
Analysis of solid-state anaerobic fermentation process of mackerel processing by-products by high-throughput sequencing
-
摘要: 为了实现易产组胺的鲭鱼 (Pneumatophorus japonicus) 加工副产物综合利用产业化技术开发,对鲭鱼加工副产物进行了不灭菌的乳杆菌固态厌氧发酵,尝试生产饲料用新型活性蛋白源。系统监测了发酵过程中组胺、酸溶蛋白、挥发性盐基氮 (Total volatile basic nitrogen, TVB-N)、pH的变化,通过高通量测序技术分析了发酵过程微生物群落的变化,对比分析了发酵前后抗原蛋白、游离氨基酸、脂肪酸成分的变化,以全面评价发酵过程中营养、活性成分和安全性的变迁。结果显示,植物乳杆菌 (Lactobacillus plantarum) HSCC-LP121在提高酸溶蛋白含量、降低组胺积累、减少挥发性盐基氮产生方面综合效果最好。发酵30 d时组胺降低到0.629 g·kg−1。这一数据和高通量测序结果表明,HSCC-LP121具有组胺降解能力,同时可抑制嗜冷杆菌属 (Psychrobacter) 和假单胞菌属 (Pseudomonas) 等潜在产组胺微生物的生长。该菌种还可抑制曲霉菌属 (Aspergillus) 等潜在危害真菌的生长,同时诱使一些生香酵母的生长。Abstract: In order to develop commercial technology for the comprehensive utilization of mackerel (Pneumatophorus japonicus) processing by-product, we carried out the non sterilized solid-state anaerobic fermentation of mackerel by-products to produce a novel protein source for feed with bioactivity. In addition, we monitored the content of histamine, acid-soluble protein and other indexes such as pH, total volatile basic nitrogen and antigenic protein; and investigated the evolution of microbial community during the fermentation by high-throughput sequencing technology, comparing the free amino acids and fatty acids before and after the fermentation. The results show that Lactobacillus plantarum HSCC-LP121 had the best comprehensive effect in increasing acid soluble protein content, reducing histamine accumulation and reducing volatile base nitrogen production. After 30 d of fermentation, the histamine content decreased to 0.629 g·kg−1, which indicates that HSCC-LP121 had the ability to degrade histamine. Moreover, HSCC-LP121 inhibited the growth of potential histamine-producing microorganisms such as Psychrobacter and Pseudomonas. Besides, the strain could also inhibit the growth of potentially harmful fungi such as Aspergillus, and induce the growth of some aroma-producing yeasts.
-
黑鲷(Acanthopagrus schiegelii)、真鲷(Pagrus major)分别为鲷科中棘鲷属和赤鲷属的名贵经济鱼类[1-2],分布于西北太平洋沿海,是我国南北沿海地区重要养殖品种[3-4]。其中黑鲷抗逆性强、适温适盐范围较广且味道鲜美,于我国南、北沿海均可养殖;真鲷个体大、生长快、体色艳丽,但适温范围较窄,主要为南方养殖。利用鲷科中这两种具不同经济特性鱼类进行的杂交选育实验或生产已有许多研究[5-10]。以黑鲷(♀)×真鲷(♂)为亲本的杂交子代(F1)首次在大规模养殖上获得成功;并且该杂交F1在表现出好于双亲本的抗逆性状的基础上,其个体1~3龄的性腺发育模式及繁殖性状与黑鲷母本相一致(另文发表)。在获得杂交F1成熟个体的基础上,开展杂交F1的自繁培育研究实验,为鱼类杂交育种研究提供更多参考依据。
仔鱼早期发育过程中,存在由内源营养过渡到摄食外源营养的混合营养期(mixed feeding stage),这一阶段也常被称为“临界期”,即由于饵料保障和仔鱼器官发育的共同作用常导致仔鱼大量死亡的危险期[11]。在苗种繁育过程中,摄食与营养对早期仔鱼培育至关重要。饥饿胁迫作为影响鱼类早期发育与存活的主要因素,是鱼类早期生活史研究的主要内容[12];相关的研究结果已为不同海水鱼类的苗种培育等提供了不少宝贵资料[13-15]。鱼类的繁育是否能成功并顺利生长发育,早期幼体的培育是关键环节,对于杂交选育尤其如此。本文通过对杂交F2早期仔鱼的发育、摄食习性及饥饿胁迫的不可逆点(point of no return,PNR)等的研究,探索杂交F2幼体期在这些方面的表现,不仅为进一步丰富杂交F2早期发育阶段的生物学基础数据,及其苗种繁育技术优化提供依据,也为鲷科鱼类的远缘杂交育种研究提供借鉴。
