
Spatio-temporal distribution of habitat patterns of Scomber japonicus in East China Sea under future climatic conditions
-
摘要: 为探讨全球气候变化对东海鲐鱼 (Scomber japonicus) 栖息地时空分布的影响,减缓气候变化对鲐鱼渔场的不利影响,基于CMIP6气候模式输出的2.5 m (Temp_2.5 m)、25 m (Temp_25 m) 和50 m (Temp_50 m) 水温数据分析3种未来气候情境下 (SSP126、SSP370和SSP585) 东海鲐鱼栖息地的时空变化。结果表明,SSP126情境下,2015—2100年渔场内所有水层温度整体变化幅度较小。SSP370和SSP585情境下,各水层温度均明显上升。3个水层温度在2015—2020、2055—2060和2095—2100年具有明显空间变化。2015—2100年东海鲐鱼渔场的栖息地指数 (Habitat suitability index, HSI) 和适宜栖息地面积比例在SSP126、SSP370和SSP585情境下整体呈下降趋势,而不适宜栖息地面积呈上升趋势。不同气候变化情境下,东海鲐鱼适宜栖息地在2015—2020、2055—2060和2095—2100年3个时间段内主要分布在122°E—126°E、28°N—30°N范围内,不适宜栖息地主要分布在渔场中部;此外,鲐鱼适宜栖息地重心具有向北移动的趋势。研究表明,在未来全球变暖的气候背景下东海海域不易形成适宜的鲐鱼栖息地。Abstract: To investigate the effects of global climate change on the habitat pattern of Scomber japonicus in the East China Sea and to slow down the adverse effects of climate change on the fishing ground of S. japonicus, based on the water temperature data of 2.5 m (Temp_2.5 m), 25 m (Temp_25 m) and 50 m (Temp_50 m) output by CMIP6 climate model, we analyzed the spatio-temporal changes of the habitat in the East China Sea under three future climatic conditions (SSP126, SSP370 and SSP585). The results show that for SSP126, Temp_2.5 m, Temp_25 m and Temp_50 m showed relatively minor changes. For SSP370 and SSP585, Temp_2.5 m, Temp_25 m and Temp_50 m showed an overall upward trend. The three factors also changed in the spatial distribution during 2015−2020, 2055−2060 and 2095−2100. The habitat suitability index (HSI) on the fishing ground and proportion of suitable habitat of S. japonicus in the East China Sea from 2015 to 2100 showed a similar downward trend under SSP126, SSP370 and SSP585 conditions. However, the proportion of unsuitable habitat area from 2015 to 2100 increased under SSP126, SSP370 and SSP585 conditions. The suitable habitats of S. japonicus were mainly distributed in the waters of 122°E−126°E and 28°N−30°N during 2015−2020, 2055−2060 and 2095−2100, while the unsuitable habitat was mainly distributed in the middle of fishing ground. In addition, the gravity center of suitable habitat of S. japonicus tended to move northward. The results suggest that the global warming is unfavorable for the formation of suitable habitat of S. japonicus in the East China Sea.
-
ABC转运子(ATP-binding cassette transporter)广泛存在于真核和原核生物中,是细胞内物质运输系统的重要组成部分,主要功能为利用ATP水解产生的能量,对底物进行逆浓度梯度的跨膜运输,转运各种细菌生长所需的营养物质,如糖类、多肽、脂质、无机离子或抗生素等,并参与许多重要生理生化过程,如摄入营养、信号转导及抗原呈递等[1-6]。
核糖转运系统对于核糖的高亲和吸收至关重要,大肠杆菌的核糖转运系统由rbs操纵子(rbsA,-B,-C,-D,-K,-R)编码的6种蛋白质组成[7]。rbs操纵子转运核糖的途径如下,首先核糖通过外膜孔蛋白扩散到革兰氏阴性菌的周质中,游离核糖被RbsB结合,随后将核糖传递到由RbsAC形成的内膜复合物中,RbsAC再以ATP依赖的方式将底物跨膜运输至细胞质[8-10],核糖被激酶RbsK磷酸化后螯合在细胞中[11]。RbsR是结合到rbs操纵子转录起始位点的阻遏物,RbsD位于RbsK上游,可催化核糖改变构型,由吡喃构象转化为呋喃构象,从而发挥其功能[12]。RbsB主要功能为结合核糖,并将其转运至细胞中。除与核糖结合相关的功能外,近年来有研究发现RbsB蛋白还可介导细菌种间通讯及参与细菌生长调控。在伴放线聚集杆菌(Aggregatibacter actinomycetemcomitans)中,RbsB被视作AI-2信号分子的受体,参与对密度感应系统(QS系统)的调控,胸膜肺炎放线杆菌的RbsB2还可结合哈维弧菌BB170培养物上清液中的AI-2信号分子。此外,rbsB基因与生物被膜的形成和在铁限制条件下生长的能力也有一定的关系[13-16]。
