
Analysis of effects of environmental factors on Sthenoteuthis oualaniensis based on structural equation model
-
摘要:
为探究鸢乌贼 (Sthenoteuthis oualaniensis) 资源变化与海洋环境因子的关系,基于2019—2020年南海围网捕捞鸢乌贼的电子渔捞日志数据,首先分析了鸢乌贼资源的月间变动特征,之后利用结构方程模型解析海洋环境因子对其资源时空分布的影响。结果表明:围网月间的平均单位捕捞努力量渔获量 (Catch per unit effort, CPUE) 变化趋势基本一致,年平均CPUE则为2020年大于2019年。南海鸢乌贼渔汛期为3—4月,高产区域集中在112°E—117°E、8°N—12°N,渔汛期CPUE呈现向东和向北偏移的趋势。海洋环境对鸢乌贼资源分布的综合影响系数为0.38,而海表盐度和光合有效辐射在海洋环境上的载荷量分别为0.87和0.82,两者是影响鸢乌贼资源分布的重要环境因子。所用结构方程模型为量化海洋环境因子与鸢乌贼资源分布之间的复杂关系提供了新的研究思路,可为鸢乌贼资源可持续利用和管理提供参考依据。
Abstract:To understand the relationship between variations in Sthenoteuthis oualaniensis resources and marine environmental factors, we conducted an analysis of monthly variations in S. oualaniensis resources based on the electronic fishing log data from purse seine operations in the South China Sea from 2019 to 2020. Then we examined the impact of marine environmental factors on the spatiotemporal distribution of S. oualaniensis resources by using a structural equation model. The results indicate that the catch per unit effort (CPUE) of purse seine average presented a consistent monthly trend but with significant interannual fluctuations, having higher average CPUE values in 2020 as compared with 2019. The fishing season for S. oualaniensis in the South China Sea is observed in March and April, primarily concentrated within the region between 112°E−117°E and 8°N−12°N, with a tendency for CPUE to shift eastward and northward during the fishing season. On the other hand, the comprehensive impact coefficient of marine environmental factors on the distribution of S. oualaniensis resources was calculated as 0.38, with sea surface salinity and photosynthetically active radiation exhibiting significant loadings of 0.87 and 0.82, respectively, indicating their importance