1. 材料与方法
1.1 初孵仔鱼来源与培育
实验在江苏省海水种苗繁育与增养殖技术中心进行。亲鱼为江苏省海洋水产研究所人工繁育、养成的黑鲷(♀)×真鲷(♂)杂交子一代3龄以上成鱼,体质量(733.2±172.42) g,体长(34.7±2.21) cm。于2017年4月取同一批次自然受精卵于水泥池网箱中进行孵化;孵化温度21.2~22.4 ℃,盐度24~25,pH约8.2,微充气,于第2天获得同步发育初孵仔鱼。
1.2 实验方法
1.2.1 仔鱼发育观察
仔鱼出膜后,每天定时取样,在SMZ18解剖镜(Nikon)下观察、拍照,测量仔鱼的全长、体高、卵黄囊直径等生长数据。卵黄囊体积V=1/6×πr2R,式中r为卵黄囊短径,R为卵黄囊长径;卵黄囊消耗率=(Vm–Vn)/V0×100%,式中V0为初孵仔鱼卵黄囊体积,m、n为孵化后的第m、第n天[16]。
1.2.2 饥饿与摄食的仔鱼生长比较
在仔鱼开口摄食前(2日龄),随机选取健康仔鱼分为饥饿组和正常投饵对照组,置于体积10 L的长方形有机玻璃水族箱中培育,每组仔鱼500尾。实验用水为经沉淀砂滤的自然海水,盐度25,水温保持在(21.8±0.4) ℃,微充气,上午、下午各换水1/3,每天记录、吸出沉底死亡仔鱼。每组各设有2个重复。正常投饵对照组投喂褶皱臂尾轮虫(Brachionus plicatilis),保持水中轮虫密度为5~10个·mL–1,每天9:00、15:00各投饵1次,投饵后2 h吸污1次并换水、添足水体;饥饿实验组不投饵,直至仔鱼100%死亡。每天定时取各组仔鱼10尾,经微量丁香酚麻醉,用SMZ18体视解剖镜观察、测量仔鱼的全长、体高及肛后体长,测量卵黄囊及油球直径等指标。仔鱼特定日增长率=(lnLm–lnLn)/(m–n)×100%,式中L为特定体全长(mm),m、n为孵化后的第m、第n天[16]。
1.2.3 初次摄食率测定
观察到仔鱼开口后(3日龄),每天定时从饥饿组中取30尾仔鱼移至1 000 mL同样海水条件的大烧杯中,投喂轮虫(密度为5~10个·mL–1),3 h后取出全部仔鱼,经微量丁香酚麻醉,用解剖镜拍照、测量仔鱼生物学指标等,逐尾解剖、观察仔鱼摄食情况,记录摄食轮虫的仔鱼数量,计算仔鱼初次摄食率。设2组重复。摄食率=(肠管内含有轮虫的仔鱼尾数/测定总仔鱼尾数)×100%。
1.2.4 不可逆点(PNR)时间测定
依据已有文献中介绍的方法[11],以孵化后天数(日龄,day after hatch,dah)表示,每天测定饥饿杂交F2仔鱼的初次摄食率,得到摄食率中的最高值后,当初次摄食率降到最高摄食率的50%以下时,即为杂交F2仔鱼的PNR时间。
1.3 数据处理
所得数据用Excel 2010软件统计处理并作图,以平行组数据的“平均值±标准差(
$\overline X \pm {\rm SD}$ )”表示,并运用F检验单因素方差分析比较饥饿组与对照组仔鱼的差异性。2. 结果
2.1 仔鱼的早期发育与卵黄囊变化
初孵仔鱼出膜后展直身体,倒悬于中上水层,鱼体透明,背、腹及尾部有较窄、无色透明的鳍膜;躯体部均匀分布点状色素,并于身体中部(第20肌节附近)色素斑点集中成簇;腹面前部有一椭球形或圆球形的卵黄囊,直径(0.85~1.17 mm)约占仔鱼体长一半,头部弯曲紧贴于卵黄囊前边,附于卵黄囊后端与身体连接处有一个扁圆形油球(直径0.22~0.31 mm,图1-a)。头部顶端至尾部末端全长2.23~2.59 mm,肛后处最大体高0.44~0.56 mm,肛后体长1.01~1.24 mm (约占全长的52%)。仔鱼发育形态特征与卵黄的吸收情况见图1。
![]() 图 1 初孵仔鱼至7日龄期的生长发育形态特征a. 初孵仔鱼;b. 1 日龄仔鱼;c. 2 日龄仔鱼;d. 3 日龄仔鱼;e. 4 日龄仔鱼;f. 5 日龄仔鱼;g. 6 日龄仔鱼;h. 7 日龄仔鱼;i. 5 日龄饥饿仔鱼;j. 6 日龄饥饿仔鱼;标尺=0.5 mmFig. 1 Morphological characteristics of newly hatched larvae to 7 daha. newly hatched larva; b. 1 dah larva; c. 2 dah larva; d. 3 dah larva; e. 4 dah larva; f. 5 dah larva; g. 6 dah larva; h. 7 dah larva; i. 5 dah starved larva; j. 6 dah starved larva; scale bar=0.5 mm
图 1 初孵仔鱼至7日龄期的生长发育形态特征a. 初孵仔鱼;b. 1 日龄仔鱼;c. 2 日龄仔鱼;d. 3 日龄仔鱼;e. 4 日龄仔鱼;f. 5 日龄仔鱼;g. 6 日龄仔鱼;h. 7 日龄仔鱼;i. 5 日龄饥饿仔鱼;j. 6 日龄饥饿仔鱼;标尺=0.5 mmFig. 1 Morphological characteristics of newly hatched larvae to 7 daha. newly hatched larva; b. 1 dah larva; c. 2 dah larva; d. 3 dah larva; e. 4 dah larva; f. 5 dah larva; g. 6 dah larva; h. 7 dah larva; i. 5 dah starved larva; j. 6 dah starved larva; scale bar=0.5 mm1日龄,仔鱼全长2.39~2.88 mm,体高0.50~0.72 mm,肛后体长1.47~1.87 mm;卵黄囊明显缩小,长径0.52~0.68 mm,短径0.46~0.58 mm。卵黄消耗体积超过初孵时的50%;腹部油球后面出现细长消化管(图1-b);仍然不能主动游泳,偶尔间歇性的向上窜动。
2日龄,仔鱼全长2.86~3.24 mm,体高0.59~0.73 mm,肛后体长1.91~2.15 mm;卵黄囊快速吸收、缩小至与腹部相平的小圆球形(图1-c),直径为0.22~0.27 mm,卵黄已消耗了80%;油球略缩小,直径0.16~0.23 mm;胸鳍外缘由半圆形伸长变成半椭圆形伸向两侧,仔鱼逐渐转为平衡游动。孵化约2.5 d后,口裂出现,随后上下颌开始微张,此时投饵3 h时未见摄食。
