
Effects of fermented Astragalus membranaceus on growth, digestion, immune function and ammonia nitrogen resistance of Epinephelus fuscoguttatus
-
摘要: 为研究开发新型的促生长、增强免疫功能的水产饲料添加剂,采用发酵黄芪 (Astragalus membranaceus) 添加比例为0% (对照组)、0.25%、0.5%、1%、2%、4%的6种饲料 (分别记为L0、L1、L2、L3、L4、L5组),养殖体质量为 (44.48±2.06) g的棕点石斑鱼 (Epinephelus fuscoguttatus) 56 d,随后进行氨氮胁迫实验,以生长、消化、免疫功能为指标,评价发酵黄芪对棕点石斑鱼的生理生化和抗逆效应,得出最佳使用比例。结果显示:1) L3和L4组与对照组相比,饲料中添加发酵黄芪可显著提高棕点石斑鱼的体质量增长率 (WGR) 和特定生长率 (SGR) (P<0.05),显著降低饲料系数 (FCR) (P<0.05),还可显著提升胃肠道中消化酶活性 (P<0.05);2) 氨氮胁迫前,除了L5组,发酵黄芪可显著提升肝脏抗氧化性能 (P<0.05),显著降低肝脏丙二醛 (MDA) 含量和血清谷草转氨酶 (GOT) 活性 (P<0.05);各实验组血清谷丙转氨酶 (GPT) 活性和葡萄糖 (GLU) 浓度,及L3、L4和L5组血清甘油三酯 (TG) 浓度均显著低于对照组 (P<0.05);3) 氨氮胁迫后,发酵黄芪未提升肝脏抗氧化性能,但相较于对照组,显著降低了肝脏MDA含量 (除L5组)、血清GOT、GPT活性和GLU浓度 (除L4、L5组) (P<0.05)。综上,棕点石斑鱼饲料中发酵黄芪的适宜添加量为1%~2%。Abstract: To develop a new type of aquatic feed additive for promoting growth and enhancing immune function, we cultured Epinephelus fuscoguttatus with body mass of (44.48±2.06) g for 56 d by adding fermented Astragalus membranaceus with proportions of 0% (Control group), 0.25%, 0.5%, 1%, 2% and 4% (L0, L1, L2, L3, L4 and L5 groups), and then conducted an ammonia nitrogen stress test. Taking growth, digestion and immune function as the indicators, we evaluated the physiological, biochemical and stress-resistant effects of fermented A. membranaceus on E. fuscoguttatus, and determined the optimal ratio. The results show that: 1) Compared with the control group, the addition of fermented A. membranaceus increased the weight gain rate (WGR), specific growth rate (SGR), and the activities of digestive enzymes in gastrointestinal tract significantly (P<0.05), but reduced the feed conversion ratio (FCR) of E. fuscoguttatus in L3 and L4 groups significantly (P<0.05). 2) Before the ammonia nitrogen stress, fermented A. membranaceus increased the liver antioxidant performance significantly (Except L5 group, P<0.05), but decreased the liver malondialdehyde (MDA) content and serum glutamic-oxalacetic transaminase (GOT) activity significantly (P<0.05). The serum glutamic-pyruvic transaminase (GPT) activity, glucose (GLU) concentration and serum triglyceride (TG) concentration in L3, L4 and L5 groups were significantly lower than those in the control group (P<0.05). 3) After the ammonia nitrogen stress, fermented A. membranaceus did not improve the antioxidant performance of liver, but reduced the MDA content in liver (Except L5 group), the activities of GOT and GPT as well as the GLU concentration in serum (Except L4 and L5 groups) significantly (P<0.05). In conclusion, the recommended feed additive proportion of fermented A. membranaceus is 1%–2%.
-
罗非鱼肉质细嫩,骨刺少,富含蛋白质和多种氨基酸,含脂量低[1-2],深受消费者欢迎。但因水分含量高,容易受酶和微生物的影响而发生腐败变质[3],常温条件下无法保存,目前多以生鲜食用和冷冻加工为主[4]。