in influencing the distribution of S. oualaniensis resources. The application of a structural equation model offers a quantitative research approach to examine the complex relationship between marine environmental factors and the distribution of S. oualaniensis. The research provides valuable scientific insights for the sustainable utilization and management of S. oualaniensis resource.
-
生物与环境的关系是生态学研究的核心内容[1],生物资源变化究竟受到哪些环境因子的影响需要从多角度考虑,目前可观测、可测量的环境变量很可能是不全面的,在建模研究分析或现场监测中很可能遗漏了某些重要的环境变量。鸢乌贼 (Sthenoteuthis oualaniensis) 是一种中上层大洋性头足类,广泛分布于印度洋和中国南海海域,具有生长周期短、资源储量大等特点[2-4]。据张俊等[5]的调查评估显示,截至2018年,南海鸢乌贼资源可捕捞量超过500 万吨。作为中国传统作业渔场捕捞对象,鸢乌贼价值高,是重要的经济捕捞物种[6]。其资源变化与海洋环境密切相关,海洋环境对鸢乌贼资源的增长、变动、分布均会产生直接影响。
目前,国内外学者已开展的相关研究主要集中于探究一种或多种环境因子对鸢乌贼资源变化的影响。范江涛等[7]基于栖息地模型,利用海表温度 (Sea surface temperature, SST)、叶绿素a浓度 (Chlorophyll a concentration, Chl a)、海表盐度 (Sea surface salinity, SSS)、海表高度 (Sea surface height, SSH) 研究了海洋环境与鸢乌贼生物量的关系。余景等[8]运用广义线性模型对鸢乌贼单位捕捞努力量渔获量 (Catch per unit effort, CPUE) 进行标准化,同时结合海表温度、叶绿素a浓度、海面风场分析了鸢乌贼资源与海洋环境的关系。目前常用的统计模型在综合分析多个环境变量与渔业资源的关系时,主要倾向于考虑线性关系和单一因子的作用。这些模型在量化未纳入模型中的其他环境因子 (如潜变量) 的影响程度上仍不理想[9-11]。结构方程模型是一种多变量统计方法,属于先验性模型,可根据已有的理论背景假设模型,再对模型进行评估和修正。它的优点在于能同时将多个环境变量纳入模型进行分析、整体拟合,并用这些可观测的环境变量构建出一个无法观测的整体潜变量 (海洋环境),进而分析海洋环境对鸢乌贼资源的影响程度,且允许自变量和因变量都包含测量误差,能进一步提高模型结果的准确性[12-13]。该模型有助于更全面了解海洋环境因子对鸢乌贼资源变化的影响。但目前结构方程模型在渔业领域上的应用较少[14],在鸢乌贼资源生态学的研究中尚未见报道。
本研究基于我国南海海域围网作业捕捞获得的鸢乌贼数据,构建结构方程模型 (Structural Equation Model, SEM),观测变量 (现实中可测量到的变量) 选择为SST、SSS、SSH、Chl a、光合有效辐射 (Photosynthetically active radiation,PAR) 以及4种不同计算方式获得的CPUE,潜变量 (无法观测到的一种抽象概念的变量) 设置为海洋环境和鸢乌贼资源。用以上5个环境因子数据表征潜变量 (海洋环境);用4种不同计算方式的CPUE表征潜变量 (鸢乌贼资源)。旨在从整体上揭示海洋环境与鸢乌贼资源之间的响应关系,为鸢乌贼资源变动的机制和可持续利用提供基础依据。
1. 材料与方法
1.1 数据来源
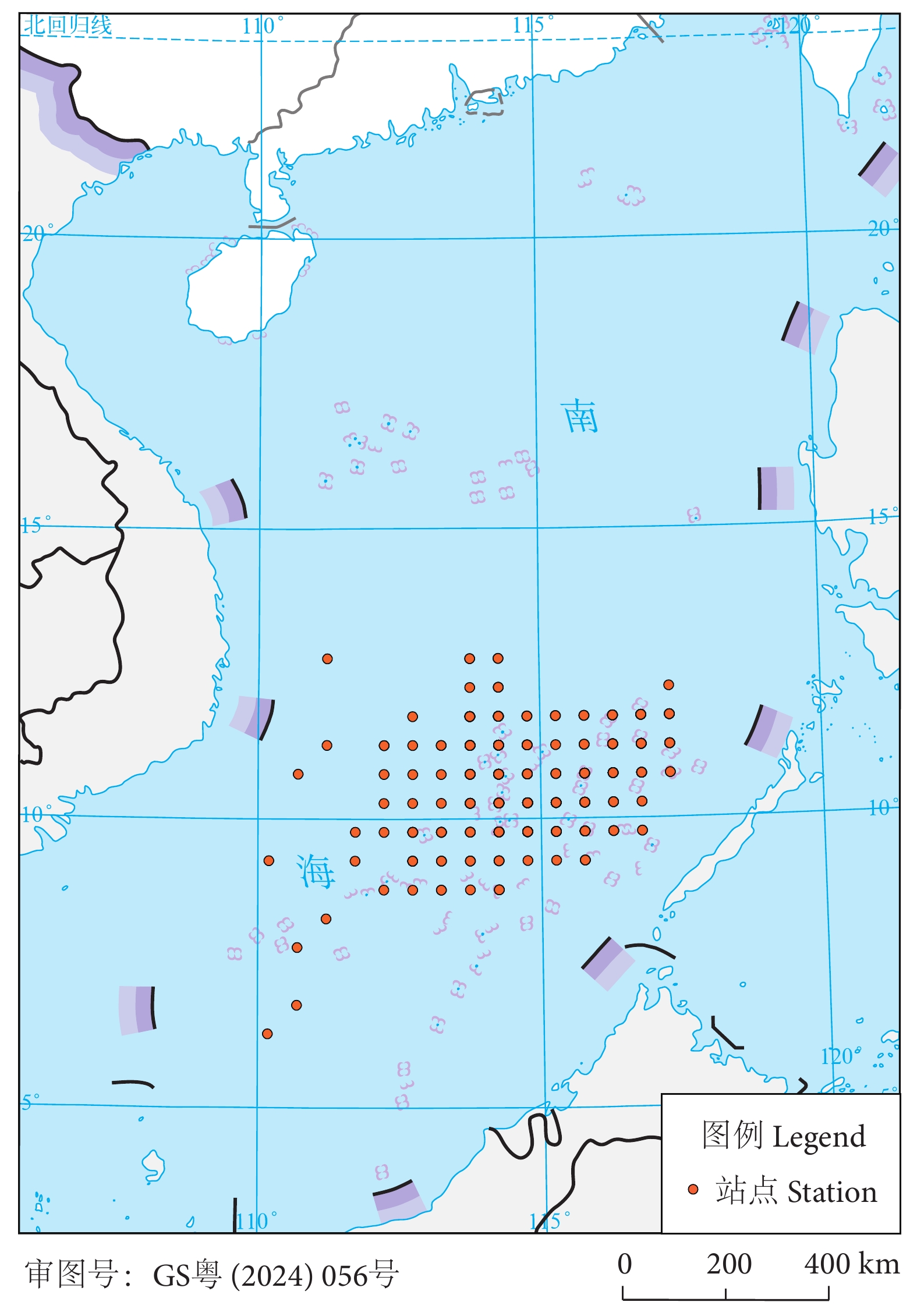