3日龄,仔鱼全长2.95~3.55 mm,体高0.66~0.75 mm,肛后体长2.02~2.17 mm,此时卵黄囊直径缩小为0.18 mm左右,与油球直径大小相近,消耗速率相对减慢,体积已减至初孵仔鱼的10%以下(图1-d);仔鱼开口增大,消化道、排泄管明显;鱼体腹部背面色素网状分布增多、加深,在肛后处也有大片色素点成簇;胸鳍增长加宽为扇形,开始自主游动,多分布于水的上层,开始有集群。第3.5 天时上下颌活动明显,下颌突出,大部分个体开始摄食,肠胃有食物。
4日龄,仔鱼全长3.17~3.48 mm,体高0.58~0.77 mm,肛后体长2.13~2.45 mm,肛后长占体长的60%以上,此时的卵黄囊及油球直径均比3日龄时又缩小近半(图1-e);卵黄囊直径约0.11 mm,油球直径0.09 mm;仔鱼鳃盖形成,脑部及腹部背面色素增多;胃部增大,肠道蠕动不停。
5日龄,仔鱼全长3.24~3.82 mm,体高0.68~0.78 mm,肛后体长2.15~2.38 mm,个体的卵黄囊已吸收、消耗了90%以上,油球直径约0.04 mm;仔鱼脑部及腹部网状色素面积增加,眼球遍布黑色素(图1-f),躯体中部的色素簇消失,出现胸鳍鳍褶;口裂增大到与头宽相当,肠胃饱满。
6日龄,仔鱼全长3.38~4.26 mm,体高达0.68~0.86 mm,肛后体长2.36~2.72 mm,约1/3个体仍有卵黄囊;仔鱼头部有局部色素集中,腹部增大,肠胃已占腹部大部分(图1-g)。
7日龄,仔鱼体长、体高快速增加,全长为3.78~4.45 mm,体高0.76~0.98 mm,肛后体长2.57~2.88 mm;卵黄囊、油球已完全消失;腹部膨大、变得更饱满,消化道背面、肛后处有大片的色素分布,多数个体腹背部内侧可见鱼鳔出现(图1-h)。
仔鱼卵黄囊在第1~第2天被快速消耗,随后转缓,油球在第3天开始明显缩小,而此时的仔鱼开始开口摄食进入混合营养阶段,部分个体的卵黄囊、油球在第6天逐渐消失,进入外源营养阶段;仔鱼完成从内源营养过渡到完全外源营养的混合营养期为4~5 d。
2.2 仔鱼的初次摄食率与PNR
在培育水温为(21.8±0.4) ℃条件下,杂交F2仔鱼孵出的第3~第7天进行首次投饵的初次摄食率变化见图2。在开口当天(3日龄),投饵3 h后,仔鱼的初次摄食率为(45.80±19.41)%,肠胃多有2~4个轮虫。4日龄饥饿仔鱼的初次摄食率为(69.5±14.08)%;5日龄饥饿仔鱼的初次摄食率提高至(77.21±14.67)%,最高达100%;6日龄时饥饿仔鱼初次摄食率下降为(29.83±4.29)%,低于最高初次摄食率的一半,仔鱼耐受饥饿的PNR即为6日龄,此时有部分个体仍有极少量的卵黄,但有过半的仔鱼因饥饿的损伤无法再恢复摄食能力;7日龄时,仔鱼的油球、卵黄已完全消失,初次摄食率为(8.87±6.83)%,此时有摄食仔鱼的活动力也已很弱。
![]()
以上结果表明,饥饿仔鱼具初次摄食能力时间为第4~第5天,最高摄食率在5日龄,随后摄食能力快速下降,PNR为6日龄(图2);此时仍有近1/3的仔鱼有初次摄食率,超过此时间点饥饿仔鱼多不能正常摄食,因此,杂交F2仔鱼最适的起始投饵时间为孵出后第3~第5天。
2.3 饥饿对仔鱼生长与成活率的影响
饥饿组仔鱼对卵黄的消耗与对照组没有明显差异。在经历了孵出后第1~第2天的90%以上的大量消耗后,杂交F2仔鱼在第3天起卵黄囊体积下降速度显著变缓,到6日龄时仅少部分个体尚有很小的卵黄囊,约有1/3个体在7日龄时才完成卵黄吸收。
完全饥饿条件下,3、4日龄仔鱼均有较好的觅食游动行为及趋光性,全长分别为(3.14±0.05) mm及(3.20±0.08) mm,与对照组差异不显著(P>0.05,图3),4日龄成活率(86.67%)略低于对照组(91.0%);5日龄仔鱼负生长,全长(3.04±0.03) mm,与对照组相比有显著差异(P<0.05),且此时不同个体卵黄囊的大小也相差较大,直径范围为0.046~0.12 mm,约半数个体有油球,直径为0.03~0.10 mm,成活率下降为71.3%,鱼体多呈现明显消瘦的形态(图1-i);到6日龄仔鱼全长为(3.10±0.01) mm,体高缩小为(0.52±0.03) mm,与对照组有显著差异(P<0.01),体色发白、腹部内陷(图1-j),游泳能力减弱、集群性差,成活率显著下降至26.67%,大约1/3的饥饿仔鱼有极少量的卵黄;7日龄时,全长为(3.01±0.03) mm,与对照组有极显著差异(P<0.01),体高缩小为(0.46±0.06) mm,赖以生存的内源营养已耗尽,饥饿仔鱼头大身瘦畸形,背部边缘粗皱、尾巴向下弯曲,在底层游动缓慢,成活率为16.67%(图1)。少数饥饿仔鱼卵黄在第7天时才完全被吸收,但7.5日龄只有几个存活的个体,成活率小于1%,第8天时全部死亡。
![]() 图 3 杂交F2仔鱼对照组与饥饿组全长随日龄变化比较*. 组间差异显著(P<0.05);**. 组间差异极显著(P<0.01)Fig. 3 Comparison of total length of hybrid F2 larvae with days after hatch between control group and starvation group*. significant difference between two groups (P<0.05); **. very significant difference between two groups (P<0.01)
