
Effects of desiccation on survival, metabolism-related enzymes and histological structure of adult red swamp crayfish (Procambarus clarkii)
-
摘要: 为研究干露对克氏原螯虾 (Procambarus clarkii) 成虾的影响,在 (16±1) ℃和相对湿度 (Relative humidity, RH) (55±5)%的条件下,对成虾进行0、3、6、12、24、36、48、60、72、96、132和144 h干露处理。结果显示,雄虾半致死时间 (LT50) 为148.36 h,雌虾LT50为144.01 h,两者体质量消耗率随干露时间的延长逐渐升高,雄虾在干露144 h后体质量消耗率达16.09%,雌虾在132 h后达15.87%。鳃、肝胰腺和肌肉组织结构随干露时间的延长损伤逐渐加剧。肝胰腺和肌肉乳酸脱氢酶 (LDH) 活力呈先上升后下降的趋势,雌、雄虾肝胰腺LDH分别在第24和第36小时达到最大活力值,肌肉LDH分别在第36和第48小时达到最大活力值,肌肉乳酸 (LA) 浓度呈现相似的变化趋势,雌、雄虾LA分别在干露第60和第48小时达到最大浓度。琥珀酸脱氢酶 (SDH) 活力在胁迫期间呈下降趋势。成虾在干露约第48 小时耐受能力达到极限,呼吸代谢由有氧呼吸逐步转换到以无氧呼吸为主,并伴有组织损伤。雌、雄虾的抗干露胁迫能力存在差异,在进行成体运输时干露时间不宜超过36 h。Abstract: To investigate the effects of desiccation on adult red swamp crayfish (Procambarus clarkii), we designed a desiccation stress for crayfish for 0, 3, 6, 12, 24, 36, 48, 60, 72, 96, 132 and 144 h under the conditions of (16±1) ℃ and (55±5) % relative humidity (RH). The results show that the half-lethal time (LT50) of male and female crayfish were 148.36 and 144.01 h, respectively. The mass consumption rate increased gradually with the extension of desiccation time; the weight consumption of male reached 16.09% of body mass after 144 h desiccation, while that of female reached 15.87% of body mass after 132 h desiccation. Besides, the histological structure damage of gill, hepatopancreas and muscle was gradually aggravated. Moreover, the activity of lactate dehydrogenase (LDH) in hepatopancreas and muscle first increased and then decreased. The maximum LDH activities of hepatopancreas in female and male were observed at 24th and 36th hour, while those of muscle in female and male were observed at 36th and 48th hour, respectively. The lactic acid (LA) content of muscle showed similar trend, with LA contents of female and male being the highest after 60 and 48 h desiccation stress, respectively. The succinate dehydrogenase (SDH) activity reduced significantly during desiccating. Furthermore, the desiccation tolerance of adult crayfish reached the limit after 48 h, and the respiratory metabolism gradually transformed from aerobic respiration to anaerobic respiration, accompanied by histological damage. In conclusion, the ability of resist desiccation between male and female is different. Thus, the desiccation time should not exceed 36 h when transporting adult crayfishes.