随着现代消费模式的改变,人们对方便速食产品的需求逐渐增加。对鱼肉进行加工,使之便于室温存放,满足开袋即食或冲泡即食的需求成为当前研究热点之一。真空冷冻干燥作为一种条件温和的干燥方式成为本研究的首选。真空冷冻干燥适用于热敏性成分原料 (果蔬和水产品) 的干燥,但因时间长、效率低、能耗高等因素限制了其在水产品中的应用。
研究表明用非热预处理或热处理后再干燥,可在一定程度上提高原料的干燥效率和干制品的复水性[5-6]。渗透脱水是一种非热预处理手段,其利用细胞内外部渗透压,促使水分迁移,改变原料对自由水的束缚能力,从而影响原料的干燥效率[7]。食盐是常用的渗透剂之一。食盐腌制不仅可降低水分含量、延缓烹饪损失、加速肌苷酸的形成并赋予产品主要滋味,还提升了整体感官品质。但食盐摄入过多会造成健康问题,有研究转向了低盐(质量分数1.5%~2.0%) 腌制[8],在低盐环境下添加复合渗透剂一定程度上弥补了因食盐用量过低导致的品质下降问题[9];张芸等[10]发现糖醇代替部分盐腌制处理,鱼肉的出品率、持水性明显提高,细胞保持完整;郑曼[11]研究表明丙二醇+丙三醇+氯化钠 (NaCl) 预处理可提高罗非鱼片的干燥速率和复水性。
笔者项目组在前期研究中发现热处理提高了细胞通透性,使细胞持水力下降,干燥时可加快水分散失并显著提升干燥速率和复水性[12-13]。姚娜娜等[14]研究表明添加剂和热处理均缩短了大果沙棘(Hippophae rhamnoides) 的干燥时间。目前有关热处理与非热处理相结合提升水产品干燥特征的研究尚未见报道。本研究以热处理样品为对照,分析了热处理与非热处理联合对提升干燥速率及复水性的影响,探讨了渗透剂对鱼肉冷冻干燥的影响机制,以期为罗非鱼肉冷冻干燥产品开发提供理论依据。
1. 材料与方法
1.1 材料与试剂
新鲜罗非鱼 (每尾400~500 g),购于广州超市。
食用盐 (广东省广盐集团股份有限公司);丙二醇 (食品级,美媛生物科技有限公司);蛋白测定试剂盒、总巯基、羰基含量测定试剂盒 (南京建成生物工程研究所);12% (体积分数) Bis-Tris预制胶、SDS 电泳缓冲液 (20×)(上海英潍捷基贸易有限公司);预染蛋白分子量标准 (6.5~270 kD)、SDS-PAGE 蛋白上样缓冲液(5×)、考马斯亮蓝染色液 (上海碧云天生物技术有限公司)。
1.2 仪器与设备
CR-400色差计 (日本柯尼卡美能达控股公司);Alpha1-4 真空冷冻干燥机 (德国Christ 公司);H1850R台式高速冷冻离心机 (湘仪离心机仪器有限公司);BS124S电子天平 (德国 Sartorius 公司);HH-4 快速恒温数显水浴箱 (常州澳华仪器公司);CT3 质构仪 (美国Brookfield公司);Mini Gel Tank PAGE 电泳槽 (美国赛默飞科技公司);Sunrise-basic Tacan酶标仪 (瑞士TECAN公司)。
1.3 实验方法
1.3.1 原料预处理
取罗非鱼背部肌肉,切成3 cm×3 cm×1 cm的小块,质量约为12 g,吸干鱼肉表面水分,在4 ℃条件下浸渍预处理1 h,浸渍液 (质量分数) 分别为:1.5%食盐、3%丙二醇、1.5%食盐+3%丙二醇,对照组不做任何处理。浸渍结束后滤纸吸去鱼肉表面水分,100 ℃沸水烫漂1 min,滤纸吸去表面水分后进行真空冷冻干燥。真空冷冻干燥:− 40 ℃冰箱预冻12 h,真空冷冻干燥冷阱温度 – 40 ℃,真空度20 Pa。
1.3.2 蒸煮损失
鱼肉经100 ℃沸水烫漂1 min取出,吸干表面水分,然后准确称质量,烫漂前后鱼肉质量差与烫漂前质量之比即为蒸煮损失率。
$$ {R}_{\mathrm{c}\mathrm{l}}=\frac{{m}_{\mathrm{b}}-{m}_{{\rm{a}}}}{{m}_{\mathrm{b}}}\times 100{\text{%}} $$ (1) 式中:Rcl为蒸煮损失率;mb、ma分别为烫漂前后鱼肉的质量 (g)。
1.3.3 干燥曲线
参考崔清亮等[15]的方法,略作修改。测定干燥过程罗非鱼肉的质量,至含水率约5%时停止干燥,计算含水率和样品干燥速率。
$$ M=\frac{{m}_{t}-{m}_{0}}{{m}_{t}}\times 100{\text{%}} $$ (2) $$ {R}_{\mathrm{d}}=\frac{{M}_{t}-{M}_{t+4}}{\Delta t} $$ (3) 式中:M为含水率 (%);mt为不同时间段样品质量 (g);m0为最终干燥后质量 (g);Rd为干燥速率 (%·h−1);Mt为不同时间段样品含水率 (%);Mt+4为 (t+4) h样品含水率 (%);Δt为4 h。
1.3.4 复水率
参考刘书成等[16]的方法,略做修改。干制样品80 ℃水浴锅复水60 min,每10 min测一次质量。
$$ {R}_{\mathrm{r}}=\frac{{m}_{1}-{m}_{0}}{m_1}\times 100{\text{%}} $$ (4) 式中:Rr为复水率 (%);m0、m1为复水前后鱼肉的质量 (g)。
1.3.5 白度
参考Chen等[17]的方法,略做修改。干制鱼肉白度 (Wn) 采用色差仪测定,室温下测定干样品的L*、a*、b*值。L*为明度,正值表示偏白,负值表示偏黑;a*为红绿值,正值表示偏红,负值表示偏绿; b*为黄蓝值,正值表示偏黄,负值表示偏蓝。
$$ {W}_{n}=100-\sqrt{{\left(100-{L^{^*}}\right)}^{2}+{a}^{*2}+{b}^{*2}} $$ (5) 1.3.6 质构的测定
用质构仪测定复水后鱼肉样品的硬度、弹性、咀嚼性和内聚性,参考吴燕燕等[18]的方法。每组实验重复6次。
1.3.7 肌原纤维蛋白分析
1) 肌原纤维蛋白的提取和分析
参考崔燕等[19]的方法,称2.0 g干制鱼肉,加入20 mL预冷Tris-maleate缓冲溶液 (50 mmol·L−1 KCl-20 mmol·L−1 Tris-maleate, pH 7.0),充分匀浆,4 ℃ 冷冻离心 (10 000 r·min−1, 10 min),除上清液,沉淀中加入20 mL预冷Tris-maleate 缓冲溶液 (0.6 mol·L−1 KCl-20 mmol·L−1 Tris-maleate, pH 7.0),匀浆后4 ℃静置提取1 h,4 ℃冷冻离心 (10 000 r·min−1, 10 min),上清液为肌原纤维蛋白溶液。Bradford 法测定肌原纤维蛋白溶液浓度。