图 3 杂交F2仔鱼对照组与饥饿组全长随日龄变化比较*. 组间差异显著(P<0.05);**. 组间差异极显著(P<0.01)Fig. 3 Comparison of total length of hybrid F2 larvae with days after hatch between control group and starvation group*. significant difference between two groups (P<0.05); **. very significant difference between two groups (P<0.01)对照组仔鱼在成活率保持较高水平(83.33%~91.0%)的同时,生长也保持较好的态势;全长(L,mm)与日龄(d)变化符合线性关系式L=0.276 1d+2.283 2 (R2=0.956 8)。对照组仔鱼的生长及卵黄囊吸收情况见表1。仔鱼全长的特定日增长率由高到低依次为2日龄、7日龄、1日龄,3~5日龄相对平缓,最低是6日龄;而饥饿组的增长率在3~4日龄逐渐下降,5日龄时为负增长,6日龄时略有上升,7日龄又表现为负增长(图4)。
表 1 杂交F2仔鱼对照组全长和卵黄囊、油球大小随日龄变化情况 (平均值±标准差)Table 1 Growth change of total length, size of yolk sac and oil globules of hybrid F2 larvae in control group with days after hatch ($\overline {\mathit{\boldsymbol{X}}} \pm {\bf SD}$ )日龄
dah全长/mm
total length体高/mm
body height肛后体长/mm
length after anus卵黄囊长径/mm
yolk long diameter卵黄囊短径/mm
yolk short diameter油球直径/mm
oil diameter0 2.40±0.09 0.47±0.05 1.14±0.15 0.95±0.08 0.63±0.04 0.24±0.03 1 2.61±0.19 0.59±0.07 1.57±0.28 0.54±0.07 0.42±0.01 0.21±0.03 2 2.98±0.12 0.68±0.10 2.06±0.06 0.25±0.03 0.23±0.03 0.18±0.02 3 3.15±0.09 0.71±0.03 2.11±0.10 0.16±0.04 0.15±0.02 0.13±0.01 4 3.31±0.20 0.70±0.19 2.26±0.08 0.12±0.04 0.10±0.01 0.09±0.00 5 3.52±0.14 0.72±0.07 2.35±0.07 0.10±0.00 0.07±0.01 0.04±0.00 6 3.63±0.08 0.73±0.08 2.41±0.09 0.06±0.00 0.03±0.00 0 7 3.95±0.06 0.84±0.09 2.57±0.05 0 0 0 ![]() 图 4 对照组和饥饿组杂交F2仔鱼1~7日龄仔鱼全长的特定日增长率Fig. 4 Comparison of specific daily growth rate of hybrid F2 larvae at 1–7 dah between control group and starvation group
图 4 对照组和饥饿组杂交F2仔鱼1~7日龄仔鱼全长的特定日增长率Fig. 4 Comparison of specific daily growth rate of hybrid F2 larvae at 1–7 dah between control group and starvation group3. 讨论
3.1 杂交F2早期仔鱼发育与摄食
仔鱼期是鱼类早期生活史中最脆弱、最敏感的时期。而早期仔鱼更是容易因营养保障与生长发育问题导致大量死亡,是鱼类早期发育的危险期,也是苗种繁育的关键阶段。本研究首次开展黑鲷(♀)×真鲷(♂)杂交子二代的苗种繁育,得到的杂交F2初孵仔鱼全长(2.23~2.59 mm)略大于同期的黑鲷仔鱼(1.90~2.25 mm),与同期的真鲷仔鱼(1.94~2.6 mm)[1]及其杂交F1亲本仔鱼(2.08~2.45 mm)大小相近。杂交F2仔鱼孵出后第1~第2天的卵黄消耗量最大,全长增长率也最大,随后的混合营养期(3~6日龄),卵黄消耗及全长增长趋势均较平缓,6日龄时卵黄基本吸收完毕,全长增长率也降至最低,至7日龄已完成转为外源营养时,仔鱼又表现出较快的生长态势。杂交F2仔鱼1~6日龄的生长及卵黄消耗特征与是否摄食没有明显相关性,这与真鲷[17]及唐鱼(Tanichthys albonubes)[18]等相似。有报道得出海水鱼类卵黄囊期仔鱼的生长可分为3个时期:1)初孵时的快速生长期;2)卵黄囊消失前后的慢速生长期;3)未能摄食外界食物的负生长期[19]。本实验中饥饿仔鱼也表现出这样的生长规律。仔鱼这种生长变化趋势是与其营养模式由内源营养-混合营养阶段-外源营养(摄食或饥饿)的变化进程相对应的结果。而杂交F2仔鱼对油球的吸收在第1~第2天相对较慢,第3天起明显加快,类似于条纹锯鮨 (Centropristis striata)[15]。在内源营养消耗后仔鱼个体的生长依靠摄食。本研究中正常摄食的杂交F2仔鱼7日龄已出现鱼鳔,其全长(3.78~4.65 mm)大于真鲷同期的仔鱼(3.5~3.7 mm)[1],也大于亲本杂交F1及黑鲷的同期仔鱼规格。这些结果说明杂交F2仔鱼发育良好,有较好的生长态势和培育可行性;取得的杂交F2鱼苗也为接下来的育种研究提供了难得的物质基础。
在仔鱼混合营养期,及时摄食是关系到早期仔鱼生长与存活的主要因素。初次摄食率的高低与最高摄食率持续时间的长短可以用来判断鱼类的摄食能力。高的初次摄食率意味着仔鱼群体具备了在饵料供应充足的情况下,获得高成活率的生物学基础[18]。许多海水鱼类仔鱼开口初次摄食率低于30%。杂交F2仔鱼开口初次摄食率为45.80%,远高于真鲷[17]、黄鲷(Dentex tumifrons)[20]及条石鲷(Oplegnathus fasciatus)[21]等多数鱼类;最高的初次摄食率为77.21%且只持续1 d,与真鲷[17]、黄鲷[20]及斜带石斑鱼(Epinephelus coioides)[22]等相近,又高于大泷六线鱼(Hexagrammos otakii)[14]、鮸鱼(Miichthys miiuy)[23]、斑鳜(Siniperca scherzeri)[24]等种类。而且,相比于开口当天不摄食的鱼类,如花尾胡椒鲷(Plectorhinchus cinctus)[25]等,杂交F2在开口当天就具有较高的摄食能力,有利于仔鱼及时建立外源性营养、提高成活率。