-
河流栖息地是对水生生物有直接或间接影响的多种尺度下物理化学条件的组合[1],随着人类涉水活动的增加,鱼类等水生生物的栖息地正遭受严重破坏。河流生物栖息地评估在河流生态修复中具有重要作用,通过栖息地评估可为河流生态修复提供基础信息和依据[2]。针对不同水动力条件下鱼类生境的变化,国内外学者进行了大量研究[3-5]。小尺度产卵场的地形特征是研究产卵场水动力特征的基础。河流的水动力特征与水下地形、地貌存在密切关系。特殊的河床形态决定了特殊的水力学特征,如坡度与底部流速存在正相关关系[6],这些因素决定了鱼类的产卵条件[7]。河流地形地貌在河流水量和水质不变的基础上,与生物群落的多样性存在正相关性,可影响其功能和结构[8]。河流地形地貌方面的研究集中在河道和河段尺度上,多为定性描述[9]。定量的、微尺度的高程、高程变异系数、地形复杂度、坡度、坡向等地形因子,多应用于海洋河口或中华鲟 (Acipenser sinensis) 等珍稀濒危鱼类的产卵场研究,而在“四大家鱼”产卵场方面的研究运用较少[7,10-12],珠江流域尚未见报道。
西江水系约有70多处经济鱼类产卵场,尤其以郁、黔、浔三江最为集中。东塔产卵场是珠江流域最大的产卵场,也是斑鳠 (Mystus guttatus)、卷口鱼 (Ptychidio jordani) 等珠江众多珍稀特有鱼类的重要栖息地,具有重要的生态功能 [13-14]。近年来受水利水电开发、航道整治、挖沙等涉水活动影响,鱼类栖息地遭到严重破坏,表现为生境丧失、破碎化和同质化,导致鱼类多样性降低、个体小型化,鱼类群落组成发生巨大变化[15-16]。主要渔业对象由80年代的鲥 (Tenualosa reevesii)、花鰶 (Clupanodon thrissa)、青鱼 (Mylopharyngodon piceus)、草鱼 (Ctenopharyngodon idella)、赤眼鳟 (Squaliobarbus curriculus) 等变为目前的广东鲂 (Megalobrama terminalis)、赤眼鳟、鲮 (Cirrhinus molitorella)、䱗 (Hemicculter Leucisculus),“四大家鱼”仔鱼资源在总鱼苗量中所占比例从46.6%下降至4.51%[17-18]。
大藤峡水利枢纽位于东塔产卵场上游约8 km,于2020年建成。该电站的运行,势必会改变下游江段的水下地形结构,导致鱼类栖息地的生境发生变化,从而影响其生存和繁衍[19-20]。要准确认识工程对鱼类栖息地的影响,必须对自然条件下栖息地状况,鱼类与栖息地地形、地貌间的关系有清楚而充分的认识。因此,有必要对该江段水下地形特征进行研究,评估水坝建设及运行对下游鱼类栖息地的影响。
目前,珠江流域关于鱼类微观栖息地地形、地貌的研究尚未见报道,相关研究仅限于定性描述[21],栖息地功能现状缺少科学评估。本研究首次采用声学方法对东塔产卵场栖息地现状进行评估,并构建了栖息地适宜性指数 (Habitat suitability index, HSI) 模型,以期为江河鱼类栖息地功能评价、生态修复提供技术支撑。
1. 材料与方法
1.1 研究区域
东塔产卵场位于浔江上游,自黔、郁两江汇合口起至东塔村止,长约7 km,为珠江最大“四大家鱼”产卵场。该研究区域上起大藤峡水利枢纽下游,下至石咀镇 (图1),具体地理位置为 110.057°E—110.16°E、23.432°N—23.467°N。研究区域长约14 km,水深介于1.5~32.0 m。采用ArcGIS 10.1软件绘制地图,数据来源于全国地理信息资源目录服务系统 (https://www.webmap.cn/,1∶25 万全国基础地理数据库,2019公众版) 。
1.2 数据采集
分别于2016年4月、2017年6月、2018年4月和2019年4月对该江段进行了声学走航探测。使用仪器为分裂波束渔探仪EY60 (Simrad,挪威),换能器中心频率120 kHz,发射功率200 W,−3 dB波束宽7.0°,脉冲宽度256 μs。在每次调查前,参照校准流程,使用碳化钨校准球 (Φ23 mm) 对系统收发增益进行校准[22]。调查船只使用当地渔船,长约8 m,换能器垂直向下固定于船舷,吃水约0.5 m。船速约4.4~5.5 kn,走航方向为顺流而下,设计航线为“之”字形,在实际航程中根据调查江段情况进行实时调整。