表面疏水性的测定参考Iiham等[20]的方法,20 mmol·L−1 pH 6.0的磷酸缓冲液将肌原纤维蛋白稀释至2 mg·mL−1,取1 mL稀释液加100 µL (1 mg·mL−1) 溴酚蓝混合均匀,并做空白对照,涡旋震荡10 min,4 ℃冷冻离心 (3 000 r· min−1, 15 min),上清液稀释50倍,用酶标仪在595 nm波长下测定吸光值。
$$ {A}_{\mathrm{BPB}}=200\times \frac{({\mathrm{O}\mathrm{D}}_{\mathrm{c}}-{\mathrm{O}\mathrm{D}}_{\mathrm{s}})}{{\mathrm{O}\mathrm{D}}_{\mathrm{c}}} $$ (6) 式中:ABPB为溴酚蓝结合量 (μg);ODc、ODs分别为对照组和样品的吸光值。
巯基、羰基按照总巯基测定试剂盒、羰基含量测定试剂盒说明书进行测定。
2) 分子间的作用力
测定鱼肉样品肌原纤维蛋白分子间作用力,参考Pan等[21]方法。鱼肉样品分别与不同缓冲溶液按1∶19体积比进行均质匀浆,缓冲溶液分别包括SA (50 mmol·L−1 NaCl)、SB (600 mmol·L−1 NaCl)、SC (600 mmol·L−1 NaCl+1.6 mol·L−1 尿素)、SD (600 mmol·L−1 NaCl+8 mol·L−1尿素)、SE (600 mmol·L−1 NaCl+8 mol·L−1尿素+1.5 mol·L−1 β-巯基乙醇)。4 ℃静置1 h,10 000 r·min−1 离心15 min,测定上清液蛋白溶液浓度确定离子键 (SB−SA)、氢键 (SC−SB)、疏水相互作用 (SD−SC)、二硫键 (SE−SD) 质量浓度。
3) SDS-PAGE 凝胶电泳分析
按3∶1体积比将肌原纤维蛋白与SDS-PAGE上样缓冲液 (5×) 混合,混合液沸水浴煮5 min,取10 μg混合体系,在12% (体积分数) Bis-Tris预制胶进行上样分析,电泳电压120 V,电泳时间约1 h,电泳结束,用考马斯亮蓝染色液对胶板进行染色、然后脱色,扫描蛋白条带进行分析。
1.4 数据分析
每组实验均取3次平行进行测定,采用SPSS 19.0软件对实验数据进行处理与显著性分析,结果以“平均值±标准差 (
$\overline { X}\pm { \rm {SD}} $ )”表示,用Duncan's 法进行组间多重比较,显著性水平P<0.05,使用Origin 8.5软件作图。2. 结果与分析
2.1 蒸煮损失
蒸煮损失可反映肉制品的保水性能[22]。热处理后样品质量均有减轻,原因是可溶性化合物溶出、脂肪水解和水分流失,其中水分流失是导致质量减轻的主要原因[23]。如图1所示,与对照组相比,不同渗透处理均可降低鱼肉的蒸煮损失 (P<0.05),复合渗透剂对蒸煮损失的影响与单一食盐渗透的效果无显著性差异 (P>0.05)。烫漂过程中蛋白质发生热变性,持水力下降,造成质量下降,蒸煮损失增大。而经过渗透处理可改变鱼肉蛋白与水的结合状态或结合程度,从而降低产品蒸煮损失。吴亮亮等[24]研究发现低浓度食盐具有保水作用,食盐添加使不易流动水与肌原纤维蛋白的结合程度增加,引起肌原纤维溶胀,保水性增强;李雨露和刘丽萍[25]发现食盐质量分数低于3%的保水性较强。丙二醇是二羟基化合物,具有良好的亲水性和吸湿性,烫漂过程中鱼肉细胞在汁液流失后从环境中吸收一部分水分,降低蒸煮损失[26]。
![]()
2.2 干燥速率曲线
罗非鱼肉含水率随干燥时间的延长逐渐降低,8~12 h处理组含水率下降显著 (图2-a)。与对照组相比,渗透处理的罗非鱼肉含水率下降趋势更显著,3组渗透处理的鱼肉干燥过程含水率变化趋势表现出相似性,干燥期间含水率差异不明显。鱼肉经渗透剂处理后,样品水分质量分数达到5%时所需时间较对照组缩短约4~5 h。由于渗透溶液中溶质的填充作用,肌肉内部与外部水分形成较好的水分扩散通道,从而促进了水分的去除[27-28]。
如图2-b所示,前期干燥速率处于加速阶段,第12小时最高,随后随时间的延长而降低。复合渗透处理19 h,食盐和丙二醇渗透处理组第20小时鱼肉含水率低于5%,此时对照组含水率为12.72%,说明渗透处理可使样品干燥的时间明显缩短。真空冷冻干燥前期的干燥速率高是因为添加物渗透作用,细胞内外为了维持渗透压平衡,胞内水分流出集中在细胞间隙,水分流失快;细胞内水分与大分子结合,不易流失,因而解吸阶段主要是结合水的升华,干燥速率降低。干燥速率随干燥时间的延长而降低,也有部分原因是水分从物料内部向表面的迁移速率与物料表面的水分蒸发速率的不一致[29]。食盐渗透处理的干燥速率略低于其他两组,可能同食盐与细胞内水分结合、传质推动力减弱使干燥速率下降有关[30],食盐渗入增加了物料内部的含水量,影响了水分的扩散和蒸发。
2.3 复水率
复水是干制品食用前的一个重要操作,复水率与细胞和结构的破坏程度密切相关。由图3可知,丙二醇与复合渗透剂可显著提高干制鱼肉的复水性 (P<0.05),丙二醇渗透处理的复水率最高 (71.11%),而经过食盐渗透处理则明显降低了干制鱼块的复水性。复水率降低是因为NaCl使蛋白质变性发生交联,肌肉组织收缩[31],增加了鱼肉在干燥过程中的组织损伤,干制后鱼块的肌肉纤维组织具有致密性,吸水能力减弱不利于复水[32]。韩珍琼和马菁[33]研究发现食盐渗入会破坏荷兰豆 (Pisum sativum var. saccharatum) 细胞组织,造成细胞质壁分离,引起脱水,从而使干制品复水性下降。丙二醇因其含有羟基基团,具有良好的亲水性,吸水能力强,复水率高[34];复合渗透剂中丙二醇对肌肉组织吸水作用力强,减弱肌肉组织的收缩性,减小组织结构的变形,有效维持细胞的完整性,因而复水率高。
![]() 图 3 渗透处理对干制罗非鱼块复水率的影响Figure 3. Effect of osmotic treatment on rehydration rate of dried tilapia fillets
图 3 渗透处理对干制罗非鱼块复水率的影响Figure 3. Effect of osmotic treatment on rehydration rate of dried tilapia fillets2.4 冻干罗非鱼品质测定