鸢乌贼数据来自中国水产科学研究院南海水产研究所北斗船位监控平台记录的2019年2月—2020年12月的电子渔捞日志,作业方式为围网,捕捞站点分布在110°E—118°E、6°N—13°N (图1)。以0.5°×0.5° 网格中心点代表捕捞渔区的经纬度。将捕捞日志中功率、作业方式、渔获量或时空信息空缺的记录删除,检查功率、作业时长或渔获量数值过大或过小,将含有异常值的记录删除。
![]() 图 1 南海2019—2020年围网捕捞站点分布图Fig. 1 Sampling stations for seine fishing in the South China Sea during 2019−2020
图 1 南海2019—2020年围网捕捞站点分布图Fig. 1 Sampling stations for seine fishing in the South China Sea during 2019−20201.2 数据处理
所选的海洋环境变量包括:海表温度、叶绿素a浓度、海表高度、海表盐度和光合有效辐射,时间分辨率为月,按渔区0.5°×0.5° 进行统计整理,同一渔区网格内数据取平均值,作为该网格中心对应的环境数据值,数据均来自夏威夷大学卫星遥感数据 (https://oceanwatch.pifsc.noaa.gov/erddap/index.html)。
使用4种不同公式计算的CPUE作为结构方程模型潜变量的观测变量,其优点是可以多角度观察鸢乌贼的资源变化,每个计算方法涵盖了资源的特定方面,反映了资源不同的变化机制,多种公式还可以减少由于测量误差或不确定性引起的问题,提高潜变量的可靠性。计算公式[15-16]为:
$$ \begin{array}{c}{\mathrm{C}\mathrm{P}\mathrm{U}\mathrm{E}}_{1}{\text{=}}\dfrac{C}{P\times T} \end{array} $$ (1) $$ \begin{array}{c}{\mathrm{C}\mathrm{P}\mathrm{U}\mathrm{E}}_{2}{\text{=}}\dfrac{C}{T} \end{array} $$ (2) $$ \begin{array}{c}{\mathrm{C}\mathrm{P}\mathrm{U}\mathrm{E}}_{3}{\text{=}}\dfrac{C}{P} \end{array} $$ (3) $$ \begin{array}{c}{\mathrm{C}\mathrm{P}\mathrm{U}\mathrm{E}}_{4}{\text{=}}\dfrac{C}{W} \end{array} $$ (4) 式中:C表示当天捕捞的总渔获量 (kg);P表示该渔船的功率 (kW);T表示当天捕捞的总作业时长 (h);W表示当天捕捞的总网次 (次)。
1.3 研究方法
1.3.1 鸢乌贼资源分析
分析作业区域内月渔获量和平均CPUE1的变化规律,以及渔汛期鸢乌贼资源CPUE2在空间上的分布情况。相对于其他3个计算公式,CPUE1综合了更多相关因素,提供了对资源变化的综合性评估,渔汛期使用CPUE2则是为了更直观表达渔汛期渔获量大小的程度。
1.3.2 结构方程模型的建立
结构方程模型是一种探究多种变量间关系及效应的方法。方程由测量模型和结构模型组成,前者用来表示无法观测到的潜变量和可观测的测量变量之间的关系,后者用来表示潜变量和潜变量之间的关系。计算公式为:
$$ \begin{array}{c}x{\text{=}}{\Lambda }_{x}\xi {\text{+}}\delta \end{array} $$ (5) $$ \begin{array}{c}y{\text{=}}{\Lambda }_{y}\eta {\text{+}}\varepsilon \end{array} $$ (6) $$ \begin{array}{c}\eta {\text{=}}{ B}\eta {\text{+}}\varGamma \xi {\text{+}}\zeta \end{array} $$ (7) 式中:方程 (5) 和 (6) 是测量模型,方程 (7) 是结构模型;x为外生可观测变量向量;y为内生可观测变量向量;ξ 为外生潜变量向量;η 为内生潜变量向量;Λx 和Λy分别为指标变量x和y的因子负荷量;B为内生潜变量的关系矩阵;Γ为外生潜变量对内生潜变量的影响;δ和ε为测量模型的误差项;ζ为结构模型的误差项。