2019年4月使用声学多普勒流速剖面仪 (ADCP,WHR600,美国) 对产卵场进行流场、地形同步监测。在测量前,记录待测量区域内上下游断面中心点的坐标,以及弯道处断面中心点坐标,通过配置引导物,画出测量区域的计划线。在实际测量时,根据导航测量软件,记录、校正测船航迹,使测船沿着计划线依次测量。该次测量断面间距为150 m,共94个断面,通过ADCP实测,监测期间浔江流量约5 000 m3·s−1。
1.3 数据分析
1.3.1 地形因子提取
测量的原始数据在ADCP辅助测量软件WinRiver以ASCII码文件导出,读取每个测量断面、流速矢量、水深、经纬度等数据,使用ArcGIS 10.1 软件构建河床不规则三角网和数字高程模型,提取产卵场高程、坡度、坡向、地形起伏度、高程变异系数、地表粗糙度共6个地形因子[11]。
地形因子之间可能存在共线性关系,分析数据时,采用 Pearson相关系数判定两者之间的相关性,当两者存在共线性关系时,仅选取其中一个变量为解释变量。
1.3.2 热点分析
热点分析用于识别具有统计显著性的高值 (热点) 和低值 (冷点) 的空间聚类,是根据在一定分析规模内的所有要素,计算每个要素Getis-Ord Gi*统计值,得到每个要素的z得分和P值。统计学上的显著性正值z得分表示热点,z得分越高,表示热点聚集越紧密;负值表示冷点,z得分越低,冷点聚集就越紧密。通过z得分和P值,可以得到产卵场鱼类资源密度高值或低值在空间上发生聚类的位置[23]。
与传统的鱼类空间分布相关研究不同,本研究侧重从地理空间视角去解释鱼类的空间聚类。首先使用声学数据后处理软件Echoview 5.4 (Myriax Pty Ltd. 澳大利亚) 对原始数据进行处理,获得不同调查时间的鱼类密度信息。其次利用ArcGIS 10.1软件中的不等边三角形TIN方法建立密度数据范围,通过建立TIN能够剔除无数据的水域;然后利用空间统计分析模块中的空间自相关和热点分析对鱼类密度数据进行分析和结果可视化输出。
1.3.3 栖息地模型建立
2016、2018和2019年采样时间均为4月,2017年为6月。为降低在热点分析和模型建立中季节变化造成的影响,本文利用2016、2018和2019年数据构建模型,以2017年数据验证模型。
本文假设2年及以上鱼类热点分布重叠区域 (网格) 为鱼类适宜分布区域,提取热点区域中的地形因子。根据频率分布法,统计各因子分布范围,然后对结果进行归一化处理,估算适宜度指数值 (Suitability index, SI),出现频次最高的值赋予SI=1,SI∈[0, 1]。
由于几何平均值考虑了各因子之间的补偿影响,对生境适宜性的估计更为保守,因此HSI模型采用几何平均法建立,计算公式为[4]:
$$ S_{\mathrm{HSI},i}=\left(\mathrm{SI}_{1 i} \times \mathrm{SI}_{2 i} \times \ldots \times \mathrm{SI}_{n i}\right)^{{}^{1} \diagup {}_{n}\;} $$ (1) 式中:
$S_{\mathrm{HSI},i}$ 为第i个网格的栖息地适宜性指数值;i为研究区域网格序号;${\mathrm{S}\mathrm{I}}_{{n}{i}}$ 为第n个地形因子第i个网格的SI值。1.4 HIS模型验证
利用2017年6月的声学调查数据及水深数据,分析该江段鱼类资源密度分布情况,验证是否HIS值越高的网格鱼类密度越高。
2. 结果
2.1 鱼类密度及空间热冷点分布格局
2016—2019年鱼类平均密度见表1,鱼类密度呈逐年下降趋势,使用非参数检验进行分析,Kurskal-Wallis结果显示,不同调查时间鱼类密度存在显著性差异 (P<0.05)。 从偏度值可以看出数据左偏,表明低密度单元格较多。
表 1 东塔产卵场不同调查时间鱼类密度分布信息Table 1. Fish density distribution in Dongta spawning ground in different periods调查时间
Survey year鱼类密度
Fish density/(尾·m−3)范围
Range偏度
Skewness峰度
Kurtosis变异系数