由表1可知,与对照组相比,添加食盐干制鱼肉白度显著降低 (P<0.05),丙二醇和复合渗透剂对鱼肉颜色的影响不显著 (P>0.05)。单独食盐处理的鱼肉褐变严重,实验观察到干制鱼肉表面呈微黄色。鱼肉的白度与鱼体内的水分和氧化发生有关,食盐添加使鱼块肌肉脱水、收缩,光折射减弱,造成鱼块色泽变暗,盐渍过程会加速脂质氧化,也可能与NaCl促进氧化使肌红蛋白氧化生成褐色的高铁肌红蛋白有关[35],腌制时间越长与环境中的氧长时间接触使L*下降[36];丙二醇有良好的亲水性,可提高鱼肉持水力和降低氧化,能有效抑制血红蛋白氧化,因而丙二醇和复合渗透剂对鱼肉白度影响不显著。
表 1 渗透处理对干制鱼肉白度和复水后质构的影响Table 1. Effect of osmotic treatment on whiteness and texture of dried fish after rehydration处理 Treatment 白度Whiteness 硬度Hardness/g 弹性Elasticity/mm 咀嚼性Chewability/mJ 内聚性Cohesion 对照 Control 20.4±0.85b 81.0±4.2a 2.87±0.18a 1.58±0.35a 0.69±0.08b 食盐 Salt 14.90±1.01a 94.3±3.9b 3.44±0.23b 2.01±0.06b 0.50±0.01a 丙二醇 Propylene glycol 21.43±0.62b 82.3±4.5a 3.21±0.27b 1.55±0.08a 0.63±0.09b 食盐+丙二醇 Salt+Propylene glycol 19.49±0.81b 92.8±4.5b 3.37±0.24b 2.27±0.05b 0.68±0.02b 注:同列字母不同表示差异显著 (P<0.05);下表同此。 Note: Different letters with in the same column indicate significant differences (P<0.05). The same case in the following table. 与对照组相比,添加食盐对鱼肉复水后的质构影响显著 (P<0.05),硬度和弹性增加,内聚性减弱;丙二醇添加对鱼肉硬度和内聚性无显著性影响,但提高了鱼肉的弹性。食盐腌制有利于产品产生高弹性、耐咀嚼[37];吴素娟等[38]研究发现鱼肉腌制后硬度、弹性增加。NaCl增大离子强度,改变肌原纤维蛋白分子表面的电荷分布,从而改变蛋白质结构,多肽链的交互作用增强,提高了凝胶特性[39]。适盐浓度有利于形成蛋白质凝胶网络,提高鱼肉制品的质构[40-41]。丙二醇分子式中含有吸水基团羟基,具有高复水性,因而对复水后组织硬度影响不显著;醇类渗透液进入到肌肉组织内部,使得肌肉间隙变小,提高鱼片的弹性及口感。复合渗透剂综合了食盐和丙二醇的特点,联合作用对鱼肉的硬度、弹性和咀嚼性均有提高,赋予鱼肉复水后良好的口感。
2.5 肌原纤维蛋白特性的测定
2.5.1 肌原纤维蛋白
渗透剂处理对鱼肉肌原纤维蛋白含量的影响显著。食盐腌制使鱼肉肌原纤维蛋白和总巯基质量摩尔浓度显著下降 (P<0.05),表面疏水性和羰基质量摩尔浓度上升(表2)。食盐增加了离子强度,影响蛋白质的表面电荷,从而促进肌原纤维蛋白溶解[42];黄彬等[43]研究发现盐水煮使鸡肉盐溶性蛋白含量降低,腌制过程食盐引起肌肉中蛋白质的氧化修饰,影响表面疏水性和羰基含量。同时Na+和Cl–进入细胞组织,破坏细胞内静电平衡,引起肌肉中蛋白质的氧化修饰,构象发生改变,使蛋白质表面的氨基酸残基受到攻击形成羰基[44-45],蛋白质分子之间的相互交联,促进巯基氧化为二硫键。
表 2 渗透处理对肌原纤维蛋白特性的影响Table 2. Effect of osmotic treatment on myofibrillar protein properties处理Treatment 肌原纤维质量浓度Myofibrillar mass
concentration/(mg·mL−1)表面疏水性Surface hydrophobicity/
μg总巯基质量摩尔浓度Total Thiol molality/
(μmol·g−1)羰基质量摩尔浓度Carbonyl molality/(nmol·mg−1 ) 对照 Control 4.22±0.09b 23.73±1.32b 3.32±0.05c 2.42±0.09b 食盐 Salt 3.90±0.12a 27.83±1.02c 2.85±0.17a 2.65±0.42c 丙二醇 Propylene glycol 4.79±0.05c 20.74±1.71a 4.11±0.26d 1.97±0.13a 食盐+丙二醇 Salt + Propylene glycol 4.16±0.07b 22.75±1.22ab 3.15±0.13b 2.50±0.29b 与对照组相比,丙二醇处理的肌原纤维质量浓度显著升高 (P<0.05),说明添加物具有抑制蛋白氧化的作用。沈春蕾等[46]研究表明虾仁经糖醇渗透处理后冻藏,贮藏过程中抑制蛋白巯基氧化转化为二硫键,维持肌肉中肌原纤维蛋白稳定;二羟基醇与肌肉组织中的水分子和蛋白质通过形成氢键而发生结合,起到稳定肌原纤维蛋白结构的作用,从而维持蛋白质功能特性,降低空间构象变化,确保蛋白质的二级和三级结构稳定。羟基与蛋白质大分子结合,使肌纤维蛋白质溶液中自由水转为结合水,因而降低肌纤维蛋白的共晶点温度,减少冰晶形成,抑制疏水性基团的释放、减缓肌原纤维蛋白质表面疏水性的增加、影响肌纤维蛋白质巯基含量,从而减弱蛋白质分子的聚集和降低蛋白质变性程度[47-48],提高鱼肉持水性,保持鱼肉品质。复配处理也有效减缓了对蛋白的氧化降解,延缓鱼肉肌原蛋白的冷冻变性,表面疏水性和羰基含量低于单一食盐添加组,提高蛋白稳定性。有研究表明,醇与NaCl复配使用显著提高了干制罗非鱼片的Ca2+-ATP酶活性和盐溶性蛋白溶解度,降低了蛋白质变性,减少了营养成分流失,从而改善了干鱼片品质[11]。
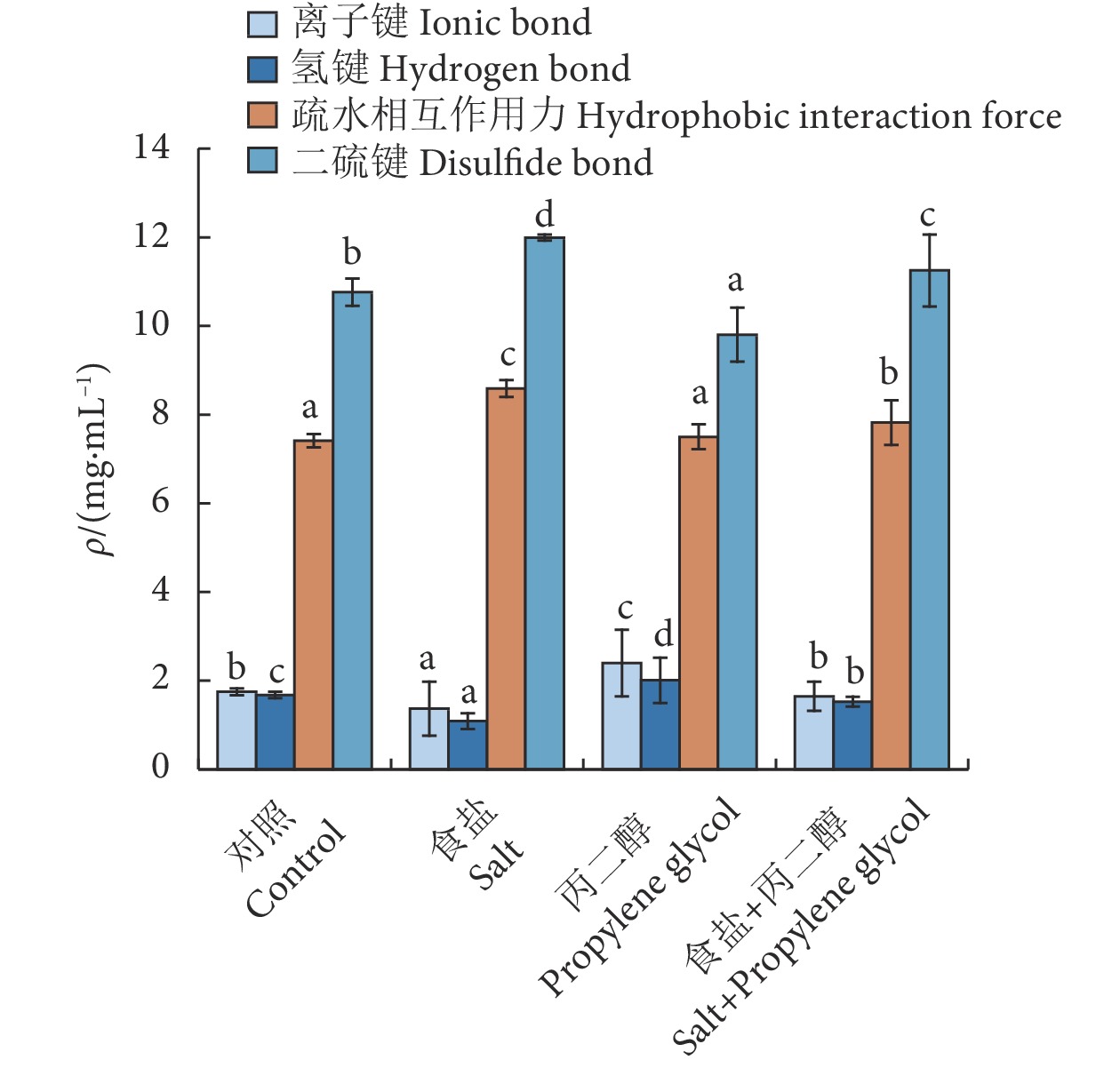
2.5.2 分子间化学作用力
离子键和氢键是维持肌原纤维蛋白天然结构的主要作用力,二者的键合力弱于疏水相互作用和二硫键[49],易受环境影响使结构发生改变。预处理和冷冻干燥过程对肌肉蛋白质产生了影响,维持蛋白质结构的化学作用力发生了一定的改变。范露等[50]研究表明火腿的腌制、发酵过程影响了蛋白质的结构和相互作用力,其离子键和氢键相对含量下降,疏水相互作用和二硫键相对含量上升。