仔鱼的初次摄食率与摄食能力受到口裂大小、游泳能力、饵料密度及光照等内部因子及水生态条件因素的影响。笔者在是否充气、轮虫投喂密度、仔鱼培育密度或环境因子等不同条件下,对饥饿仔鱼在1 000 mL水体的烧杯中的初次摄食率进行了单一因素的比较实验,结果表明:1)不充气的效果好于微充气的,第1、第2天的3 h初次摄食率分别达50%及85%以上,这可能是在不充气的条件下,轮虫与仔鱼一样均趋光集群到一起,使仔鱼易于捕食到食物;2)仔鱼密度为20个·L–1或30个·L–1的初次摄食率无明显差异;3)投放轮虫密度由5~10个·mL–1增加到15~20个·mL–1,并没有提高仔鱼摄食效果;4)在仔鱼开口当天,投喂相同时间的同批次仔鱼,实验组仔鱼的初次摄食率比水泥池培育的低约25%,这可能与实验小水体与水泥池大水体在水质生态及取样偏差等方面有关。另外,笔者在检测同样实验条件下的另一批次仔鱼时,发现开口当天投饵2 h后的最高初次摄食率为20%,第3小时为70%以上,到第4小时达100%,明显好于第一次实验的结果;且相应的饥饿实验成活率也相对较高,这可能与仔鱼本身活力、受精卵质量乃至亲鱼质量等因素相关。彭志兰等[23]分析认为亲鱼经历的环境压力、积温、营养积累和所产卵的营养成分等直接影响到鮸鱼仔鱼的初次摄食率和PNR。
3.2 仔鱼的饥饿与PNR
饥饿是影响早期仔鱼的生长发育及成活率的主要因素[13-16,26]。虽然杂交F2仔鱼具较高的初次摄食能力,不过耐饥饿能力较差,饥饿仔鱼在开口的第3天起,全长出现负增长,开口后第4天饥饿仔鱼的成活率低于30%。类似的情况,开口后3日龄和PNR是真鲷仔鱼对外界环境的敏感期,其时高温和低盐耐力很低[27]。这可能与仔鱼体内能量不能满足其代谢耗能和正常的生长发育相关[28-29];随着饥饿时间的延长,对仔鱼危害不断加重,致使成活率不断下降。杂交鱼苗种生长虽然有可能存在杂种优势,但其苗种的培育能否成功存在不确定性;而且杂交鱼幼体发育受到多种复杂的生理、生化或免疫力的制约[30],其饵料投喂或生态环境调控尤其需要关注与探索。
PNR是衡量仔鱼饥饿耐力的指标,是指初次摄食期仔鱼达到耐受饥饿的时间临界点。抵达PNR后,饥饿仔鱼虽能存活一些时间,但已无法恢复摄食能力,所以也称“不可逆转饥饿(irreversible starvation)”或“生态死亡(ecological death)”[11]。从初次摄食期到PNR的这段时间是鱼类构建外源性摄食的关键时期。抵达PNR时间长,表明仔鱼耐饥饿能力强,有利于提高仔鱼建立外源性营养关系的可能性;反之,则耐饥饿能力弱[18]。仔鱼的发育与水温、卵质量等因素相关[31-32];仔鱼的PNR时间同样受水温及卵质量等因素影响[33],且种间甚至同种的不同种群间也有很大差异[11]。杂交F2仔鱼PNR在开口后第4天(6日龄),从PNR出现到全部仔鱼死亡仅1~2 d的时间,与真鲷[17]及其他多数海水鱼类相似,如花尾胡椒鲷[24]、黄鲷[20]、卵形鲳鲹(Trachinotus ovatus)[19]、太平洋鳕[34]及条纹锯鮨[15]等,属于易遭受饥饿胁迫鱼类;好于有些鱼类,仔鱼从开口到PNR时间只有2.5~3 d时间,如条石鲷[21]、牙鲆(Paralichthys olivaceus)[17]、点带石斑鱼(E.coioides)[35]及斜带石斑鱼[22]等。总之,大多数海水鱼类的PNR出现在开口后的第2~第4天[18],PNR之后,仔鱼已错过了正常构建外源营养吸收代谢关系的时机,因此抓住在临界点之前的只有2~3 d时间里及时投喂对育苗非常关键。
-
![]()
图 1 不同乳杆菌菌株发酵对鲭鱼加工副产物理化指标的影响
注:L0、L1、L2和L3分别为无任何处理的对照组、接种嗜酸乳杆菌、接种植物乳杆菌和接种贝氏乳杆菌的实验组;不同字母之间表示存在显著性差异 (P<0.05),图 2 同此。
Figure 1. Effects of different lactic acid bacteria fermentation on physicochemical indexe of processed by-products of mackerel
Note: L0, L1, L2 and L3 are the control group without any treatment, groups inoculated with L. acidophilus, L. plantarum and L. beijerinck, respectively. Different letters indicate significant difference (P<0.05). The same case in Fig. 2.
![]()
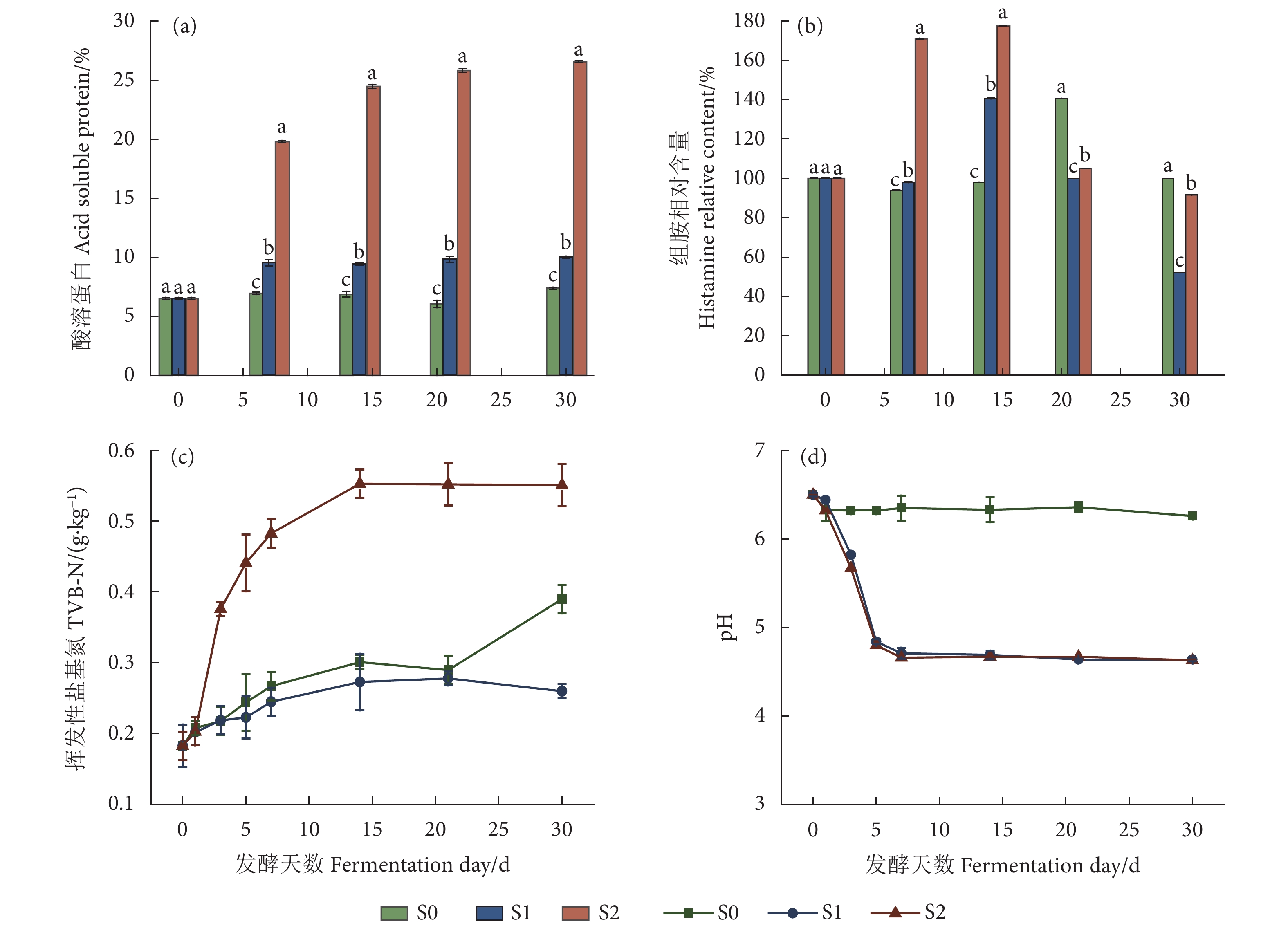
图 2 灭菌与不灭菌发酵对鲭鱼加工副产物理化指标的影响
注:S0、S1、S2 分别为灭菌不接种乳杆菌、灭菌接种乳杆菌、不灭菌接种乳杆菌;组胺相对含量由每一组组胺实际含量除以第 0 天未发酵的组胺实际含量计算得出。
Figure 2. Effects of sterilization and non-sterilization fermentation on physicochemical indexes of processed by-products of mackerel
Note: S0, S1 and S2 represent sterilization without inoculation of lactic acid bacteria, and non-sterilization with inoculation of lactic acid bacteria, respectively. The relative histamine content is calculated by the actual histamine content of each group by the actual histamine content of the sterilization group on 0th day.