Coefficient of variance2016年 0.057 4±0.128 7 (0.000 3, 1.018 4) 6.146 41.269 2.24 2017年 0.043 4±0.056 5 (0.007 2, 0.405 3) 3.744 17.185 1.30 2018年 0.031 6±0.037 7 (0.000 4, 0.196 7) 2.004 4.265 1.19 2019年 0.009 4±0.007 1 (0.000 4, 0.035 6) 1.492 2.536 0.75 将鱼类密度导入GIS中进行空间自相关和热点分析,得到鱼类在东塔产卵场的热点分布图 (图2)。空间上,研究区域鱼类资源的空间分布具有一定的聚集特征,即不同年份均出现“热点区域”(红色区域),标志为z>+1.65 (P<0.1),如东塔产卵场部分区域3年结果均为“热点区域”;而大藤峡下游及石咀段出现“冷点区域”(蓝色区域);其他江段绝大部分z值介于−1.65~+1.65,这些区域内渔业资源密度高值和低值之间的空间自相关性弱,为随机性分布。总体来看,东塔产卵场部分区域局部自相关性强,整体自相关性表现并不强烈,鱼类资源密度呈现“热”“冷”不均的局部性分布特征。
![]() 图 2 不同年份东塔产卵场鱼类资源空间热点分布图Figure 2. Hotspots of fsh resources in Dongta spawning ground in different years
图 2 不同年份东塔产卵场鱼类资源空间热点分布图Figure 2. Hotspots of fsh resources in Dongta spawning ground in different years2.2 适宜度曲线构建
根据上述方法,从鱼类“热点分布”区域中选取2年及以上重合区域网格,共80个,提取各网格地形因子,利用频度分布,拟合建立地形因子的单因子适宜度曲线。
本文共获取了6个解释变量,包括高程、坡度、坡向、地形起伏度、高程变异系数、地表粗糙度。高程变异系数、地形起伏度和坡度的相关系数大于0.85,因高程变异系数和地形起伏度是基于高程 (水深) 计算得出的,因此只保留坡度进行分析;地表粗糙度数据主要集中在1左右,无法拟合。因此选择水深、坡度、坡向3个地形因子作为预测变量参与分析。由图3可见,热点区域水深介于1.5~25.0 m,鱼类适宜水深为5.0~8.0 m;坡度介于0°~27°,鱼类适宜范围介于1°~1.5°,依据坡度等级,鱼类适宜区域为微斜坡;鱼类适宜的坡向介于90°~180°,即东南方向。
2.3 HSI值计算
根据公式 (1) 计算得出鱼类密度的HSI值。5 000 m3·s−1流量下HSI值分布介于0.02~0.95,平均值为0.31±0.22。其中,HSI值大于0.5的区域占总数的26.67%。由图4可知,研究江段HSI最大值出现在产卵场沙洲两侧,河流两侧也有零星分布;另外,主航道基本上为HSI低值区域。
![]() 图 4 东塔产卵场鱼类栖息地适宜性指数分布图Figure 4. Habitat suitability index distribution of Dongta spawning ground
图 4 东塔产卵场鱼类栖息地适宜性指数分布图Figure 4. Habitat suitability index distribution of Dongta spawning ground2.4 栖息地模型验证
使用2017年6月声学数据对HSI模型进行验证。通过Pearson相关性分析可知,鱼类密度与HSI值呈正相关关系 (R=0.794, P=0.011)。拟合结果显示,HSI与鱼类密度的关系为:
$ y=0.02x+ 0.04 $ (R2=0.63, RMSE=0.005),符合本文假设。3. 讨论
3.1 地形因子的选择性特征
东塔产卵场位于郁江、黔江和浔江三江交汇口,鱼类种类丰富,物种多样性高。文献资料显示,该江段鱼类优势种为䱗、鲤 (Cyprinus carpio)、鲮、赤眼鳟、广东鲂、卷口鱼等[15-16]。河流物种多样性和河床特征存在着一定程度的响应关系,同时生物群落的结构和功能受之影响。然而,目前尚不清楚地形因子如何影响鱼类[24]。本研究提取了水深、坡度、坡向、地形粗糙度、高程变异系数、曲率等地形因子进行分析,通过判定各因子之间的相关性及各因子取值的分布情况,筛选出了水深、坡度、坡向作为构建适宜性模型的因子。
水深是鱼类生活空间的反映,鱼的体型大小、生活习性等决定了鱼类对水深的需求[25]。Miranda和Killgore [26]研究表明,鱼类分布与水深呈非随机关系,约75%的物种分布在12 m以浅。Sheldon[27]认为鱼类分布与水深有着紧密联系,随季节和种类的不同而变化。本研究区域最大水深为31.0 m,鱼类适宜水深介于5.0~8.0 m。从图1可以看出,深水区主要为航道,航道在扩能升级过程中改变了河床结构,破坏了鱼类栖息地;另外,鱼类会对航道中的行船噪声产生回避行为[28-29],因此深水区基本为HSI低值区。
水下地形结构影响着鱼类的分布和多样性[24]。相关研究表明,疏浚通过改变水下地形,显著影响鱼类群落及空间分布。鱼类丰度和多样性从与自然河口陡峭的坡度相关,转变为与疏浚河口平缓的坡度相关[12]。西江航道经过二次扩能升级,河流水下地形地貌已发生了变化[30],因此鱼类偏好栖息于坡度较缓的区域[31]。
