
Analysis of phytoplankton community in aquaculture crab pond using morphological and metagenomics methods
-
摘要:
为全面准确评估中华绒螯蟹 (Eriocheir sinensis) 养殖池塘中后期水体浮游植物群落结构特征及其与环境因子的关系,于2022年6—10月逐月监测水质指标,同时采用基于形态学和宏基因组学的人工镜检和高通量测序两种方法分析浮游植物的群落结构特征,探讨浮游植物优势种与环境因子的关系。结果显示,人工镜检鉴定的浮游植物共4门65种,而高通量测序数据分析所得浮游植物共4门1463种,其中人工镜检鉴定的优势种为蓝藻门的水华微囊藻 (Microcystis flos-aquae)、细小隐球藻 (Aphanocapsa elachista) 以及硅藻门的中肋骨条藻 (Skeletonema costatum),而高通量测序数据分析所得优势种为蓝藻门的水华束丝藻 (Aphanizomenon flos-aquae)、铜绿微囊藻 (M. aeruginosa)。人工镜检方法计算所得多样性指数普遍低于高通量测序结果。两种方法共同分析得出溶解氧(DO)、叶绿素 a (Chl-a)、总氮 (TN) 和氨氮 (NH3-N) 是影响浮游植物群落结构的关键环境因子。建议采用人工镜检与高通量测序法相结合的方式分析养殖期间的浮游植物群落结构,以便更全面、准确地评估养殖生态系统中的浮游植物群落结构特征及其与环境因子的关系。
Abstract:In order to fully and accurately evaluate the characteristics of phytoplankton community structure and its relationship with environmental factors in the middle and late periods of Chinese mitten crab (Eriocheir sinensis) pond culture, we had monitored the water quality indexes and phytoplankton community of aquaculture crab ponds from June to October 2022, and identified the phytoplankton community by artificial microscopy and high-throughput sequencing based on morphological and metagenomics methods. Then we discussed the relationship between dominant phytoplankton species and environmental factors. The results show that 65 species in four phyla were identified by artificial microscopy, and 1 463 species by metagenomic data analysis, in which the dominant species identified by artificial microscopy were Microcystis flos-aquae, Aphanocapsa elachista and Skeletonema costatum, and the dominant species identified by metagenomic data analysis were Aphanizomenon flos-aquae and M. aeruginosa. The diversity index calculated by manual microscopy was generally lower than that of metagenomic results. Dissolved oxygen, chlorophyll a, total nitrogen and ammonia nitrogen were the key environmental factors affecting phytoplankton community structure. It is suggested to simutaneously use artificial microscopy and high-throughput sequencing methods to analyze phytoplankton community structure in aquacultural water, so as to evaluate the characteristics of phytoplankton community structure and its relationship with environmental factors in aquaculture ecosystems more comprehensively and accurately.
-
方斑东风螺 (Babylonia areolata)属软体动物门、腹足纲、蛾螺科,俗称“花螺”、“海猪螺”和“南风螺”,分布于中国东南沿海、东南亚及日本等地,是中国东南沿海主要的养殖海珍品和经济贝类,因其肉质鲜美、营养丰富,成为畅销国内外且具有推广前景的优质海水养殖贝类品种[1]。近年来,方斑东风螺的养殖生产在中国部分地区如广东、广西和海南等有较快发展,但在养殖过程中常受到氨氮 (NH4-N)毒性的影响。积累的氨以离子氨 (