近年来,哈维弧菌(Vibrio harveyi)是中国南方沿海地区水产养殖业中最主要的病原菌之一,感染对象广泛,包括海洋鱼类[17-19]、甲壳类[20]、软体动物[21]和珊瑚[22]等,给沿海养殖户带来重大经济损失。为探究RbsB在哈维弧菌中的功能,本研究以课题组前期测序得到的哈维弧菌rbsB片段序列为基础,克隆哈维弧菌rbsB基因全长,并对其氨基酸序列进行预测分析结构功能,多序列比对比较与其他弧菌RbsB氨基酸差异,构建原核表达载体并对其进行诱导表达获得重组蛋白,为进一步研究其致病性及疫苗制备奠定基础。
1. 材料与方法
1.1 菌株来源与培养
哈维弧菌354菌株为中国水产科学研究院南海水产研究所渔业生物病害防治研究室于2013年10月从深圳南澳患病珍珠龙趸 [鞍带石斑鱼(Epinephelus lanceolatus)♂×棕点石斑鱼(E.fuscoguttatus)♀] 体内分离保种[23]。接种于LBS液体培养基(酵母提取粉0.5%,胰蛋白胨1%,NaCl 3%),28 ℃、140 r·min–1振荡培养16 h。
1.2 rbsB基因的同源扩增
根据前期对哈维弧菌354菌株进行全基因组测序所得的rbsB基因序列,设计1对引物,引物两端分别加限制性酶BamH I和Xho I酶切位点(由上海英潍捷基合成),正向引物:5′ GATCTGGTTCCGCGTGGATCCATGAAAAAACTCGCAACTCTTATC 3′,反向引物:5′ GTCACGATGCGGCCGCTCGAGCTTAGTCACGACTTTAAGCGGAA 3′。PCR反应为50 μL体系,反应条件为95 ℃预变性5 min;95 ℃变性30 s,55 ℃退火40 s,72 ℃延伸1 min,共35个循环;72 ℃延伸10 min,4 ℃保存。扩增产物经1.2%琼脂糖凝胶电泳检测后,切胶回收并连接转化,挑取阳性菌送上海英潍捷基生物技术有限公司测序。
1.3 rbsB基因序列分析
用软件ApE分析哈维弧菌rbsB基因序列及其开放阅读框,并将翻译后的氨基酸序列应用NCBI中的BLAST程序(https://blast.ncbi.nlm.nih.gov/)进行同源性检索;应用SMART及ExPaSy-Prosite对RbsB蛋白进行功能位点和功能域的预测;在线工具SignalP 4.0 Server及TMHMM 2.0预测蛋白的信号肽和跨膜结构域;在线(http://www.cbs.dtu.dk/services/NetPhos/)对RbsB氨基酸序列进行磷酸化位点分析;应用ExPASy的ProtParam程序对RbsB氨基酸序列进行氨基酸数量、分子量、等电点等基本理化性质分析;ExPaSy-SOPMA分析蛋白质的二级结构,对蛋白质三级结构采用SWISS-MODEL进行预测。用Clustal W 2.0进行多重序列比对,用MAGA 7.0构建NJ系统进化树。
1.4 重组质粒的构建
按照质粒DNA提取试剂盒说明提取rbsB-DH5α和pGEX-4t-1两种质粒,构建pGEX-rbsB重组质粒,随后将重组质粒转化入大肠杆菌BL21(DE3)感受态中,阳性菌以BamH I、Xho I双酶切后进行PCR反应,经1.2%琼脂糖凝胶电泳检测后,送上海英潍捷基生物技术有限公司测序以验证插入序列的正确性。
1.5 重组蛋白表达条件优化
将测序验证结果正确的阳性菌菌液按1∶100的比例接种到含100 μg·mL–1Amp的LB液体培养基中,37 ℃ 、180 r·min–1振荡培养至OD600为0.4~0.6时,加入异丙基硫代半乳糖甘(IPTG)诱导表达,IPTG的浓度分别为0、0.2 mmol·L–1、0.4 mmol·L–1、0.6 mmol·L–1、0.8 mmol·L–1和1 mmol·L–1,在30 ℃和37 ℃ 2个温度下180 r·min–1继续诱导培养8 h,分别收集诱导(0 h,2 h,4 h,6 h,8 h)菌体2 mL,菌液4 ℃离心5 min后弃上清,加入50 μL 5×SDS-PAGE Loading buffer和200 μL 1×PBS悬浮菌体,100 ℃煮沸10 min,4 ℃离心3 min后进行SDS-PAGE电泳,考马斯亮蓝R-250过夜染色后进行分析。同时进行Western blot,以Anti GST-Tag Rabbit HRP为抗体,最后用HRP-DAB (辣根过氧化物酶)底物显色试剂盒进行显色处理。
1.6 铁离子对rbs基因表达的影响
将哈维弧菌354接种于T1N1 (胰蛋白胨1%,酵母提取粉0.5%,NaCl 1%)培养基中140 r·min–1振荡过夜培养,按1∶1 000的比例重新接种于T1N1培养基中,实验组额外加入终浓度为7 mmol·L–1的柠檬酸铁,以加入菌液后的时间记为第0小时,分别培养4 h、8 h、12 h、24 h、48 h后按照RNA提取试剂盒说明提取RNA,根据逆转录试剂盒的说明合成cDNA用于qRT-PCR。采用qRT-PCR检测基因表达,设计1对qRT-PCR引物(正向引物:5′ GTGCAGAAGCGAAAGCGAAA 3′,反向引物:5′ GCACCACGGTCTAGTGTCAA 3′),以哈维弧菌16S rDNA作为内参基因(正向引物:5′ CCTACGGGAGGCAGCAG 3′,反向引物:5′ ATTACCGCGGCTGCTGG 3′)。反应体系为10 μL,反应条件为94 ℃预变性1 min;94 ℃变性10 s,60 ℃退火延伸40 s,共40个循环。通过熔解曲线和琼脂糖凝胶电泳共同验证PCR产物的特异性。同时扩增目的基因rbsB与内参基因16S rDNA,以各时间段的16S rDNA的表达量做△Ct校正样,以对照组培养第4小时的Ct值为△△Ct校正样,第0小时重新接种后的培养液数据作0处理,设为坐标原点,使用2–△△Ct的方法[24]计算rbsB基因不同时间段的相对拷贝数,作图分析。
2. 结果
2.1 rbsB全长核苷酸序列及其推导的氨基酸序列分析
克隆得到rbsB基因(GenBank序列号MF797015)测序后经DNAstar分析,得到开放阅读框(ORF)大小为879 bp (图1),G+C含量为49%。该序列可编码292个氨基酸,分子量为30.7 kD,理论等电点为5.05,在1~23 aa处有信号区域,N-末端为Met。
![]() 图 1 哈维弧菌rbsB基因序列及氨基酸序列起始密码子(ATG)和终止密码子(TAA)大写标明,磷酸位点加粗并用下划线标出;以“<”和“>”标出Peripla_BP_4结构域的起始和终止位置Figure 1. Gene and amino acid sequence of rbsB gene from V.harveyiThe start and stop codons are capitalized; phosphate sites are bold and underlined; the start and end positions of the Peripla_BP_4 domain are identified by "<" and ">".