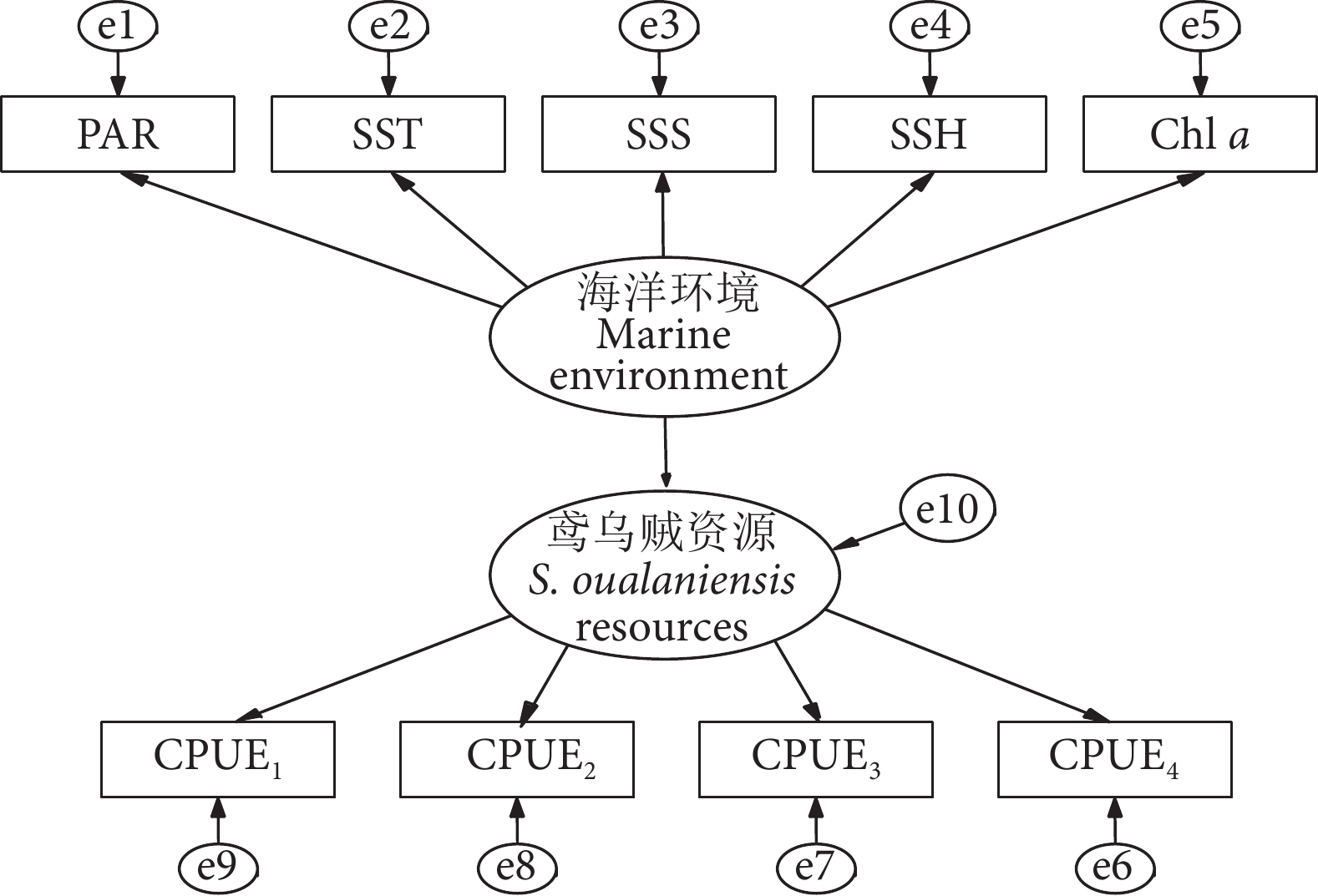
研究表明,鸢乌贼的生物量受到海洋环境的影响[17-18]。例如海表温度[19]通过影响头足类饵料食物的丰度[20-21],进而引起鸢乌贼资源的变化。叶绿素a浓度决定了浮游动物的分布,在一定程度上影响了鸢乌贼的生物量[22-23]。此外,海表盐度、海表高度、光合有效辐射等均对鸢乌贼生物量产生了影响[24-27]。这些变量与鸢乌贼的分布、繁殖和生态习性等密切相关,因此在模型中选择上述变量作为潜变量海洋环境的指标,可满足方程对于测量指标之间相关性和解释力的要求。基于上述考虑和数据内容,建立关于海洋环境和鸢乌贼资源关系的概念模型 (图2),模型包含2个潜变量,9个观测变量,e1—e10是残差变量,假设潜变量海洋环境对鸢乌贼资源具有正向影响。使用R语言“Lavaan”程序包运算模型,使用Amos 24.0绘制模型路径图。运算后对模型进行拟合指标评价,当卡方检验P>0.05,CFI (比较拟合指数,Comparative fit index) 和GFI (拟合优度指数,Goodness of fit index) 大于0.9,RMSEA (近似误差均方根,Root mean square error of approximation) 和SRMR (标准化残差均方和平方根,Standardized root mean square residual) 小于0.08时,模型拟合较为理想[28]。
![]() 图 2 南海海洋环境与鸢乌贼资源响应关系概念模型Fig. 2 Conceptual model of the relationship between marine environment and S. oualaniensis resources in South China Sea
图 2 南海海洋环境与鸢乌贼资源响应关系概念模型Fig. 2 Conceptual model of the relationship between marine environment and S. oualaniensis resources in South China Sea2. 结果
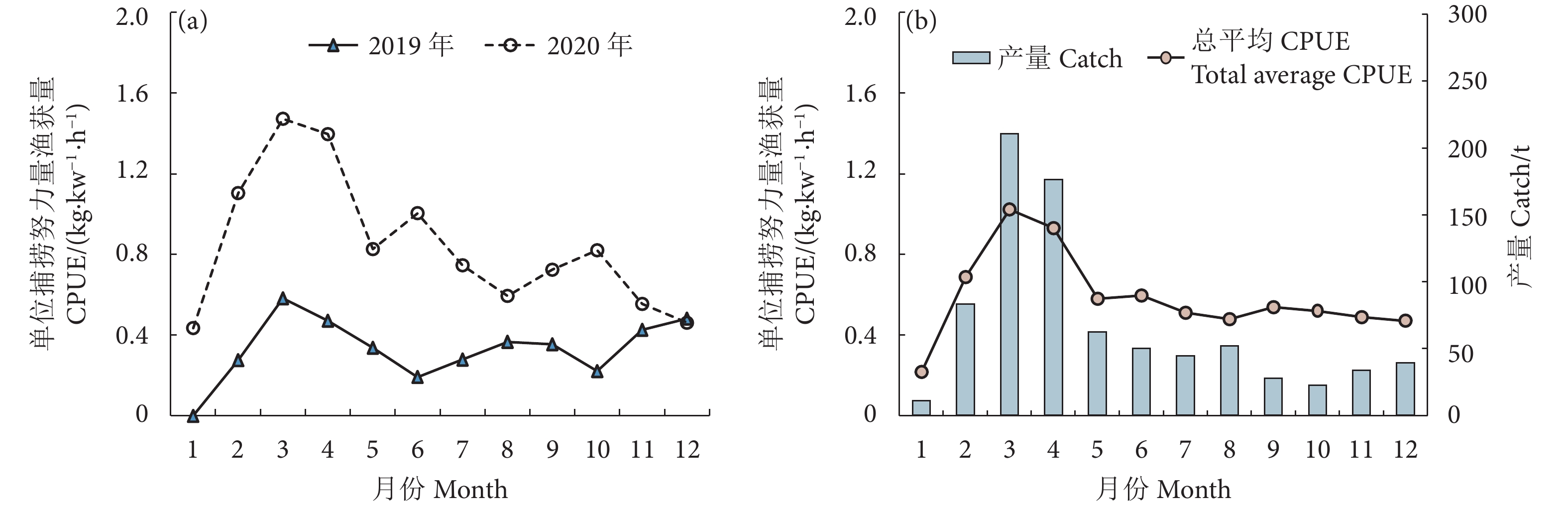
2.1 月均CPUE1和平均产量的变化规律
围网CPUE (kg·kW−1·h−1) 在两年中差异较大,在月间的变化趋势基本一致 (图3-a)。其中:2020年的围网平均CPUE整体上大于2019年,每年CPUE均在3、4月处于高位。从总平均产量分布和总平均CPUE变化趋势看 (图3-b),南海鸢乌贼的渔汛期主要集中在3、4月。围网鸢乌贼产量和CPUE在1—3月呈上升趋势,1月产量和CPUE为全年最低 (产量为10.82 t,CPUE为0.22 kg·kW−1·h−1),在3月达到峰值 (产量为210.62 t,CPUE为1.03 kg·kW−1·h−1),4—5 月下降幅度较大,之后至12 月之间趋于平缓。
![]() 图 3 2019—2020年围网平均单位捕捞努力量渔获量 (a) 和总平均产量、总平均单位捕捞努力量渔获量分布 (b)Fig. 3 Distribution of average CPUE (a), total average catch and total average CPUE (b) for seine during 2019−2020
图 3 2019—2020年围网平均单位捕捞努力量渔获量 (a) 和总平均产量、总平均单位捕捞努力量渔获量分布 (b)Fig. 3 Distribution of average CPUE (a), total average catch and total average CPUE (b) for seine during 2019−20202.2 渔汛期CPUE2的空间分布
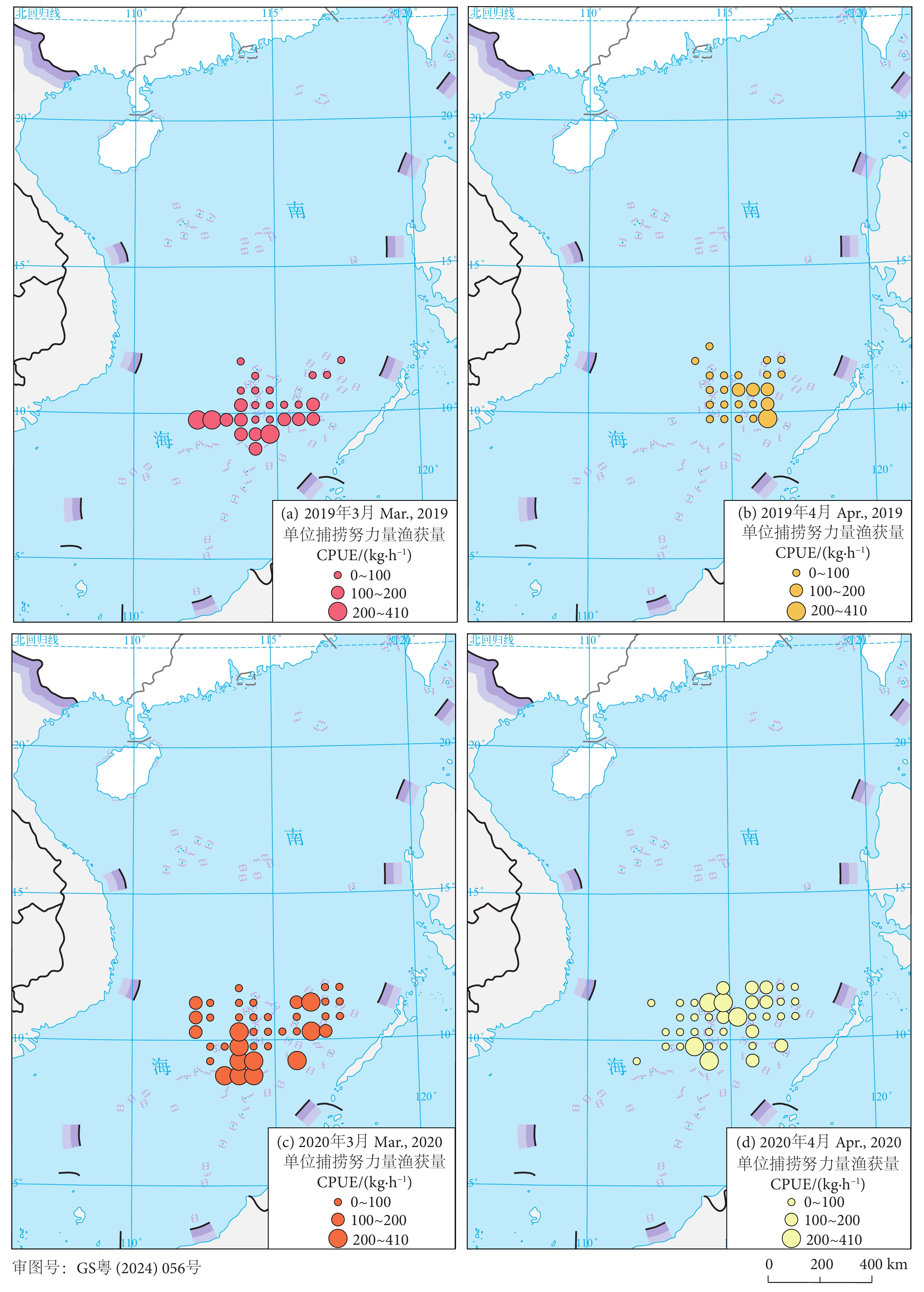