![]()
图 3 不同发酵阶段的细菌群落相对丰度
注:保藏 270 d 指发酵后保存在常温下 270 d;图 4 同此。
Figure 3. Relative abundance of bacterial communities at different fermentation stages
Note: Storage of 270 d means storing at room temperature for 270 d after fermentation; the same case in Fig. 4.
![]()
图 4 不同发酵阶段真菌群落的相对丰度
Figure 4. Relative abundance of fungal communities at different fermentation stages
![]()
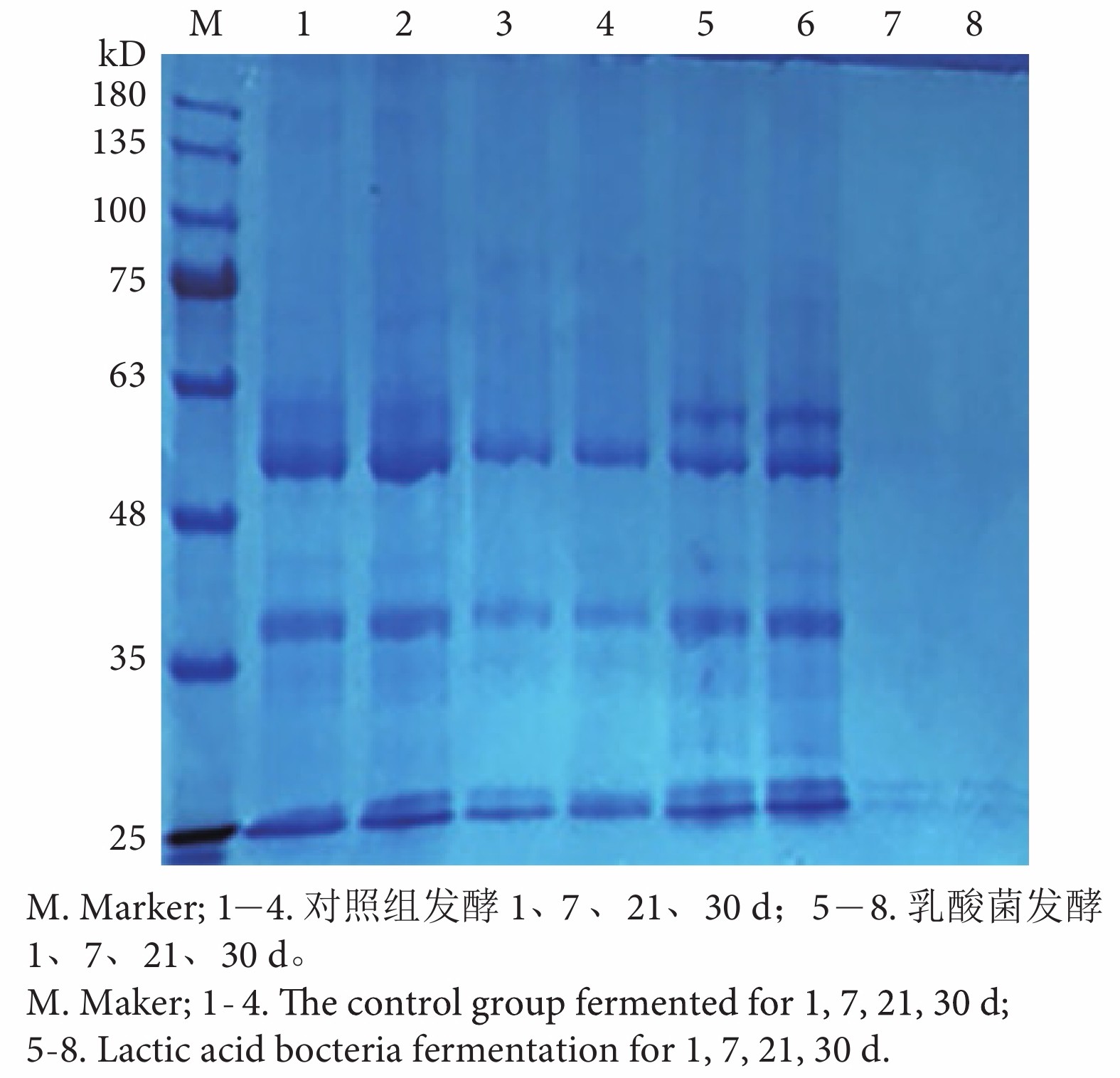
图 5 乳杆菌发酵降解抗原蛋白的SDS-PAGE蛋白电泳图谱
Figure 5. SDS-PAGE electrophoretic pattern of lactobacilli protein degraded by fermentation
表 1 植物乳杆菌发酵鲭鱼加工副产物的Alpha多样性指数
Table 1 Alpha diversity index of processed by-products of mackerel fermented by L. plantarum
样品
SampleACE 指数
ACE indexChao1指数
Chao1 index香农指数
Shannon index覆盖率
Goods coverage对照组
Control group第1天 1st day 123.735 123.438 0.905 1 第30天 30th day 151.349 149.636 1.072 1 发酵组
Fermentation group第1天 1st day 128.206 121.5 1.371 1 第30天 30th day 114.553 110.286 1.668 1 第270天 270th day 57.176 56.0 0.634 1  下载: 导出CSV
下载: 导出CSV
表 2 乳杆菌发酵鲭鱼加工副产物游离氨基酸组成的变化
Table 2 Changes of free amino acid composition of processed by-products of mackerel fermented by lactic acid bacteria
mg·g−1 氨基酸名称
Amino acid name发酵前
Before fermentation对照组第30天
Control group on 30th day乳杆菌发酵第30天
Lactic acid bacteria fermentation on 30th day天冬氨酸 Asparagine 3.742 4.674 4.701 苏氨酸 Threonine 1.420 1.565 1.493 丝氨酸 Serine 1.828 2.079 2.152 谷氨酸 Glutamic acid 7.111 8.030 7.897 甘氨酸 Glycine 2.874 2.863 3.034 丙氨酸 Alanine 2.315 2.411 2.567 胱氨酸 Cystine 0.357 0.313 0.410 缬氨酸 Valine 1.814 1.702 1.953 蛋氨酸 Methionine 0.186 0.118 0.184 异亮氨酸 Isoleucine 1.529 1.489 1.881 亮氨酸 Leucine 2.799 3.049 2.358 酪氨酸 Tyrosine 0.971 1.131 1.246 苯丙氨酸 Phenylalanine 1.896 2.036 1.893 赖氨酸 Lysine 1.107 1.197 1.081 组氨酸 Hlstidine 2.182 1.881 2.266 精氨酸 Argnine 2.940 2.898 2.938 脯氨酸 Proline 2.082 2.407 3.275 总和 Total 37.153 40.175 41.331
下载: 导出CSV
表 3 乳杆菌发酵前后脂肪酸组成变化
Table 3 Changes of fatty acid composition before and after lactic acid bacteria fermentation
% 脂肪酸种类
Fatty acid type发酵前占比
Proportion before fermentation乳杆菌发酵第30天占比
Proportion of lactic acid bacteria fermentation on 30th day十三烷酸 C13:0 9.34 7.51 棕榈酸 C16:0 24.13 25.13 棕榈油酸 C16:1 7.32 7.13 油酸 C18:1 2.79 3.02 亚油酸 C18:2 12.95 15.11 亚麻酸 C18:3 2.81 2.96 γ-亚麻酸 C18:3 8.15 9.53 花生酸 C20:0 2.16 2.15 二十碳二烯酸 C20:2 3.74 3.27 二十碳五烯酸 C20:5 8.83 7.75 芥子酸 C22:1 0.56 0.55 二十二碳二烯酸 C22:2 4.28 3.89 木蜡酸 C24:0 9.11 8.33 二十二碳六烯酸 C22:6 3.84 3.67 单不饱和脂肪酸 MUFA 10.67 10.70 多不饱和脂肪酸 PUFA 44.60 46.18 饱和脂肪酸 SFA 44.74 43.12
下载: 导出CSV
-