$ {\rm{NH}}_4^ + $ )或非离子氨 (NH3)的形式存在于水体,从而影响水生动物的生长[2]。NH4-N是水产养殖水环境中的主要有毒物质和重要的水体环境指标之一,被认为是水产养殖中的常见胁迫因子,其浓度受养殖水体中动物排泄物、分泌物、残饵、动植物尸体等含氮有机物分解的影响[3-4]。研究表明,多数水生生物对NH4-N毒性非常敏感。关于NH4-N对鱼类、虾类等水生生物消化系统和抗氧化系统酶活力影响的研究已有不少报道[5-8],但NH4-N胁迫对方斑东风螺消化酶活力影响的研究仍属空白。徐武杰等[9]研究NH4-N胁迫对三疣梭子蟹 (Portunus trituberculatus) 消化酶活力的影响时发现,在NH4-N胁迫下其中肠腺中的胃蛋白酶和脂肪酶活力表现出明显的诱导作用,而对3种消化器官中淀粉酶活力表现出明显的抑制作用;胡炜等[10]研究发现慢性NH4-N胁迫会对刺参 (Stichopus japonicus) 摄食、消化与生长产生不利影响,在质量浓度低于4 mg·L–1的NH4-N胁迫下,刺参消化酶活力短期可被显著诱导上调,高NH4-N质量浓度对消化酶活力起抑制效应并可导致生理紊乱,且该响应存在体质量、规格上的差异;王程昊等[11]研究发现,NH4-N胁迫也会影响泥蚶 (Arca granosa)体内的几种免疫酶活力。NH4-N还可降低水生动物生长速度,影响其血液生化指标并伤害免疫功能、组织机构及繁殖能力[12-13]。当水体中NH4-N质量浓度低于生物体耐受限度时,机体能够自行调节其体内相关酶的活力从而适应外界环境的变化,但当一定质量浓度的NH4-N持续长时间刺激且超过机体调节限度时,机体的非特异性免疫系统及消化系统就会受到破坏,相关酶活力下降[14-16]。另外,NH4-N胁迫还会增加水生动物的致病性[17],从而影响水生动物健康。为探究方斑东风螺在NH4-N刺激下其体内消化酶活力变化,进而揭示其消化酶应对急性胁迫时的调节机制,本文通过开展方斑东风螺不同质量浓度的NH4-N胁迫实验,测定NH4-N对其常见消化酶活力的影响,以期充实NH4-N对贝类毒理实验消化酶层面的基础数据,并为其他贝类NH4-N胁迫实验提供科学依据。
1. 材料与方法
1.1 实验材料
方斑东风螺由中国水产科学研究院南海水产研究所热带水产研究开发中心 (海南陵水)提供,并在该基地进行实验。实验所用方斑东风螺平均体质量为 (0.392±0.080) g,体长为(0.65±0.15) cm,实验开始前先暂养2 d。养殖过程中水质参数为温度 (26.0±1.0) ℃,盐度33±0.8,溶解氧 (DO)质量浓度大于6.5 mg·L–1,亚硝酸盐质量浓度小于0.04 mg·L–1,NH4-N质量浓度小于0.01 mg·L–1,pH为8.0±0.2。实验用水为过滤的海水。
1.2 实验方法
1.2.1 饲养管理及实验设计
将氯化铵 (NH4Cl,AR)配置成质量浓度为0~500 mg·L–1的溶液,按实验要求放入健康状况良好且大小均一的方斑东风螺,观察其行为、活动及存活状况,得到24 h和 96 h 100%死亡浓度 [ (LC100, 24 h)和 (LC100, 96 h)],根据实验结果确定出实验液NH4-N质量浓度的上、下限。实验过程中停止投饵。
根据所确定的上、下限设置5个NH4-N质量浓度,分别为0 mg·L–1、22 mg·L–1、47 mg·L–1、102 mg·L–1和220 mg·L–1,各组均为3个平行,每个实验容器 (3 L)内投放30只健康状况良好的方斑东风螺,分别于第6、第12、第24、第36、第48、第72和第96小时从各养殖桶中随机取样3只,检测其消化酶活力的变化情况。同时,每隔3 h从每个容器中取水样,及时调整至设定的表观质量浓度。
每日换水1次,每次换水50%,并将养殖桶中的排泄物以及死去的方斑东风螺及时除去。间隔观察并记录各实验组方斑东风螺的状态、行为活动、中毒症状及死亡率。判断死亡的标准为实验东风螺置于塑料板上无明显活动迹象,用解剖刀触碰无反应。
1.2.2 样品处理
用0.2 mol·L–1生理盐水将各实验组所取样品 (整个螺)按1∶2 (m∶V)进行研磨,研磨液于5 000 r·min–1、4 ℃下离心10 min,取1 mL上清液于洁净EP (eppendorf)管中,−80 ℃保存待测,分别采用相关试剂盒测定蛋白及各消化酶活力,试剂盒购自南京建成生物工程研究所。
1.2.3 数据处理与分析
利用SPSS 21.0对实验数据进行统计分析,先对数据作单因素方差分析 (One-Way ANOVA),处理间若有显著差异,再用Duncan法比较均值间的差异显著性 (P<0.05),本文数据均采用“平均值±标准差 (
$\overline X \pm {\rm SD}$ )”表示。2. 结果
2.1 NH4-N胁迫对方斑东风螺的急性毒性作用
NH4Cl处理组中,随着NH4Cl浓度的升高,方斑东风螺呈现不同的应激行为甚至死亡,主要表现为运动缓慢,对外界刺激反应迟钝,爬壁运动减少,逐渐翻背,沉于水桶底部,身体僵硬直至死亡。死亡状态的东风螺,吻管向外凸出,螺肉外翻、惨白僵硬。300 mg∙L–1 NH4Cl处理组第48小时开始出现死亡,第96小时成活率为 (60.0±1.6)%,而500 mg·L–1 NH4-N处理组于第24 小时便出现 (20.3±2.1)%的死亡率,死亡个体吻稍微张开、出现体色变红现象,且第96小时成活率为0,其余各组成活率为100% (表1)。可见,NH4-N浓度越大,其毒性作用越强,方斑东风螺死亡率越高;相同条件下,NH4-N胁迫时间越久,方斑东风螺死亡率也越高。
表 1 氨氮对方斑东风螺急性毒性实验结果Table 1. Acute toxicity experiment of NH4-N to B. areolataρ(NH4Cl)/mg·L–1 死亡率/% mortality rate 第24小时 第48小时 第72小时 第96小时 对照 control 0 0 0 0 25 0 0 0 0 50 0 0 0 0 75 0 0 0 0 100 0 0 0 0 200 0 0 0 0 300 0 21.4±3.3 40.1±2.9 60.0±1.6 500 20.3±2.1 50.0±2.7 73.6±4.2 100.0 2.2 NH4-N胁迫对方斑东风螺溶菌酶活力的影响
不同浓度NH4-N对方斑东风螺溶菌酶活力的影响显著 (P<0.05),而处理时间对方斑东风螺溶菌酶活力影响极显著 (P<0.01)。第6小时时各NH4-N浓度处理组个体溶菌酶活力与对照组相比均呈降低趋势,但随着时间的延长,溶菌酶活力呈先增后降 (22 mg·L–1 和102 mg·L–1处理组)或先增后降再增 (47 mg·L–1和220 mg·L–1处理组)的趋势 (图1-a)。综上,溶菌酶活力表现出“抑制−诱导”的趋势。
![]() 图 1 氨氮胁迫对方斑东风螺4种消化酶活力的影响不同字母表示同一组不同处理时间之间存在显著性差异,字母下标数字代表组号,分别为22 mg∙L–1、47 mg∙L–1、102 mg∙L–1和220 mg∙L–1Figure 1. Effect of NH4-N stress on activities of four digestive enzymes of B. areolataDifferent letters indicate significant difference. The subscript letters represent group No., which are 122 mg∙L–1, 47 mg∙L–1, 102 mg∙L–1 and 220 mg∙L–1, respectively.
