
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