[1] 佘文熙, 曾宇, 赵俐娜, 等. 酵母葡聚糖鲭鱼鱼丸的研制及其质构特性和防腐效果的研究[J]. 食品工业, 2015, 36(9): 133-135. [2] 农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 中国渔业统计年鉴[R]. 北京: 中国农业出版社, 2020: 178. [3] ÁLVAREZ C, LÉLU P, LYNCH S A, et al. Optimised protein recovery from mackerel whole fish by using sequential acid/alkaline isoelectric solubilization precipitation (ISP) extraction assisted by ultrasound[J]. LWT, 2018, 88: 210-216. doi: 10.1016/j.lwt.2017.09.045
[4] 刘在军, 岑剑伟, 李来好, 等. 罗非鱼血液综合利用的研究思路及展望[J]. 南方水产科学, 2012, 8(2): 76-80. doi: 10.3969/j.issn.2095-0780.2012.02.012 [5] COLOMBO F M, CATTANEO P, CONFALONIERI E, et al. Histamine food poisonings: a systematic review and meta-analysis[J]. Crit Rev Food Sci Nutr, 2018, 58(7): 1131-1151. doi: 10.1080/10408398.2016.1242476
[6] ZHANG J B, JI C F, HAN J, et al. Inhibition of biogenic amines accumulation during Yucha fermentation by autochthonous Lactobacillus plantarum strains[J]. J Food Process Pres, 2021, 45(9): 15291.
[7] LIM E. Inhibitory effect of bacteriocin-producing lactic acid bacteria against histamine-forming bacteria isolated from Myeolchijeot[J]. Fish Aquat Sci, 2016, 19(1): 42. doi: 10.1186/s41240-016-0040-x
[8] 柳佳娜, 张宾, 刘宇, 等. 高效降解生物胺乳酸菌的筛选、鉴定及特性研究[J]. 中国酿造, 2021, 40(7): 141-147. doi: 10.11882/j.issn.0254-5071.2021.07.026 [9] 张栩. 不同乳酸菌发酵对酸菜的风味物质形成及品质指标的影响[J]. 中国酿造, 2021, 40(4): 133-137. doi: 10.11882/j.issn.0254-5071.2021.04.025 [10] WANG Q, YAMABE K, NARITA J, et al. Suppression of growth of putrefactive and food poisoning bacteria by lactic acid fermentation of kitchen waste[J]. Process Biochem, 2001, 37(4): 351-357. doi: 10.1016/S0032-9592(01)00217-5
[11] 沈颖莹, 吴燕燕, 李来好, 等. 发酵鳜鱼营养成分和安全性评价[J]. 南方水产科学, 2020, 16(3): 103-112. doi: 10.12131/2090247 [12] 肖志明, 李丽蓓, 邓涛, 等. 饲料原料中酸溶蛋白的测定方法研究[J]. 中国畜牧杂志, 2016, 52(2): 72-75,80. doi: 10.3969/j.issn.0258-7033.2016.02.015 [13] 李旺军, 方华, 季春源. 豆粕发酵蛋白中抗原蛋白和不良寡糖的检测[J]. 粮食与饲料工业, 2013(4): 61-65. [14] 王建, 林秋萍. 气相色谱-质谱法测定鱼油脂肪酸[J]. 中国饲料, 2001(17): 19-20. doi: 10.3969/j.issn.1004-3314.2001.17.011 [15] 王兴春, 杨致荣, 王敏, 等. 高通量测序技术及其应用[J]. 中国生物工程杂志, 2012, 32(1): 109-114. [16] 高林, 王新伟, 申国明, 等. 不同连作年限植烟土壤细菌和真菌群落结构差异[J]. 中国农业科技导报, 2019, 21(8): 147-152. [17] 曹荣, 林瑞环, 赵玲, 等. 基于高通量测序技术分析凡纳滨对虾(Litopenaeus vannamei)冷藏过程菌群结构变化[J]. 食品安全质量检测学报, 2019, 10(8): 2127-2131. doi: 10.3969/j.issn.2095-0381.2019.08.007 [18] AMATO K R, YEOMAN C J, KENT A, et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes[J]. ISME J, 2013, 7(7): 1344-1353. doi: 10.1038/ismej.2013.16
[19] CAPORASO J G, KUCZYNSKI J, STOMBAUGH J, et al. QIIME allows analysis of high-throughput community sequencing data[J]. Nat Methods, 2010, 7(5): 335-336. doi: 10.1038/nmeth.f.303
[20] FENG L, LI Y, TUO Y, et al. The bacterial diversity of sun-dried spanish mackerel in Dalian and application of Lactobacillus plantarum X23 as a bio-preservative[J]. J Food Prot, 2021, 84(12):2133-2142.