鸢乌贼渔汛期 (3—4月) CPUE在空间上出现较大变动。2019年作业渔场出现自西向东偏移的趋势 (图4-a—4-b),3月主要作业区域集中在112°E—117°E、9°N—11°N,CPUE峰值为218.75 kg·h−1;4月较3月作业区域向东偏移,主要区域在115°E—117°E、9°N—11°N,CPUE峰值为215.35 kg·h−1。
![]() 图 4 2019—2020年春季鸢乌贼渔汛期单位捕捞努力量渔获量空间分布Fig. 4 Spatial distribution of spring CPUE for S. oualaniensis during 2019−2020
图 4 2019—2020年春季鸢乌贼渔汛期单位捕捞努力量渔获量空间分布Fig. 4 Spatial distribution of spring CPUE for S. oualaniensis during 2019−20202020年鸢乌贼作业渔场出现自南向北偏移的趋势 (图4-c—4-d),3月主要作业区域集中在112°E—117°E、8°N—12°N,CPUE峰值为406.25 kg·h−1;4月较3月作业区域向北偏移,主要区域在113°E—117°E、9°N—12°N,CPUE峰值为347.92 kg·h−1。
2.3 SEM模型计算结果分析
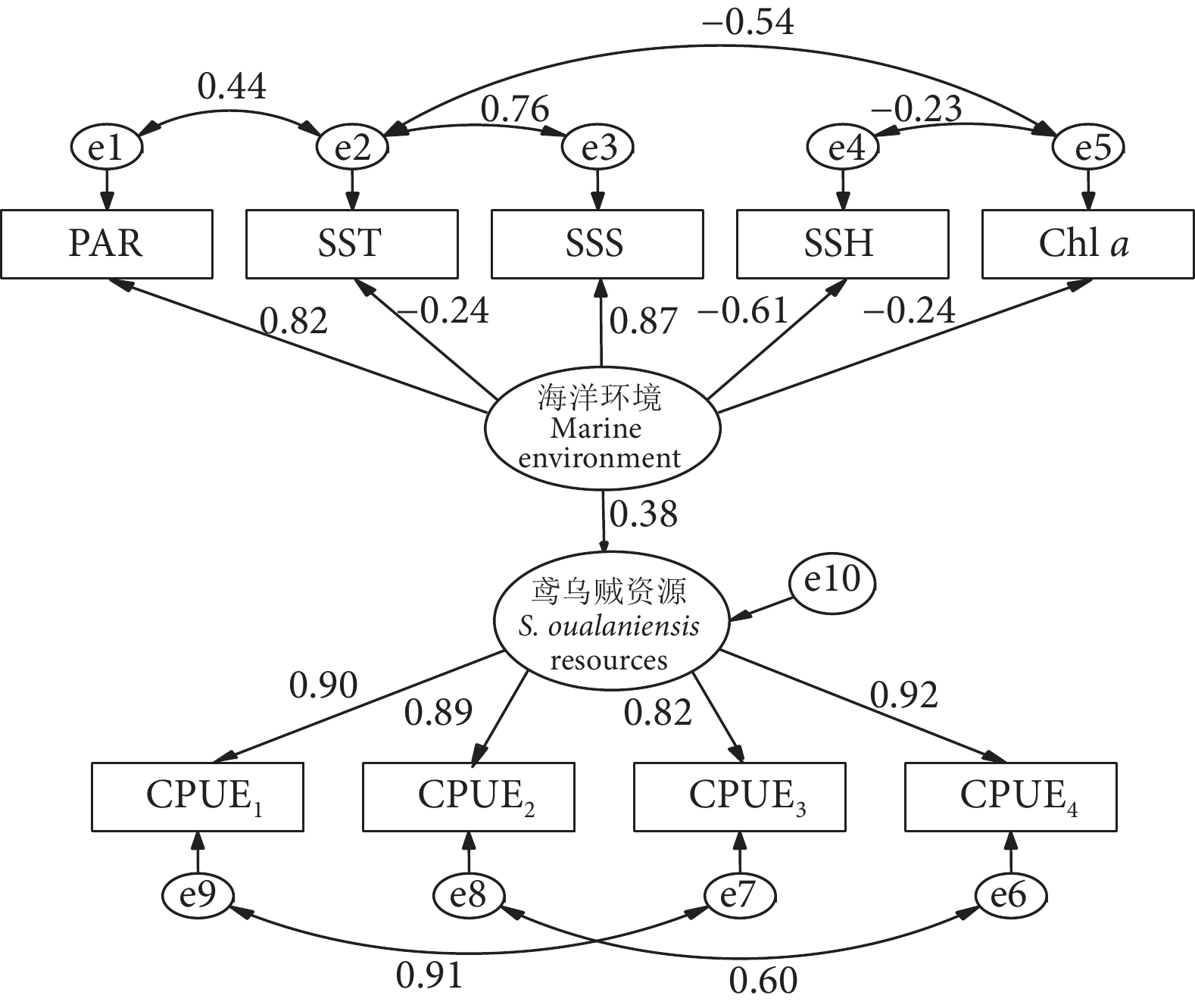
建模完成后,围网捕捞作业数据的模型评价指标如下:显著性概率P=0.01<0.05、RMSEA=0.057<0.08、SRMR=0.07<0.08、CFI=0.99>0.9、GFI=0.98>0.9。表明结果符合事实逻辑,模型拟合指标良好,潜变量与观测变量设置合理,因此接受该模型,模型假设成立。即潜变量海洋环境对鸢乌贼资源具有正向影响 (环境越适宜,资源分布越集中)。
在结构模型中,潜变量海洋环境对鸢乌贼资源具有正向影响,影响路径系数为0.38 (图5)。影响路径系数是变量之间直接关系的度量,箭头上的数值及方向表示一个变量对另一个变量的直接影响强度,范围介于 −1~1 (负数为负相关,正数为正相关),绝对值越大表示变量之间的关系越强烈。图中海洋环境与鸢乌贼资源呈正相关关系,一个标准单位的海洋环境发生变化会引起0.38个标准单位的鸢乌贼资源发生变化。
![]() 图 5 南海海洋环境与鸢乌贼资源响应关系修正模型Fig. 5 Revised model of response relationship between marine environment and S. oualaniensis resources in South China Sea
图 5 南海海洋环境与鸢乌贼资源响应关系修正模型Fig. 5 Revised model of response relationship between marine environment and S. oualaniensis resources in South China Sea在测量模型中,潜变量海洋环境上载荷量最大的因子为海表盐度,其次为光合有效辐射,载荷量分别是0.87和0.82。载荷量是表示潜变量和与之相关的观测变量之间连接强度的数值,范围介于−1~1 (负数为负向影响,正数为正向影响),绝对值越大表示观测变量对潜变量的解释力越强。因此海表盐度和光合有效辐射对潜变量的影响最大,从而在对鸢乌贼资源的影响中作用最强。其余环境因子影响程度从大到小依次为海表高度、海表温度和叶绿素a浓度 (−0.61、−0.24 和 −0.24)。潜变量鸢乌贼资源上4种不同CPUE的载荷量均较高,分别为0.90、0.89、0.82、0.92,较高的载荷量表明相关的CPUE变量与鸢乌贼资源潜变量之间的关系非常密切,观测变量能很好地反映出潜变量的变化。
3. 讨论
3.1 鸢乌贼资源变动的影响因素
围网是基于鸢乌贼具有趋光性的原理[29],针对中上层鱼类资源,其所获的数据能较好地反映出鸢乌贼资源变动的规律。从平均CPUE及产量变化看,年间差异明显,月间变化趋势基本一致。这可能与鸢乌贼的繁殖规律以及大尺度范围下的海洋环境变化有关,从而影响其所在区域的丰度和分布。已有研究表明海表温度[30]、海表盐度[31]、海表高度[32]、叶绿素a浓度[33]等均能对鸢乌贼资源变化产生影响。鸢乌贼渔汛期集中在3—4月且2019年呈现向东偏移的趋势,2020年呈现向北偏移的趋势。这可能是由于南海地区春季时海洋环境条件相对稳定,水温适中、光照充足,浮游生物和其他饵料资源也相对较丰富。适宜的生长条件促使鸢乌贼在特定的地点进行交配和产卵[34-36],导致其所在地区的丰度增加,从而增加了渔民捕捞到鸢乌贼的机会。此外,周艳波等[19]研究显示2013—2018年南海鸢乌贼渔场月重心1月主要分布在110.4°E、9.6°N;2—5月开始大幅度向东北方向移动,平均渔场重心在112.8°E、12.4°N,这期间鸢乌贼CPUE最高,作业范围最广。此发现与本研究结果基本一致。