与对照组相比 (图4),食盐处理的离子键和氢键质量浓度显著降低 (P<0.05),疏水相互作用和二硫键质量浓度增加,表明食盐腌制破坏蛋白结构。离子键和氢键含量降低可能与食盐渗透作用导致细胞内汁液和细胞间水分的流失有关,鱼肉腌制过程中离子键和氢键断裂,含量降低[50];渗透过程蛋白发生聚集产生交联作用[51],内部的疏水性基团暴露在蛋白质分子表面,使蛋白质表面疏水性发生改变,肌原纤维蛋白空间结构改变,巯基暴露被氧化生成二硫键[52-53]。
![]() 图 4 渗透处理对肌原纤维蛋白分子间化学作用力的影响注:同色方柱不同字母表示组间差异显著 (P<0.05)。Figure 4. Effect of osmotic treatment on intermolecular chemical force of myofibrillar proteinNote: Different letters on the same color bar indicate significant difference among the groups (P<0.05).
图 4 渗透处理对肌原纤维蛋白分子间化学作用力的影响注:同色方柱不同字母表示组间差异显著 (P<0.05)。Figure 4. Effect of osmotic treatment on intermolecular chemical force of myofibrillar proteinNote: Different letters on the same color bar indicate significant difference among the groups (P<0.05).与对照组相比,添加丙二醇的离子键和氢键质量浓度显著提高 (P<0.05),二硫键质量浓度显著降低 (P<0.05),蛋白稳定性提高。亲水性添加剂可以与水分子氢键结合,增强氢键作用力[54],蛋白质表面的部分水分子被代替,减少蛋白质氢键连接点的暴露,使蛋白质高级结构维持稳定;羟基醇通过与大分子结合在极性条件下具有蛋白保护作用,预冻过程中抑制冰晶体的形成,延缓组织中蛋白质的降解和变性。复合渗透剂中丙二醇与水结合力增强,蛋白质分子聚集减弱,抑制疏水基团释放,蛋白质变性程度降低,细胞组织结构稳定,复合渗透剂有效保护肌纤维排列的有序性,维持蛋白质构象。
2.5.3 SDS-PAGE
肌原纤维蛋白主要包含肌球蛋白重链、肌动蛋白、原肌球蛋白和一些调节性结构蛋白。对照组和复合渗透处理的鱼肉中肌球蛋白重链和一些大分子蛋白条带 (95~270 kD) 变浅,单独食盐渗透处理组鱼肉肌球蛋白重链和一些大分子条带消失,单一丙二醇渗透处理的肌原纤维蛋白条带效果最好 (图5)。说明经过食盐渗透处理肌原纤维蛋白发生了一定程度的降解,这与吴林洁等[55]的研究结果一致。有研究表明添加NaCl可使蛋白质部分结构展开,变性焓值降低[56],蛋白发生降解,肌球蛋白重链消失和一些大分子条带变浅。丙二醇渗透剂形成的“玻璃态”可抑制冰晶的形成[57],抑制蛋白质的降解,减轻对蛋白质结构的破坏。混合处理组蛋白条带丰度虽不及单一醇处理组,与对照组相比无明显减弱迹象,但明显高于盐处理组样品。
![]() 图 5 渗透处理对鱼肉肌原纤维蛋白SDS-PAGE的影响Figure 5. Effect of osmotic treatment on SDS-PAGE in fish myofibrillar protein
图 5 渗透处理对鱼肉肌原纤维蛋白SDS-PAGE的影响Figure 5. Effect of osmotic treatment on SDS-PAGE in fish myofibrillar protein3. 结论
渗透处理可有效降低蒸煮损失,提高干燥速率。与对照组 (24 h) 相比,复合渗透剂处理的鱼肉冷冻干燥时间最短 (19 h),食盐和丙二醇渗透处理后干燥时间明显缩短 (20 h)。
食盐腌制的鱼肉复水后硬度提高,具有较好的咀嚼性,但食盐会破坏蛋白结构,使蛋白分子展开,促进二硫键的形成,使羰基和总巯基浓度降低,导致蛋白质变性,SDS-PAGE中肌球蛋白重链消失和其他大分子蛋白条带变浅。丙二醇渗透处理后干制品复水效果最佳,复水后鱼肉的弹性提高,且具有良好的口感,丙二醇中的羟基对蛋白有保护作用,减少了蛋白氧化变性。
复合渗透处理综合了盐和丙二醇处理的优势,复合处理后可有效减少蒸煮损失,在提高产品干燥速率的同时,保持了干制品良好的色泽,并提高了复水后鱼肉的硬度、弹性和咀嚼性;同时起到降低蛋白氧化程度、维持蛋白结构稳定的作用,可作为鱼肉冷冻干燥渗透处理的首选。
-
![]()
图 1 发酵黄芪对棕点石斑鱼胃蛋白酶、前肠胰蛋白酶、淀粉酶和脂肪酶活性的影响
注:方柱上不同小写字母表示有显著性差异 (P<0.05)。
Figure 1. Effects of fermented A. membranaceus on pepsin in stomach and trypsin, amylase and lipase activities in foregut of E. fuscoguttatus
Note: Different lowercase letters indicate significant differences (P<0.05).
![]()
图 2 发酵黄芪对棕点石斑鱼氨氮胁迫前后肝脏过氧化氢酶活性、总抗氧化能力、超氧化物歧化酶活性和丙二醛浓度的影响
注:方柱上不同小写字母表示胁迫前差异显著 (P<0.05),不同大写字母表示胁迫后差异显著 (P<0.05),图3同此。
Figure 2. Effects of fermented A. membranaceus on CAT activity, T-AOC, SOD activity and MDA concentration in liver of E. fuscoguttatus before and after ammonia nitrogen stress
Note: Different lowercase and uppercase letters on the bars indicate significant differences before and after ammonia nitrite stress, respectively (P<0.05). The same case in Fig. 3.