图 1 哈维弧菌rbsB基因序列及氨基酸序列起始密码子(ATG)和终止密码子(TAA)大写标明,磷酸位点加粗并用下划线标出;以“<”和“>”标出Peripla_BP_4结构域的起始和终止位置Figure 1. Gene and amino acid sequence of rbsB gene from V.harveyiThe start and stop codons are capitalized; phosphate sites are bold and underlined; the start and end positions of the Peripla_BP_4 domain are identified by "<" and ">".2.2 RbsB蛋白理化性质分析
RbsB原子总数为4 362个,分子式为C1338H2215N371O428S10,不稳定系数为19.36,属于稳定蛋白。脂溶系数为102.26,亲水性系数为0.043。分析氨基酸组成可知RbsB蛋白含酸性氨基酸38个,碱性氨基酸30个,极性氨基酸224个。
2.3 RbsB结构分析和功能预测
TargetP 1.1 Server预测结果表明RbsB蛋白无线粒体目标肽,但有一分泌途径信号肽。采用Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)中的Gram-negative protein程序进行亚细胞定位,推测RbsB蛋白最可能定位于细胞质中,为周质蛋白。
在Swiss-Prot和TrEMB的模式序列进行搜索没有得到与已知序列匹配的功能位点。InterProscan结合Smart分析RbsB蛋白的结构功能域,结果表明RbsB蛋白中27~278 aa存在Peripla_BP_4结构域(图1)。采用TMHMM 2.0 Server分析RbsB无跨膜区域,为非跨膜蛋白。
RbsB蛋白共含有23个蛋白质磷酸化位点,包括9个苏氨酸位点、12个丝氨酸位点、2个酪氨酸位点(图1)。RbsB蛋白的二级结构主要由4种形式组成,其中α-螺旋占43.84%,无规则卷曲占22.60%,延伸链占21.58%,β-转角占11.99%,三维结构见图2。由此推测,RbsB蛋白主要在运载结合中发挥作用。
2.4 RbsB多重序列比对和系统进化树构建
将获得的哈维弧菌RbsB氨基酸序列与其他弧菌RbsB氨基酸序列(表1)应用Clustal W 2.0进行多重序列比对(图3)。根据比对情况利用MEGA 7.0构建NJ系统进化树(图4),结果显示哈维弧菌和欧文弧菌(V.owensii CAIM 1854)关系最近,与远青弧菌(V.azureus NBRC 104587)关系较远。
表 1 哈维弧菌354菌株的RbsB与其他细菌的氨基酸同源性分析Table 1. Homology analysis of amino acids of RbsB from V.harveyi strain 354 and other bacterial species菌种
species同源性/%
homology注册号
accession No.哈维弧菌 (V.harveyi ATCC 33843) 100 AIV07307.1 欧文弧菌 (V. owensii CAIM 1854) 100 KIF46393.1 坎氏弧菌 (V.campbellii ATCC BAA-1116) 98 AGU97393.1 轮虫弧菌 (V.rotiferianus HM-10) 98 OHY94262.1 溶藻弧菌 (V.alginolyticus ATCC 17749) 97 GAD69766.1 副溶血弧菌 (V.parahaemolyticus ATCC 17802) 97 KKI09273.1 需钠弧菌 (V.natriegens ATCC 14048) 97 EPM40141.1 贻贝弧菌 (V.mytili CAIM 528) 96 KIN10841.1 创伤弧菌 (V.vulnificus ATCC 33147) 92 KGK71697.1 远青弧菌 (V.azureus NBRC 104587) 92 GAD74752.1 ![]() 图 3 哈维弧菌RbsB氨基酸序列与其他已知菌种的比对分析“*”表示所有序列中都一致的氨基酸;“:”表示保守的氨基酸;“.”表示半保守的氨基酸,方框内为Peripla_BP_4结构域Figure 3. Sequence alignment of predicted RbsB amino acid from V.harveyi and other known bacterial species“*” means that the residues or nucleotides are identical in all sequences in the alignment; “:” indicates conserved substitution; “.” indicates semi-conserved substitution; Peripla_BP_4 domain is in box.