图 1 氨氮胁迫对方斑东风螺4种消化酶活力的影响不同字母表示同一组不同处理时间之间存在显著性差异,字母下标数字代表组号,分别为22 mg∙L–1、47 mg∙L–1、102 mg∙L–1和220 mg∙L–1Figure 1. Effect of NH4-N stress on activities of four digestive enzymes of B. areolataDifferent letters indicate significant difference. The subscript letters represent group No., which are 122 mg∙L–1, 47 mg∙L–1, 102 mg∙L–1 and 220 mg∙L–1, respectively.2.3 NH4-N胁迫对方斑东风螺胃蛋白酶活力的影响
处理时间和不同质量浓度的NH4-N均极显著影响方斑东风螺胃蛋白酶活力 (P<0.01)。相比对照组,22 mg·L–1处理组个体胃蛋白酶活力随时间的延长呈“诱导−抑制”变化趋势,但47 mg·L–1处理组个体胃蛋白酶活力随时间的延长却呈现“抑制−诱导−抑制”变化趋势,而其他各处理组个体胃蛋白酶活力随时间的变化趋势为“抑制−诱导”且峰值均出现在第72 小时,谷值均出现在第6小时 (图1-b)。
2.4 NH4-N胁迫对方斑东风螺脂肪酶活力的影响
处理时间对方斑东风螺脂肪酶活力影响极显著 (P<0.01),不同浓度的NH4-N胁迫导致各组处理个体脂肪酶活力变化趋势也存在显著差异 (P<0.05)。各处理组脂肪酶活力较对照组在总体上呈抑制作用或与对照组呈相似水平,但中质量浓度处理组 (47 mg·L–1、102 mg·L–1)个体脂肪酶活力在第6小时时表现出诱导作用,低、高质量浓度处理组 (22 mg·L–1、220 mg·L–1)个体脂肪酶活力于第48小时时表现出诱导作用 (图1-c)。
2.5 NH4-N胁迫对方斑东风螺淀粉酶活力的影响
处理时间对方斑东风螺淀粉酶活力影响极显著 (P<0.01),不同浓度的NH4-N胁迫导致各组处理个体淀粉酶活力变化趋势存在明显差异 (P<0.05)。各处理组淀粉酶活力相比对照组在处理时间较短的情况下并无太大的诱导或抑制作用,NH4-N质量浓度为47 mg·L–1和102 mg·L–1的处理组在第72和第96小时才表现出明显的诱导作用,而NH4-N质量浓度为220 mg·L–1的处理组在第6小时表现出明显的抑制作用,并在第36和第96小时时具有一定的诱导作用 (图1-d)。
3. 讨论
3.1 NH4-N对方斑东风螺生存状况和成活率的影响
研究表明,对水生动物造成危害甚至死亡的NH4-N成分主要是NH3[2]。NH3因不带电荷且具有较强的脂溶性而对水生动物有极大毒性,能够穿透贝体细胞膜表现出毒性效应,但
${\rm{NH}}_4^ + $ 对水生动物毒性很小[7,11,18]。本实验结果显示,方斑东风螺对NH4-N具有一定的耐受性,因此高质量浓度 (500 mg∙L–1) NH4-N处理24 h仅 (20.3±2.1)%的死亡率,但在96 h后全部死亡,说明该NH4-N浓度已超过方斑东风螺最大耐受浓度。师尚丽等[2]指出,NH4-N的浓度越大,其毒性作用越强,与本实验结果一致。因此,养殖生产中应尽可能地降低养殖水体中NH4-N的浓度,及时清理投喂的冰鲜鱼、虾蟹肉等残渣及死亡的东风螺等,防止含氮有机物分解产生NH4-N。