3.2 结构方程模型结果分析
鸢乌贼生活在海域的中上层,受到环境的影响较大。结构方程模型显示每个观测变量均能对鸢乌贼资源产生影响。在测量模型中,海表盐度和光合有效辐射具有较高的载荷量 (0.87和0.82),是影响鸢乌贼生长、繁殖和分布的最重要因素。这主要是由于南海受到春季东南季风和夏季西南季风的影响,形成较强的上升流区域,上升流的交汇会导致盐度等环境因子改变,致使深海水体涌至表层,在带来大量营养物质的同时,提高了浮游生物量。而光合有效辐射是海洋初级生产力水平的重要驱动因子,因此聚集了大量的中上层鱼类,给鸢乌贼提供了充足的饵料食物,促使其形成鸢乌贼重要的生长和繁殖场所[8,24,37]。因此海表盐度和光合有效辐射对于衡量海洋生态环境起着关键作用。其余的环境因子如海表温度、海表高度和叶绿素a浓度的载荷量相对较低 (分别为−0.24、−0.61和−0.24),原因可能是这些指标与其他海洋环境指标存在较高的相关性,独立性较低。在测量模型中,如果指标之间存在高度相关性,其载荷量可能会降低。原因是它们无法提供独立的信息[38],且只是海洋环境变化的一部分,无法全面反映鸢乌贼资源分布的关键因素,从而导致其载荷量较低[39]。但是这些环境指标仍然可能在特定条件下对鸢乌贼的分布产生重要影响,需要进一步研究。
在结构模型中,海洋环境对鸢乌贼资源的正向影响系数为0.38,表明海洋环境的变化可能会直接或间接地影响鸢乌贼的生存、繁殖和分布。在鸢乌贼的生态食物链中,海洋环境如海表温度的变化可能会影响浮游动物的生长和分布,从而改变鸢乌贼的食物可获得性。叶绿素a浓度与浮游动物的数量和种类密切相关,这直接影响了鸢乌贼的食物供应。饵料生物资源对鸢乌贼的生长繁殖起着至关重要的作用,其中饵料生物的可用性和种类之间的相互关系,对鸢乌贼的生物学特性有着显著影响,海洋环境通过影响饵料生物从而间接影响鸢乌贼的资源变化。因此海洋环境是鸢乌贼资源变化的一个重要解释因素,能解释其变化的38%。其余未解释部分可能是受到其他因素,如天敌捕食、人类捕捞等的影响。张梅[40]利用GAM模型分析表明,海表温度和海表盐度会对鸢乌贼CPUE产生显著性影响。周艳波等[19]研究发现通过分析各月份的海表高度和叶绿素a浓度适宜范围,可以有效表征鸢乌贼资源密度和渔场分布。温健等[24]认为基于海表温度和光合有效辐射的栖息地模型能够有效预测西北太平洋柔鱼的栖息地热点海域。这表明鸢乌贼资源变化与海洋环境变化具有非常紧密的联系,与本文结论相符。
综上所述,海洋环境是一个复杂的自然系统,描述海洋环境因子对鸢乌贼资源的影响时,不能仅从一对一的直接关系加以描述,更要考虑各变量间的相互关系,从整体进行分析,才能更科学地揭示海洋环境与鸢乌贼资源的响应机理。相对于传统的统计模型,结构方程模型在解决生态系统这些复杂的关系上具有更明显的优势,它在运算时可囊括所有变量进行分析,而非变量间的单独运算。还可以使用一些容易观测到的指标来估计那些不易测量到的潜变量,进而揭示各潜变量间的相互关系,以直接作用和间接作用来描述[41],为揭示海洋环境与鸢乌贼资源的响应关系提供了独特的整体视角。研究表明结构方程模型能够直观反映出海洋环境对鸢乌贼资源的影响。但由于数据的局限性,后续研究中应加入更多的观测指标以增强潜变量的解释力,加入更多的潜变量以揭示鸢乌贼资源变化的内在机理。
-
![]()
图 1 南海2019—2020年围网捕捞站点分布图
Figure 1. Sampling stations for seine fishing in the South China Sea during 2019−2020
![]()
图 2 南海海洋环境与鸢乌贼资源响应关系概念模型