图 3 哈维弧菌RbsB氨基酸序列与其他已知菌种的比对分析“*”表示所有序列中都一致的氨基酸;“:”表示保守的氨基酸;“.”表示半保守的氨基酸,方框内为Peripla_BP_4结构域Figure 3. Sequence alignment of predicted RbsB amino acid from V.harveyi and other known bacterial species“*” means that the residues or nucleotides are identical in all sequences in the alignment; “:” indicates conserved substitution; “.” indicates semi-conserved substitution; Peripla_BP_4 domain is in box.![]() 图 4 利用Clustal W 2.0和MEGA 7.0构建的rbsB基因的氨基酸序列系统进化树Figure 4. Phylogenetic tree of amino sequence of rbsB gene by Clustal W 2.0 and MEGA 7.0
图 4 利用Clustal W 2.0和MEGA 7.0构建的rbsB基因的氨基酸序列系统进化树Figure 4. Phylogenetic tree of amino sequence of rbsB gene by Clustal W 2.0 and MEGA 7.02.5 重组表达载体的构建与鉴定
将PCR扩增产物和原核表达pGEX-4t-1质粒经XhoI、BamH I双酶切后构建重组质粒pGEX-4t-1-rbsB,并将其转化到大肠埃希菌BL21 (DE3)感受态中,经IPTG诱导后进行SDS-PAGE电泳,观察到诱导组在59 kD处有一条带。Western bolt结果表明重组蛋白诱导表达成功(图5)。37 ℃条件下诱导8 h表达量最高(图6),IPTG浓度对蛋白的表达量没有影响。对载体pGEX-4t-1-rbsB菌液测序后,与NCBI中V.harveyi CAIM 1792的rbsB基因序列进行比对,序列一致。
![]() 图 6 温度及时间对诱导RbsB蛋白的表达量的影响M. Marker;1. pGEX-4t-1菌液;2. 诱导0 h后菌液;3~5. 30 ℃诱导4 h、6 h、8 h后菌液;6~8. 37 ℃诱导4 h、6 h、8 h后菌液Figure 6. Effect of temperature and time on expression of RbsB proteinM. Marker; 1. pGEX-4t-1 plasmid; 2. 0 h after IPTG induction; 3–5. 4 h, 6 h and 8 h after IPTG induction at 30 ℃, respectively; 6–8. 4 h, 6 h and 8 h after IPTG induction at 37 ℃, respectively
图 6 温度及时间对诱导RbsB蛋白的表达量的影响M. Marker;1. pGEX-4t-1菌液;2. 诱导0 h后菌液;3~5. 30 ℃诱导4 h、6 h、8 h后菌液;6~8. 37 ℃诱导4 h、6 h、8 h后菌液Figure 6. Effect of temperature and time on expression of RbsB proteinM. Marker; 1. pGEX-4t-1 plasmid; 2. 0 h after IPTG induction; 3–5. 4 h, 6 h and 8 h after IPTG induction at 30 ℃, respectively; 6–8. 4 h, 6 h and 8 h after IPTG induction at 37 ℃, respectively2.6 铁离子对rbsB基因表达的影响