另外,方斑东风螺对NH4-N的应激行为表现为运动缓慢、对外界刺激反应迟钝和爬壁运动减少等,可能是NH4-N影响了东风螺的组织器官,如影响肌肉伸缩能力和神经介质传递。研究表明,水体中的NH4-N会影响鱼、贝类等水产动物的生长摄食、组织器官和免疫机能等 [12,19-23]。洪美玲等[24]在研究NH4-N对中华绒螯蟹 (Eriocheir sinensis)幼体的免疫指标及肝胰腺组织结构影响时指出,NH4-N不仅损伤机体非特异性免疫防御系统,还对机体细胞和组织造成伤害。王琨[25]研究发现,在NH4-N的影响下鲤 (Cyprinus carpio)幼鱼的各个组织器官 (鳃、肠、心肌、肾脏、脾脏和肝脏)均受到不同程度的损伤,且高浓度的NH4-N使鲤幼鱼的红细胞数量和血红蛋白浓度下降,而组织器官的损伤和免疫机能的下降正是造成鲤容易患病、死亡的主要原因。
3.2 NH4-N对方斑东风螺个体消化酶活力的影响
大量研究表明,NH4-N作为养殖水质污染源之一,除了对鱼、贝类等水产动物机体的血液指标、组织结构和免疫机能造成损害,还能降低鱼虾贝类的产卵能力[26],引起血淋巴理化因子和抗病能力的变化[27-28],使血淋巴中血细胞 (如透明细胞、颗粒细胞和半颗粒细胞)数量下降[7],还会对水生生物消化系统酶活力造成影响[5]。本研究结果表明,不同质量浓度的NH4-N对消化酶活力均产生影响,且随着处理时间的延长表现出不同的诱导或抑制作用。NH4-N对方斑东风螺个体溶菌酶活力表现出“抑制−诱导”的趋势,说明贝类溶菌酶活力的提高可能也是一种防御反应外的被动病理显示[29]。相比对照组,22 mg·L–1处理组个体胃蛋白酶活力随时间的延长呈“诱导−抑制”变化趋势,但47 mg·L–1处理组个体胃蛋白酶活力随时间的延长呈“抑制−诱导−抑制”变化趋势;而其他各处理组个体胃蛋白酶活力均在NH4-N处理一段时间后表现出抑制作用。可以看出在NH4-N急性胁迫下,由于NH4-N的毒性作用,使得机体的相关代谢酶活力降低,而机体的应激反应又在短时间内诱导酶活力上升,其恢复正常水平的时间反映了机体对环境的适应能力。最终酶活力的显著降低说明对处理个体已经造成毒害作用。而不同NH4-N质量浓度对方斑东风螺处理个体脂肪酶活力的总体抑制,更表明了NH4-N的毒害作用。赵海涛[30]通过南方鲶 (Silurus meridionalis)幼鱼的NH4-N胁迫实验得出,经NH4-N胁迫后南方鲶幼鱼的造血机能受损,机体无法将氧 (O2)顺利携带进入,猜测NH4-N急性中毒引起死亡的主要原因是由机体组织缺氧造成。而乔顺风等[31-32]认为,NH4-N的毒性大小取决于存在状态,当水体中的NH4-N以NH3状态存在时,便会对水生动物产生很强的神经性毒害,造成急性氨中毒。李波等[33]在研究亚硝酸 (HNO2)和NH4-N对黄颡鱼 (Pelteobagrus fulvidraco)的急性毒性时发现,NH4-N对黄颡鱼的毒性明显大于HNO2对黄颡鱼的毒性,因而把NH4-N作为黄颡鱼养殖过程中毒性危害的重要影响因子。
此外,虽然各消化酶活力随着NH4-N处理浓度显著变化,但并不存在明显的线性关系或一致规律,变化规律较特异,这可能与东风螺的高抗逆性等生理习性或所处发育阶段不同有关,也可能是NH4-N的胁迫对其消化系统造成了一定的紊乱效应,但并未引起死亡是因为胁迫处理计量仍在其适应范围内。有研究表明,NH4-N胁迫程度的差异性与对象规格、养殖密度和个体差异等有关[34]。NH4-N处理组方斑东风螺消化酶活力的变化规律很好地指引了养殖过程中水体NH4-N的控制范围,从而防止因超过最大承受能力而造成重大养殖损失。