![]()
图 3 发酵黄芪对棕点石斑鱼氨氮胁迫前后血清谷草转氨酶、谷丙转氨酶、碱性磷酸酶活性和甘油三酯、葡萄糖浓度的影响
Figure 3. Effects of fermented A. membranaceus on GOT, GPT, AKP activity and TG, GLU concentrations in serum of E. fuscoguttatus before and after ammonia nitrogen stress
表 1 发酵黄芪对棕点石斑鱼生长性能的影响
Table 1 Effects of fermented A. membranaceus on growth performance of E. fuscoguttatus
组别Group 成活率SR/% 体质量增长率WGR/% 特定生长率SGR/(%·d−1) 饲料系数FCR 肥满度CF/(g·cm−3) 脏体比VSI/% 肠脂比IPF/% L0 (0%) 98.89±1.92 75.09±6.38c 1.00±0.06c 1.31±0.17a 0.10±0.01ab 8.27±1.55 2.38±0.42 L1 (0.25%) 98.89±1.92 84.27±5.65bc 1.09±0.06bc 1.23±0.18a 0.07±0.05b 9.21±1.75 3.02±1.73 L2 (0.5%) 98.89±1.92 77.76±8.16bc 1.03±0.08bc 1.40±0.18a 0.11±0.05ab 8.73±1.06 2.27±0.87 L3 (1%) 100±0.00 90.18±9.53b 1.15±0.10b 0.93±0.03b 0.09±0.04ab 8.68±1.44 2.17±0.98 L4 (2%) 100±0.00 105.00±7.10a 1.28±0.06a 0.93±0.07b 0.09±0.04ab 8.21±1.49 2.25±0.72 L5 (4%) 100±0.00 79.42±5.13bc 1.04±0.05bc 1.24±0.04a 0.11±0.05a 8.65±2.29 2.56±0.64 注:同列数据不同上标小写字母表示差异显著 (P<0.05)。 Note: Values with different lowercase letters within the same column are significantly different (P<0.05).  下载: 导出CSV
下载: 导出CSV
-
[1] 孙晓飞, 郭伟良, 谢珍玉, 等. 棕点石斑鱼中草药免疫增强剂的快速筛选[J]. 渔业科学进展, 2015, 36(1): 54-60. [2] 孙颖. 棕点石斑鱼(♀)×鞍带石斑鱼(♂)杂交F1代生长优势的转录组学研究[D]. 广州: 中山大学, 2016: 2. [3] 蔡岩. 三种中草药对棕点石斑鱼的免疫调控及其作用机制研究[D]. 海口: 海南大学, 2017: 1. [4] IP Y K, CHEW S F. Air-breathing and excretory nitrogen metabolism in fishes[J]. Acta Histochem, 2018, 120(7): 680-690. doi: 10.1016/j.acthis.2018.08.013
[5] 郑洪武. 氨氮胁迫对大口黑鲈相关器官及免疫因子的影响研究[D]. 舟山: 浙江海洋大学, 2020: 1. [6] 吴进喜. 氨氮、亚硝酸盐和姜黄素对海鲈生长代谢及抗病相关基因的影响[D]. 湛江: 广东海洋大学, 2020: 1-3. [7] 王美姿. 氨氮胁迫下饲料蛋白质水平对吉富罗非鱼生长、血清生化指标和组织结构的影响[D]. 上海: 上海海洋大学, 2018: 1-7. [8] PU H Y, LI X Y, DU Q B, et al. Research progress in the application of Chinese herbal medicines in aquaculture: a review[J]. Engineering, 2017, 3(5): 731-737. doi: 10.1016/J.ENG.2017.03.017
[9] 付天祥. 试论中草药饲料添加剂在水产养殖中的应用[J]. 农家参谋, 2018(16): 113. [10] HAI N V. The use of medicinal plants as immunostimulants in aquaculture: a review[J]. Aquaculture, 2015, 446: 88-96. doi: 10.1016/j.aquaculture.2015.03.014
[11] 柴新娥, 项东, 项颖, 等. 中草药饲料添加剂在养殖中的应用进展[J]. 现代畜牧兽医, 2022(6): 85-88. [12] 张淑娟, 张育贵, 牛江涛, 等. 黄芪的研究进展及其质量标志物预测分析[J]. 中华中医药学刊, 2022, 40(2): 151-155. [13] 薛倩倩, 刘晓节, 李科, 等. 黄芪药材化学成分差异的研究进展[J]. 山西医科大学学报, 2018, 49(10): 1259-1263. [14] 倪慧艳, 陈伟, 宋文静. 黄芪多糖抗氧化作用研究[J]. 中医学报, 2017, 32(9): 1705-1707. [15] 白东清, 吴旋, 郭永军, 等. 长期投喂黄芪多糖对黄颡鱼抗氧化及非特异性免疫指标的影响[J]. 动物营养学报, 2011, 23(9): 1622-1630. [16] 刘金海, 陈恒, 罗小丽, 等. 半滑舌鳎鱼血液免疫因子对黄芪多糖的免疫应答[J]. 饲料研究, 2020, 43(9): 40-45. [17] LI Y, RAN C, WEI K J, et al. The effect of Astragalus polysaccharide on growth, gut and liver health, and anti-viral immunity of zebrafish[J]. Aquaculture, 2021, 540: 736677. doi: 10.1016/j.aquaculture.2021.736677