用qRT-PCR对rbsB在富铁培养条件下的表达量进行分析(图7),结果显示rbsB的表达具有明显的差异性。在限铁培养条件下,rbsB表达量在培养第4小时达到最高(6.22),随着培养时间的增加表达量逐步降低,第48小时为0.35;富铁培养条件下则相反,第4小时表达量仅为0.45,培养第48小时rbsB的表达量为5.48。
3. 讨论
在革兰氏阴性菌中,周质结合蛋白溶解在细胞周质中,底物进入周质后与周质结合蛋白可逆性结合,从而通过构象上的改变,进一步与膜结合蛋白相互作用,最终使底物进入胞浆发挥功能作用[25]。RbsB作为周质结合蛋白,被认为可能参与细菌种间通讯及细菌生长调控,在细菌毒力因子的表达中发挥重要作用。
本研究首次克隆出哈维弧菌rbsB基因全长,二级结构预测含有α-螺旋、无规则卷曲、延伸链、β-转角4种结构。SDS-PAGE和Western blot检测结果表明,Vh-RbsB蛋白与已报道的豌豆根瘤菌(Rhizobium leguminosarum)[16]、伴放线聚集杆菌[26]的RbsB蛋白分子量相似,为非分泌型蛋白,均含有N-端信号肽,亚细胞定位为周质蛋白。具核梭杆菌(Fusobacterium nucleatum)中另一种AI-2信号分子D-半乳糖结合蛋白Gbp也显示出与RbsB序列的相似性[27]。然而豌豆根瘤菌RbsB蛋白与哈维弧菌LuxP蛋白共享结构域,其N-端信号肽可被I型分泌系统所识别,研究比对哈维弧菌354株RbsB与LuxP的结构域后,并未发现如上特征,这可能是不同种属间RbsB氨基酸序列差异所导致的功能差异。
结合Pfam蛋白数据库和SMART分析发现Vh-RbsB蛋白包含Peripla_BP_4结构域,Peripla_BP_4结构域所属的第2周质结合蛋白家族簇中包含目前为止发现的2种具有AI-2结合能力的周质结合蛋白AI-2受体,分别是哈维弧菌的LuxP型受体和鼠伤寒沙门菌的LsrB型受体[28-29]。在非流行性流感嗜血杆菌(Nontypeable Haemophilus influenzae,NTHI)中,rbsB编码的蛋白对NTHI AI-2信号系统起着至关重要的作用,可促进体外生物膜的形成及慢性感染,rbsB的失活显著降低NTHI对AI-2和AI-2前体二羟基戊二酮(dihydroxypentanedione,DPD)的吸收,被视作AI-2的调节基因[30]。胸膜肺炎放线杆菌中RbsB与哈维弧菌的LuxP有27%相似性,能够结合哈维弧菌AI-2分子[15],James等[14]、Shao等[26]实验也表明在限铁培养条件下伴放线聚集杆菌中rbsB缺失株与luxS缺失株表型类似。以上结果显示RbsB蛋白在部分细菌中起到结合AI-2的功能。因此推测在哈维弧菌354株中,RbsB蛋白可能与密度感应系统有关,为一种潜在的AI-2受体,与LuxP协同或竞争性结合AI-2信号分子,参与信号结合及对密度感应系统的调控。
RbsB蛋白同AI-2的活性密切相关,在铁限制培养条件下rbsB基因表达量的变化同细菌的生长及AI-2的活性有明显的联系。本文通过qRT-PCR分析哈维弧菌354株在富铁与限铁条件下rbsB基因表达量的变化,发现限铁条件下rbsB表达量在对数生长早期就达到最高值,而后持续下降,在培养第24小时达到最低;富铁条件下rbsB表达量在对数生长后期(第12小时)达到最低,而后显著上升,在培养第48小时达到最高。James等[14]证实在限铁条件下,伴放线聚集杆菌rbsB缺失株生长水平仅为野生株的30%,rbsB的失活影响着铁吸收基因的表达。鳗弧菌与嗜水气单胞菌在对数生长前期6 h就表现出明显AI-2活性[31-32]。这些研究结果意味着Vh-RbsB在限铁条件下可能在生长早期就发挥其生理学功能,富铁条件下Vh-RbsB在高细胞密度下即在生长的稳定期发挥功能。由于本研究并未同步构建rbsB基因缺失株及回补株,并对其他铁摄取和代谢相关的基因进行表达差异的分析,结果具有部分局限性。
对于一些细菌而言,通过在培养液中添加核糖,RbsB介导抑制哈维弧菌发光是可逆的,说明核糖与AI-2分子共同竞争RbsB结合位点,核糖可影响AI-2的活性及生物膜的形成[15,26];然而对于某些细菌,AI-2前体DPD的摄取并未受到核糖或其他戊糖的竞争性抑制[30]。本研究成功构建pGEX-4t-1-rbsB重组质粒并转化至大肠埃希菌BL21 (DE3)中,得到重组蛋白的相对分子量约为59 kD,IPTG浓度不影响重组蛋白的表达。本研究可为研制哈维弧菌亚单位疫苗提供理论基础。