-
![]()
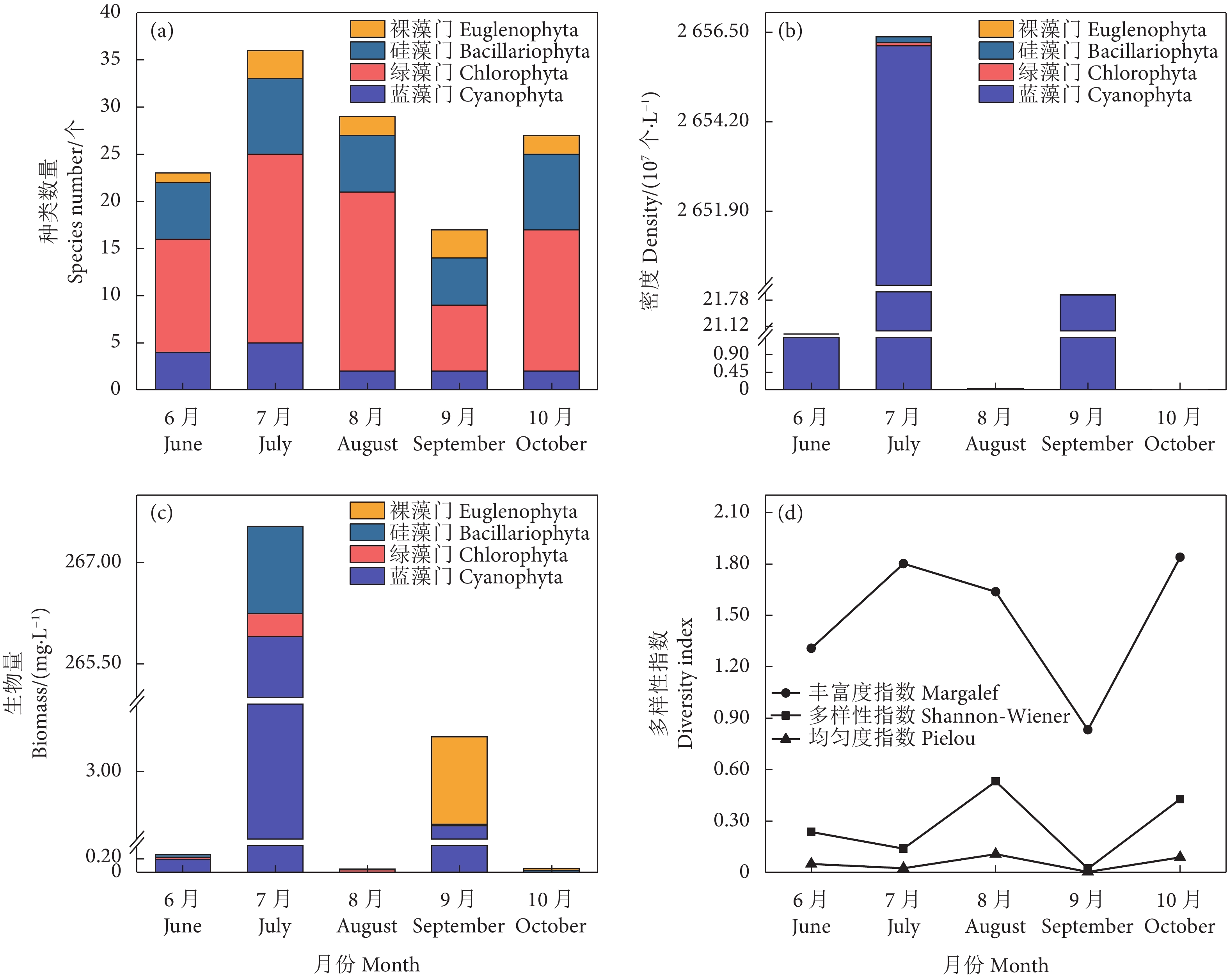
图 1 不同月份的浮游植物种类数量、密度、生物量和多样性指数
Figure 1. Species composition, density, biomass and diversity index of plankton in different months
![]()
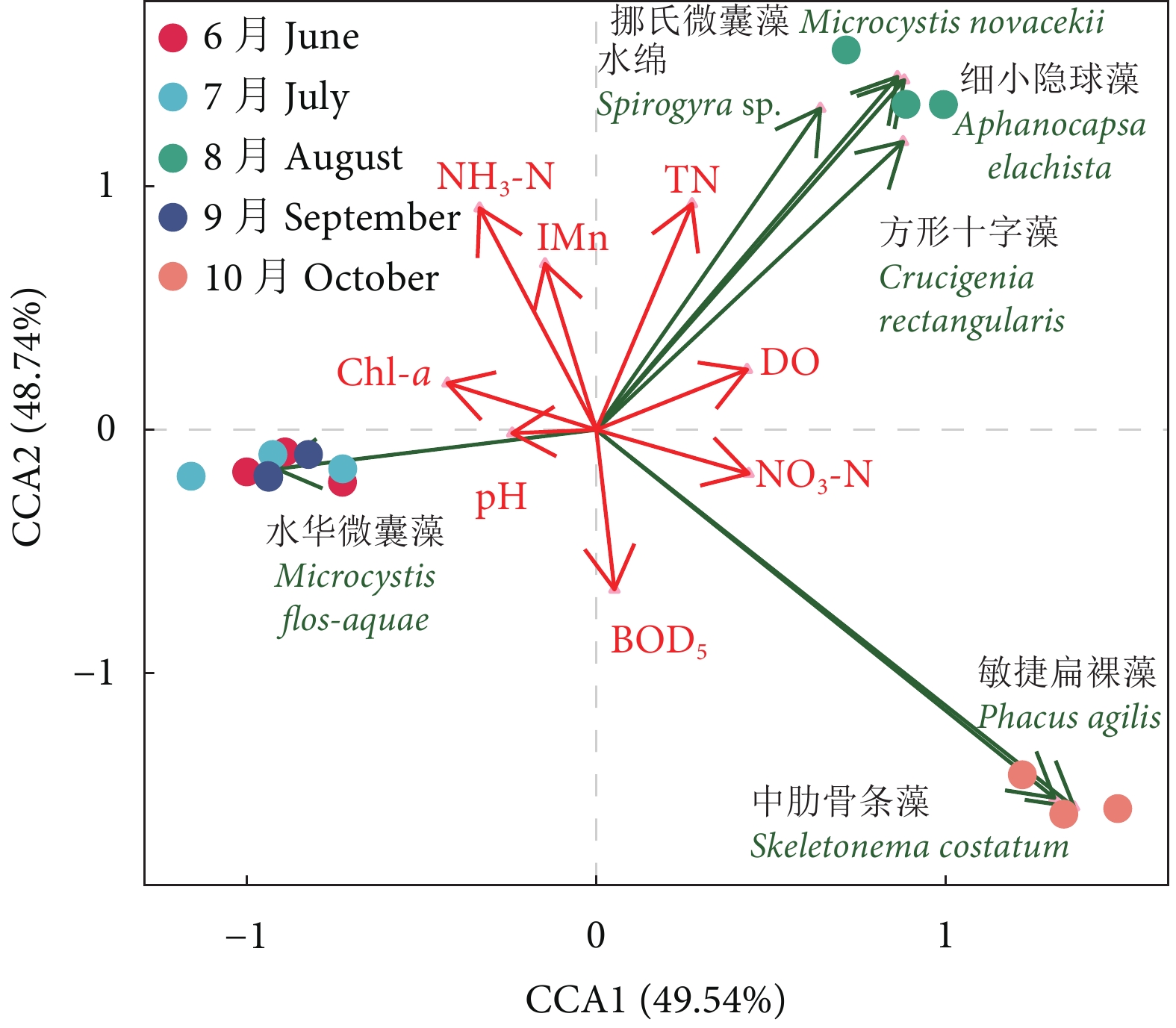
图 2 养殖蟹塘浮游植物优势种与环境因子的CCA排序分析
Figure 2. CCA ordination of phytoplankton dominant species and environmental factors in aquaculture pond of E. sinensis
![]()
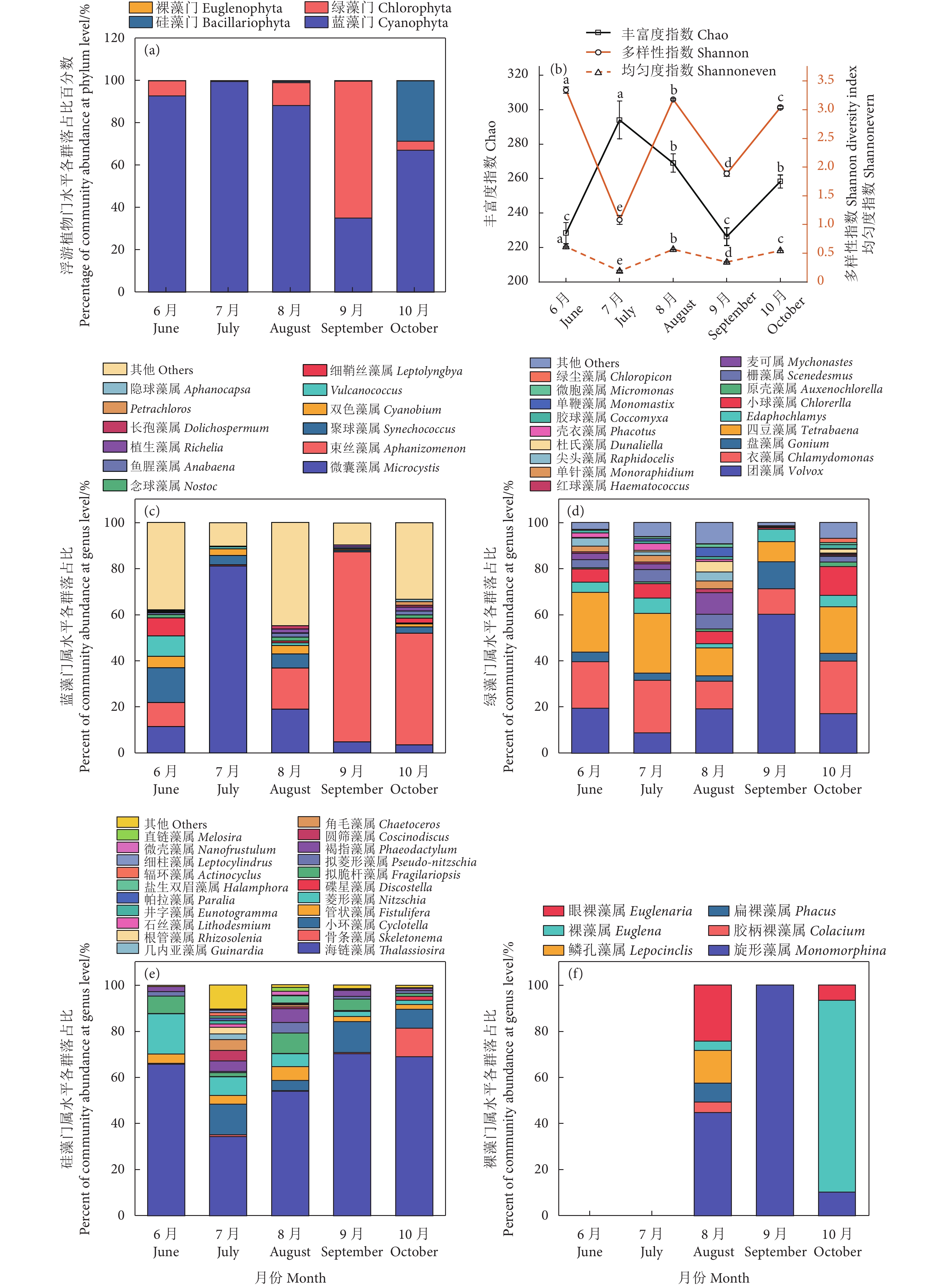
图 3 浮游植物门水平各群落组成(a)、多样性指数(b)、蓝藻门、绿藻门、硅藻门和裸藻门属水平各群落组成(c—f)
Figure 3. Percent ages of community abundance at phylum level (a) and diversity indices of plankton (b), percent of community abundance at genus level of Cyanophyta, Chlorophyta, Bacillariophyta, and Euglenophyta (c–f) in different months
![]()
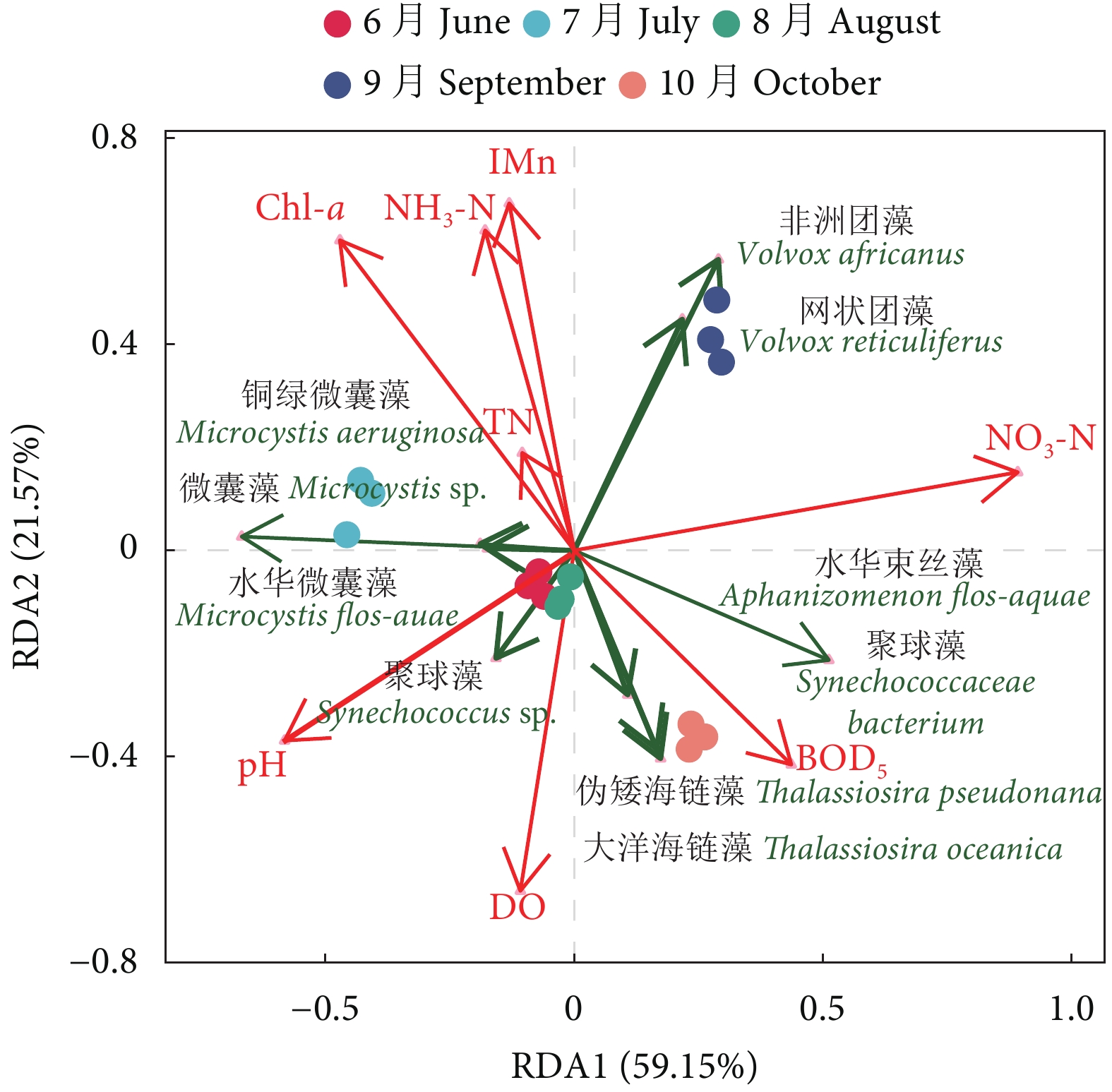
图 4 养殖蟹塘浮游植物优势种与环境因子的RDA排序分析
Figure 4. RDA ordination of phytoplankton dominant species and environmental factors in aquaculture pond of E. sinensis
表 1 中华绒螯蟹养殖池塘水体理化指标
Table 1 Physicochemical parameters in water bodies of E. sinensis aquaculture ponds
指标
Index6月
June7月
July8月
August9月
September10月
OctoberpH 9.97±0.06a 9.15±0.25b 8.65±0.44b 7.92±0.39c 8.62±0.13b 水温 Water temperature/℃ 33.75±0.43a 33.78±1.27a 29.63±0.04b 22.00±0.02c 20.04±0.54d 溶解氧 DO/(mg·L−1) 11.88±1.61a 6.61±1.00bc 11.07±1.45a 5.23±0.95c 9.37±2.62ab 叶绿素 a Chl-a/(μg·L−1) 1.33±0.58d 50.67±2.08a 20.67±8.08c 33.33±6.11b 