坡向是坡度所面对的方向,用于识别表面上某一位置处的最陡下坡方向。相关研究表明,中华鲟产卵场平均坡向分别为219.92°和207.63°,长江四大家鱼产卵场平均坡向介于150.16°~200.6°[10-11],本研究中栖息地坡向适宜范围介于90°~180°。上述产卵场坡向分布范围不同,但根据河流地形走势来看,坡向均为水流方向;因此推测鱼类偏好栖息于河流中向着水流方向的微斜坡。
3.2 栖息地适宜度
HSI模型是通过适宜度方程评价各个生态因子对某一特定物种的适宜度值和综合值,以此来评价环境对这个特定物种生存和繁殖的适合程度。以往研究使用的生物因子来源于专家经验或者室内实验[3]。近年来,相关研究越来越重视探讨在自然状态下鱼类分布与生境因子之间的关系,以此来确定鱼类适宜栖息地范围[31]。本研究根据鱼类对生境的利用情况来绘制适宜度曲线,鱼类时空分布及水下地形数据均为实测,地形因子的频率分布是基于3年声学调查结果,利用局部空间自相关方法对鱼类空间分布进行分析,定位到鱼类分布的高密度区域,能较为准确地反映野外实际情况。因此栖息地适宜性指数具有较高的可靠度。另外,模型验证结果显示,HSI值较高的区域,鱼类密度较大,表明该模型适用于东塔江段鱼类栖息地适宜性评价。
在数据分析中,经常会遇到少量的鱼类高密度值,这些值是研究者关注的热点;但从经典统计学角度讲,这些值却是造成评估不确定性的原因[32]。鱼类以个体、种群、群落的形式分布在特定空间上,具有高度的空间自相关性。经典统计学受基本假设的限制,在研究个体、种群和群落空间自相关方面具有较多缺陷[23]。热点分析可划分鱼类资源高低属性值的空间聚集,能够直观清晰地表现渔业资源分布情况,结果具有统计学意义。在渔业生态学中,热点分析已应用于栖息地变动趋势、物种丰富度等方面[33-35]。本研究尝试应用该法来识别鱼类适宜栖息地,取得了较好的效果。由于能够较为直接清晰地表现渔业资源分布情况,热点分析的应用前景将会被不断拓展。
3.3 研究不足及未来展望
本研究以河流地形特征作为变量研究鱼类栖息地适宜情况。坡度、坡向为常量,不会随流量的变化而改变。水深作为表征高程的因子,会随径流量的变化而增减。随着流量增大,水深加深,河流两岸漫滩及中心沙洲会被淹没,适宜鱼类栖息的区域也会发生改变。但是本研究区域边界是基于5 000 m3·s−1流量建立的,河流边界无法随流量的增加而变化。因此本文所建模型无法模拟高流量下的河流栖息地状况。
未来在开展此类工作时,野外采样应选择在水位较高时,尽可能将河流两岸漫滩及中心沙洲淹没。漫滩作为河流生态系统的重要组成部分,为鱼类提供了多样化的栖息生境,具有高生产力和微生境异质性,对维持区域鱼类生物多样性具有重要作用[36-37]。本研究探明了大藤峡水利枢纽蓄水前东塔产卵场水下地形特征及鱼类时空分布,今后仍需持续监测,以评估该电站运行对坝下江段鱼类栖息地的影响。
-
![]()
图 2 干露胁迫下克氏原螯虾的体质量消耗率
大写字母不同表示雄虾不同胁迫时间组的差异显著 (P<0.05),小写字母不同表示雌虾不同胁迫时间组的差异显著 (P<0.05)。
Figure 2. Mass consumption rate of P. clarkii in air exposure periods
Values with different uppercase and lowercase letters indicate significantly difference in male and female crayfish among different groups, respectively (P<0.05).
![]()
图 3 干露胁迫对克氏原螯虾雄虾 (a) 和雌虾 (b) 肝胰腺乳酸脱氢酶活力的影响
*. 各组与0 h组的差异显著 (P<0.05);不同字母上标表示除0 h组外不同胁迫时间组的差异显著 (P<0.05);后图同此。
Figure 3. Effect of dry exposure on hepatopancreas LDH activity of male (a) and female (b) P. clarkii
*. Significant difference between each group and 0 h group (P<0.05); different letters represent significant difference among different groups except 0 h group (P<0.05). The same case in the following figures.
![]()
图 4 干露胁迫对克氏原螯虾雄虾 (a) 和雌虾 (b) 肌肉乳酸脱氢酶活力的影响