-
![]()
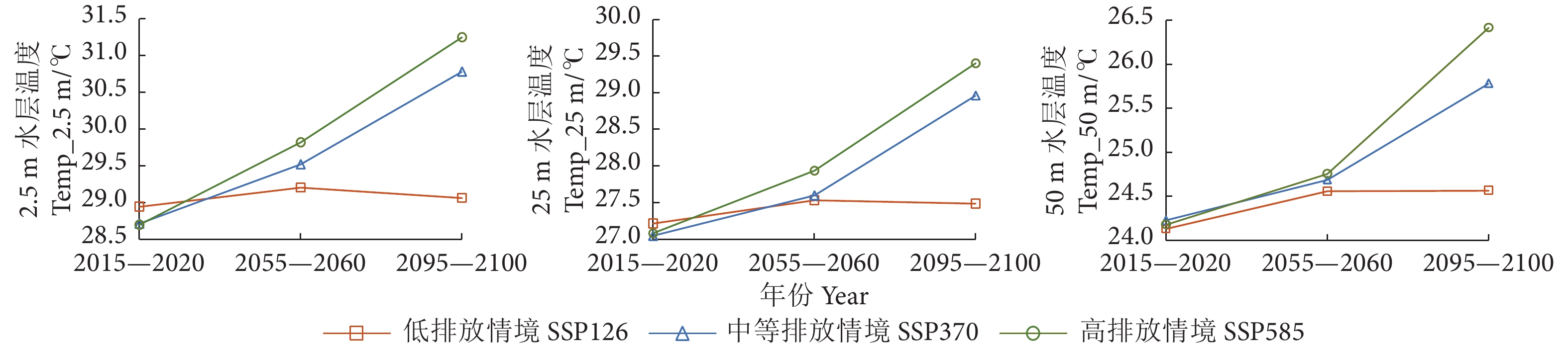
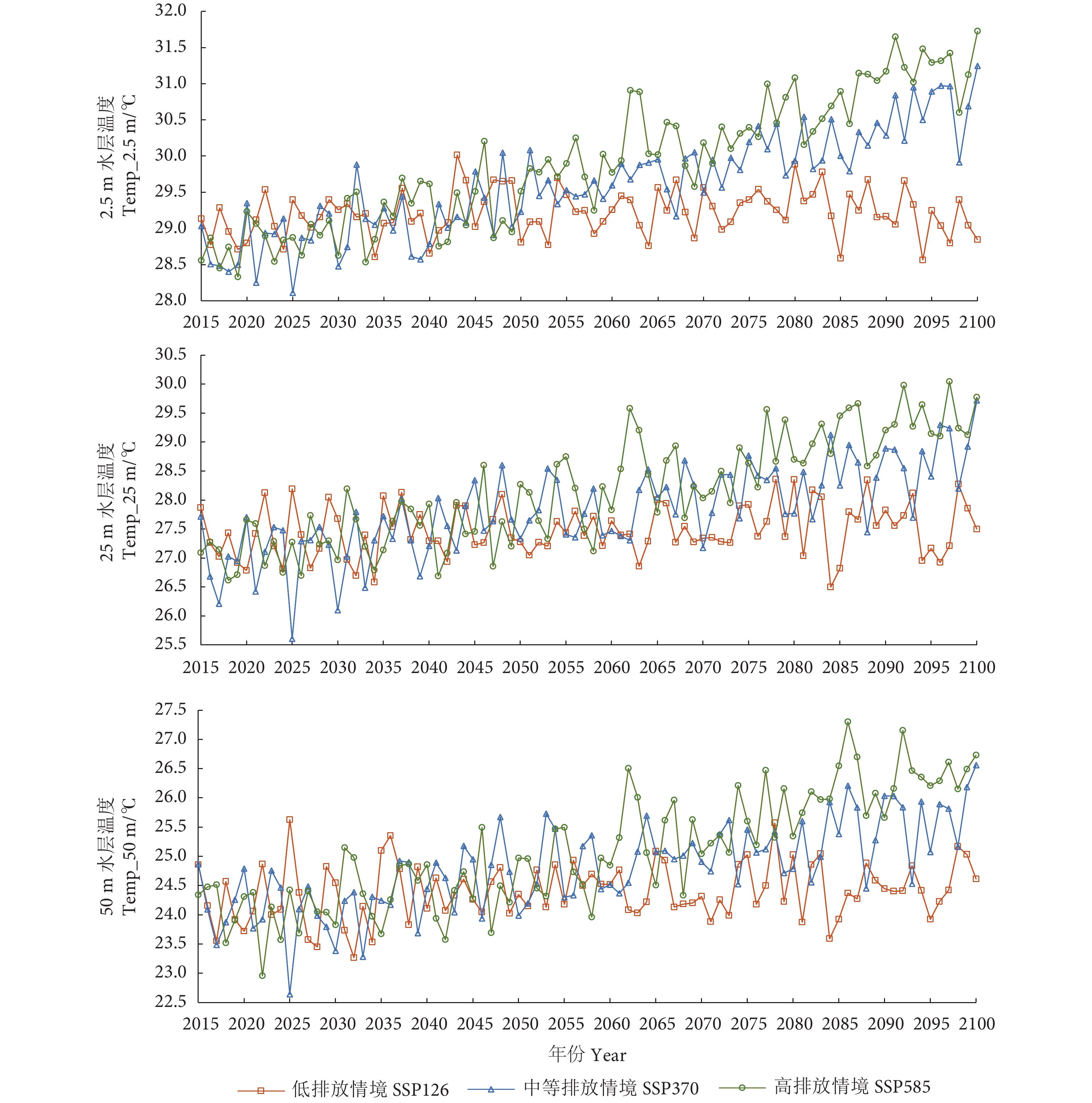
图 1 不同气候情境下2015—2100年鲐鱼渔场内不同水层水温的年际变化
Figure 1. Interannual variation of water temperature at different depths in fishing ground of S. japonicus during 2015−2100 under different climatic conditions
![]()
图 2 不同气候情境下2015—2020、2055—2060和2095—2100年内鲐鱼渔场内不同水层水温的平均值
Figure 2. Average water temperature at different depths in fishing ground of S. japonicus during 2015−2020, 2055−2060 and 2095−2100 under different climatic conditions
![]()
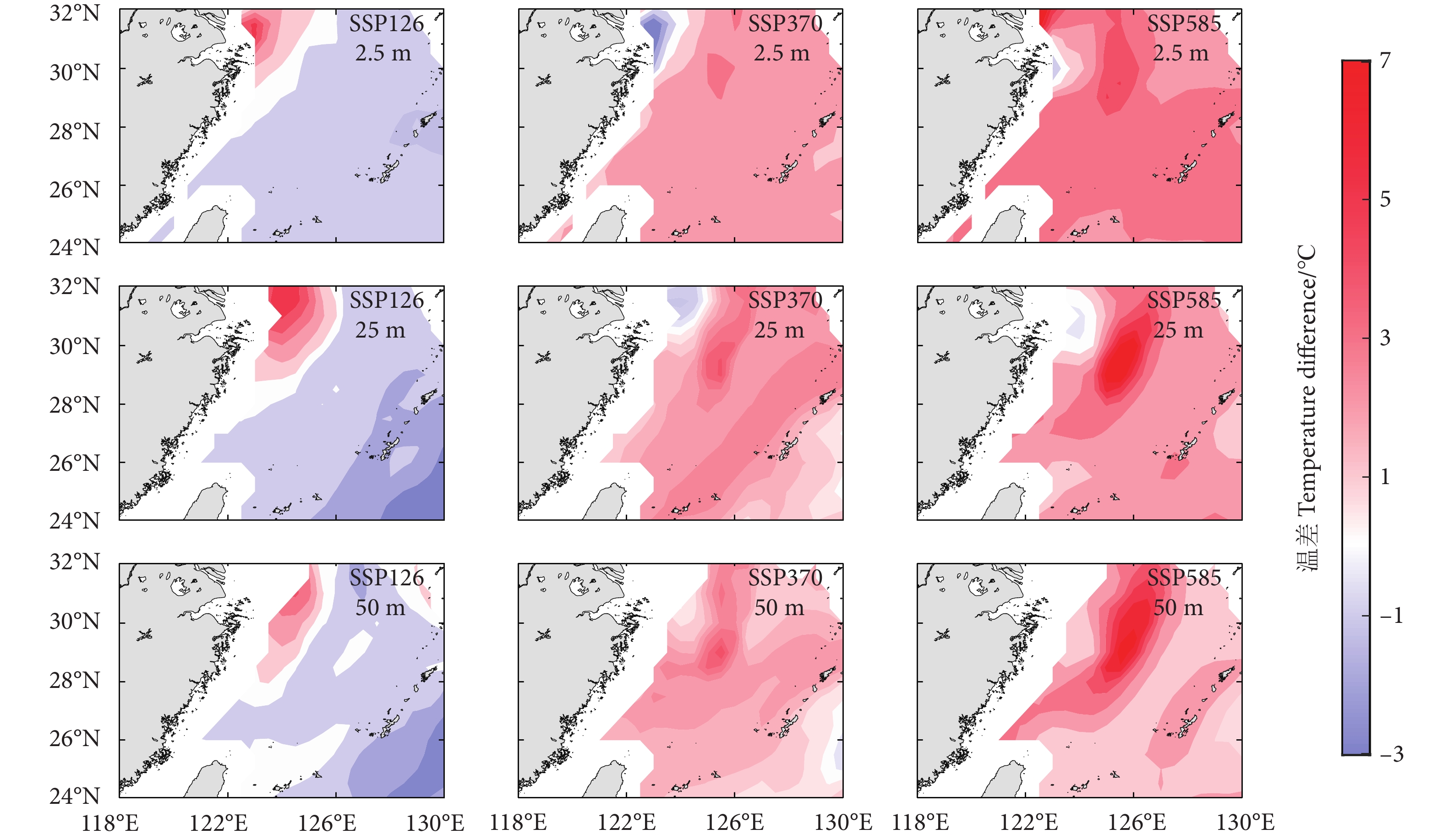
图 3 不同气候情境下2100和2015年鲐鱼渔场内不同水层水温差值的空间分布
Figure 3. Spatial distribution of water temperature difference at different depths in fishing ground of S. japonicus in 2100 and 2015 under different climate conditions
![]()
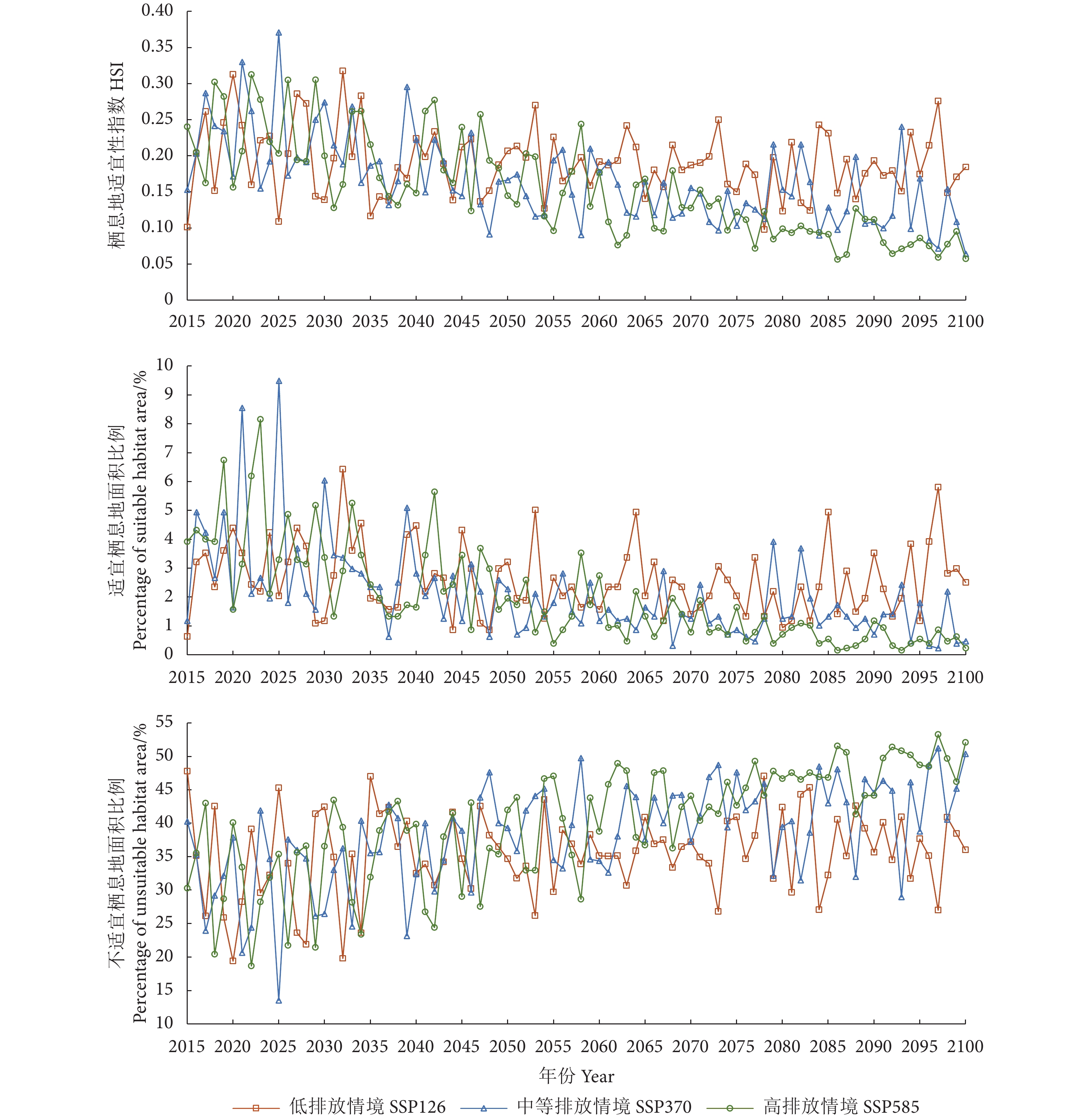
图 4 不同气候情境下2015—2100年东海鲐鱼平均栖息地指数、适宜栖息地及不适宜栖息地面积比例
Figure 4. Interannual variation of average habitat suitability index, percentages of suitable and unsuitable habitat area of S. japonicus in East China Sea under different climatic conditions during 2015−2100
![]()
图 5 不同气候情境下东海鲐鱼栖息地指数在2015—2020、2055—2060和2095—2100年的空间分布
Figure 5. Spatial distribution of habitat suitability index of S. japonicus in East China Sea during 2015−2020, 2055−2060 and 2095−2100 under different climatic conditions
![]()
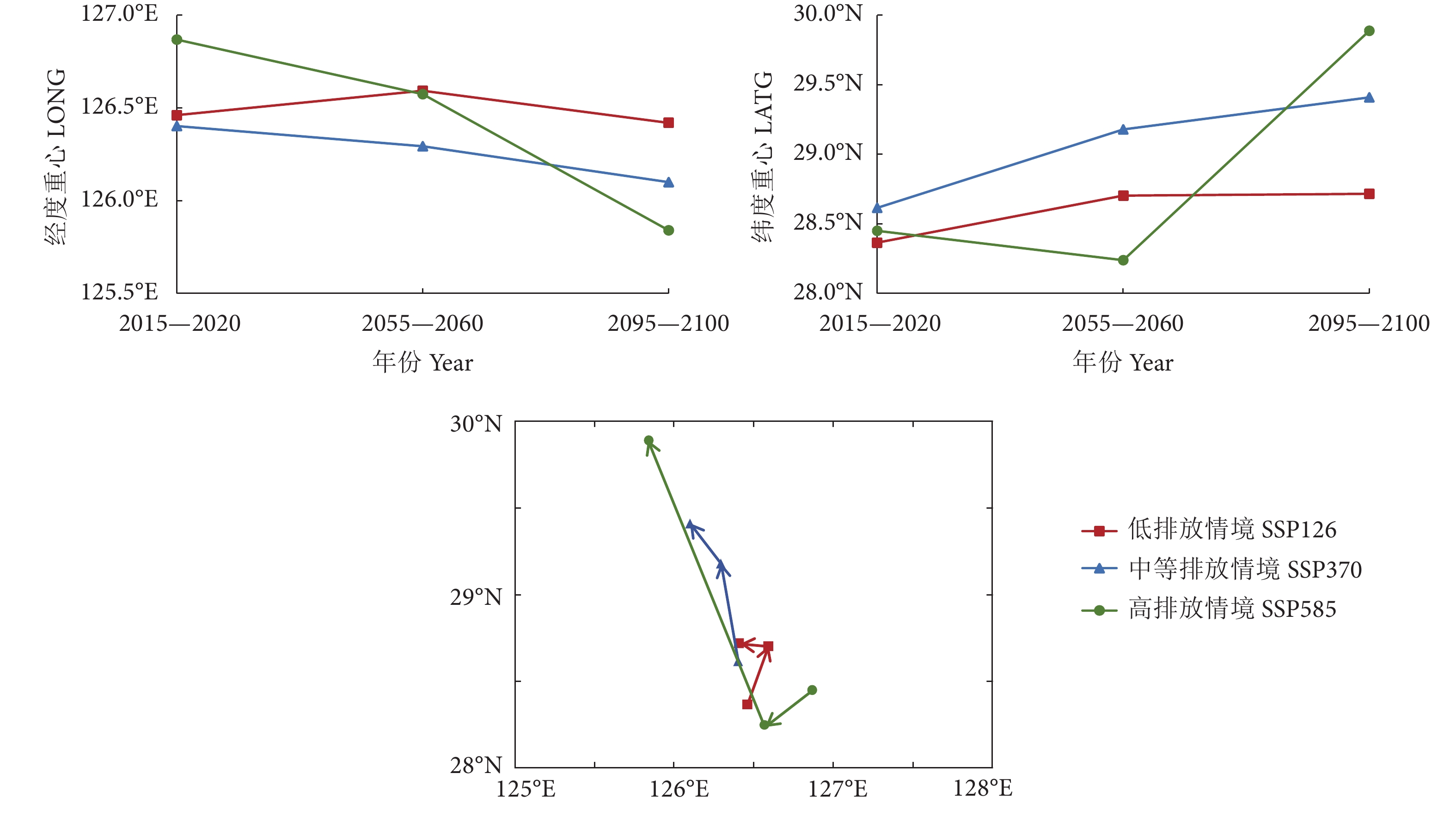
图 6 不同气候情境下2015—2020、2055—2060和2095—2100年时间段内东海鲐鱼适宜栖息地经纬度重心及其空间分布图
Figure 6. Longitude and latitude center of gravity and its spatial distribution of suitable habitat of S. japonicus in East China Sea during 2015−2020, 2055−2060 and 2095−2100 under different climatic conditions
![]()
图 7 不同气候情境下2015—2020、2055—2060和2095—2100年内各环境因子适宜性指数 (SI≥0.6) 重心
Figure 7. Suitability index (SI≥0.6) center for each environmental factor during 2015−2020, 2055−2060 and 2095−2100 under different climatic conditions
表 1 不同气候情境的不同时间段内东海鲐鱼正常与不适宜栖息地面积变化