7.00±1.00d 总磷 TP/(mg·L−1) 0.11±0.01d 0.23±0.02c 0.34±0.01b 0.43±0.04a 0.09±0.01d 磷酸盐 PO4/(mg·L−1) 0.08±0.01c 0.16±0.03b 0.30±0.02a 0.28±0.02a 0.03±0.00d 总氮 TN/(mg·L−1) 0.89±0.03c 1.14±0.09b 2.41±0.24a 1.25±0.10b 0.78±0.02c 氨氮 NH3-N/(mg·L−1) 0.65±0.01c 0.55±0.04d 0.88±0.07a 0.75±0.04b 0.21±0.01e 亚硝态氮 NO2-N/(mg·L−1) 0.01±0.00b 0.01±0.00b 0.05±0.00a 0.01±0.00b 0.01±0.00b 硝态氮 NO3-N/(mg·L−1) 0.02±0.01d 0.01±0.01d 0.23±0.02c 0.46±0.04a 0.37±0.04b 高锰酸钾指数 IMn/(mg·L−1) 5.59±0.07b 7.43±0.20a 8.04±0.39a 8.06±1.20a 5.53±0.15b 生物需氧量 BOD5/(mg·L−1) 5.20±0.00a 3.50±0.10c 3.57±0.06c 4.07±0.25b 5.00±0.53a 注:同行数据不同上标字母表示组间存在显著性差异 (P<0.05);n=3。 Note: Values with different superscripts within each row are significantly different (P<0.05); n=3.  下载: 导出CSV
下载: 导出CSV
表 2 中华绒螯蟹养殖池塘浮游植物优势种优势度
Table 2 Dominance index of phytoplankton dominant species in aquaculture pond of E. sinensis
浮游植物
Phytoplankton优势种
Dominant species6月
June7月
July8月
August9月
September10月
October蓝藻门 Cyanophyta 水华微囊藻 Microcystis flos-aquae 0.98 1.00 — 1.00 — 细小隐球藻 Aphanocapsa elachista — — 0.82 — — 挪氏微囊藻 Microcystis novacekii — — 0.04 — — 绿藻门 Chlorophyta 水绵 Spirogyra sp. — — 0.04 — — 方形十字藻 Crucigenia rectangularis — — 0.04 — — 硅藻门 Bacillariophyta 中肋骨条藻 Skeletonema costatum — — — — 0.88 裸藻门 Euglenophyta 敏捷扁裸藻 Phacus agilis — — — — 0.06 注:数字表示物种优势度指数;“—”表示物种未达到优势度0.02。 Note: The numbers represent dominance index of the species; "—" represents the species has not reached the dominance standard of 0.02.