[18] 边亚彬. 发酵黄芪多糖的制备及其对小鼠树突状细胞成熟相关信号通路的影响[D]. 北京: 中国农业科学院, 2017: 13. [19] 苏贵龙, 张景艳, 张凯, 等. 益生菌发酵提高黄芪根、茎、叶活性成分含量的研究[J]. 中国畜牧兽医, 2017, 44(6): 1877-1883. [20] 乔宏兴, 史洪涛, 白静, 等. 植物乳杆菌和屎肠球菌协同固体发酵黄芪的互作及在断奶仔猪上的应用[J]. 中国兽医学报, 2018, 38(10): 1982-1988. [21] 李继开, 贾琳, 石建存, 等. 益生菌发酵中药饲料添加剂工艺的优化[J]. 饲料研究, 2021, 44(16): 59-62. [22] QIAO H X, SONG Y Z, SHI H T, et al. Fermented Astragalus in diet altered the composition of fecal microbiota in broiler chickens[J]. Amb Expr, 2018, 8(1): 151. doi: 10.1186/s13568-018-0682-4
[23] SHI H T, ZHAO S Z, WANG K L, et al. Effects of dietary Astragalus Membranaceus supplementation on growth performance, and intestinal morphology, microbiota and metabolism in common carp (Cyprinus carpio)[J]. Aquac Rep, 2022, 22: 100955. doi: 10.1016/j.aqrep.2021.100955
[24] 谢炎福. 复方中药发酵工艺优化及防治鲫鱼出血病的初步研究[J]. 饲料研究, 2015(13): 54-58. [25] 刘洋, 金顺义, 常娟, 等. 复合益生菌发酵中草药前后活性成分变化[J]. 安徽农业科学, 2017, 45(34): 123-125. doi: 10.3969/j.issn.0517-6611.2017.34.039 [26] 王坛, 华雪铭, 朱伟星, 等. 饲料溶菌酶添加水平对氨氮应激下吉富罗非鱼血清生化指标、抗菌性能和肝脏抗氧化能力的影响[J]. 水产学报, 2016, 40(5): 740-750. [27] HEGAZI M M, ATTIA Z I, HEGAZI M A M, et al. Metabolic consequences of chronic sublethal ammonia exposure at cellular and subcellular levels in Nile tilapia brain[J]. Aquaculture, 2010, 299(1/2/3/4): 149-156.
[28] 王贞杰, 叶保民, 常青, 等. 饲料维生素C含量对圆斑星鲽幼鱼抗氨氮胁迫能力的影响[J]. 动物营养学报, 2016, 28(12): 4054-4062. [29] HUSSAIN A, BOSE S, WANG J H, et al. Fermentation, a feasible strategy for enhancing bioactivity of herbal medicines[J]. Food Res Int, 2016, 81: 1-16. doi: 10.1016/j.foodres.2015.12.026
[30] 赵倩, 陈玉春, 高绪娜, 等. 枯草芽孢杆菌发酵中药制剂对鲤鱼生长性能、生化指标、抗氧化指标及抗感染能力的影响[J]. 中国畜牧兽医, 2017, 44(3): 724-731. [31] 谢炎福, 代春梅, 杜亚. 益生菌发酵复方中药对黄河鲤生长性能和免疫性能的影响[J]. 黑龙江畜牧兽医, 2015(15): 198-200. [32] TANG T, TONG F L, ZHAO S N, et al. Effects of fermented Broussonetia papyrifera on growth, intestinal antioxidant, inflammation and microbiota of grass carp (Ctenopharyngodon idella)[J]. Aquac Rep, 2021, 20: 100673. doi: 10.1016/j.aqrep.2021.100673
[33] 农林生, 黎建斌. 发酵黄芪对山瑞鳖稚鳖生长性能的影响[J]. 河北渔业, 2019(5): 8-10. doi: 10.3969/j.issn.1004-6755.2019.05.002 [34] 王煜恒, 徐孝宙, 王会聪, 等. 黄芪多糖对杂交鳢生长性能、免疫能力、抗氧化能力和抗病力的影响[J]. 动物营养学报, 2018, 30(4): 1447-1456. doi: 10.3969/j.issn.1006-267x.2018.04.028 [35] 向枭, 陈建, 周兴华, 等. 黄芪多糖对齐口裂腹鱼生长、体组成和免疫指标的影响[J]. 水生生物学报, 2011, 35(2): 291-299. [36] 韦海明. 黄芪和维生素C对大黄鱼抗应激的影响[D]. 青岛: 中国海洋大学, 2014: 27. [37] 李秋月, 林连兵, 杨雪娇, 等. 微生物发酵中草药的研究现状[J]. 微生物学通报, 2021, 48(6): 2232-2244. [38] 彭凯, 萧鸿发, 莫文艳, 等. 黑水虻幼虫粉替代鱼粉对加州鲈生长性能、形体指标、体成分及营养物质沉积率的影响[J]. 动物营养学报, 2021, 33(11): 6340-6348. doi: 10.3969/j.issn.1006-267x.2021.11.034 [39] ZHANG Z F, LYU G Y, PAN H J, et al. Production of powerful antioxidant supplements via solid-state fermentation of wheat (Triticum aestivum Linn.) by Cordyceps militaris[J]. Food Technol Biotechnol, 2012, 50(1): 32-39.