Figure 4. Effect of dry exposure on muscle LDH activity of male (a) and female (b) P. clarkii
![]()
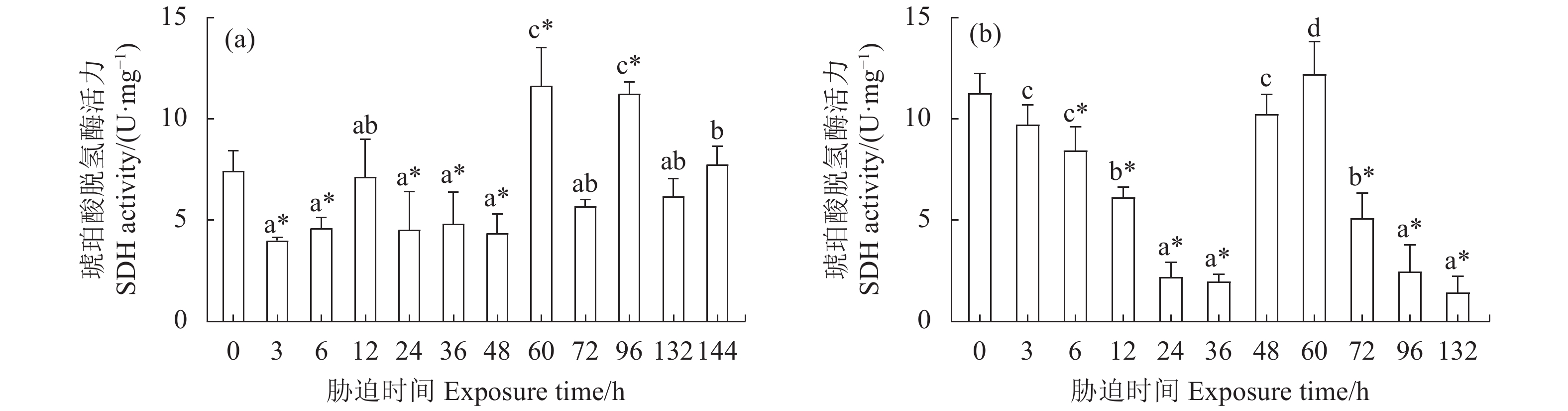
图 5 干露胁迫对克氏原螯虾雄虾 (a) 和雌虾 (b) 肝胰腺琥珀酸脱氢酶活力的影响
Figure 5. Effect of dry exposure on hepatopancreas SDH activity of male (a) and female (b) P. clarkii
![]()
图 6 干露胁迫对克氏原螯虾雄虾 (a) 和雌虾 (b) 肌肉琥珀酸脱氢酶活力的影响
Figure 6. Effect of dry exposure on muscle SDH activity of male (a) and female (b) P. clarkii
![]()
图 7 干露胁迫对克氏原螯虾雄虾 (a) 和雌虾 (b) 肌肉乳酸质量摩尔浓度的影响
Figure 7. Effect of dry exposure on muscle LA molar concentrations of male (a) and female (b) P. clarkii
![]()
图 8 干露胁迫对雄性克氏原螯虾肝胰腺显微结构的影响
a—i分别代表干露胁迫0、12、24、36、48、60、72、96和132 h;B. B细胞;R. R细胞;F. F细胞;L. 管腔;BM. 基膜;TV. 转运泡;图9同此。
Figure 8. Effect of dry exposure on hepatopancreas microstructure of male P. clarkii
a–i represent dry exposure of 0, 12, 24, 36, 48, 60, 72, 96 and 132 h, respectively; B. B cell; R. R cell; F. F cell; L. Lumen; BM. Basement membrane; TV. Transferred vacuoles; the same case in Figure 9.
![]()
图 9 干露胁迫对雌性克氏原螯虾肝胰腺显微结构的影响
Figure 9. Effect of dry exposure on hepatopancreas microstructure of female P. clarkii
![]()
![]()
图 11 干露胁迫对雌性克氏原螯虾鳃显微结构的影响
Figure 11. Effect of dry exposure on gill microstructure of female P. clarkii
![]()
图 12 干露胁迫对雄性克氏原螯虾肌肉显微结构的影响
Figure 12. Effect of dry exposure on muscle microstructure of male P. clarkii
-
[1] 堵南山. 甲壳动物学 下册[M]. 北京: 科学出版社, 1993: 821-822. [2] 农业农村部渔业渔政管理局. 2020 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2020: 24. [3] 谢佳彦, 朱爱意. 几种重要水产品活体运输技术研究[J]. 水产科学, 2010, 29(9): 532-536. doi: 10.3969/j.issn.1003-1111.2010.09.007 [4] PAITAL B. Antioxidant and oxidative stress parameters in brain of Heteropneustes fossilis under air exposure condition; role of mitochondrial electron transport chain[J]. Ecotoxicol Environ Saf, 2013, 95: 69-77. doi: 10.1016/j.ecoenv.2013.05.016
[5] ABELE D, PHILIPP E, GONZALEZ P M, et al. Marine invertebrate mitochondria and oxidative stress[J]. Front Biosci, 2007, 12(3): 933-946. doi: 10.2741/2115
[6] MARTINE-ALVAREZ R M, MORALES A E, SANZ A. Antioxidant defenses in fish: biotic and abiotic factors[J]. Rev Fish Biol Fish, 2005, 15(1/2): 75-88.