[21] DOMINGOSLOPES M F, PSTANTON C, ROSS R P, et al. Histamine and cholesterol lowering abilities of lactic acid bacteria isolated from artisanal Pico cheese[J]. J Appl Microbiol, 2020, 129(6): 1428-1440. doi: 10.1111/jam.14733
[22] 吕文竹, 王洋, 方玉美, 等. 不同发酵条件对构树饲料酸溶蛋白含量的影响[J]. 饲料研究, 2021, 44(11): 75-78. [23] 谢全喜, 侯楠楠, 王梅. 高产苯乳酸菌株的筛选及其在豆粕发酵中的应用[J]. 中国酿造, 2021, 40(7): 65-70. doi: 10.11882/j.issn.0254-5071.2021.07.013 [24] ZHOU Y Q, WU S M, PENG Y L. Effect of lactic acid bacteria on mackerel (Pneumatophorus japonicus) seasoning quality and flavor during fermentation[J]. Food Biosci, 2021, 41: 100971. doi: 10.1016/j.fbio.2021.100971
[25] JAYACHITRA J, SIVASAKTHIVELAN P, BABU E. Inhibitory effect of bacteriocin-producing lactic acid bacteria against histamine-producing bacteria isolated from fish[J]. Res J Biotechnol, 2021, 16(3): 55-61.
[26] KOBAYASHI T S, WANG X G, SHIGE T N, et al. Distribution of histamine-producing lactic acid bacteria in canned salted anchovies and their histamine production behavior[J]. Ann Microbiol, 2016, 66(3): 1277-1284. doi: 10.1007/s13213-016-1213-7
[27] ZANG J H, XU Y S, XIA W S, et al. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing[J]. Food Res Int, 2018, 111: 565-573. doi: 10.1016/j.foodres.2018.05.076
[28] 王苾, 马明博, 朱弘焱, 等. 大骨鸡不同产蛋水平粪便微生物群落特征[J]. 饲料研究, 2021, 44(5): 75-81. [29] 江艳华, 王联珠, 许东勤, 等. 基于高通量测序分析虾夷扇贝柱菌群结构及腐败优势菌[J]. 食品科学, 2018, 39(20): 140-145. doi: 10.7506/spkx1002-6630-201820021 [30] KUNG H F, LE Y H, CHANG S C, et al. Histamine contents and histamine-forming bacteria in sufu products in Taiwan[J]. Food Control, 2007, 18(5): 381-386. doi: 10.1016/j.foodcont.2006.02.012
[31] LIU B Y, HUAN H L, GU H R, et al. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages[J]. Bioresour Technol, 2019, 273: 212-219. doi: 10.1016/j.biortech.2018.10.041
[32] 李倩, 赵颖, 朱晓嫚, 等. 天然产物抑制黄曲霉生长及其产毒的研究进展[J]. 河南工业大学学报(自然科学版), 2021, 42(1): 132-140. [33] 刘闪, 曹星星, 吴攀, 等. 酸性矿山废水影响下水库真菌群落特征与环境因子研究[J]. 环境科学与技术, 2021, 44(2): 1-8. [34] BAWIN T, SEYE F, BOUKRAA S, et al. Histopathological effects of Aspergillus clavatus on larvae of the southern house mosquito, Culex quinquefasciatus[J]. Fungal Biol, 2016: 489-499.
[35] KIRK P M, PEGLER D N, SUTTON B C. Dictionary of the Fungi[M]. 10th ed. Trowbridge: Cromwell Press, 2008: 315-351.
[36] 白小燕, 邱树毅, 雷安亮, 等. 酱香白酒酿造过程中产多元醇功能酵母的筛选[J]. 中国酿造, 2017, 36(5): 58-62. doi: 10.11882/j.issn.0254-5071.2017.05.012 [37] 肖辰, 陆震鸣, 张晓娟, 等. 泸型酒中层酒醅真菌群落的发酵演替规律[J]. 应用与环境生物学报, 2018, 24(5): 1081-1086. [38] LEE Y C, TSENG P H, HWANG C C, et al. Effect of vacuum packaging on histamine production in Japanese Spanish mackerel (Scomberomorus niphonius) stored at various temperatures[J]. J Food Protect, 2019, 82(11): 1931-1937. doi: 10.4315/0362-028X.JFP-19-143
[39] 顾赛麒, 唐锦晶, 周绪霞, 等. 腌腊鱼传统日晒干制过程中品质变化与香气形成[J]. 食品科学, 2019, 40(17): 36-44. doi: 10.7506/spkx1002-6630-20180716-201 [40] 王悦齐, 李来好, 蔡秋杏, 等. 分离自腌干鱼的抗氧化发酵菌株的筛选及鉴定[J]. 南方水产科学, 2016, 12(3): 74-83. doi: 10.3969/j.issn.2095-0780.2016.03.010 [41] 田建军, 张开屏, 景智波, 等. 瑞士乳杆菌与木糖葡萄球菌对发酵香肠蛋白质分解和游离脂肪酸释放的影响[J]. 中国食品学报, 2019, 19(3): 141-147. [42] 王章存, 袁路阳, 张露, 等. 生物酶解法去除大豆蛋白抗原性研究进展[J]. 食品工业科技, 2018, 39(15): 317-321. -
期刊类型引用(2)
1. 胡重华,刘斌,黄雅贞,张家海,连海,范紫嫣,张万昌,卢新民,方园. 光倒刺鲃胚胎及仔、稚鱼发育研究. 南昌大学学报(理科版). 2025(02): 208-214 .  百度学术
百度学术
2. 韩豪祥,廉杰,金洪宇,马波. 额尔齐斯河黑鲫胚胎及早期仔鱼发育特征. 中国水产科学. 2024(06): 684-692 . 百度学术
其他类型引用(0)









计量
- 文章访问数: 602
- HTML全文浏览量: 216
- PDF下载量: 57
- 被引次数: 2
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