Figure 2. Conceptual model of the relationship between marine environment and S. oualaniensis resources in South China Sea
![]()
图 3 2019—2020年围网平均单位捕捞努力量渔获量 (a) 和总平均产量、总平均单位捕捞努力量渔获量分布 (b)
Figure 3. Distribution of average CPUE (a), total average catch and total average CPUE (b) for seine during 2019−2020
![]()
图 4 2019—2020年春季鸢乌贼渔汛期单位捕捞努力量渔获量空间分布
Figure 4. Spatial distribution of spring CPUE for S. oualaniensis during 2019−2020
-
[1] 马克明, 傅伯杰, 黎晓亚, 等. 区域生态安全格局: 概念与理论基础[J]. 生态学报, 2004(4): 761-768. [2] 范江涛, 冯雪, 邱永松, 等. 南海鸢乌贼生物学研究进展[J]. 广东农业科学, 2013, 40(23): 122-128. [3] 冯波, 颜云榕, 张宇美, 等. 南海鸢乌贼 (Sthenoteuthis oualaniensis) 资源评估的新方法[J]. 渔业科学进展, 2014, 35(4): 1-6. [4] 陈新军, 叶旭昌. 印度洋西北部海域鸢乌贼渔场与海洋环境因子关系的初步分析[J]. 上海水产大学学报, 2005(1): 55-60. [5] 张俊, 邱永松, 陈作志, 等. 南海外海大洋性渔业资源调查评估进展[J]. 南方水产科学, 2018, 14(6): 118-127. [6] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. [7] 范江涛, 张俊, 冯雪, 等. 基于栖息地模型的南沙海域鸢乌贼渔情预报研究[J]. 南方水产科学, 2015, 11(5): 20-26. [8] 余景, 胡启伟, 李纯厚, 等. 西沙—中沙海域春季鸢乌贼资源与海洋环境的关系[J]. 海洋学报, 2017, 39(6): 62-73. [9] 袁帅, 付和平, 武晓东, 等. 基于结构方程模型分析荒漠啮齿动物优势种对不同放牧干扰的响应[J]. 生态学报, 2017, 37(14): 4795-4806. [10] MOUCHET M A, VILLÉGER S, MASON N W H, et al. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules[J]. Funct Ecol, 2010, 24(4): 867-876. doi: 10.1111/j.1365-2435.2010.01695.x
[11] 温纯, 金光泽. 功能多样性对典型阔叶红松林生产力的影响[J]. 植物生态学报, 2019, 43(2): 94-106. [12] 刘冉. 基于结构方程模型的浮游植物与环境因子关系的研究[D]. 北京: 华北电力大学, 2018: 4-9. [13] DUNCAN T E, DUNCAN S C. An introduction to latent growth curve modeling[J]. Behav Ther, 2004, 35(2): 333-363. doi: 10.1016/S0005-7894(04)80042-X
[14] 石亚飞, 石善恒, 黄晓敏. 基于R的结构方程模型在生态学中的应用[J]. 生态学杂志, 2022, 41(5): 1015-1023. [15] YANG Z, WANG X F, ZENG J W, et al. Effects of environmental factors on the spatio-temporal patterns of bigeyes (Priacanthus spp.) in the northern South China Sea[J]. Indian J Fish, 2021, 68(3): 27-37.