[7] 连春盎, 李健, 李吉涛, 等. 干露胁迫对脊尾白虾 (Exopalaemon carinicauda) 呼吸代谢相关酶的影响[J]. 渔业科学进展, 2017, 38(4): 53-60. [8] 段亚飞, 董宏标, 王芸, 等. 干露胁迫对日本囊对虾抗氧化酶活性的影响[J]. 南方水产科学, 2015, 11(4): 102-108. doi: 10.3969/j.issn.2095-0780.2015.04.015 [9] 姜令绪, 刘群, 王仁杰, 等. 三疣梭子蟹 (Portunus trituberculatus) 幼体不同干露温度下死亡率的研究[J]. 海洋与湖沼, 2012, 43(1): 127-131. doi: 10.11693/hyhz201201021021 [10] 王明焰. 提高小龙虾虾苗长途运输成活率技术要点[J]. 科学养鱼, 2020(5): 11-12. doi: 10.3969/j.issn.1004-843X.2020.05.006 [11] 马贵范, 杨凤, 郭文学, 等. 中国蛤蜊和四角蛤蜊对干露和淡水浸泡的耐受性比较[J]. 大连海洋大学学报, 2013, 28(1): 44-48. doi: 10.3969/j.issn.2095-1388.2013.01.009 [12] 温小波, 库夭梅, 罗静波. 克氏原螯虾耗氧率及窒息点的研究[J]. 大连水产学院学报, 2003, 18(3): 170-174. [13] 平洪领, 李玉全. 干露对天津厚蟹水分代谢和死亡率的影响[J]. 湖北农业科学, 2014, 53(18): 4377-4380. [14] 王海锋, 成永旭, 李京昊, 等. 干露和再入水对克氏原螯虾抗氧化应激能力的影响[J]. 南方水产科学, 2019, 15(5): 69-76. doi: 10.12131/20190059 [15] OMORI K, IRAWAN B, KIKUTANI Y. Studies on the salinity and desiccation tolerances of Helice tridens and Helice japonica (Decapoda: Grapsidae)[J]. Hydrobiologia, 1998, 386(1/2/3): 27-36. doi: 10.1023/A:1003461911201
[16] CARDINAUD M, OFFRET C, HUCHETTE S, et al. The impacts of handling and air exposure on immune parameters, gene expression, and susceptibility to vibriosis of European abalone Haliotis tuberculata[J]. Fish Shellfish Immunol, 2014, 36(1): 1-8. doi: 10.1016/j.fsi.2013.09.034
[17] ABUMANDOUR M M. Gill morphology in the red swamp freshwater crayfish Procambarus clarkii (Crustacea: Decapoda: Cambarids) (Girard 1852) from the river Nile and its branches in Egypt[J]. Int J Morphol, 2016, 34(1): 168-178. doi: 10.4067/S0717-95022016000100025
[18] 曾朝曙, 李少菁. 温度对锯缘青蟹幼体存活与发育的影响[J]. 水产学报, 1992, 16(3): 213-221. [19] VIRU M. Differences in effects of various training regimens on metabolism of skeletal muscles[J]. J Sports Med Phys Fit, 1994, 34(3): 217-227.
[20] 陶易凡, 强俊, 王辉, 等. 低pH胁迫对克氏原螯虾鳃和肝胰腺酶活力及组织结构的影响[J]. 中国水产科学, 2016, 23(6): 1279-1289. [21] 段亚飞, 董宏标, 王芸, 等. 干露胁迫对日本囊对虾呼吸代谢酶活性和RNA/DNA比值的影响[J]. 海洋渔业, 2016, 38(1): 42-50. doi: 10.3969/j.issn.1004-2490.2016.01.006 [22] KAJA S, PAYNE A J, SINGH T, et al. An optimized lactate dehydrogenase release assay for screening of drug candidates in neuroscience[J]. J Pharmacol Toxicol Meth, 2015, 73: 1-6.
[23] RUSTIN P, CHRETIEN D, BOURGERON T, et al. Biochemical and molecular investigations in respiratory chain deficiencies[J]. Clinica Chimica Acta, 1994, 228(1): 35-51. doi: 10.1016/0009-8981(94)90055-8
[24] GAO X L, ZHANG M, LI X, et al. Effects of LED light quality on the growth, metabolism, and energy budgets of Haliotis discus discus[J]. Aquaculture, 2016, 453: 31-39. doi: 10.1016/j.aquaculture.2015.11.033
[25] OLIVEIRA G T, EICHLER P, ROSSI I C, et al. Hepatopancreas gluconeogenesis during anoxia and post-anoxia recovery in Chasmagnathus granulata crabs maintained on high-protein or carbohydrate-rich diets[J]. J Exp Zool Pt A, 2004, 301(3): 240-248. doi: 10.1002/jez.a.20026
[26] 赵卫红, 王资生, 张余霞, 等. 雌二醇对日本沼虾肝胰腺的脂肪酸含量及组织结构的影响[J]. 海洋渔业, 2014, 36(6): 542-548. doi: 10.3969/j.issn.1004-2490.2014.06.010 [27] 张晴, 姜明慧, 田元勇, 等. 虾夷扇贝闭壳肌蛋白溶解性与三磷酸腺苷的关联性[J]. 水产科学, 2020, 39(4): 476-482. [28] 李娜, 赵永强, 李来好, 等. 冰藏过程中罗非鱼鱼片肌肉蛋白质变化[J]. 南方水产科学, 2016, 12(2): 88-94. doi: 10.3969/j.issn.2095-0780.2016.02.013 [29] SOLLID J, NILSSON G E. Plasticity of respiratory structures: adaptive remodeling of fish gills induced by ambient oxygen and temperature[J]. Respir Physiol Neurobiol, 2006, 154(1): 241-251.