[40] GAO J, WANG R, LIU J X, et al. Effects of novel microecologics combined with traditional Chinese medicine and probiotics on growth performance and health of broilers[J]. Poult Sci, 2022, 101(2): 101412. doi: 10.1016/j.psj.2021.101412
[41] 张武肖, 孙盛明, 戈贤平, 等. 急性氨氮胁迫及毒后恢复对团头鲂幼鱼鳃、肝和肾组织结构的影响[J]. 水产学报, 2015, 39(2): 233-244. [42] 刘明阳. 茶树精油对罗氏沼虾生长、免疫及抗氨氮应激的影响[D]. 南京: 南京农业大学, 2020: 5. [43] 亢玉静, 郎明远, 赵文. 水生生物体内抗氧化酶及其影响因素研究进展[J]. 微生物学杂志, 2013, 33(3): 75-80. [44] 王芸, 李正, 段亚飞, 等. 红景天提取物对凡纳滨对虾抗氧化系统及抗低盐度胁迫的影响[J]. 南方水产科学, 2018, 14(1): 9-19. [45] 夏磊, 赵明军, 张洪玉, 等. 不同比例复合益生菌对凡纳滨对虾生长、免疫及抗氨氮能力的影响[J]. 中国水产科学, 2015, 22(6): 1299-1307. [46] 刘成荣, 刘丽香. 香菇多糖对氨氮胁迫泥鳅抗氧化功能的影响[J]. 生态学杂志, 2021, 40(11): 3701-3710. [47] 刘雨, 丁炜东, 曹哲明, 等. 急性氨氮胁迫对翘嘴鳜幼鱼抗氧化酶活性及炎症反应相关基因表达的影响[J]. 南方农业学报, 2019, 50(8): 1860-1868. doi: 10.3969/j.issn.2095-1191.2019.08.29 [48] LI J T, LI W T, ZHANG X M. Effects of dissolved oxygen, starvation, temperature, and salinity on the locomotive ability of juvenile Chinese shrimp Fenneropenaeus chinensis[J]. Ethol Ecol Evol, 2018, 31(2): 1-18.
[49] LI T Y, LI E C, SUO Y T, et al. Energy metabolism and metabolomics response of Pacific white shrimp Litopenaeus vannamei to sulfide toxicity[J]. Aquat Toxicol, 2017, 183: 28-37. doi: 10.1016/j.aquatox.2016.12.010
[50] 何玲, 王佩, 罗来婷, 等. 螺旋藻对中华鳖生长和体组成及血清生化指标的影响[J]. 湖南农业大学学报 (自然科学版), 2019, 45(5): 536-540, 559. [51] 刘梦迪, 周楠楠, 王腾飞, 等. 银鲳卵巢发育与磷酸酶ACP及AKP活性的关系[J]. 宁波大学学报 (理工版), 2020, 33(6): 13-19. [52] 刘卫红, 颜贤忠, 张蕾, 等. 代谢组学技术在高脂血症及动脉粥样硬化痰瘀演变研究中的应用[J]. 辽宁中医杂志, 2008(5): 684-686. doi: 10.3969/j.issn.1000-1719.2008.05.021 [53] ZHANG M Z, LI M, WANG R X, et al. Effects of acute ammonia toxicity on oxidative stress, immune response and apoptosis of juvenile yellow catfish Pelteobagrus fulvidraco and the mitigation of exogenous taurine[J]. Fish Shellfish Immunol, 2018, 79: 313-320. doi: 10.1016/j.fsi.2018.05.036
[54] dos SANTOS SILVA M J, BATISTA da COSTA F F, PAES LEME F, et al. Biological responses of neotropical freshwater fish Lophiosilurus alexandri exposed to ammonia and nitrite[J]. Sci Total Environ, 2018, 616/617: 1566-1575. doi: 10.1016/j.scitotenv.2017.10.157
-
期刊类型引用(6)
1. 朱文婷,李文嘉,宣雄智,赵娟. 渔用中草药种类、作用机理及应用效果研究进展. 中国饲料. 2024(05): 94-100 .  百度学术
百度学术
2. 杨希文,祁效林,戴衍朋,金国栋,李爱堂,张金霞,冯坤蓉,覃志江. 黄芪中有效成分的提取、生物活性功能及其在动物生产中的应用. 饲料研究. 2024(03): 149-153 . 百度学术
3. 张燕,贾阳,杜涓,刘娜,王园,齐景伟,安晓萍. 偏最小二乘法和近红外光谱技术快速测定发酵黄芪茎叶中黄酮含量及抗氧化活性. 饲料研究. 2024(18): 70-75 . 百度学术
4. 孟睦涵,马林,尤宏争,李明泽,夏苏东,孙学亮,毕相东. 黄芪及其多糖在水产养殖中的应用研究进展. 现代畜牧兽医. 2024(12): 77-81 . 百度学术
5. 章晓磊,毕相东,王鑫宇,戴伟. 复方中草药茎叶好氧发酵条件优化研究. 饲料研究. 2024(23): 119-124 . 百度学术
6. 曹雪,孙佳,杨质楠,梁爽,李月红. 中草药在水产动物养殖中的研究进展. 饲料研究. 2023(24): 133-137 . 百度学术
其他类型引用(1)
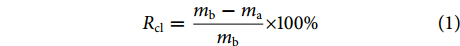
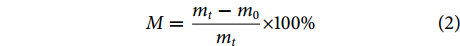
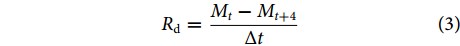
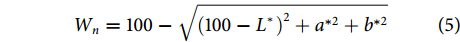
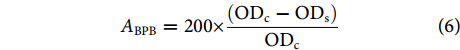







计量
- 文章访问数: 605
- HTML全文浏览量: 176
- PDF下载量: 56
- 被引次数: 7
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