Table 1 Changes of normal and unsuitable habitat area of S. japonicus in East China Sea in different time periods under different climatic conditions
模态SSP 年份Year 渔场正常栖息地面积HS/% 面积增减ID_HS/% 渔场不适宜栖息地面积UNSH/% 面积增减ID_UNSH/% SSP126 2015—2020 24.38 — 32.80 — 2055—2060 21.70 −10.99 35.48 +8.17 2095—2100 21.33 −12.51 35.84 +9.27 SSP370 2015—2020 24.07 — 33.11 — 2055—2060 19.48 −19.07 37.70 +13.86 2095—2100 11.39 −52.68 45.79 +38.29 SSP585 2015—2020 24.18 — 32.99 — 2055—2060 18.16 −24.89 39.02 +18.28 2095—2100 7.45 −69.19 49.73 +50.74 注:HS代表渔场正常栖息地面积;ID_SH代表2055—2060和2095—2100年渔场正常栖息地面积相对于2015—2020年正常栖息地面积的增减;UNSH代表渔场不适宜栖息地面积;ID_UNSH代表2055—2060和2095—2100年渔场不适宜栖息地面积相对于2015—2020年不适宜栖息地面积的增减。 Note: HS represents the area of normal habitat of fishing ground; ID_SH represents the increase or decrease of the normal habitat area of the fishing ground during 2055–2060 and 2095–2100 compared with the normal habitat area during 2015–2020; UNSH represents the area of unsuitable habitat of fishing ground; ID_UNSH represents the increase or decrease of the area of unsuitable habitat of fishing ground during 2055–2060 and 2095–2100 compared with that during 2015–2020.  下载: 导出CSV
下载: 导出CSV
-
[1] 李曰嵩, 邢宇娜, 潘灵芝, 等. 鲐鱼生活史及模型应用研究进展[J]. 大连海洋大学学报, 2021, 36(4): 694-705. doi: 10.16535/j.cnki.dlhyxb.2020-227 [2] 刘雅丹. 浅谈气候变化对渔业和水产养殖的影响[J]. 中国水产, 2022(1): 68-71. [3] GENNER M J, SIMS D W, WEARMOUTH V J, et al. Regional climatic warming drives long-term community changes of British marine fish[J]. Proc R Soc B, 2004, 271(1539): 655-661. doi: 10.1098/rspb.2003.2651
[4] PERRY A L, LOW P J, ELLIS J R, et al. Climate change and distribution shifts in marine fishes[J]. Science, 2005, 308(5730): 1912-1915. doi: 10.1126/science.1111322
[5] 周天军, 邹立维, 陈晓龙. 第六次国际耦合模式比较计划 (CMIP6) 评述[J]. 气候变化研究进展, 2019, 15(5): 445-456. [6] KIPARISSIS S, TSERPES G, TSIMENIDIS N. Aspects on the demography of chub mackerel (Scomber japonicus Houttuyn, 1782) in the Hellenic Seas[J]. Belg J Zool. 2000, 130: 5-9.