[16] 肖戈, 徐博, 张衡, 等. 阿拉伯海外海鸢乌贼渔场时空分布与海洋环境要素的研究[J]. 南方水产科学, 2022, 18(4): 10-19. [17] 陆化杰, 王洪浩, 何静茹, 等. 季风变化对西北印度洋鸢乌贼渔业生物学及渔场学影响的研究进展[J]. 中国水产科学, 2022, 29(11): 1669-1678. [18] 招春旭. 南海鸢乌贼渔场时空分布及其预报模型构建[D]. 湛江: 广东海洋大学, 2017: 30-32. [19] 周艳波, 谢恩阁, 吴洽儿, 等. 南海外海鸢乌贼渔场范围与海洋环境的关系[J]. 海洋学报, 2021, 43(2): 38-48. [20] CHEN X J, LIU B L, TIAN S Q, et al. Fishery biology of purpleback squid, Sthenoteuthis oualaniensis, in the northwest Indian Ocean[J]. Fish Res, 2007, 83(1): 98-104. doi: 10.1016/j.fishres.2006.09.005
[21] CHEN X J, ZHAO X H, CHEN Y. Influence of El Niño/La Niña on the western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean[J]. Ices J Mar Sci, 2007, 64(6): 1152-1160. doi: 10.1093/icesjms/fsm103
[22] MOHAMED K S, SAJIKUMAR K K, RAGESH N, et al. Relating abundance of purpleback flying squid Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) to environmental parameters using GIS and GAM in south-eastern Arabian Sea[J]. J Nat Hist, 2018, 52(29/30): 1869-1882.
[23] LIU K K, CHAO S Y, SHAW P T, et al. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study[J]. Deep-Sea Res I, 2002, 49(8): 1387-1412. doi: 10.1016/S0967-0637(02)00035-3
[24] 温健, 陆鑫一, 陈新军, 等. 基于海表温度和光合有效辐射的西北太平洋柔鱼冬春生群体栖息地热点预测[J]. 上海海洋大学学报, 2019, 28(3): 456-463. [25] YU W, CHEN X J, YI Q, et al. Variability of suitable habitat of western winter-spring cohort for neon flying squid in the Northwest Pacific under anomalous environments[J]. PLoS One, 2015, 10(4): e0122997. doi: 10.1371/journal.pone.0122997
[26] SANCHEZ P, DEMESTRE M, RECASENS L, et al. Combining GIS and GAMs to identify potential habitats of squid Loligo vulgaris in the Northwestern Mediterranean[J]. Hydrobiologia, 2008, 612(1): 91-98. doi: 10.1007/s10750-008-9487-9
[27] 温利红, 张衡, 方舟, 等. 印度洋北部鸢乌贼CPUE标准化初步研究[J]. 海洋湖沼通报, 2022, 44(4): 89-97. [28] SHEN H, DONG S K, LI S, et al. Grazing enhances plant photosynthetic capacity by altering soil nitrogen in alpine grasslands on the Qinghai-Tibetan plateau[J]. Agr Ecosyst Environ, 2019, 280: 161-168. doi: 10.1016/j.agee.2019.04.029
[29] 张宇美, 颜云榕, 卢伙胜, 等. 西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究[J]. 广东海洋大学学报, 2013, 33(3): 56-64. [30] 徐红云. 南海外海鸢乌贼栖息地分布与关键环境因子分析[D]. 上海: 上海海洋大学, 2017: 32-33. [31] 余为, 陈新军. 印度洋西北海域鸢乌贼9-10月栖息地适宜指数研究[J]. 广东海洋大学学报, 2012, 32(6): 74-80. [32] 范秀梅, 崔雪森, 唐峰华, 等. 基于PCA-GAM的阿拉伯海公海鸢乌贼资源量空间分布预测模型研究[J]. 水产学报, 2022, 46(12): 2340-2348. [33] 范江涛, 余为, 马胜伟, 等. 春季南海鸢乌贼栖息地时空分布及其年际差异分析[J]. 南方水产科学, 2022, 18(1): 1-9. [34] 颜云榕, 邱星宇, 张丽姿, 等. 南沙海域鸢乌贼繁殖生物学特性[J]. 广东海洋大学学报, 2021, 41(3): 20-27. [35] VIJAI D, SAKAI M, KAMEI Y, et al. Spawning pattern of the neon flying squid Ommastrephes bartramii (Cephalopoda: Oegopsida) around the Hawaiian Islands[J]. Sci Mar, 2014, 78(4): 511-519. doi: 10.3989/scimar.04112.27B
[36] NIGMATULLIN C M, MARKAIDA U. Oocyte development, fecundity and spawning strategy of large sized jumbo squid Dosidicus gigas (Oegopsida: Ommastrephinae)[J]. J Mar Biol Assoc UK, 2009, 89(4): 789-801. doi: 10.1017/S0025315408002853
[37] 范江涛, 张俊, 冯雪, 等. 基于地统计学的南沙海域鸢乌贼渔场分析[J]. 生态学杂志, 2017, 36(2): 442-446. [38] 骆雷. PLS-SEM多变量统计分析在赛事观众研究领域中的应用[J]. 上海体育学院学报, 2020, 44(11): 86-94. [39] 张沥今, 陆嘉琦, 魏夏琰, 等. 贝叶斯结构方程模型及其研究现状[J]. 心理科学进展, 2019, 27(11): 1812-1825. [40] 张梅. 南海外海鸢乌贼渔场时空分布与渔情预报模型研究[D]. 西安: 西安电子科技大学, 2021: 25-31. [41] JONSSON M, WARDLE D A. Structural equation modelling reveals plant-community drivers of carbon storage in boreal forest ecosystems[J]. Biol Lett, 2010, 6(1): 116-119. doi: 10.1098/rsbl.2009.0613
-
期刊类型引用(0)
其他类型引用(1)
 下载:
下载:
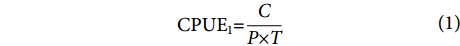
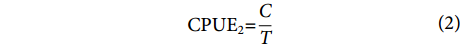
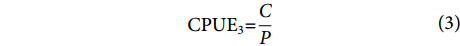

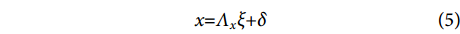
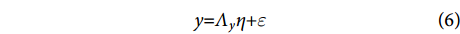
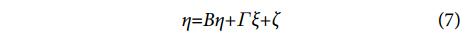


计量
- 文章访问数: 229
- HTML全文浏览量: 46
- PDF下载量: 57
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