下载: 导出CSV
表 3 中华绒螯蟹养殖池塘浮游植物优势种占比
Table 3 Composition of phytoplankton dominant species in aquaculture pond of E. sinensis
浮游植物
Phytoplankton优势种
Dominant species占比前三的种
Top three species/%6月
June7月
July8月
August9月
September10月
October蓝藻门 Cyanophyta 水华束丝藻 Aphanizomenon flos-aquae 9.63 15.73 28.86 32.52 聚球藻 Synechococcus sp. 8.06 铜绿微囊藻 Microcystis aeruginosa 6.79 52.26 10.52 微囊藻 Microcystis sp. 4.00 水华微囊藻 Microcystis flos-aquae 3.70 聚球藻 Synechococcaceae bacterium 7.63 绿藻门 Chlorophyta 非洲团藻 Volvox africanus 15.84 网状团藻 Volvox reticuliferus 12.79 硅藻门 Bacillariophyta 大洋海链藻 Thalassiosira oceanica 9.82 伪矮海链藻 Thalassiosira pseudonana 9.75
下载: 导出CSV
-
[1] 周永昌, 姜晓东, 龙晓文, 等. 中草药对中华绒螯蟹幼蟹生长及免疫力的影响[J]. 水产科学, 2019, 38(2): 188-193. [2] KATMOKO G M D, RISJANI Y, MASITHAH E D. Analysis of phytoplankton structure community, water quality and cultivation performance in Litopenaeus vannamei intensive pond located in Tembokrejo Village, Muncar, Banyuwangi[J]. J Exp Life Sci, 2021, 11(3): 68-76. doi: 10.21776/ub.jels.2021.011.03.01
[3] 孙军, 宋煜尧, 刘海娇. 海洋浮游植物快速鉴定与监测技术[J]. 海洋科学进展, 2022, 40(4): 701-716. [4] 侯德昌, 张莹莹, 魏文志. 不同中华鳖养殖模式浮游植物功能群特征及水环境评价[J]. 安徽农业科学, 2022, 50(9): 96-99, 170. doi: 10.3969/j.issn.0517-6611.2022.09.023 [5] MERCURIO A L, QUERIJERO B, CHING J. Phytoplankton community in aquaculture and non-aquaculture sites of Taal Lake, Batangas, Philippines[J]. J Exp Biol Agric Sci, 2016, 4(1): 66-73. doi: 10.18006/2016.4(1).66.73
[6] 蒲炜佳, 董世鹏, 张东旭, 等. 三疣梭子蟹池塘综合养殖系统浮游植物群落结构及其与环境因子的关系[J]. 中国水产科学, 2022, 29(4): 549-561. [7] MANOYLOV K M. Taxonomic identification of algae (morphological and molecular): species concepts, methodologies, and their implications for ecological bioassessment[J]. J Phycol, 2014, 50(3): 409-424. doi: 10.1111/jpy.12183
[8] 方圆, 黎慧, 王李宝, 等. 凡纳滨对虾小型温棚养殖模式水体微生物群落的研究[J]. 南方水产科学, 2023, 19(3): 29-41. [9] 陈琼, 李贵阳, 罗坤, 等. 凡纳滨对虾 (Litopenaeus vannamei) 亲虾繁殖期水体微生物多样性[J]. 海洋与湖沼, 2017, 48(1): 130-138. [10] 高权新, 李云莉, 齐占会, 等. 基于宏基因组学的中国沿海密集养殖水域秋季底质细菌多样性研究[J]. 海洋渔业, 2019, 41(3): 354-363. [11] 卫鹏, 毕相东, 戴伟, 等. 淡水养殖池塘微型和超微型浮游植物的群落结构组成[J]. 大连海洋大学学报, 2022, 37(1): 113-119. [12] 乔玲, 常志强, 李健, 等. 基于形态学和高通量测序的海水池塘生态养殖系统中浮游植物多样性比较[J]. 渔业科学进展, 2022, 43(2): 32-43. [13] 国家环境保护总局水和废水监测分析方法编委会. 水和废水监测分析方法[M]. 北京: 中国环境出版社集团, 2013: 88-284. [14] 赵文. 水生生物学[M]. 北京: 中国农业出版社, 2015: 16-126, 493-498. [15] FU R B, LI Z P, ZHOU R, et al. The mechanism of intestinal flora dysregulation mediated by intestinal bacterial biofilm to induce constipation [J]. Bioengineered, 2021, 12(1): 6484-6498.