[7] 范秀梅, 杨胜龙, 张胜茂, 等. 基于栖息地指数的阿拉伯海鲐鱼渔情预报模型构建[J]. 南方水产科学, 2020, 16(4): 8-17. doi: 10.12131/20190255 [8] 李纲, 陈新军. 东海鲐鱼资源和渔场时空分布特征的研究[J]. 中国海洋大学学报 (自然科学版), 2007(6): 921-926. [9] 郭爱, 余为, 陈新军, 等. 中国近海鲐鱼资源时空分布与海洋净初级生产力的关系研究[J]. 海洋学报, 2018, 40(8): 42-52. [10] 李宜锴, 方星楠, 余为, 等. 2005—2016年中国东海鲐鱼渔场的时空分布及与海表面温度的关联[J]. 上海海洋大学学报, 2022, 31(3): 710-720. doi: 10.12024/jsou.20210503429 [11] 郭爱. 气候与海洋环境变化对东黄海鲐鱼栖息地时空变动的影响[D]. 上海: 上海海洋大学, 2020: 2-4. [12] 杨胜龙, 范秀梅, 伍玉梅, 等. 基于GAM模型的阿拉伯海鲐鱼渔场分布与环境关系[J]. 生态学杂志, 2019, 38(8): 2466-2470. doi: 10.13292/j.1000-4890.201908.032 [13] 李曰嵩. 东海鲐鱼 (Scomber japonica) 早期生活史过程的生态动力学模拟研究[D]. 上海: 上海海洋大学, 2012: 41-42. [14] YU W, WEN J, CHEN X J, et al. Effects of climate variability on habitat range and distribution of chub mackerel in the East China Sea[J]. J Ocean Univ China, 2021, 20(6): 1483-1494. doi: 10.1007/s11802-021-4760-x
[15] 刘红红, 朱玉贵. 气候变化对海洋渔业的影响与对策研究[J]. 现代农业科技, 2019(10): 244-247. doi: 10.3969/j.issn.1007-5739.2019.10.149 [16] CHEUNG W W L, LAM V W Y, SARMIENTO J L, et al. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change[J]. Glob Chang Biol, 2010, 16(1): 24-35. doi: 10.1111/j.1365-2486.2009.01995.x
[17] 苏杭, 陈新军, 汪金涛. 海表水温变动对东、黄海鲐鱼栖息地分布的影响[J]. 海洋学报, 2015, 37(6): 88-96. [18] 张孝威. 鲐鱼[M]. 北京: 农业出版社, 1983: 42-46. [19] SHULTZ A D, ZUCKERMAN Z C, TEWART H A, et al. Seasonal blood chemistry response of sub-tropical nearshore fishes to climate change[J]. Conserv Physiol, 2014, 2(1): 1-12.
[20] YASUDA T, NAGANO N, KITANO H. Diel vertical migration of chub mackerel: preliminary evidence from a biologging study[J]. Mar Ecol Prog Ser, 2018, 598: 147-151. doi: 10.3354/meps12636
[21] CHEN, X J, TIAN S Q, GUAN W J. Variations of oceanic fronts and their influence on the fishing grounds of Ommastrephes bartramii in the Northwest Pacific[J]. Acta Oceanol Sin, 2014, 33(4): 45-54. doi: 10.1007/s13131-014-0452-3
[22] 易炜, 郭爱, 陈新军. 不同环境因子权重对东海鲐鱼栖息地模型的影响研究[J]. 海洋学报, 2017, 39(12): 90-97. [23] 宋利明, 许回, 陈明锐. 毛里塔尼亚海域日本鲭时空分布与海洋环境的关系[J]. 上海海洋大学学报, 2020, 29(6): 868-877. doi: 10.12024/jsou.20190702746 [24] 官文江, 陈新军, 高峰, 等. 海洋环境对东、黄海鲐鱼灯光围网捕捞效率的影响[J]. 中国水产科学, 2009, 16(6): 949-958. doi: 10.3321/j.issn:1005-8737.2009.06.016 [25] 王子鸣. 基于气候变化分析对海洋的影响[J]. 湖北农机化, 2020(5): 25. [26] 何越. 全球变暖背景下热带太平洋及中国近海气候变化动力降尺度预估[D]. 厦门: 厦门大学, 2017: 64-65. [27] STOUFFER R J, BROCCOLI A J, DELWORTH T L, et al. GFDL's CM2 Global Coupled Climate Models. Part IV: idealized climate response[J]. J Clim, 2015, 19(5): 723-740.
[28] LEE H C. Impact of atmospheric CO2 doubling on the North Pacific subtropical mode water[J]. Geophys Res Lett, 2009, 36(6): 295-311.
[29] 官文江. 基于海洋遥感的东、黄海鲐鱼渔场与资源研究[D]. 上海: 华东师范大学, 2008: 14-15. [30] 马超, 庄之栋, 刘勇, 等. 西北太平洋公海灯光敷网渔获组成及主要种类渔业生物学特征研究[J]. 渔业研究, 2018, 40(2): 141-147. [31] SASSA C, KAWAGUCHI K, TAKI K. Larval mesopelagic fish assemblages in the Kuroshio-Oyashio transition region of the western North Pacific[J]. Mar Biol, 2007, 150(6): 1403-1415. doi: 10.1007/s00227-006-0434-x
[32] YATSU A, SASSA C, MOKU M, et al. Night-time vertical distribution and abundance of small epipelagic and mesopelagic fishes in the upper 100 m layer of the Kuroshio-Oyashio Transition Zone in spring[J]. Fish Sci, 2005, 71(6): 1280-1286. doi: 10.1111/j.1444-2906.2005.01094.x
[33] TIAN Y, UENO Y, SUDA M, et al. Decadal variability in the abundance of Pacific saury and its response to climatic/oceanic regime shifts in the northwestern subtropical Pacific during the last half century[J]. J Mar Syst, 2004, 52: 235-257. doi: 10.1016/j.jmarsys.2004.04.004
[34] DING X, HU B, LI J, et al. Late Holocene Orbital forcing and solar activity on the Kuroshio Current of subtropical North Pacific at different timescales[J]. Front Earth Sci, 2022, 10: 1-11.
[35] 龚彩霞. 未来气候变化情景下西北太平洋柔鱼资源变动[D]. 上海: 上海海洋大学, 2020: 69-70. -
期刊类型引用(1)
1. 薛尊,石建高,张文阳. 中国重力式深水网箱研究进展和展望. 渔业研究. 2024(04): 393-402 .  百度学术
百度学术
其他类型引用(1)









计量
- 文章访问数: 390
- HTML全文浏览量: 160
- PDF下载量: 51
- 被引次数: 2
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