[30] 王权, 王建国, 陆宏达, 等. 硫酸锌慢性毒性胁迫下克氏原螯虾的组织病理[J]. 中国水产科学, 2012, 19(1): 126-137. [31] 丁正峰, 薛晖, 王晓丰, 等. 毒死蜱 (CPF) 对克氏原螯虾的急性毒性及组织病理观察[J]. 生态与农村环境学报, 2012, 28(4): 462-467. doi: 10.3969/j.issn.1673-4831.2012.04.023 [32] WEI K Q, YANG J X. Histological alterations and immune response in the crayfish Procambarus clarkii given rVP28-incorporated diets[J]. Fish Shellfish Immunol, 2011, 31(6): 1122-1128. doi: 10.1016/j.fsi.2011.10.002
[33] 韩晓琳, 高保全, 王好锋, 等. 低盐胁迫对三疣梭子蟹鳃和肝胰腺显微结构及家系存活的影响[J]. 渔业科学进展, 2014, 35(1): 104-110. doi: 10.3969/j.issn.1000-7075.2014.01.015 [34] 洪美玲, 陈立侨, 顾顺樟, 等. 氨氮胁迫对中华绒螯蟹免疫指标及肝胰腺组织结构的影响[J]. 中国水产科学, 2007, 14(3): 412-418. doi: 10.3321/j.issn:1005-8737.2007.03.010 [35] JONG-MOREAU L, BRUNET M, CASANOVA J P, et al. Comparative structure and ultrastructure of the midgut and hepatopancreas of five species of Mysidacea (Crustacea): functional implications[J]. Can J Zool, 2000, 78(5): 822-834. doi: 10.1139/z99-263
[36] 李二超, 陈立侨, 曾嶒, 等. 不同盐度下饵料蛋白质含量对凡纳滨对虾生长、体成份和肝胰腺组织结构的影响[J]. 水产学报, 2008, 32(3): 425-433. -
期刊类型引用(9)
1. 傅建军,安睿,朱文彬,王兰梅,罗明坤,董在杰. 套养池塘主要养殖阶段浮游生物的群落结构特征. 水产学杂志. 2024(01): 104-112 .  百度学术
百度学术
2. 向劲,吴启藩,宋锐,彭治桃,李金龙,高峰,谢敏,程小飞. 循环水养殖池塘中浮游动植物功能群特征及水质评价. 水产学杂志. 2024(01): 81-89 . 百度学术
3. 闵文武,王龙燕,陈飞雄,周其椿. 集装箱养殖生态净化池塘中浮游植物群落结构多样性分析. 水产科技情报. 2024(05): 302-310 . 百度学术
4. 郑颖强,代梨梨,张辉,彭亮,陶玲,李谷,柴毅,廖咏玲. 池塘循环水养殖系统净化单元对水体浮游植物的影响. 中国水产科学. 2024(12): 1497-1510 . 百度学术
5. 王小冬,车轩,刘兴国,顾兆俊,丁艳青,陈晓龙. 曝气扰动促进高营养的饲料腐烂液中绿藻生长. 上海海洋大学学报. 2023(01): 142-149 . 百度学术
6. 闵文武,王龙燕,陈飞雄,周其椿,赵瑞平. 生态净化池塘浮游动物群落结构特征及其与环境因子的关系. 贵州农业科学. 2023(02): 66-75 . 百度学术
7. 赵秀侠,方婷,陈金良,高娜,陈诚,卢文轩. 陆基推水集装箱循环水养殖系统中浮游植物群落结构特征. 农业环境科学学报. 2023(04): 869-878 . 百度学术
8. 王龙燕,闵文武,王金乐,陈飞雄,周其椿,吴俣学. 集装箱循环水养殖系统中浮游植物群落结构特征及影响因素. 渔业现代化. 2022(04): 26-36 . 百度学术
9. 赵宇曦,刘兴国,周润锋,肖述文,孙照云. 池塘多营养级养殖水体的初级生产力及影响因子分析. 渔业现代化. 2022(06): 91-99 . 百度学术
其他类型引用(4)

 下载:
下载:



















计量
- 文章访问数: 725
- HTML全文浏览量: 235
- PDF下载量: 55
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