[16] STALDER T, BARRAUD O, JOVÉ T, et al. Quantitative and qualitative impact of hospital effluent on dissemination of the integron pool[J]. ISME J, 2014, 8(4): 768-777. doi: 10.1038/ismej.2013.189
[17] LIU J F, LAI L Y, LIN J J, et al. Ranitidine and finasteride inhibit the synthesis and release of trimethylamine N-oxide and mitigates its cardiovascular and renal damage through modulating gut microbiota[J]. Int J Biol Sci, 2020, 16(5): 790-802. doi: 10.7150/ijbs.40934
[18] 刘淑娟. 中肋骨条藻 (Skeletonema costatum) 赤潮生消规律及其对浮游生物群落结构的影响[D]. 舟山: 浙江海洋大学, 2019: 3-6. [19] 赵艳民, 秦延文, 张雷, 等. 基于GIS的近30年长江口及其邻近海域赤潮时空分布特征研究[J]. 海洋科学, 2021, 45(12): 39-46. [20] 胡婧文, 弋钰昕, 尤庆敏, 等. 骨条藻属硅藻的分类、生态及生理研究进展[J]. 上海师范大学学报(自然科学版), 2020, 49(6): 698-708. [21] 韩志男. 崇明岛海水入侵特征及趋势分析[D]. 青岛: 国家海洋局第一海洋研究所, 2013: 1-6. [22] 董立新, 周绪申. 浮游植物多样性指数在内陆水体污染类型评价中的应用简述[J]. 海河水利, 2017(5): 57-60. [23] 龚进玲, 李学梅, 何勇凤, 等. 青鱼—鲢、鳙生态池塘浮游生物群落结构特征及其与环境因子的关系[J]. 水产科学, 2022, 41(4): 517-526. [24] 孟得微, 孙博, 张立民, 等. 东北盐碱池塘浮游植物群落结构特征与环境因子的关系[J]. 安徽农业科学, 2021, 49(21): 110-115. [25] 温旭, 马旭洲, 范伟, 等. 不同面积芦苇稻幼蟹塘浮游植物功能类群的结构特征[J]. 浙江大学学报(农业与生命科学版), 2019, 45(1): 85-94. [26] FAN L M, HU G D, QIU L P, et al. Variations in bacterioplankton communities in aquaculture ponds and the influencing factors during the peak period of culture[J]. Environ Pollut, 2020, 258: 113656. doi: 10.1016/j.envpol.2019.113656
[27] 夏德军, 隋燚, 王云平, 等. 稻田推水增氧养殖小龙虾新技术[J]. 中国水产, 2019(1): 88-90. [28] NONG X Z, SHAO D G, SHANG Y M, et al. Analysis of spatio-temporal variation in phytoplankton and its relationship with water quality parameters in the South-to-North Water Diversion Project of China[J]. Environ Monit Assess, 2021, 193(9): 593. doi: 10.1007/s10661-021-09391-6
[29] JIANG Y J, HE W, LIU W X, et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu)[J]. Ecol Indic, 2014, 40: 58-67. doi: 10.1016/j.ecolind.2014.01.006
[30] 刘乾甫, 赖子尼, 高原, 等. 珠江三角洲地区精养淡水鱼塘浮游植物功能群特征[J]. 中国水产科学, 2018, 25(1): 124-136. [31] SIPAOBA-TAVARES L H, DIAS S G. Water quality and communities associated with macrophytes in a shallow water-supply reservoir on an aquaculture farm[J]. Braz J Biol, 2014, 74(2): 420-428. doi: 10.1590/1519-6984.27212
[32] 俞焰, 刘德富, 杨正健, 等. 千岛湖溶解氧与浮游植物垂向分层特征及其影响因素[J]. 环境科学, 2017, 38(4): 1393-1402. [33] 田盼, 宋林旭, 纪道斌, 等. 三峡库区神农溪不同时期溶解氧与叶绿素a垂向分布特征[J]. 水生态学杂志, 2022, 43(2): 1-8. [34] TAN X, GU H H, RUAN Y N, et al. Effects of nitrogen on interspecific competition between two cell-size cyanobacteria: Microcystis aeruginosa and Synechococcus sp.[J]. Harmful Algae, 2019, 89: 101661. doi: 10.1016/j.hal.2019.101661
-
期刊类型引用(10)
1. 孙阿君,丁炜东,曹丽萍,曹哲明,邴旭文. 氨氮胁迫对翘嘴鳜幼鱼抗氧化酶、消化酶活性及应激相关基因表达的影响. 水产科技情报. 2024(01): 44-51 .  百度学术
百度学术
2. 王筱,冼健安,张秀霞,张泽龙,李军涛,郑佩华,吴恒梅,鲁耀鹏. 方斑东风螺人工养殖、环境生理和营养需求研究进展. 中国饲料. 2024(03): 83-88+117 . 百度学术
3. 张钰伟,赵旺,邓正华,黄星美,温为庚,孙敬锋,王瑞旋. 芽孢杆菌对方斑东风螺生长、存活、免疫及消化酶活性的影响. 南方水产科学. 2023(01): 106-115 . 本站查看
4. 梁晶,邢诒炫,吕布,臧战,刘子岭,唐贤明,於锋,Hebert Ely Vasquez,战欣,郑兴,顾志峰. 底砂粒径对方斑东风螺生长、消化、抗氧化及底质的影响. 水产科技情报. 2023(03): 137-145 . 百度学术
5. 梅泊承,张晓东,赵淳朴,徐继林,王丹丽,郭春阳. 串联养殖模式下的凡纳滨对虾(Litopenaeus vannamei)与缢蛏(Sinonovacula constricta)生长特性、消化免疫及水生态效应. 海洋与湖沼. 2023(03): 907-920 . 百度学术
6. 赵旺,温为庚,谭春明,黄星美,杨蕊,陈明强,杨其彬,陈旭. 饥饿胁迫对猛虾蛄溶菌酶和消化酶活性的影响. 水产科技情报. 2022(01): 30-35 . 百度学术
7. 谭春明,赵旺,马振华,于刚. 红腹海参消化道指标、组织学和酶活性的季节变化. 南方水产科学. 2022(05): 39-45 . 本站查看
8. 周建聪,顾志峰,叶丙聪,刘闯,羊玉梅,刘春胜,王爱民,石耀华. 盐度和氨氮对方斑东风螺存活和能量收支的影响. 海洋科学. 2022(10): 104-112 . 百度学术
9. 韩朝婕,陈屹洋,贺振楠,张严匀,周文礼,高金伟,贾旭颖. 氨氮胁迫对水产动物生长、消化酶及免疫影响的研究进展. 河北渔业. 2021(05): 32-35 . 百度学术
10. 丁炜东,曹丽萍,曹哲明,邴旭文. 氨氮胁迫对翘嘴鳜幼鱼鳃、消化道酶活力的影响. 南方水产科学. 2020(03): 31-37 . 本站查看
其他类型引用(6)




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量: 0
- 被引次数: 16
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
